
Abstract
Cliffs contain one of the least known plant communities, which has been overlooked in biodiversity assessments due to the inherent inaccessibility. Our study adopted the unmanned aerial vehicle (UAV) with the telephoto camera to remotely clarify floristic variability across unreachable cliffs. Studied cliffs comprised 17 coastal and 13 inland cliffs in Gageodo of South Korea, among which 9 and 5 cliffs were grazed by the introduced cliff-dwelling goats. The UAV telephotography showed 154 and 166 plant species from coastal and inland cliffs, respectively. Inland cliffs contained more vascular plant species (P < 0.001), increased proportions of fern and woody species (P < 0.05), and decreased proportion of herbaceous species (P < 0.001) than coastal cliffs. It was also found that coastal and inland cliffs differed in the species composition (P < 0.001) rather than taxonomic beta diversity (P = 0.29). Furthermore, grazed coastal cliffs featured the elevated proportions of alien and annual herb species than ungrazed coastal cliffs (P < 0.05). This suggests that coastal cliffs might not be totally immune to grazing if the introduced herbivores are able to access cliff microhabitats; therefore, such anthropogenic introduction of cliff-dwelling herbivores should be excluded to conserve the native cliff plant communities.
Similar content being viewed by others


Introduction
Cliff is one of the valuable ecosystems in terms of its unique topography and ecological integrity1. Inherent inaccessibility of cliffs has been traditionally considered to provide refuges for many species vulnerable to competitions and land use changes2,3,4. Environmental gradients along the vertical cliff landscape result in heterogenous taxonomic and lifeform compositions and high biodiversity within a relatively small area5,6,7. Recently, plant ecologists further emphasize the necessity of studying cliff flora, given morphological differences from the non-cliff flora8, specialized interactions with the surrounding fauna and microclimate9,10, biological invasions of alien species11, and impacts from leisure climbing and grazing of introduced ungulates12,13.
Cliff plants are among the least known plant communities, which has been overlooked in biodiversity assessments due to the poor accessibility14. Most of the cliff studies have depended on the engagements of climbing and rappelling experts, which requires risky, time-consuming, and labor-intensive works15. Such direct investigations are also affected by the bias to the limited sampling points on accessible and visible cliffs, and consequently cause the knowledge gap regarding unreachable cliff habitats16,17,18,19. Moreover, the use of climbing tools during the direct investigations may disturb cliff microtopography, and hinder the health and structure of the studied plant communities20,21.
Rapid developments of unmanned aerial vehicle (UAV) act as a remote sensing platform to revolutionize monitoring methodologies for unreachable cliff ecosystems22. First of all, smart-controlled positioning systems enabled semi-automated flight and image sampling along the extremely steep landscapes23. The UAV studies have also collaborated with 3D modeling24, AI technology25, GIS database26, and robotics27,28,29 to enlarge the applicability and usefulness of UAV remote sensing. In addition, preliminary field trials with a close-range photography provided a guideline for the UAV-based image sampling on the harsh, vertical ecosystems8,30. The attachment of high-quality gimbal and telephoto lens can further improve such close-range UAV photography by allowing more detailed species identification from the safer distance, although it was limitedly piloted for insect ecology31.
Despite these innovations, UAV-based researches on cliff plant communities are scarce because the majority of UAV applications have focused on technical aspects of remote sensing and data analysis rather than hypothesis-driven ecological studies22; correspondingly, only few UAV studies targeted on the structure and function of cliff plant communities. For example, Zhou et al.8 reported an elevating number of plant species with increasing cliff height and decreasing distance from stream across subtropical inland cliffs. Kim et al.30 found that north-facing cliffs contained fewer evergreen species and more deciduous species than south-facing cliffs along temperate island coasts. Other studies surveyed the microhabitat characteristics of endangered cliff-dwelling plants23,26. However, such UAV studies principally addressed the counts of detected individuals and species but did not adopt the beta diversity index, which has become a key concept to understand ecological processes in relation to various natural and anthropogenic factors32.
Our study used UAV telephotography to remotely unveil the structure and beta diversity of plant communities along unreachable cliff landscapes. An island under grazing of introduced goats was selected to specifically compare floristic composition and beta diversity depending on cliff topography and cliff-dwelling herbivores. Given the previous reports on the effects of cliff plant communities8,13,30,32, we set two null hypotheses as follows: (1) cliffs directly exposed to the ocean surface (coastal cliffs) would differ in the flora and beta diversity from cliffs sheltered by vegetated, terrestrial landscapes (inland cliffs) and (2) grazing of introduced cliff-dwelling herbivores would reorganize the species composition and beta diversity by accelerating biological invasions into the native cliff plant communities.
Results
Flora taxa
A total of 203 plant species from 81 families were detected throughout all studied cliffs (Table A1). Asteraceae (24 species), Poaceae (16 species), Rosaceae (8 species), Asparagaceae (7 species), and Crassulaceae (6 species) were the most prevalent families, accounting for 30.0% of the detected plant species. Hemerocallis hongdoensis (7.2%) occupied the highest incidence ratio, followed by Carex wahuensis var. robusta (6.6%) and Peucedanum terebinthaceum (4.3%) (Fig. 1a). If based on the cliff type, coastal and inland cliffs contained 154 and 166 species, respectively. C. wahuensis var. robusta (11.7%) and Selaginella tamariscina (6.9%) showed the highest incidence ratio in coastal and inland cliff types, respectively (Fig. 1a). There were four endangered, four Korea endemic, and 17 alien species in the studied cliffs (Table A1). Of the four endangered species (Fig. 1b), Neofinetia falcata was found exclusively in one of the coastal cliffs, while Dendrobium moniliforme, Bulbophyllum drymoglossum, and Bulbophyllum inconspicuum were observed exclusively in the inland cliffs. All these four endangered species were protected as CITES appendix II. Full list of the detected species is provided in Table A1.

Incidence ratio of the abundant plant species (a) and UAV telephoto images of endangered plant species detected from the studied cliffs (b). Black bars in panel (a) infer the species abundant at both cliff types. Incidence ratio indicates the percentage of counted individuals for each species within the total amount of the individuals (3841 for coastal cliffs, 4620 for inland cliffs).
Generalized linear mixed model (GLMM) demonstrated that cliff type (P < 0.001), cliff height (P < 0.001), and their interaction (P < 0.001) had a significant effect on the number of detected plant species (Fig. 2). Grazing had no significant effect on the number of detected plant species (P = 0.43). The inland cliffs generally involved more plant species than the coastal cliffs, although the number of detected plant species increased with cliff height (r = 0.73–0.84). The gradient of linear regression between the number of detected plant species and cliff height was steeper in the inland cliffs than in the coastal cliffs (Fig. 2). Meanwhile, cliff type (P < 0.001) and cliff height (P < 0.001) had a significant effect on Shannon diversity index, while grazing and their interactions had no significant effect (P = 0.07–0.33). Shannon diversity index (mean ± standard error) was generally higher in the inland cliffs (4.10 ± 0.06) than in the coastal cliffs (3.79 ± 0.06).

Comparison of the number of detected plant species between coastal and inland cliffs. Box graph with vertical error bars indicates average value and standard error (n = 17 for coastal cliffs and 13 for inland cliffs). Dots and dashed lines show the relationship between the number of plant species and cliff height. Letters above the box graph inform statistically significant difference based on Tukey’s HSD test (P < 0.05).
Of the 17 alien plant species, 16 and 7 were found in the coastal and inland cliffs, respectively (Table A1). The GLMM also exhibited that cliff type (P < 0.001), grazing (P < 0.05), and their interaction (P < 0.05) had a significant effect on the proportion of alien species (Fig. 3). Cliff height had no significant effect on the proportion of alien species (P = 0.09). The coastal cliff (5.3–9.8%) generally had the higher proportions of alien species than the inland cliffs (0.6–1.4%). Grazing elevated the proportion of alien species only in the coastal cliffs rather than the inland cliffs (Fig. 3), reflecting that the coastal cliffs were potentially more vulnerable to grazing interventions than the inland cliffs.

Proportion of alien species of coastal and inland cliffs with or without grazing. Error bars indicate standard error. Letters above the error bars inform statistically significant difference based on Tukey’s HSD test (P < 0.05) (n = 8 for ungrazed coastal cliffs, 9 for grazed coastal cliffs, 8 for ungrazed inland cliffs, and 5 for grazed inland cliffs).
Lifeform composition
The coastal cliffs featured the higher proportions of herbaceous (annual and perennial) species but lower proportions of fern and woody (deciduous and evergreen) species than the inland cliffs (Table 2). The GLMM demonstrated that cliff type (P < 0.001), grazing (P < 0.01) and its interaction (P < 0.001) had a significant effect on the proportion of annual herb species (Table 2). The post-hoc test also showed that grazing increased the proportion of annual herb species only in the coastal cliffs, while such difference from grazing was not detected in the inland cliffs. Conversely, only cliff type (P < 0.05) had a significant effect on the proportions of other lifeforms while the effect of grazing and cliff height had no consistent pattern.
Herbaceous (annual: 45.9%, perennial: 37.8%) species accounted for the largest proportion within the 37 species exclusively detected in the coastal cliffs, and no fern species was exclusive to the coastal cliff (Fig. 4a). Inversely, deciduous woody species (32.7%) recorded the largest proportion within the 49 species exclusively detected in the inland cliffs, and only 1 annual herb species was exclusive to the inland cliffs (Fig. 4a). All 5 major exclusive species (based on incidence ratio) were herbaceous in the coastal cliffs, whereas 2, 2, and 1 of them were perennial herb, evergreen woody, and fern species in the inland cliffs, respectively (Fig. 4b).

Number of plant species exclusively detected in coastal or inland cliffs (a) and incidence ratio of the major species exclusively detected in either cliff type (b). Incidence ratio indicates the percentage of counted individuals for each species within the total amount of the individuals (3841 for coastal cliffs, 4620 for inland cliffs).
Species composition and beta diversity
First, second, and third axes of principal coordinates analysis (PCoA) explained 24.7, 8.2, and 7.3% of multivariate dispersions of the species composition, respectively (Fig. 5a). While the first axis clearly separated the inland cliffs from the coastal cliffs, the second axis slightly reflected the variations between grazed and ungrazed cliffs (Fig. 5a). The third axis was correlated to cliff height (r = 0.67, P < 0.01). Permutational multivariate analysis of variance (PERMANOVA) showed that cliff type (P < 0.001) and cliff height (P < 0.05) had a significant effect on the multivariate group centroids of species composition (Fig. 5a). However, permutational analysis of multivariate dispersion (PERMDISP) reported no significant effect of cliff type (P = 0.29) and grazing (P = 0.43) on the multivariate dispersions of species composition (Fig. 5a). Consistently, the beta diversity was similar across the cliff types and grazing conditions, which was mainly attributed to turnover component (βJTU: 0.826–0.876) instead of nestedness resultant component (βJNE: 0.028–0.043) (Fig. 5b, c).

Principal coordinates analysis (PCoA), permutational multivariate analysis of variance (PERMANOVA), and permutational analysis of multivariate dispersion (PERMDISP) on beta diversity with Jaccard dissimilarity index (a) and proportion of turnover (βJTU) and nestedness-resultant (βJNE) components for the beta diversity according to cliff type (b) and grazing (c). Color and shape of dots in panel (a) represent cliff type (coastal cliff: black, inland cliff: white) and grazing (grazed: triangle, ungrazed: circle), respectively. PERMANOVA and PERMDISP are based on 9999 permutations.
Discussion
Effects of cliff type
Regarding the first hypothesis, our results show that the inland cliffs contained more vascular plant species than the coastal cliffs (Fig. 2). Flora of the two cliff types also differed in lifeform composition, among which higher proportions of fern and woody species were found in the inland cliffs than in the coastal cliffs (Table 1). This might occur because inland cliffs potentially experience plant migration from the surrounding non-cliff forests and grasslands5, whereas the ocean could physically prevent such species input to coastal cliffs. Direct salt deposition by the ocean spray could also decrease the fitness of any plants without enough salt-tolerance1,30, by which only limited number of species could survive at microhabitats within the coastal cliffs compared to those within the inland cliffs.
The two cliff types were further distinguishable in terms of plant species composition, reflected by the different multivariate group centroids (Fig. 5a). It was found that only the flora of the coastal cliffs was characterized by the abundance of halophytic species such as C. wahuensis var robusta, Aster hispidus, Peucedanum japonicum, and Tetragonia tetragonoides (Figs. 3a, 6b)33. This pattern confirms the potential differentiation of plant communities between the coastal and inland cliffs according to the salt-tolerance1. Conversely, the inland cliffs featured the abundance of an epiphytic fern (S. tamariscina, Fig. 1a) and 5 fern species undetected in the coastal cliffs (Fig. 4a). Considering that epiphytes on the cliffs strongly rely on substrate conditions16, such difference indicates that saline microhabitats within the coastal cliffs might lower the fitness of epiphytic ferns.

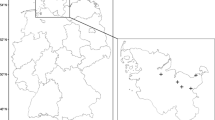
(Source: https://map.ngii.go.kr/ms/map/NlipMap.do, accessed on 8 December 2023).
Location of the studied cliff areas (a) and examples of coastal (b) and inland cliffs (c). The map used in panel (a) is taken from the open-source database of National Geographic Information Institute of South Korea
Another interesting finding is the effects of cliff type being dependent on cliff height. Particularly, the difference in the number of plant species between the coastal and inland cliffs was magnified with increasing cliff height (Fig. 2), although elevated cliff height provides larger surface for the survival of diverse plant species regardless of the cliff type8,30. In fact, coastal cliffs are known by their susceptibility to mass waste by the direct exposure to tidal impacts from the ocean34 and to soil erosion at the cliff top by rainfall and wind1,35. These extreme disturbances around coastal cliffs might reduce the availability of stable microhabitats to establish plant communities relative to inland cliffs18,36, and accordingly induce the slower increase in the number of plant species with the height of the coastal cliffs.
The multivariate dispersions of species composition, however, did not vary between the cliff types (Fig. 5a), inferring no significant differentiation in the taxonomic beta diversity37. This pattern is unexpected because the gradients of salt deposition are known to affect plant beta diversity along the coasts38,39. Such lack of the differences in the beta diversity possibly occurred because both the studied cliff types have a similar annual precipitation and parent material of Gageodo island40, which are the major factors controlling moisture availability and beta diversity of cliff plant communities16,41. The dominance of βJTU in both the cliff types also assists this idea (Fig. 5b), considering that the turnover components of beta diversity are the function of spatial, environmental variations within each group of comparison, not only the difference in species composition42.
Effects of grazing interventions
Anthropogenic introduction of herbivores has threatened island plant communities as it frequently leads to overgrazing, soil degradation, and disposal of invasive plant species43,44,45. In this context, cliffs in island ecosystems are considered as refuges for native plants to avoid such threats from the anthropogenic changes30,46. Regarding the second hypothesis, our results demonstrate that the grazed coastal cliffs involved the increased proportion of alien plant species than the ungrazed coastal cliffs (Fig. 3). Similar pattern was also observed by the elevated proportion of annual herb species under grazing of the coastal cliffs (Table 1). These patterns are in line with Thomson et al.45, who informed the invasion of non-native, fast-growing species and the suppression of native, slow-growing species by introduced herbivores in island plant communities. Our findings further suggest that cliff plant communities might not be immune to biological invasion and grazing if the introduced herbivores are able to access the cliff microhabitats. It supports previous reports on the alterations in cliff ledge flora after inclusion of cliff-dwelling ungulates13.
Interestingly, the effects of grazing were undetected in the inland cliffs (Fig. 3, Table 1), illustrating different vulnerabilities to the cliff-dwelling herbivores between the two cliff types. It might be related to the proximities to coast because repetitive natural disturbances around coastal areas promote the competitive replacement and invasibility of alien plant species47,48. This unstable environment might boost the spread of fast-growing, short-living, and non-native species only into the coastal cliffs under grazing interventions45. The relatively high proportion of woody species in the inland cliffs might also counteract the effects of grazing (Table 1), given that invasion into the woody plant communities requires the sufficient shade-tolerance49.
There was no distinct change in the species composition, beta diversity, and number of detected species under grazing in both the cliff types (Figs. 4, 7), unlike the proportions of annual herb and alien species. Our results are inconsistent with the previous reports on the negative impacts of introduced herbivores and alien species invasion on plant diversity across cliffs11 and other island ecosystems45,50. It reflects that the magnitude of such interventions across the studied cliffs might not be heavy enough to distort the overall taxonomic structure and beta diversity at the current stage. Otherwise, no effect on the species composition and beta diversity might occur as decrease in native plant diversity by biological invasions becomes more distinct at fine quadrate-scale investigations than landscape-scale approaches32. As the effects of grazing interventions could be altered by grazing intensity and elapsed time13,44, long-term, repetitive monitoring should be necessary to ensure whether the species composition and beta diversity will remain unchanged in the future.

Examples of transects to collect UAV image data (a), UAV images taken by wide-angle lens (b) and by telephoto lens for detailed species identification (c; Pyrrosia hastata).
Implications
The overall results allow us to partly accept the null hypotheses regarding the effects of cliff types and grazing on cliff flora. The coastal and inland cliffs appeared to differ in the number of detected species, lifeform composition, the proportion of alien species, and overall species composition with no clear differentiation in the beta diversity. Similarly, the grazed and ungrazed coastal cliffs were distinguishable according to the proportions of alien and annual herb species. In terms of biodiversity conservation, these findings indicate that cliff plant communities along the coasts might be sensitive to biological invasion of alien plants11, which could be magnified by grazing of the introduced cliff-dwelling ungulates. In fact, the proportion of alien species was similar between the grazed coastal cliffs (9.8%) and non-cliff areas (8.9%)51,52,53,54, both of which were higher than those in the ungrazed coastal cliffs (5.3%) as well as the inland cliffs (0.6–1.4%). Considering that coastal cliffs of the neighboring, well-preserved island reserve featured the lower proportion of alien plant species (2.1%)30, the anthropogenically introduced herbivores should be excluded to conserve the native plant communities along the coastal cliffs. Given the high turnover components within the beta diversity across the studied cliffs (Fig. 5), we also suggest that individual cliffs potentially have unique plant communities requiring multi-site conservation strategies rather than protection of a single habitat with the highest species abundance32. As only one southeastern coastal cliff contained the habitat of the endangered species N. falcata, particular attentions should be paid to that cliff site to prevent excessive grazing and illegal poaching.
Beyond the null hypotheses, our results exhibit the usefulness of UAV to study plant communities on the inaccessible ecosystems. For example, our UAV telephotography found the epiphytic fern, Pyrrosia hastata from inaccessible cliff microhabitats (Fig. 7c in “Methods” section), though previous studies on traditionally accessible sites have informed that this species does not occur in the study area51. The endangered orchid species, N. falcata has also remained undetected after the last observation by Yun et al.52 until the present study (Fig. 1b). Furthermore, the obtained telephoto images showed the natural reproduction of P. macrophyllus populations on the unreachable coastal cliff faces (Fig. A2), supporting the idea that this subtropical conifer might be native to the study area53. These floristic findings affirm the necessity of exploring inaccessible microhabitats to fully shed light on regional biodiversity by overcoming the uncertainties from sampling bias to traditionally reachable plant communities, even though the studied inaccessible cliffs contained only 37.5% of the vascular plant species in the non-cliff sites51. Similar utilizations of the UAV telephotography will help improve the reliability of the original close-range UAV approaches8,30, especially when identifying small plant species under other challenging conditions.
Several future collaborations may reinforce the usefulness of UAV for ecological researches. Particularly, the attachment of mechanical samplers will enable to collect eDNA from inaccessible ecosystems for the detailed phylogenetic analyses on any species without noticeable morphological traits27,28. Developing vertical auto flight methodologies may be helpful to reduce time–cost and enhance long-term monitoring consistency compared to the current UAV approaches based on manual flight23. The use of multispectral UAV sensors will deepen our understanding of plant ecophysiology at the harsh, challenging microhabitats, and relieve the confounding effects of spatial sampling scale by comparing with satellite imageries and ground-level observations9. Although grazing intensity can be an important factor controlling the cliff vegetation43, our study on the inaccessible cliffs did not quantitatively account for the grazing intensity. Thus, future studies should consider the gradients of herbivore population sizes and the associated grazing intensities for experimental design. Wind exposure, aspect, and other microtopographic factors may also be other critical factors influencing the floristic patterns across cliffs30, which further UAV investigations should take into consideration.
Conclusions
Cliffs are one of the least known ecosystems because of the inherent inaccessibility and harsh environmental conditions. Here, the UAV telephotography was adopted to overcome such challenges and describe its applicability to clarifying the influential factors for cliff plant communities. Results show that plant communities in the coastal cliffs featured the lower number of species, higher proportions of herbaceous and alien species, and the abundance of halophytic species than those in the inland cliffs. These differences between the cliff types are more on the overall species composition of plant communities (multivariate group centroids) rather than the beta diversity (multivariate group dispersion). In addition, grazing of the introduced goats elevated the proportions of annual herbs and alien species especially in the coastal cliffs. This indicates that coastal cliffs might be not totally immune to the anthropogenic interventions by grazing, requiring the appropriate exclusion treatments of the introduced cliff-dwelling herbivores. We expect that similar uses of the UAV telephotography will enhance our understanding of cliff plant communities as well as other unstudied ecosystems because of the extreme inaccessibility.
Methods
Study area
Studied cliff areas are located in Gageodo, the island at the southwesten end of South Korea (34°04′N, 125°06′E) (Fig. 6a). Distance from the main Korean peninsula is 127 km, and the average annual temperature and precipitation are 13.8 °C and 1120 mm51. Size of the island is approximately 9 km2, with the maximum altitude of 639 m54. The coasts of Gageodo feature the steep cliffs directly exposed to the ocean surface (Fig. 6b), and 95.8% of the lands comprise mountainous forests with rocky cliffs and ledges (Fig. 6c)55. The parent materials are igneous rocks, mainly consisting of andesite with a minor proportion of rhyolite40.
Natural vegetation of the inner mountains in Gageodo is characterized as broadleaved evergreen forests under the warm-temperate climate, which is dominated by Castanopsis sieboldii and Machilus thunbergii56. The latest studies on the accessible, non-cliff areas reported 541 vascular plant species51, of which 11, 6, and 48 were Korea endemic, endangered, and alien plant species, respectively52,53,54. This island is the northernmost distributional limit of the evergreen shrub, Viburnum japonicum52, and debatably considered as the only natural Korean habitat of the subtropical conifer, Podocarpus macrophyllus53. However, the vegetation of Gageodo has been impacted by anthropogenic interventions, such as overgrazed goats and cattle, recreational fishing, and long-term breakwater construction55. Due to the absence of native predators, overgrazed goats were accidentally naturalized throughout the entire island, and become invasive even to cliff and crag landscapes (Fig. A1).
Studied cliffs and UAV investigations
A UAV (Mavic 3 Enterprise, DJI, China) containing wide-angle (48 megapixel sensor with f/2.8 lens) and telephoto (12 megapixel sensor with f/4.4 lens; up to 7× optical and 56× hybrid zooms) cameras was utilized in the present study. This UAV weighs approximately 920 g and can fly up to 45 min. There are infrared and omni-directional sensing systems to detect surrounding obstacles for flight safety. It is also equipped with a real time kinematic (RTK) module, ensuring the positioning and hovering accuracies of 1 cm ± 1 ppm and ± 0.1 m for horizontal direction and 1.5 cm ± 0.1 ppm and ± 0.1 m for vertical direction, respectively.
Our UAV investigations targeted on a total of 30 cliff sites with different cliff types and grazing conditions (Table 2). Of them, 17 were categorized as the coastal cliffs that was directly exposed to the ocean surface (Fig. 6b), while the others were classified as the inland cliffs that was sheltered by vegetated mountainous lands (Fig. 6c)1. The studied cliffs were subdivided in accordance with the occurrence and feeding trait of cliff-dwelling goat populations (Fig. A1) to address the effect of grazing by the introduced herbivores (Table 2). Here, feeding traits and damages by goats in the UAV image data (e.g., Fig. A1a, b) were considered as the occurrence of the cliff-dwelling goat populations, to minimize potential bias due to the temporal absence of the herbivores during the UAV investigations. Height of the studied cliffs were 20–290 m for the coastal cliffs and 20–140 m for the inland cliffs, respectively (Table 2).
UAV image was sampled three times (April, June, and August 2023) for each cliff to collect clear morphological data of the detected plant species. The UAV image was taken through the multiple diagonal transects on cliff face area (Fig. 7a), as described by30. A wide-angle lens photograph was taken for each sampling point at 3–6 m away from the cliff surface (safe distance to protect UAV)8 (Fig. 7b), and additional photographs were taken using the telephoto lens for any plants requiring the detailed species identification (Fig. 7c). Then, the UAV flew 10 m diagonally upward following the transects and hovered until stabilization for image samplings at the next sampling point. Such procedure was continued unless the UAV reached the cliff top area.
Species identification and categorization
Species identification for UAV images was based on Zhou et al.8 and Kim et al.30. Three regional botanical experts independently identified the plant species within each wide-angle lens image by using the associated telephoto lens images. The experts primarily identified the detected plants into the species level; however, if there was no clear morphological trait data during all three sampling times, the taxa were classified up to the genus level only (1 species in the present study, Table A1). When the experts recorded different names for the same species, the morphological traits from the telephoto lens images were rechecked according to the botanical specimen studies on Gageodo52,53,54 and the standard illustration of Korean plants57,58. International Plant Name Index (IPNI) and World Checklist of Selected Plant Families (WCSP) were used for the taxonomy and nomenclature of the detected plant species. In addition, lifeform of the detected plant species was divided into herbaceous (annual or perennial), fern, and woody (deciduous or evergreen) species as described by the latest study on Gageodo51 for ease of direct comparisons. The occurrence of endangered species was checked using the list of the Ministry of Environment of South Korea, and Alien (archaeophyte and naturalized plants of Kang et al.59) and Korea endemic60 species were informed as well (Table A1).
Data analyses
Incidence ratio was calculated to quantify the relative abundance for each plant species as follows8,30:
where Ix and Itotal represent the number of incidences of plant individuals for a given species and total number of plant incidences throughout all analyzed UAV images (coastal cliff: 3841, inland cliff: 4620). In the present study, such incidence of plant individuals was counted once for each species per each wide-angle lens photograph (i.e., sampling point) because UAV image data provided no belowground information to confirm the actual number of individuals 8. Shannon diversity index was also estimated using the vegan61 package in R 4.2.1 (R Core Team, 2022) to enable direct comparisons of the obtained alpha diversity with traditional studies regarding plant diversity.
Normality and homoscedasticity were tested by Shapiro–Wilk and Leven’s tests for the number of detected species and the proportions of alien species and each lifeform, given the unbalanced experimental design (n = 8 for ungrazed coastal cliffs, 9 for grazed coastal cliffs, 8 for ungrazed inland cliffs, and 5 for grazed inland cliffs) (α = 0.05). Leven’s test confirmed the homogeneity of variance for all variables (P > 0.05); however, the proportion data were square root-transformed for normalization considering the result of Shapiro–Wilk test (P < 0.05). Then, generalized linear mixed model (GLMM) and Tukey’s HSD test were conducted to assess the effects of the two categorical variables (cliff type and grazing), one continuous variable (cliff height), and their interactions on the number of detected species and the proportions of alien species and each lifeform (α = 0.05). Pearson correlation and linear regression were also used to describe any pairwise relationships (α = 0.05). These analyses were performed using the Agricolae62 package in R 4.2.1 (R Core Team, 2022).
Taxonomic beta diversity with Jaccard dissimilarity index was estimated using the binomial occurrence data (absence: 0, presence: 1) of each species. Principal coordinates analysis (PCoA) with Jaccard dissimilarity was carried out to display multivariate patterns in the species composition and beta diversity. Permutational multivariate analysis of variance (PERMANOVA) and permutational analysis of multivariate dispersion (PERMDISP) with 9999 permutations were also conducted to evaluate the effects of cliff type, cliff height, and grazing on multivariate group centroids and dispersions (α = 0.05). Moreover, turnover (βJTU) and nestedness-resultant (βJNE) components within the beta diversity were separated to reveal the ratio between the dissimilarities associated with species replacements and losses. These analyses on the species composition and beta diversity were performed using vegan61, ecodist63, pairwiseAdonis64, and betapart65 packages in R 4.2.1 (R Core Team, 2022).
Data availability
Raw data used for statistical analyses are available from the first and corresponding authors. Full list of detected plant species is available at Table A1 in the attached supplementary materials file.
References
Larson, D. W., Matthes, U. & Kelly, P. E. Cliff Ecology: Pattern and Process in Cliff Ecosystems (Cambridge University Press, 2000).
Davis, P. H. Cliff vegetation in the eastern Mediterranean. J. Ecol. 39, 63–93 (1951).
Goldsmith, F. B. The sea-cliff vegetation of Shetland. J. Biogeogr. 2, 297–308 (1975).
Goldsmith, F. B. Interaction (competition) studies as a step towards the synthesis of sea-cliff vegetation. J. Ecol. 66, 921–931 (1978).
Graham, L. & Knight, R. L. Multi-scale comparisons of cliff vegetation in Colorado. Plant Ecol. 170, 223–234 (2004).
Haig, A. R., Matthes, U. & Larson, D. W. Effects of natural habitat fragmentation on the species richness, diversity, and composition of cliff vegetation. Can. J. Bot. 78, 786–797 (2000).
Wilson, J. B. & Cullen, C. Coastal cliff vegetation of the Catlines region Otago, South Island, New Zealand. N. Z. J. Bot. 24, 567–574 (1986).
Zhou, H. et al. Opening a new era of investigating unreachable cliff flora using smart UAVs. Remote Sens. Ecol. Conserv. 7, 638–648 (2021).
Vandvik, V. et al. Plant traits and associated data from a warming experiment, a seabird colony, and along elevation in Svalbard. Sci. Data 10, 578 (2023).
Zwolicki, A., Zmudczyńska-Skarbek, K., Weydmann-Zwolicka, A. & Stempniewicz, L. Ecological niche overlap in the Arctic vegetation influenced by seabirds. Sci. Rep. 13, 4405 (2023).
Mugnai, M. et al. Impacts of the invasive alien Carpobrotus spp. on coastal habitat on a Mediterranean island (Giglio Island, Central Italy). Plants 11, 2802 (2022).
Clark-Tapia, R. et al. Effects of habitat loss on the ecology of Pachyphytum caesium (Crassulaceae), a specialized cliff-dwelling endemic species in Mexico. Diversity 13, 421 (2021).
Watts, S. H., Griffith, A. & Mackinlay, L. Grazing exclusion and vegetation change in an upland grassland with patches of tall herbs. Appl. Veg. Sci. 22, 383–393 (2019).
March-Salas, M., Lorite, J., Bossdorf, O. & Scheepens, J. F. Cliff as priority ecosystems. Conserv. Biol. 37, e14166 (2023).
Boggess, L. M., Harrison, G. R. & Bishop, G. Impacts of rock climbing on cliff vegetation: A methods review and best practices. Appl. Veg. Sci. 24, e12583 (2021).
Aho, K., Weaver, T. & Eversman, S. Water and substrate control of cliff communities: Patterns among species and phyla. J. Veg. Sci. 25, 1381–1393 (2014).
Harrison, G. R., Boggess, L. M., Budke, J. M. & Madritch, M. D. Rock-climbing shifts cliff-face vegetation community composition based on site characteristics. Appl. Veg. Sci. 25, e12667 (2022).
Jung, S. H., Kim, A. R., Lim, B. S., Seol, J. W. & Lee, C. S. Spatial distribution of vegetation along the environmental gradient on the coastal cliff and plateau of Janggi peninsula (Homigot), southeastern Korea. J. Ecol. Environ. 43, 14 (2019).
Mifsud, S. Distribution of some rare or endemic chasmophytic and rupestral species growing along the coastal cliffs of the Maltese Islands. Webbia 68, 35–50 (2013).
Hepenstrick, D., Bergamini, A. & Holderegger, R. The distribution of climbing chalk on climbed boulders and its impact on rock-dwelling fern and moss species. Ecol. Evol. 10, 11362–11371 (2020).
March-Salas, M. et al. Rock climbing affects cliff-plant communities by reducing species diversity and altering species coexistence patterns. Biodivers. Conserv. 32, 1617–1638 (2023).
Sun, Z. et al. UAVs as remote sensing platforms in plant ecology: Review of applications and challenges. J. Plant Ecol. 14, 1003–1023 (2021).
Gao, D.-Z. et al. A new approach to surveying cliff-dwelling endangered plants using drone-based nap-of-the-object photography: A case study of Clematis acerifolia. Glob. Ecol. Conserv. 49, e02769 (2024).
Li, M. et al. A novel method for cliff vegetation estimation based on the unmanned aerial vehicle 3D modeling. Front. Plant Sci. 13, 1006795 (2022).
Reckling, W., Mitasova, H., Wegmann, K., Kauffman, G. & Reid, R. Efficient drone-based rare plant monitoring using a species distribution model and AI-based object detection. Drones 5, 110 (2021).
Strumia, S., Buonanno, M., Aronne, G., Santo, A. & Santangelo, A. Monitoring of plant species and communities on coastal cliffs: Is the use of unmanned aerial vehicles suitable?. Diversity 12, 149 (2020).
Aucone, E. et al. Drone-assisted collection of environmental DNA from tree branches for biodiversity monitoring. Sci. Robot. 8, eadd5762 (2023).
La Vigne, H. et al. Collecting critically endangered cliff plants using a drone-based sampling manipulator. Sci. Rep. 12, 14827 (2022).
Liu, Z. et al. Mini-drone assisted tree canopy sampling: A low-cost and high-precision solution. Front. Plant Sci. 14, 1272418 (2023).
Kim, S. et al. Piloting an unmanned aerial vehicle to explore the floristic variations of inaccessible cliffs along island coasts. Drones 7, 140 (2023).
Niwa, H. & Hirata, T. A new method for surveying the world’s smallest class of dragonfly in wetlands using unoccupied aerial vehicles. Drones 6, 427 (2022).
Socolar, J. B., Gilroy, J. J., Kunin, W. E. & Edwards, D. P. How should beta-diversity inform biodiversity conservation?. Trends Ecol. Evol. 31, 67–80 (2016).
Lee, M., Kim, S. & Jung, A. H. Distribution patterns of halophytes in the coastal area in Korea. J. Korean Soc. Oceanogr. 24, 139–159 (2019) (in Korean with English abstract).
Alessio, P. & Keller, E. A. Short-erm patterns and processes of coastal cliff erosion in Santa Barbara, California. Geomorphology 353, 106994 (2020).
Young, A. P. et al. Three years of weekly observations of coastal cliff erosion by waves and rainfall. Geomorphology 375, 107545 (2021).
Garner, K. L. et al. Impacts of sea level rise and climate change on coastal plant species in the central California coast. PeerJ 3, e958 (2015).
Anderson, M. J., Ellingsen, K. E. & McArdle, B. H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 9, 683–693 (2006).
Canepuccia, A. D., Pérez, C. F., Farina, J. L., Alemany, D. & Iribarne, O. O. Dissimilarity in plant species diversity between salt marsh and neighboring environments decreases as environmental harshness increases. Mar. Ecol. Prog. Ser. 494, 135–148 (2013).
Wrubel, E. & Parker, V. T. Local patterns of diversity in California northern coastal scrub. Ecol. Evol. 8, 7250–7260 (2018).
Yoo, H.-S. Gelogy of Kago-do (Soheogsan-do). J. Korean Isl. 7, 115–122 (1996) (in Korean with English abstract).
Melo, E. A. & Waechter, J. L. Beta diversity patterns of Bromeliaceae growing on rocky cliffs within the Atlantic Forest in southern Brazil. Biota Neotrop. 20, e20190846 (2020).
Anderson, M. J. et al. Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist. Ecol. Lett. 14, 19–28 (2011).
Cai, Y. et al. The fertile island effect collapses under extreme overgrazing: Evidence from a shrub-encroached grassland. Plant Soil 448, 201–212 (2020).
Drissen, T. et al. Plant composition and diversity in a semi-natural Mediterranean island landscape: The importance of environmental factors. Plant Biosyst. 153, 756–766 (2019).
Thomson, D. M. et al. Diverse native island flora shows rapid initial passive recovery after exotic herbivore removal on Santa Rosa Island, California. Biol. Invasions 24, 1635–1649 (2022).
Sangüesa-Barred, G. et al. Vertical cliff harbor millennia-old junipers in the Canary Islands. Ecology 103, e3633 (2022).
Dimitrakopoulos, P. G., Koukoulas, S., Michelaki, C. & Galanidis, A. Anthropogenic and environmental determinants of alien plant species spatial distribution on an island scale. Sci. Total Environ. 805, 150314 (2022).
Ren, J. et al. An invasive species erodes the performance of coastal wetland protected areas. Sci. Adv. 7, eabi8943 (2021).
Martin, P. H., Canham, C. D. & Marks, P. L. Why forests appear resistant to exotic plant invations: Intentional introductions, stand dynamics, and the role of shade tolerance. Front. Ecol. Environ. 7, 142–149 (2008).
Son, D., Waldman, B. & Song, U. Effects of land-use types and the exotic species, Hypochaeris radiata, on plant diversity in human-transformed landscapes of the biosphere reserve, Jeju Island, Korea. Plant Divers. 45, 685–693 (2023).
Kim, H.-H., Kim, D. B., Song, H. H., Hwang, G. Y. & Kong, W. S. Phytogeographical characteristics of outermost islands in the Korean Peninsula. J. Korean Geogr. Soc. 53, 117–132 (2018) (in Korean with English abstract).
Yun, K.-W., Hwang, Y., So, S. & Kim, M. Flora of island Gageo in Jellonamdo, Korea. Korean J. Environ. Ecol. 26, 139–155 (2012) (in Korean with English abstract).
Kim, J.-H., Nam, G.-H. & Kim, J.-S. The vascular plants in Is. Gageo (Jeollanam-do). J. Environ. Sci. 21, 437–450 (2012) (in Korean with English abstract).
Yang, S.-G. et al. Floristic study of Gageodo in Korea. Korean J. Plant Res. 26, 597–612 (2013) (in Korean with English abstract).
Lee, J. et al. Vegetation structures of warm temperate evergreen broad-leaved forest in Gageodo, Korea. J. Korean Soc. Environ. Restor. Technol. 13, 75–86 (2010) (in Korean with English abstract).
Lee, J.-Y., Lee, S.-M., Kim, H.-S., Oh, J.-G. & Myeong, H.-H. A study on the current distribution of vegetation of Gageodo, Jeollanamdo. J. Korean Isl. 27, 195–210 (2015) (in Korean with English abstract).
Lee, T. B. Coloured Flora of Korea (Hyangmun Publ. Co., 2003) (in Korean).
Lee, W. T. Standard Illustrations of Korean Plants (Academy Press, 1996) (in Korean).
Kang, E. S. et al. Comprehensive review about alien plants in Korea. Korean J. Plant Taxon. 50, 89–119 (2020) (in Korean with English abstract).
Chung, G. Y. et al. A checklist of endemic plants on the Korean Peninsula II. Korean J. Plant Taxon. 53, 79–101 (2023).
Oksanen, J. et al. The vegan package. Community Ecol. Package 10, 631–637 (2007).
de Mendiburu, F. & Simson, R. Agricolae—ten years of an open source statistical tool for experiments in breeding, agriculture and biology. PeerJ Preprints 3, e1404v1 (2015).
Goslee, S. C. & Urban, D. L. The ecodist package for dissimilarity-based analysis of ecological data. J. Stat. Softw. 22, 1–19 (2007).
Arbizu, P. M. PairwiseAdonis: Pairwise multilevel comparison using adonis. R Package Version 0.4 (2020).
Baselga, A. & Orme, C. D. L. Betapart: An R package for the Study of beta diversity. Methods Ecol. Evol. 3, 808–812 (2012).
Acknowledgements
This study was supported by the National Institute of Ecology (NIE-B-2024-49) of South Korea.
Author information
Authors and Affiliations
Contributions
Conceptualization: S.K. Formal analysis: S.K., C.W.L. Funding acquisition: S.K. Investigation: S.K., C.W.L., H.J.P. Methodology: S.K., C.W.L. Supervision: S.K., C.W.L. Writing – original draft: S.K. Writing – review and editing: C.W.L., H.J.P., J.E.H., H.B.P, Y.J.Y., Y.J.K.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kim, S., Lee, C.W., Park, HJ. et al. UAV telephotography elucidates floristic variability and beta diversity of island cliffs under grazing interventions. Sci Rep 14, 15465 (2024). https://doi.org/10.1038/s41598-024-66446-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-66446-7
- Springer Nature Limited