
Abstract
Previous studies arguing for parental care in dinosaurs have been primarily based on fossil accumulations of adults and hatchlings, perinatal and post-hatchlings in nests and nest areas, and evidence of brooding, the majority of which date to the Late Cretaceous. Similarly, the general body proportions of preserved embryonic skeletons of the much older Early Jurassic Massospondylus have been used to suggest that hatchlings were unable to forage for themselves. Here, we approach the question of parental care in dinosaurs by using a combined morphological, chemical, and biomechanical approach to compare early embryonic and hatchling bones of the Early Jurassic sauropodomorph Lufengosaurus with those of extant avian taxa with known levels of parental care. We compare femora, the main weight-bearing limb bone, at various embryonic and post-embryonic stages in a precocious and an altricial extant avian dinosaur with those of embryonic and hatchling Lufengosaurus, and find that the rate and degree of bone development in Lufengosaurus is closer to that of the highly altricial Columba (pigeon) than the precocious Gallus (chicken), providing strong support for the hypothesis that Lufengosaurus was fully altricial. We suggest that the limb bones of Lufengosaurus hatchlings were not strong enough to forage for themselves and would likely need parental feeding.
Similar content being viewed by others


Introduction
Parental care involving brooding, feeding, and protection has been the subject of scientific investigations for generations, but in the fossil record it did not attract significant attention until the discovery of Late Cretaceous dinosaur nesting sites preserving embryos, hatchlings and adults together1,2,3,4. Hypotheses of parental care in ornithischian dinosaurs were then based on this associational evidence given the difficulty in preserving direct records of behaviour5,6,7,8,9. It was the discovery of the Late Cretaceous oviraptorosaur found atop a nest that led to the reasonable interpretation as providing protection and possibly brooding behaviour10. The hypothesis of parental care based on body proportions like allometric limb proportions and a shift in the center of mass cranially of embryos and hatchlings was first proposed for the Early Jurassic sauropodomorph Massospondylus, although this could represent circumstantial evidence as no hatchling skeletons were found at the time11,12,13,14,15,16.
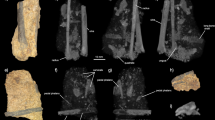
The Early Jurassic dinosaur Lufengosaurus is unique in the fossil record because representatives of various stages of embryonic development have been found in an embryonic bonebed in addition to dozens of subadult and adult skeletons at the same classic locality near the village of Dawa, Lufeng, Yunnan Province, China. Such an ontogenetic record makes it possible to compare these skeletal elements with those of altricial and precocious extant birds (avian dinosaurs) with well-documented embryologic ontogenies and known levels of parental care. Altricial birds are those that cannot fend for themselves shortly after hatching and are reliant on parental care, while precocious birds are those that can, and do not rely on parental care17. The discovery of a partial skeleton of a hatchling (Chuxiong Prefectural Museum C2019 2A233-2003) was the impetus for the present study (Fig. 1, Supplementary Table 1, Supplementary Fig. 1).

Diameter of Lufengosaurus embryonic and hatchling femora. (a), Photograph and (b), semi-transparent rendering of partial semi-articulated hatchling Lufengosaurus skeleton, including femur (Chuxiong Prefectural Museum, no.C2019 2A233-2003). (c–h), Lufengosaurus femora (Chuxiong Prefectural Museum, no.C2019 2A233a-e) where the smallest diameter is c and the largest being h, which is a µCT rendering. Abbreviations: fe femur.
It is usually difficult to determine whether this relatively small skeleton represents a very late embryo, or a hatchling, but the difference in size between the known embryonic materials and this new skeleton would most likely indicate that this skeleton represents a hatchling. This interpretation is supported by the size and level of ossification of the skeletal elements, in particular the proximal preserved end of the femur, and the level of development of the dentition, which in contrast to the embryonic materials are protruding well beyond the edge of the jaw bones.
Here, we use three lines of evidence: hard tissue morphology at the histological level, chemical analyses of bone composition, and biomechanical strength of embryonic and hatchling limb bones to compare extant avian dinosaurs with known reproductive behaviours to Lufengosaurus. This approach takes interpretations of extinct dinosaur parental care out of the realm of association and into comparative and quantitative evaluations of locomotory abilities.
Embryonic long bones of dinosaurs, both extinct and extant, have extensive Primary Tubular Cavities (PTC), large cavities within developing endochondral bone and these are generally regarded as typical for fast-growing non-remodeled bone14,15. For example, the %PTC of the femur is directly related to the amount of apatite group minerals present in the shaft portion of the bone, with higher %PTC values indicating less hard tissue than that in the shafts with a lower %PTC. The amount of bone tissue as represented by apatite minerals is directly related to bone strength, as more robust long bones have thicker cortical bone and a smaller PTC. The rapidly forming cortical bone of embryonic femora in both extant birds (avian dinosaurs) and Lufengosaurus can be directly compared to each other, and the varying levels of %PTC, representing the varying levels of bone/hard tissue building apatite and the resulting increases in bone strength, can be traced throughout their embryonic development.
The amount of apatite in the bones of an animal can be calculated by its chemical composition through the concentration of phosphorus because the dominant mineral components of both extant and extinct tetrapod bones are carbonatoapatite-A and carbonatoapatite-B18,19,20, with phosphorus atoms in various distorted tetrahedral environments21. The complicated nature of the bio-apatite group minerals in bone makes the use of the omnipresent phosphorus preferable for evaluating mineral content. Interestingly, as previously shown22,23,24, Rare Earth Elements (REE) are readily incorporated into these apatite group minerals in fossilized bones, and ICP-MS (Inductively Coupled Plasma Mass Spectrometry) analyses have shown that there is a direct relationship between the amount of REE and the proportion of phosphorus in Lufengosaurus adult and embryonic dinosaur bones at this locality25. It was also possible to determine P concentrations in embryos and hatchlings of Gallus and Columba, the common chicken and pigeon, using similar methods, allowing for direct comparisons between these extant taxa with Lufengosaurus.
Micro CT (µ-CT) imaging has been successfully used in bone quantifications in different animal models26,27,28, with recent advanced imaging technology of µ-CT offering high spatial resolution down to micrometer and nanometer scales. It was therefore possible to derive several quantitative parameters from µ-CT images to characterize femoral trabecular and cortical bones. These parameters for bone quality and strength characterization include bone mineral density, percent bone volume, trabecular segment number, and cortical bone fraction (CF, Supplementary S2, S3, Table 5, Supplementary Data 1–3). Quantitative values were extracted from µ-CT images and used non-destructively to study the biomechanical bone strength of the dinosaur Lufengosaurus and extant birds, and include cortical bone fraction, effective bone strength (EBS, Supplementary S2, S3, Table 5, Supplementary Data 1–3), and percent bone volume (PBV or PTC, where PBV = 1-%PTC). Lower EBS values correspond to weaker bone strength more typical of an altricial condition, while higher EBS values correspond to greater bone strength more typical of the precocious condition.
Results
Since the embryos of the precocious Gallus have much lower %PTC values than altricial Columba at each stage of embryonic life (Supplementary Table 1), we can calculate the rate of growth in material strength of the femora in animals with vastly different locomotory abilities at hatching and evaluate comparatively their locomotory capabilities to those of embryos and hatchlings of extinct dinosaurs. For example, the mid-shaft of the Lufengosaurus femora also had high %PTC (Fig. 2, Supplementary Table 1), ranging from 56—65%, likely indicating not only very fast embryonic growth, but also relatively little hard tissue formation.

Morphological comparison of the mid-shaft of DaWa Lufengosaurus, Gallus and Columba embryonic femora. (a), 2µm resolution µ-CT scan of Lufengosaurus (Chuxiong Prefectural Museum, no.C2019 2A233) femur. (b), Neutron scan of femur at 6.7µm resolution of Lufengosaurus. (c–e), Bone and Primary Tubular Cavity (PTC) rendering as 3D distribution in embryos of Lufengosaurus, Gallus and Columba, respectively, with apatite group minerals (bone) shown in color. Top row show longitudinal and lower row show transverse views. Extant birds embryos were one day before hatching. (f), 3D Second Harmonic Generation (SHG) image of Lufengosaurus embryonic femur. (g), 3D Third Harmonic Generation (THG) image of Lufengosaurus embryonic femur. (h), Partially enlarged combination of SHG and THG images (yellow boxes) of Lufengosaurus embryonic femur.
Morphology
Our 3D µ-CT and neutron scan30 (Fig. 2, Supplementary Table 1) data of several Lufengosaurus femora, undertaken for this study, confirm the presence of high %PTC values (average of 53.9%) in 3D. Similar µ-CT analysis of Columba embryonic femora also showed high 3D %PTC values (average of 56.2%).
Multiple Harmonic Generation Microscopy (MHGM)31 (Supplementary S1) also confirmed this pattern of a highly porous bone structure, and hence low effective bone strength. In sharp contrast, µ-CT data of Gallus showed much lower 3D %PTC values (average of 32.6%), and hence more hard tissues (bio-apatites) for greater biomechanical strength. These 3D %PTC values provide direct morphological support to the hypothesis that Lufengosaurus embryonic limbs were similar to the altricial Columba embryos, and significantly weaker bone structure than the precocious Gallus. Although it has been suggested that Lufengosaurus and other early Jurassic sauropodomorphs were quadrupedal as hatchlings, and transitioned to a bipedal posture and locomotion as they grew to adulthood, we do not feel that this would have an effect on the results.
Chemical analyses
The chemical composition and amount of hard tissues in fossil and extant bone have been successfully calculated, in particular the value of phosphorus18,19,20,21,22. In addition, Rare Earth Elements (REE) found in Lufengosaurus, have been incorporated into the apatite group minerals of the fossilized bones with a direct relationship between the amount of REE and the proportion of phosphorus23,24,25. Although the embryonic growth series of Lufengosaurus femora were not available for destructive sampling for such an analysis, when adult and other embryonic bones were subjected to REE analyses from the Dawa locality, it was found that the total La-REE amount of the adult Lufengosaurus bone was about ten times higher than that of the embryonic bones24 (Supplementary Table 2), confirming the contrast between them, as embryonic bone had a high %PTC and relatively little hard tissue (and little phosphorus), while PTC were largely absent in adult bone, and massive cortical hard tissue was present instead.
Once laid, the total amount of phosphorus inside the egg is fixed, limiting the total phosphorus available for bone formation in the embryo. Once hatched, additional phosphorus can be recruited from food, and this pattern can be used to look at rates of bone formation26 (and bone strength) in embryonic and postembryonic dinosaurs (including birds). The differences in %PTC values shown above suggest that embryonic Lufengosaurus Columba femora used less phosphorus for ossification than in embryonic Gallus.
Building on this, the phosphorus ratios of the adult to the embryonic Columba and Gallus were measured at various stages before and after the hatching (Fig. 3). The results show that during embryonic development (Fig. 3, left side), increases of phosphorus in the Gallus femur (red line) are much more rapid than in Columba (blue line). Once hatched, the phosphorus in Gallus increases slowly in the first week, while Columba shows a very significant increase in the rate of phosphorus concentration. These patterns, indicative of the rate of bone hard tissue formation26, are significantly different in these precocious and altricial birds. Gallus undergoes rapid bone mineral deposition during embryology, and then slows down after hatching and during consequent food foraging. In contrast, Columba has lower relative levels of mineral deposition and bone formation during embryology but shows rapid bone formation during parental feeding. These different trajectories seem related to different types of parental care.

Ratios of phosphorus (P) concentration change for Columba and Gallus. Incubation ratio represents percentage of embryonic development, where 1 is the point of hatching, this is done because the actual duration of embryonic development varies between the two bird species. The rest of the x-axis post hatching (> 1) is the number of days. Hatching point is indicated by the thick vertical line, value of 1 along the X-axis. P values increase in embryos of Gallus faster with a much steeper trend line (red) than that of Columba, which is nearly flat trend line (blue). Ratios of adult to embryonic P concentration is listed in the subset chart together with those in Gallus and Columba. Although incubation ratio is unknown for Lufengosaurus and thus is not plottable, the ratio between P concentration of known adult specimens and, hatchling specimens is listed in the subset chart and is closer to the ratio in Columba and distinct from Gallus. The P data are available in Supplementary Table 2.
The phosphorus concentration ratio between Lufengosaurus adult and embryo materials is also generally closer to the ratio in Columba and distinct from that in the precocial Gallus, the latter showing a 52-fold gain of phosphorus between embryos and adults, while altricial Columba shows a 336-fold gain. This suggests that bone formation was much slower in the embryonic Columba than in Gallus, but much more rapid post-hatching. The available evidence indicates that Lufengosaurus (of unknown incubation days, but sampled at the largest known embryonic sample14,25 had a 218-fold gain between the embryo and adult, and closer to that seen in the altricial Columba.
Biomechanical analysis
Micro CT (µ-CT) imaging for bone quantifications in different animal models27,28,29,30, with recent advanced imaging technology for high spatial resolution31,32 successfully produced several quantitative parameters to characterize femoral bone density. These are correlated to the biomechanical strength of femoral cortical bones and include cortical bone fraction (CF), percent bone volume (PBV), and effective bone strength (EBS) (Supplementary S2, S3, Supplementary Table 5, ementary Data 1, 2).
The quantitative approach to determine bone strength combines whole bone strength with internal density. The former Deals with the strength of the overall limb element structurally, while the latter analyzes the building material density of the internal structures (CF and PBV). They are then combined for a meaningful biomechanical strength of the whole embryonic femur, called the Effective Bone Strength (EBS, 0—100), with higher values representing a stronger limb bone. The calculated value of EBSGallus at hatching is 35.1, the average EBSLufengosaurus of approximately late incubation stage is 28.9 while EBSColumba at hatching is 25.4, as shown in Fig. 4a (ementary Tables 3, 4). Thus, the EBS of Lufengosaurus is much more similar to that of Columba than to Gallus, with both also preserving similar slopes of EBS to proportional femur diameter (Fig. 4a). They are further distinguished from Gallus by their highest pre-hatching EBS values being less than the lowest embryonic values of Gallus (Fig. 4, Supplementary S3). This pattern is largely re-capitulated in the %PTC values (Fig. 4ba), where Lufengosaurus is intermediate in %PTC between Columba and Gallus, but more similar to Columba in both %PTC pattern through embryonic development and in possessing higher absolute %PTC values at hatching (comparable to the early embryonic stages of Gallus). The absolute differences in EBS and %PTC observed in these taxa indicate that Lufengosaurus did not have enough bone strength to forage for themselves at hatching, similar to Columba.

Comparisons of biomechanical measures against life stage. (a), Plot of effective bone strength (EBS) against proportional femur diameter, calculated as the femoral diameter of a specimen (X) divided by the diameter at hatching (Dperinatal/hatchling), and used as a relative indicator of incubation/life stage. The vertical dashed line at proportional femur diameter = 1 represents the approximate condition at hatching. (b), Violin plot, showing density and distribution of %PTC measurements for each taxon, with incubation/life stage indicated. The absolute values and relative data distributions of Lufengosaurus and Columba are more similar to each other than either is to Gallus. EBS and %PTC data available in Supplementary Tables 3&4. Additional information is provided in Supplementary Fig. 2.
Discussion and conclusion
Morphological, chemical and biomechanical evidence comparing altricial and precocious extant birds to Lufengosarus show that the embryonic femur of this dinosaur, the main weight-bearing element in the skeleton, is more similar to the altricial Columba than to the precocial Gallus. We propose that in Lufengosaurus, where a limited amount of phosphorus is available during the embryonic bone-building stage, only the most important bone framework directed at rapid growth was built, with extensive primary tubular cavities being present (high %PTC). As similarities with the altricial Columba are extensive, we propose that hatchling Lufengosaurus would obtain additional needed phosphorus for building bone strength from the food provided by its parents, as the hatchlings would lack the bone strength to do so for themselves. This multidisciplinary comparative study therefore provides evidence that Lufengosaurus hatchlings were likely altricial, and parental feeding may have been required until they acquired enough bone strength to forage for themselves. This current study, restricted to two extant avian taxa and an early Jurassic sauropodomorph dinosaur is limited in scope, and its purpose is to provide a new, novel approach to the study of dinosaur behaviour. We recognize that there are important differences between the extant birds of this study and the sauropodomorph Lufengosaurus in terms of likely rates of growth, with birds having determinant growth in contrast to the non-determinant growth of non-avian dinosaurs. Interestingly, the size differences between the eggs, embryos, and early hatchlings of Gallus and Lufengosaurus are surprisingly small. Previous studies have shown that the eggs of early Jurassic sauropodomorphs are surprisingly small, sub-spherical in shape and ranging from about 5-7 cm in diameter. We would like to suggest that the comparisons between the taxa used in this study are therefore appropriate. It is hoped that it will lead to other expanded studies using similar methodologies. For example, this approach may also apply to embryos and hatchlings of other morphologically similar early sauropodomorphs like Massospondylus, which had awkwardly proportioned bodies with huge heads on a long slender neck and long forelimbs, generally indicative of poor locomotory capabilities.
Materials and methods
Lufengosaurus embryonic femora (ChuXiong Prefecture Museum, Catalogue no. C2019 2A233 and C2019 2A233-2003) were used in this study from a single site, and they ranged in sample size from one for the earliest known embryonic stage and one for the hatchling, and 15 femora at various intermediate embryonic stages for both histological and µ-CT scans. In addition, the femora of extant Columba and Gallus were used for µ-CT scans, and all were visualized using OsiriX, Drishti, Avizo Lite, and ImageJ/Fiji for post-processing. 3D %PTC used in this study is defined as the total %PTC inside a given volume in the mid-shaft section of an embryonic femur cortical bone. Ten or more µ-CT image slices were used for the measurements and calculations, as shown in Supplementary Table 1, %PTC comparison. Neutron Tomography scans were done at DINGO station of ANSTO OPAL reactor at 6.7 µm resolution. The ICP-MS work was carried out by using Agilent 7500ce, which can detect 75 elements at National Tsing Hwa University in Taiwan and other labs for the elemental analyses, including REE and P. Biomechanical analyses were done using µ-CT scan images.
The eggs of Gallus and Columba were collected and stored at 16 °C.
70% humidity to ensure that there is no embryonic development prior to experiment. For incubation, eggs were gradually warmed to a temperature to 37 °C for a few hours to allow acclimation. Then, the eggs were maintained at 38 °C and 60–70% humidity with frequent rotation.
during the incubation process. On days 7 and 14, the eggs were examined for determining the internal state of embryonic development through candling. In total 5 femoral bones at each ontogenetic stage were collected for comparisons for the different embryonic developmental stage of this study.
The biomechanical analyses combined macro and micro views for evaluating whole bone strength of the femora. Cortical bone fraction was determined at macro view using the outer bone diameter and that of the inner medullary cavity, while the micro view used high resolution µ-CT scans for 3D % PTC measurements and analysis of the biomechanical strength of the cortical bone (For more detailed information, see Supplementary S2).
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information files.
References
Horner, J. R. & Makela, R. Nest of juveniles provides evidence of family structure among dinosaur. Nature https://doi.org/10.1038/282296a0 (1979).
Horner, J. R. Evidence of colonial nesting and ‘site fidelity’among ornithischian dinosaurs. Nature 297, 675–676 (1982).
Fastovsky, D. E. et al. A Nest of Protoceratops andrewsi (Dinosauria, Orinthischia). J. Paleontol. 85(6), 1035–1041 (2011).
Meng, Q. J., Liu, J. Y., Varricchio, D. J., Huang, T. D. & Gao, C. L. Parental Care in an Ornithischian Dinosaur. Nature 431, 145–146 (2004).
Coria, R. A. & Chiappe, L. M. Embryonic skin from Late Cretaceous sauropods (Dinosauria) of Auca Mahuevo, Patagonia, Argentina. J. Paleontol. 81(6), 1528–1532 (2007).
Amiot, R. et al. δ18O-derived incubation temperatures of oviraptorosaur eggs. Palaeontology 60, 633–647 (2017).
Varricchio, D. J. et al. Avian paternal care had dinosaur origin. Science 322, 1826–1828 (2008).
Jackson, F. D., Horner, J. R. & Varricchio, D. J. A study of a Troodon egg containing embryonic remains using epifluorescence microscopy and other techniques. Cret. Res. 31, 255–262 (2010).
Starck, M. J. & Ricklefs, R. E. Avian Growth and Development (Evolution within the altricial precocial spectrum (Oxford University Press, 1998).
Clark, J. M., Norell, M. A. & Chiappe, L. M. An oviraptorid skeleton from the Late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avian-like brooding position over an oviraptorid nest. Am. Mus. Novit. 3265, 1–36 (1999).
Reisz, R. R., Evans, D. C., Sues, H. D. & Scott, D. Embryonic Skeletal Anatomy of the Sauropodomorph Dinosaur Massospondylus from the Lower Jurassic of South Africa. J. Vert. Paleontol. 30(6), 1653–1664 (2010).
Reisz, R. R., Evans, D. C., Roberts, E. M., Sues, H. D. & Yates, A. M. Oldest known dinosaurian nesting site and reproductive biology of the Early Jurassic sauropodomorph Massospondylus. Proc. Nat. Acad. Sci. 109(7), 2428–2433 (2012).
Reisz, R. R., Scott, D., Sues, H.-D., Evans, D. C. & Raath, M. A. Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance. Science https://doi.org/10.1126/science.1114942 (2005).
Reisz, R. R. et al. Embryology of Early Jurassic dinosaur from China with evidence of preserved organic remains. Nature 496, 210–214 (2013).
Fu, R., Liu, C., Yan, Y., Li, Q. & Huang, R. L. Bone defect reconstruction via endochondral ossification: A developmental engineering strategy. J. Tissue Eng. 12, 1–22 (2021).
Horner, J. R., Padian, K. & de Ricqles, A. Comparative osteohistology of some embryonic and perinatal archosaurs: developmental and behavioral implications for dinosaurs. Paleobiology 27(1), 39–58 (2001).
Scheiber, I. B. R., Weiß, B. M., Kingma, S. A. & Komdeur, J. The importance of the altricial - precocial spectrum for social complexity in mammals and birds - a review. Front Zool. 14, 3 (2017).
Becker, J. S. et al. Determination of phosphorus in small amounts of protein samples by ICP-MS. Anal. Bioanal. Chem. 375(4), 561–566 (2003).
Faibish, D., Ott, S. M. & Boskey, A. L. Mineral changes in osteoporosis: A review. Clin. Orthop. Relat. Res. 443, 28–38 (2006).
Helliwell, T. R. et al. Elemental analysis of femoral bone from patients with fractured neck of femur or osteoarthrosis. Bone 18(2), 151–157 (1996).
Lee, Y. C. et al. Evidence of preserved collagen in an Early Jurassic sauropodomorph dinosaur revealed by synchrotron FTIR microspectroscopy. Nat. Comm. 8, 14220 (2019).
Kaflak, A. Efficiency of 1H–31P NMR cross-polarization in bone apatite and its mineral standards. Solid State Nuclear Magnetic Resonance https://doi.org/10.1016/j.ssnmr.2005.11.005 (2006).
Trueman, C. N. & Benton, M. J. A geochemical method to trace the taphonomic history of reworked bones in sedimentary settings. Geology 25(3), 263–266 (1997).
Trueman, C. N. & Palmer, M. R. Diagenetic Origin of REE in Vertebrate Apatite: A Reconsideration of Samoilov and Benjamini, 1996. Palaios 12(5), 495–497 (1997).
Wang, S. et al. Rare earth elements in dinosaur bones across the embryo-adult spectrum. Front. Earth Sci. 8, 230 (2020).
Bouxsein, M. L. et al. Guidelines for assessment of bone microstructure in rodents using micro-computed tomography. J. Bone Min. Res. 25(7), 1468–1486 (2010).
Cotti, S. et al. More Bone with Less Minerals? The Effects of Dietary Phosphorus on the Post-Cranial Skeleton in Zebrafish. Int. J. Mol. Sci. 21(15), 5429. https://doi.org/10.3390/ijms21155429 (2020).
Tu, S. J., Wang, S. P., Cheng, F. C. & Chen, Y. J. Extraction of gray-scale intensity distributions from micro computed tomography imaging for femoral cortical bone differentiation between low-magnesium and normal diets in a laboratory mouse model. Sci. Rep. https://doi.org/10.1038/s41598-019-44610-8 (2019).
Tu, S. J. et al. Attenuating trabecular morphology associated with low magnesium diet evaluated using micro computed tomography. PLoS One https://doi.org/10.1371/journal.pone.0174806 (2017).
Tu, S. J., Wu, S. Y., Wang, F. S. & Ma, Y. H. Retention assessment of magnetic nanoparticles in rat arteries with micro-computed tomography. Phys. in Med. and Biol. 59(5), 1271–1281 (2014).
Mays, C., Bevitt, J.J. & Stilwell, J. Pushing the limits of neutron tomography in palaeontology: three-dimentional modelling of in situ resin within fossil plants. Palaeontol Electronica 20 (3.57 A), 1–12 (2017).
Chen, Y.-C. et al. Third-harmonic generation microscopy reveals dental anatomy in ancient fossils. Optics Lett. https://doi.org/10.1364/OL.40.001354 (2015).
Acknowledgements
We would like to thank CK Sun of Taiwan University for the use of MHGM. Dr. Tibor Pasinszki of Fiji National University, Suva, Fiji for the31P MAS NMR. We thank Thomas Cullen for aiding in the production of figure 4. This research was supported in part by grants MOST-104-2313-B-005-043-MY3 from the Ministry of Science and Technology, ATU-106-S0508 and MOE-108-S-0023 from the Ministry of Education, National Geographic Foundation for Science and Exploration—Asia-25-16, NSERC Canada, University of Toronto Mississauga, and Jilin University, China.
Author information
Authors and Affiliations
Contributions
TDH and RRR developed the project and supervised its progress with assistance from CMC. TDH, RRR wrote the paper. TDH, CMC, THT, SJT, and JJB contributed to the experiments and discussions. EDM worked on the illustrations, SJT did biomechanics calculations and analyses. CMC and TCT prepared the embryonic chicken and pigeon bone materials, ICP-MS and µ-CT data. SMZ contributed to field work in Lufeng with TDH, RRR. All authors reviewed manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Reisz, R.R., Huang, T.D., Chen, CM. et al. Parental feeding in the dinosaur Lufengosaurus revealed through multidisciplinary comparisons with altricial and precocious birds. Sci Rep 14, 20309 (2024). https://doi.org/10.1038/s41598-024-70981-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-70981-8
- Springer Nature Limited