
Abstract
Record mean sea surface temperatures (SST) during the past decades and marine heatwaves have been identified as responsible for severe impacts on marine ecosystems, but the role of changes in the patterns of temporal variability under global warming has been much less studied. We compare descriptors of two time series of SST, encompassing extirpations (i.e. local extinctions) of six cold-temperate macroalgae species at their trailing range edge. We decompose the effects of gradual warming, extreme events and intrinsic variability (e.g. seasonality). We also relate the main factors determining macroalgae range shifts with their life cycles characteristics and thermal tolerance. We found extirpations of macroalgae were related to stretches of coast where autumn SST underwent warming, increased temperature seasonality, and decreased skewness over time. Regardless of the species, the persisting populations shared a common environmental domain, which was clearly differentiated from those experiencing local extinction. However, macroalgae species responded to temperature components in different ways, showing dissimilar resilience. Consideration of multiple thermal manifestations of climate change is needed to better understand local extinctions of habitat-forming species. Our study provides a framework for the incorporation of unused measures of environmental variability while analyzing the distributions of coastal species.
Similar content being viewed by others

Introduction
Ocean warming has been increasing at an unprecedented rate since the 1950s and this pattern is projected to continue in the future with uncertain consequences on centennial timescales1. In this context, range boundaries of marine species are changing2,3, and the community composition of marine ecosystems has been altered4. Given that ocean warming is not uniformly occurring across the oceans at a constant pace1,5, it is unclear in which direction species and communities will shift6 or if they will be able to cope with climate change through plasticity/adaptation strategies7. The patterns of fluctuation around average sea warming trends are also changing, with recent rises of extreme events. Marine heatwaves (MHWs) were longer and more frequent over the past century8 and this trend is projected to further increase9,10. MHWs have been identified as responsible for alterations in species distributions and biodiversity patterns3,11, mass mortalities12, biological changes at individual, population and community levels13, and various impacts on ecosystem services14. However, most of these effects have been observed in regions where MHWs frequently reach categories higher than “moderate” (category I according to Ref.15). Since species distribution predictions are mostly based on warming trends, the need for improved understanding of the implications of extreme events and long-term pervasive climate change has become imperative to predict their impacts on marine species11,16.
Much less is known about the influence of temporal patterns of temperature variability on species range shifts. Marine organisms regularly experience environmental variability, prompting questions about how these dynamics may affect species’ susceptibility to climate change. In disturbed ecosystems, a range of variability patterns in environmental factors may turn into stressors. For example, the temporal variance is known to interact with the mean intensity of climatic events to modify the diversity and structure of benthic assemblages17,18. As a result, there is a growing interest among aquatic ecologists in the effects of changes in the patterns of variability under climate warming, and research including descriptors for the dynamics and complexity of abiotic factors is rising19,20. Among the different measures of variability suitable to examine ecological changes, the environmental predictability, estimated by both the seasonality and the colour of environmental noise, has long been considered a driver of life-history evolution, population dynamics and biodiversity patterns (Refs.21,22 and references therein). Both components provide insights of different aspects of predictability, while the seasonality is associated to the regularity of fluctuations in data over seasons (i.e. a year; Ref.22) the environmental noise (i.e. environmental colour, colour of noise) estimates the temporal autocorrelation between successive values in the time series23. Temporal variability can be also assessed using common indices such as the coefficient of variation (CV), and the consecutive disparity index (D) that considers the chronological order of time series data24. Finally, changes in the skewness (the degree of asymmetry of probability distributions estimated from time series data) can be an effective early warning signal of ecosystem regime shifts25. However, the application of these metrics, including components of variability, has been scattered in ecological systems, and they have never been utilized jointly to evaluate their impact on the range shifts and local extinction of species.
Regarding how species are impacted by thermal fluctuations, responses vary widely among marine organisms, which have been observed to shift in phenology and/or acclimate, show tolerance, and accumulate positive or negative effects20,26. Species responses to same patterns of temporal variability and environmental events are probably modulated by species functional traits, e.g. life-history and ecophysiological responses. The relevance of using functional traits to investigate which marine species will be more vulnerable to different characteristics of MHWs has already been highlighted27. Species with distinct lifespan and generation times experience the same patterns of environmental fluctuations differently and so, their different “ecological memories” would influence their responses to warming28. Also, between-species variation in physiological thresholds may arise even in organisms that currently share similar geographic distributions, possibly because of evolutionary legacies29. To anticipate how projected ocean warming will affect marine ecosystems it is necessary to better understand the influence of species’ trait diversity in their responses to environmental changes.
At a rapidly changing environment, there are three distinct outcomes for populations: persistence through adaptation to new conditions at existing places, persistence through migration to other places tracking the same conditions, and extirpation (i.e. local extinction)30. Currently, the responses of marine canopy-forming species to thermal stressors are not fully understood and causal factors leading to local extinctions (i.e. extirpations) remain speculative. Habitat forming seaweeds (mostly species from the orders Fucales, Laminariales and Tilopteridales), provide the foundation of highly diverse and productive coastal ecosystems. These large brown algae, commonly named fucoids and kelps, form intertidal and subtidal forests in temperate and subpolar rocky reefs, being the dominant primary producers and supplying food and shelter for a diverse assemblage of associated species31. These marine macrophytes are globally in decline, particularly the populations at their warm trailing range edges14. There are well documented range contractions of native macroalgae, recognizing the crucial role played by the warming trend in coastal sea surface temperature (SST) and MHWs (e.g.3,14,32,33,34). However, there are still a lot of unanswered questions surrounding the SST components particularly responsible for the local extinction or persistence of macroalgae with similar worldwide distributions. Distinct responses of species may result from differences in their thermal thresholds and thus in their sensitiveness to the absolute magnitude of change, but sometimes the highest temperatures do not trigger the strongest biological impact on macroalgae, suggesting that species resilience is dependent on combinations between prolonged warming and short-term stress events35. Still, other components describing the variation in the long-term temporal distribution of SST (i.e. warming trends, SST predictability, etc.) and temporal changes in frequency, intensity, and duration of extreme events remain fairly unexplored for macroalgae. For example, marine cold spells (MCSs) are often understudied despite their impacts on marine ecosystems and foundational species36. Also, extreme events can have different impact depending on the preceding temperatures to each event (“preconditioning factors”; Ref.15), and on the biological processes related to the macroalgae life cycles at each season.
Here, we investigated the sea surface temperature-related factors that explain persistence and local extinction (i.e. extirpation) of six macroalgae species at their warm range edges, in the North of the Iberian Peninsula. In this region, there is a longitudinal SST gradient from cooler (West) to warmer (East) waters and the recent intertidal and subtidal rocky shore macroalgae retractions (mostly in the eastern coast) have been attributed primarily to changes in SST (e.g.33,34,37,38,39,40,41,42). We decomposed the effects of climate change on macrophytes extirpations using SST time series data corresponding to a baseline survey and a resurvey of macroalgae. To account with the previously mentioned factors, we analyzed the effects of: (i) annual and seasonal warming trends; (ii) annual and seasonal frequency, intensity, and duration of MHWs and MCSs; (iii) temporal variability measures; and (iv) species thermal thresholds for survival. This study seeks to determine which components of thermal temporal patterns are driving range shifts (local extirpations) and persistence of macroalgae along a longitudinal SST gradient at their warm range edges. We also determined whether every species is responding in the same way attending to its life cycle and thermal tolerance. This study aims to provide new insights into the understanding of the effects of climate change on macrophytes, with the incorporation of previously unused measures of thermal variability.
Results
In the north of the Iberian Peninsula, the retraction of six cold-temperate brown algae species was examined during two time spans, the baseline and resurvey periods (Fig. 1). Depending on the species, 29 to 39 from the initial 85 variables were not correlated and used for the hierarchical partitioning analyses. After running the iterative hierarchical partitioning analyses, from 8 to 11 variables were used in the final hierarchical partitioning analysis computed for each species. The most relevant variables discriminating between persistence and extirpation for each macroalgae were considered to be those showing an independent contribution above the median (I > 0.5 percentile) in the final hierarchical partitioning analysis (Fig. 2; see Fig. S1 Supporting information for the complete results). Change in autumn mean SST was the only variable found relevant for all species. Two variables measuring the temporal variability of SST, change in seasonality and change in skewness, also showed a high I value for five and four species, respectively. The rest of variables discriminated differently between persistence and extirpation depending on the species. Many variables related to changes in MHWs and MCSs showed I values below the 0.5 percentile, with some exception, i.e. the MHWs mean duration (Fig. S1; Supporting information). The dendrogram showed L. ochroleuca and L. hyperborea were related to each other (i.e. their clusters were close; Fig. 2), suggesting extirpation/persistence patterns were explained by similar factors, which differed from those of other species. For example, the thermal thresholds were relevant discriminants only for Laminaria species (Fig. 2). Similarities were also found between F. serratus and H. elongata, with MCSs in summer being relevant for only those two species, and in another cluster formed by S. polyschides and F. vesiculosus.

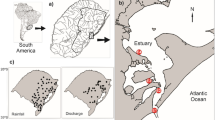
Persistence and local extinction (extirpation) of macroalgae populations between two surveyed periods along the Northern Spanish coast. Populations of Fucus serratus, F. vesiculosus, Himanthalia elongata, Laminaria hyperborea, L. ochroleuca and Saccorhiza polyschides were surveyed at two periods: baseline (1980–1990) and resurvey (2013–2015) and persistence/extirpation of populations analyzed using sea surface temperature series data. Distributional limits in the resurvey period are shown as vertical bars (for S. polyschides, there were still a few sparsely populations east of the given location). The change in the variables (mean sea surface temperature (SST) of autumn, seasonality, and skewness) represents the anomaly (difference) between the two periods.

Most relevant variables discriminating between sites with persistent and extirpated populations for each macroalgae species. Heatmap shows the independent contribution (I) of variables obtaining an I value above the median (0.5 percentile) after the iterative hierarchical partitioning analysis. The light colors represent high values of I, while the dark colors represent low values. The column dendrogram shows clustering of species whose extirpation/persistence patterns are explained by similar factors. See Table S1 in Supporting Information for a description of the variables, and Fig. S1 in Supporting information for complete results also showing the variables with I values below the 0.5 percentile in the hierarchical partitioning analysis. SST sea surface temperature.
The two-dimensional NMDS provided a reliable representation (stress value = 0.084; Fig. 3) of persistence and extirpation of macroalgae populations across the environmental space, defined by all the variables selected by the iterative process above mentioned and used in the final hierarchical partitioning analysis (Fig. S2; Supporting information). Axis 1 of the NMDS (NMDS1) discriminated persistent from extirpated populations of the six macroalgae species. Regions of extirpation were mainly related with greater temporal changes in seasonality and autumn mean SST, as well as with lower anomalies in skewness with respect to persistence grids (Figs. 3, 4, Table S2; Supporting information).

Non-metric multidimensional scaling (NMDS) ordination of the sites (i.e. locations, grids) where extirpation or persistence has occurred over the baseline and resurvey periods in the Northern Spanish coast. Smooth surfaces of the change in autumn mean sea surface temperature, seasonality and skewness fitted by means of generalized additive models showed a gradient direction able to explain the extirpation-persistence distribution (e.g. extirpated populations tend to increase when the seasonality anomaly increases over time). Macroalgae populations studied are Fucus serratus, F. vesiculosus, Himanthalia elongata, Laminaria hyperborea, L. ochroleuca and Saccorhiza polyschides. See Supporting information for a representation showing the vectors of all the variables used (Fig. S2), a description of the variables (Table S1), and the NMDS scores (Table S2). SST sea surface temperature.

Change in autumn mean SST, seasonality and skewness between the baseline (1982–1990) and resurvey (1991–2015) periods at extirpated and persistent populations of macroalgae. Anomalies of variables were standardized into z-scores to provide a better comparison. For each species, circles indicate the mean z-score of all sites and error bars indicate standard errors. SST sea surface temperature.
Variables related to changes in extreme events (MHWs and MCSs) between the baseline and resurvey periods showed relevance discriminating persistence and extirpation in the hierarchical partitioning analyses of some species (Fig. 2, Fig. S1; Supporting information) and in the NMDS (Fig. 3, Table S2; Supporting information). Remarkably, extirpated populations showed negative anomalies (reductions) in mean duration of MHWs between the analyzed time periods, while most persistent populations had positive anomalies (increases, Fig. 5). MHWs frequency and intensity also showed reductions in extirpated populations (Figs. S2, S3; Supporting information). In general, MCSs showed less relevant relationships with extirpation/persistence patterns (Figs. 2, 3). The anomalies in MCSs were positive in areas with local extinctions while in persistence areas no changes or very reduced anomalies were detected (Fig. S3; Supporting information).

Laminaria ochroleuca populations exhibiting the most differing anomalies in mean duration (days/event) of marine heatwave (MHW) events. The most positive change between periods in MHW mean duration was found in persistent populations in La Coruña (Galicia; blue circle), where the MHW event with the highest cumulative intensity occurred in September 2014, and the event with maximum intensity in May 2008 (category II). On the other hand, negative anomalies in MHW mean duration were found in extirpated populations in Pais Vasco (red circle), which showed the MHW with highest cumulative intensity in March 1990, and the maximum intensity in September 2006 (category II). Height of the lolliplots represent the duration of the MHW event and the color shows the events’ cumulative intensity. Bottom panels illustrate the MHW with the highest values of maximum intensity detected from 1982 to 2015 in the persistence (left; max. intensity: 2.8683, date: 2008-05-19) and extirpation (right; max. intensity: 4.1367, date: 2006-07-18) regions.
In average, extirpated populations suffered a higher positive anomaly between periods in autumn mean SST (mean ± SD = 0.36 °C ± 0.017) than persistent populations (mean = 0.23 °C ± 0.03) (Fig. 4, Table S3; Supporting information). Overall, skewness was positive at the studied area over time (mean skewness baseline = 0.36 ± 0.08; mean skewness resurvey = 0.37 ± 0.07). However, extirpation regions experimented a lower change in skewness (mean = 0.023 ± 0.016) than persistent populations (mean = 0.067 ± 0.004) (Figs. 3, 4, 6, Fig. S3, Table S3; Supporting information). Anomaly in seasonality was positive in extirpated populations (mean = 1.39 ± 0.34) but slightly negative in persistence regions (mean = −0.29 ± 0.26) (Figs. 3, 4, 6, Fig. S3, Table S3; Supporting information). D index was only found relevant for H. elongata (Fig. 2). D is higher in the persistent populations, and it has been decreasing over time in all the study area, but particularly in the extirpation zone, indicating a reduction in disparity (i.e. SST data becomes less pulsed over time) (Fig. 6, Figs. S3, S4; Supporting information). We did not find the colour of environmental noise to be a relevant discriminant of persistence/extirpation.

Performance comparison of variability metrics at locations where macroalgae persistence and local extirpation occurred between the baseline (1982–1990) and resurvey (1991–2015) periods. SST sea surface temperature.
Discussion
Our study identified associations between the persistence and extirpation of macroalgae populations and variables related to gradual warming and temporal variability of sea surface temperature (SST). We found local disappearance of populations are associated to the temporal variability in SST, estimated with variables not typically considered in studies on how macroalgae are affected by climate change. Populations that suffered extirpation were in regions with greater SST warming in autumn (NE Spain, i.e. the inner Bay of Biscay), and have experienced an increase in seasonality (while most persistent populations showed a decrease), and a decrease in skewness over time (Figs. 3, 4). Species responses to environmental change may be complex and dependent on distinct aspects of a factor, involving not only its trend but also the variability19,20,35. Integrating historical occurrence data with two surveys at different periods allowed us to decompose the effects of SST on distributional shifts at the southern European range edge of these habitat forming species. The study area (Northern Spanish coast), with a SST gradient longitudinally oriented at a similar latitude, provides an ideal testing ground for a fine analysis of the effects of SST, as environmental changes presumably occur faster here than along a latitudinal gradient.
Autumn SST anomalies between the resurvey and historical periods were found to be a relevant factor explaining persistence/extirpation of all species (Figs. 1, 2). This season has been seldom considered when analyzing species distributions and the ecological impacts of climate change on marine macrophytes. Winter and/or summer sea temperatures are the commonly referred drivers of geographical boundaries of North Atlantic macroalgae by limiting growth and reproduction or by lethal effects43,44,45. However, autumn may be more important than previously assumed, in particular at the warming range margin of cold-temperate macroalgae. SST is increasing at higher rates in autumn than in other seasons in NW Iberia46 and autumn SST was found a relevant factor determining the fine-grain distribution of F. serratus in Atlantic rias of NW Spain47. Warmer autumns would extend the heat stress already experienced by seaweeds during the summer, with sublethal and lethal consequences in individuals and populations. Shorter reproductive periods, mainly in spring-early summer, and more variable reproductive output were detected in marginal populations of Fucus species in N Spain and Portugal48,49,50. The success of new macrorecruits in late summer-autumn may become critical for the persistence of fucoids with a short lifespan (2 to few years) such as H. elongata and Fucus spp., particularly if the reproductive window narrows. Furthermore, frond damage and weakening observed in adult plants of already extirpated populations of F. serratus in N Spain during autumn (authors, pers. observation) can cause individuals to be detached by storms more easily. In the annual kelp S. polyschides, sporophyte growth continues until autumn, when biomass in N Spain reaches its highest; afterwards spore release, germination and gametophyte formation occur42,51. Thus, prolonged, cumulative heat stress in autumn may reduce or collapse S. polyschides sporophyte growth and reproduction. Greater tissue damage in autumn-winter than in spring-summer was also detected in sporophytes of intertidal populations of a Laminaria species in British shores, suggesting chronic effects of high air and seawater temperatures during summer-autumn52.
Interestingly, range contractions of these seaweeds were also explained by anomalies in SST temporal variability, particularly by seasonality, and skewness. Seasonality (i.e. the predictability in timing and magnitude of seasonal fluctuations) was comparatively higher in regions with extirpations (eastern shores) than in regions of persistence (westernmost coast) both during baseline and resurvey periods. But the variance in seasonality increased even more across time where extirpations occurred, i.e. increasing the amplitude of seasonal SST fluctuations. This increase in seasonality implies higher predictability but also a greater exposure to thermal extremes and fluctuations that might have affected macroalgae populations (Figs. 3, 4, 6). On the other hand, persistence regions had lower seasonality also showing a tendency to become less seasonal over time (i.e. less regular and predictable), implying a smaller oscillation of SST which confers a temperature-buffered character to these localities. A major component of the NW Iberian Peninsula’s longitudinal SST gradient is the presence of intense spring-summer upwellings53. The lower seasonality in this region is probably related to the reduction in the amplitude of the SST seasonal cycle caused by the rise of cold and nutrient-rich water to the surface in spring-summer, as observed in this and other upwelling systems54,55. Further, the decrease in seasonality with time might suggest an intensification of the upwelling from 1982–1990 to 1991–2015 that could have favored the persistence of macroalgae populations in NW Iberian Peninsula. Nevertheless, the detected reduction in seasonality was small, and the present and future trends of the coastal upwelling in NW Iberian Peninsula are currently under discussion. Several studies have indicated a weakening of the upwelling in this region over the last decades (e.g.56,57) but not others58. According to Ref.59, future sea surface warming will enhance the stratification of the upper layers, diminishing upwelling and lowering the inflow of nutrients from the surface. Future monitoring should assess if upwelling trends could or not lead to further macroalgae extirpations. Regarding skewness, in the study area there is a general pattern in which SST is right skewed (positively skewed, i.e. with dominance of low SST values). Nevertheless, the persistence region showed an increase in skewness across time; while data are balanced towards higher SST values over the years (i.e. skewness decrease) in the extirpation locations. Hence, the increase in skewness in the NW Iberian Peninsula reflects a predominance of low SST values over time that benefited the persistence of large, brown canopy-forming algae of cold-temperate affinities in this region. Conversely, skewness reduction in the extirpation shorelines illustrates a prevailing pattern of rising SST, that could have contributed to the local extinctions of these six species. Changes in the asymmetry of time series data, quantified by changes in the skewness, have previously been referred to as an indicator of regime shifts, and together with other changes in variability can alert us to identify a reduction of resilience in the ecosystems25.
Despite pertaining to close taxonomic groups, the analyzed macroalgae, kelps and fucoids, showed different vulnerability to warming effects at their southern limits of distributions in the N Iberian Peninsula, evidencing distinct response to the temperature components (Fig. 2), which may be linked to variations in their traits such as lifespans or thermal tolerances. We found similarities in the SST components relevant for F. serratus and H. elongata, the two intertidal species that exhibited clear range shifts in the studied area. As explained above, both species have short lifespans (H. elongata is biennial and F. serratus life expectancy reaches up to 4–5 years in wave-sheltered places60). Their apparent sensitivity to environmental changes could be linked to these short life spans and their dependence on sexual reproduction, since adult plants of F. serratus also lack vegetative regeneration61. F. vesiculosus and S. polyschides are resilient species within fucoids and kelps, respectively, still showing disperse populations along the studied region, and both species seemed affected by the same SST factors. The higher resilience of F. vesiculosus relies in its higher thermal tolerance compared to other fucoids such as F. serratus62 and its ability to resprout from the holdfast63. The annual sporophytes of S. polyschides exhibit a greater ability to acclimatize to changing environmental conditions than those of perennial Laminaria species, in particular L. ochroleuca, which presented a similar geographic distribution64. Also, the bigger surface area of adhesion of the gametophyte of the annual kelp might allow to remain viable for more than one year, facilitating fast recruitment65. All these aspects might give S. polyschides lower susceptibility to warming and greater resilience than do the perennial Laminaria kelps. On the other hand, we found thermal thresholds were only relevant for the two studied perennial kelps (Fig. 2, Fig. S5 Supporting information). A recent study in Baja California also found a greater sensitivity of the giant kelp Macrocystis pyrifera to exceeding absolute thresholds than to relative changes in temperature66. The importance of the temporal thermal profile in determining responses of macroalgae, and the specificity of these responses have been recently acknowledged35. Further experimental studies are however necessary to disentangle the traits influencing specific responses.
Contrary to our expectations, changes in extreme events characteristics over time (both MHWs and MCSs) did not support causal inferences regarding persistence/extirpation. We did not find key metrics of extreme events explained the local extirpations, even examining their effect by seasons, as local extinctions occurred with negative anomalies (declines over time) in mean duration, frequency and intensity of MHWs (Fig. 5, Fig. S2; Supporting information). Despite the lack of explanatory capacity of MHWs in our study, we found the extirpation region was already affected by more frequent, intense and longer MHWs than the persistence area during the baseline period (1982–1990, Fig. 5). MHWs have been related to mass mortality events of marine benthic species in the Mediterranean Sea12, Australia11,15, see13 for a review. However, MHWs reached greater categories in these marine locations than in our study area, where only MHWs category II were recorded from 1982 to 2015. In our examined region, Smith et al.13 did not find notable MHW events compared to other temperate oceanic regions. So extreme heat events variation present in our study area could presumably not be sufficient to produce a measurable response in macroalgae. Besides, extreme events detection is known to be affected by the spatial and temporal resolution of SST data15 and shows differences with in situ datasets67. However, other authors68 using in situ nearshore SST time series data from 1998 to 2019 in two coastal areas in the middle Cantabrian Sea, were also unable to identify any significant trends in the frequency and duration of marine heatwaves. In general, the anomalies in marine cold spells (MCSs) were found less relevant discriminating the extirpation-persistence pattern of these populations close to their warmest latitudinal range edge, but further research might indicate if extremely cold water events could be a more relevant factor when analyzing extirpations at their poleward edges. Alongside from finding or not anomalies that could have caused local extinctions of macroalgae, our results raise concerns over the change in the extreme events regime in persistence coastlines. The increase in MHWs duration, frequency, and intensity in the persistence region (Galicia) could reflect an early sign of regime shift for the NW Iberian Peninsula, which has been a persistence region for macroalgae over decades.
This study set out to determine the effects of different components of temperature on macroalgae persistence-extirpation, since SST has been pointed out as the main responsible of seaweed retractions in the N Iberian Coast33,34,37,39,40,42,69, and worldwide (e.g.11,14,32). The relationship between variability of SST and the temporal patterning of occurrence under climate change is still understudied for macroalgae, and our research analyzing rarely used components of variability contributes to our understanding of local extinctions. Despite its exploratory nature, this study found strong associations between changes in SST components and the persistence and extirpation patterns. The persistent populations were benefited by less SST fluctuations and overall colder SST, whilst extirpated populations were affected by rising SST, with increases in autumn anomalies and seasonality. Although we did not find environmental noise to be a relevant explanatory factor, it is possible this variable might discriminate better within wider study areas, by covering all the variation that seawater can show, and covering a wider range of species with different life cycles and lifespan. Further research should be undertaken to investigate other factors not explored in this manuscript such as herbivorous pressure (e.g.70), the role of nutrient supply—a future nutrient depletion is linked to projections of upwelling weakening in the NW Iberian Peninsula59,71—wave action, water turbidity, or the impact of aerial stress in the case of intertidal species, which could also cause synergistic effects with SST. Also note that the biological response of the species can differ between populations and its recovery from stress depend on its continuity (e.g. the number of consecutive days that thermal thresholds were exceeded was not measured). Given expected increase in MHWs extremity with ocean warming9,10, it is important to continue monitoring its effect over time. Future monitoring should assess if the increase in this trend could lead to further macroalgae extirpations. Critical effects of temperature may be cumulative in time, and impacts may involve more than one life stage. Treatments applying a combination of sustained warming and MHWs produced different responses in forest-forming seaweeds35, suggesting the synergistic effects between different components also need further investigation. Temperature is a key factor for habitat-forming macrophytes and this work is a novel approach to assist in our understanding of the role of the different aspects of warming on distributional shifts.
Methods
Study area and biological data
The distribution of six cold-temperate brown algae species in the North of the Iberian Peninsula were compared between two time periods from 1980 to 2015: two species from the Laminariales order (Laminaria hyperborea and Laminaria ochroleuca), one from Tilopteridales order (Saccorhiza polyschides), and three from the Fucales order (Himanthalia elongata, Fucus serratus and Fucus vesiculosus). These macroalgae have two southern boundaries in the Iberian Peninsula, one of them is in N Spain, and they are among the most relevant habitat-formers of the Northern Spanish coast subtidal and intertidal systems. Life cycles of kelps and fucoids clearly differ, since kelps have heteromorphic diplohaplontic cycles, with two alternate stages (the diploid macroscopic sporophyte and the microscopic gametophytes), while fucoids have monophasic life cycles. Lifespan also differ between the studied species. The fucoid H. elongata is biennial72 and Fucus spp. are perennial but with individuals living a few years (in F. serratus up to 5 years but only in sheltered places, see Rees60), while some Laminaria species, such as L. hyperborea have maximum ages around 15 years (see Schiel and Foster31 and references therein). The sporophyte of S. polyschides is annual, but in both annual and perennial species of kelps the microscopic gametophytes have unknown longevity65.
The studied area covered from the Miño River mouth in the south of Pontevedra (41° 52ʹ N, 8° 52ʹ W) to the France-Spain border (43° 22ʹ N, 1° 47ʹ W) (Fig. 1), a region affected by a longitudinal SST gradient from cooler (West) to warmer waters (East) (Fig. S6; Supporting information). We used a georeferenced database compiled from recent field surveys recording the presence of the species in the North coast of Spain, in combination with historical data obtained from the literature and technical reports (for all references, see Table 1 in Casado-Amezúa et al.33 review). The database used is openly available in Zenodo (see Data availability section). To assess the effect of seawater temperature on the distribution of the species, available information of presence/absence for all seaweeds was grouped into two time periods, following Casado-Amezúa et al.33: (i) the baseline period (1980–1990), and (ii) the resurvey in 2013–2015. These periods of biological data were examined using time series data. Since the daily OISST v2.1 data is only available from September 1, 1981 (see “Long term sea surface temperature data” section), we selected January 1, 1982 as the beginning of the SST series data to be analyzed. Thus, we examined the difference between the 1982–1990 SST time series data to describe the baseline period, and the 1991–2015 SST time series data to describe the resurvey period.
The same localities were consistently recorded in both periods at different times of the year during asynchronous visits for the different species. Intertidal species were surveyed at low tides and Laminariales and S. polyschides subtidal shallow-water populations (down to a maximal depth of about 20 m) by snorkelling and diving. The resurvey period represents a snapshot within an almost constant update of the species distributions in N Spain by the research group and a network of collaborators. For instance, in the case of Fucus serratus and Himanthalia elongata two field surveys were carried out in 2004–2006 and 2008–2009 by the research group34,73. All the targeted species gradually disappeared from the early 2000’s onwards in numerous localities of N Spain. Subsequent visits since 2015 to most of the locations confirm the local extinctions, except for S. polyschides, of which reduced populations have been recently found at some of the localities recorded as absences by 2015.
Long term sea surface temperature data
Time series of daily sea surface temperature (SST) for the Northern Spanish coast were retrieved from 1982 to the end of 2015, encompassing the same time periods as the biological data sets. We used the NOAA 1/4° daily Optimum Interpolation SST data (daily OISST v2.1 data74) which is available from September 1981. We examined the difference between the 1982 and 1990 SST time series data to describe the baseline period of seaweed data, and the 1991–2015 SST time series data to describe the resurvey period in which persistence and local extinctions were detected. This long-term climate data combines observations from diverse platforms (buoys, satellites, ships and Argo floats) offering a high quality control (https://www.ncdc.noaa.gov/). Despite being the daily OISST data at a relative coarse resolution and showing limitations in near-coastal zone (e.g. pixels that are distant from the coast), it facilitates comparisons with other related studies on extreme events as it is a standard methodology and improves reproducibility at any geographical space. Besides, daily OISST data offers a high temporal resolution, needed to obtain the recommended 30 year time series data to estimate the climatology15.
To estimate long term warming, we calculated basic statistics from the daily OISST time series for each spatial grid point and time window (baseline and 2015 resurvey, henceforth). We measured the annual and seasonal mean of the SST time series for each pixel at the baseline (1982–1990) and 2015 resurvey (1991–2015). Seasons were related to the Northern Hemisphere, being summer June to August, and winter December to February. Daily SST data were retrieved using the “heatwaveR” package75.
Extreme events detection and characterization
We detected extreme events using the SST data described above and following the consistent existing framework. Marine heatwaves (MHWs) are discrete prolonged anomalously warm water events and were detected according to the definition in Hobday et al.15, which establishes a period of 5 or more days above the 90th percentile threshold of the SST data. Both threshold and the baseline mean vary through the input time series, as they are both calculated within an 11-day window. MHWs are usually described by their frequency, intensity, and duration11,14,15. Furthermore, to test the relationship of trend changes in upwelling events with macroalgae, we examined marine cold-spells (MCSs). MCSs were detected following Schlegel et al.36 which determines the anomaly should persist for at least five days below the 10th percentile SST threshold. Both MHWs and MCSs and their derived metrics were identified using the “heatwaveR” package75. MHWs and MCSs were detected on the entire SST time series data. For each pixel, we detected extreme events (MHWs and MCSs) using the complete daily SST time series data (1982–2015—34 years) as an unique climatology period. Afterwards, we divided the information obtained into two time periods that coupled seaweed data surveys. Thus, we calculated a set of metrics describing the events of each period (1982–1990 and 1991–2015): the yearly frequency, the mean duration per event, the intensity (absolute maximum, mean maximum, and mean intensity), and the mean and maximum cumulative intensity (intended as the integral of intensity over the duration of the extreme event) (see15 for details). These metrics were estimated both as a mean value per year and seasonally for each period.
Sea surface temperature variability and skewness
We calculated variability measures from the daily OISST time series for each spatial grid point and time period. We estimated two components of environmental predictability in each time window and grid cell: the predictability of seasonal trend and the colour of environmental noise21,22 using the “envPred” R package76. The predictability of seasonal trend, related to the concept of seasonality sensu Marshall and Burguess22, reflects the variance in time series due to the predictable seasonal pattern within a year. The seasonal fluctuations in data (“seasonality”), determined as the regularity in the periodicity and variance in the average environmental state over seasons (i.e. a predictable repeating pattern within a year), is associated to the predictability of the mean environmental state22. We computed the “unbounded” seasonality (a/b), which is the ratio between the variance of the seasonal trend (“a”) and the variance of the time series once the linear trends are removed (residuals, “b”)76. The seasonal trend (a) is the monthly average of SST across the time series, interpolated afterwards to reproduce the seasonal time series at the same timescale of the original data (days, in this case). The colour of environmental noise measures the predictability between consecutive points in the time series, ranging from predictable or temporal autocorrelated environments (“brown and red noise”) to those experiencing continuous changes (“white noise”)23. Marine environments are considered as some of the most “reddened” (i.e. low-frequency cycled) systems23. To estimate the colour of noise, a spectral analysis is performed on the detrended time series22. The linear regression of the natural log of spectral density as a function of the natural log of frequency is estimated. The resultant negative slope (β) indicates the colour of noise from white (0 ≤ β ≤ 0.5), red (0.5 < β ≤ 1.5) to brown (1.5 < β ≤ 2)23. Brown noise corresponds to a time series dominated by low-frequency cycles, having then a higher predictability (correlation).
Additionally, we estimated the variation of SST for each period. We used the coefficient of variation (CV) of SST for each period (CV = standard deviation × mean−1 × 100) as a measure of the variability or dispersion of SST in relation to the mean. A greater CV would indicate a higher variability of SST. This is the most common index for assessing variability, though it has some limitations. For instance, CV is dependent on the mean value and the length of the time series and is sensitive to very rare, extreme values, typical of skewed distributions. This is why we also used the consecutive disparity index (D), which assesses the consecutive variations in SST as an indicator of temporal variability. D is considered a more suitable metric of temporal variability for ecological studies in comparison with other indices as it is not sensitive to the mean and length of time series. Furthermore, it considers the chronological order of the time series data and thus its autocorrelation structure24. It has been used, for example, to measure the average rate of change of soil moisture due to varied precipitation treatments77. D produces a logarithmic proportional comparison of consecutive values (i.e. daily SST). We calculated D index for each temporal block and grid cell following Fernández‐Martínez et al.24:
where pi is the series value at time i, and n is the series length. D value ranges from 0 to infinite, and in our case, a higher consecutive disparity (D value) would indicate a maintained pulsed nature of daily SST over time.
Finally, we used the skewness as a measure of asymmetrical distribution of SST series around its mean value. A time series with zero skewness would show a symmetric distribution of the values around the mean. In our case, a positively skewed SST time series would show a predominance of small SST values respect to the mean (right skewed; a right tail in the density distribution), while negative skewed time series would indicate a dominance of large SST values. The skewness (to test if SST was positively or negatively skewed) was calculated using the TSA package78 in R.
Thermal thresholds
For each time interval, we calculated the number of days per year exceeding the species thermal limit. Thermal thresholds were retrieved from the literature and corresponded to physiological experiments. If lethal maximum temperature differed between studies, we chose the most conservative value (i.e. the lowest temperature). Thus, we used 25 °C as the threshold for F. serratus62, 18 °C for H. elongata79, 23 °C for L. ochroleuca80, 24 °C for S. polyschides81,82, and 20 °C for L. hyperborea62,80 (Fig. S5 Supporting information). The thermal threshold of F. vesiculosus (28 °C62) was above the maximum SST registered in the time series (25.3 °C) and could not be integrated in the analyses.
Statistical analyses
We analyzed the persistence and extirpations of the macroalgae species in each site across time (baseline and resurvey periods). To do that, observations on the populations were categorized as “one (1)” if the species persisted over the two periods of time (or was observed only during the second period; just a few cases), and “zero (0)” if the species was present in the first period but absent in the second time period (we also regarded it as zero for F. vesiculosus if it has virtually vanished from semi-exposed shores and was only found in very small populations in wave sheltered locations).
To reflect the change in climate conditions over time, we calculated the difference (anomaly) between the resurvey period (1991–2015) and the baseline period (1982–1990) for all independent variables related to average trends, extreme events and temporal variability of SST (Table S1; Supporting information). In the case of the consecutive disparity index (D), we estimated the percentage of change between periods. Thus, a decrease in D (i.e. a decrease in the disparity of SST) from one period to the next is reflected as a negative percentage change.
The complete set of anomalies (85 variables) were initially assessed using a Pearson correlation test to discard high positive correlations in all cells of the study area (r ≥ 0.70, p < 0.001; i.e. variables showing a very similar longitudinal pattern of change). Since many variables derived from extreme events (MHWs, MCSs) were correlated to each other, in these cases we performed an additional correlation analysis to discard the least significant variables according to our species distributions. We first excluded the extreme events derived anomalies with the highest variance inflation factor (VIF). Then, we tested their correlations only in the cells including species data to exclude the one of each correlated pair with the largest AIC (Akaike’s Information Criterion) in a bivariate model (“binomial” family) performed for each species. This analysis was performed using the “corSelect” function of the “fuzzySim” R package83. From 18 to 29 extreme events related variables were retained for each species for subsequent analyses.
We used hierarchical partitioning analysis to determine how the change of each variable relates to the distributional shifts of macroalgae between periods. This analysis determines the relative importance of each independent variable to each species84. Hierarchical partitioning uses all regression models in a hierarchy to identify the explanatory variables more independently correlated to the dependent variables, thus producing better deductions on the most likely causal factors than regressions85. We estimated the hierarchical partitioning algorithm sensu Chevan and Sutherland84 using the “hier.part” R package86. The independent contribution (I) of each variable to the pattern of persistence/local extinction for each species was estimated using a randomized routine of 100 repetitions specifying the binomial family in the function “rand.hp” and assessing the significance of the analysis using the upper 0.95 confidence limit. Because the maximum number of variables that can be analyzed in each partition is 1286, we used hierarchical partitioning analysis in an iterative process for each species. Since the variables seasonally characterizing extreme events were very numerous, we first used hierarchical partitioning to select the most relevant variables (with the highest I, up to 12) per species and type of extreme event (MHW, MCS). Afterwards, the statistically significant variables with I ≥ 0.75 quantile related to each type of seasonal extreme events were selected to be analyzed together with the rest of the variables in the final hierarchical partitioning analysis per species. Hierarchical partitioning results were represented generating a heat map and calculating a hierarchical clustering displayed as a dendrogram using the function heatmap.2 of the R package “gplots”87.
We further explored the overall relationship between persistence/extirpation and the variables selected by the previous hierarchical partitioning analyses, using a non-metric multidimensional scaling (NMDS) ordination. We computed a two-dimensional NMDS using 100 random iterations and Euclidean distances to calculate the dissimilarity index among sites (i.e. locations, grids) with persistent/extirpated populations of each species. We calculated the dissimilarity among sites with observed persistence and/or extirpation of seaweed species based on the change in SST metrics between time periods. We fit the vectors of the environmental variables to the ordination setting 999 permutations. A generalized additive model was also used to assess how well the variables fit on a smooth surface within the NMDS. We performed the analyses with the functions metaMDS, ordisurf and envfit of the R package “vegan”88.
Data availability
The data that support the findings of this study are openly available in Zenodo at https://doi.org/10.5281/zenodo.8414685, reference number md5:a07a87983a73f1ccdef85417ec652b8a.
References
Cheng, L. et al. Past and future ocean warming. Nat. Rev. Earth Environ. 3, 776–794 (2022).
Poloczanska, E. S. et al. Global imprint of climate change on marine life. Nat. Clim. Change 3, 919–925 (2013).
Wernberg, T. et al. Climate-driven regime shift of a temperate marine ecosystem. Science 353, 169–172 (2016).
Burrows, M. T. et al. Ocean community warming responses explained by thermal affinities and temperature gradients. Nat. Clim. Change 9, 959–963 (2019).
Burrows, M. T. et al. The pace of shifting climate in marine and terrestrial ecosystems. Science 1979(334), 652–655 (2011).
Pecl, G. T. et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 355, 9214 (2017).
Donelson, J. M. et al. Understanding interactions between plasticity, adaptation and range shifts in response to marine environmental change. Philos. Trans. R. Soc. B 374, 20180186 (2019).
Oliver, E. C. J. et al. Longer and more frequent marine heatwaves over the past century. Nat. Commun. 9, 1324 (2018).
Frölicher, T. L., Fischer, E. M. & Gruber, N. Marine heatwaves under global warming. Nature 560, 360–364 (2018).
Collins, M. et al. Extremes, Abrupt Changes and Managing Risk (2019).
Wernberg, T. et al. An extreme climatic event alters marine ecosystem structure in a global biodiversity hotspot. Nat. Clim. Change 3, 78–82 (2013).
Garrabou, J. et al. Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob. Change Biol. 28, 5708–5725 (2022).
Smith, K. E. et al. Biological impacts of marine heatwaves. Annu. Rev. Mar. Sci. 15, 119–145 (2023).
Smale, D. A. et al. Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat. Clim. Change 9, 306 (2019).
Hobday, A. J. et al. A hierarchical approach to defining marine heatwaves. Prog. Oceanogr. 141, 227–238 (2016).
Mieszkowska, N., Burrows, M. T., Hawkins, S. J. & Sugden, H. Impacts of pervasive climate change and extreme events on rocky intertidal communities: Evidence from long-term data. Front. Mar. Sci. 8, 1 (2021).
Benedetti-Cecchi, L., Bertocci, I., Vaselli, S. & Maggi, E. Temporal variance reverses the impact of high mean intensity of stress in climate change experiments. Ecology 87, 2489–2499 (2006).
Vaselli, S., Bertocci, I., Maggi, E. & Benedetti-Cecchi, L. Effects of mean intensity and temporal variance of sediment scouring events on assemblages of rocky shores. Mar. Ecol. Prog. Ser. 364, 57–66 (2008).
Kroeker, K. J. et al. Ecological change in dynamic environments: Accounting for temporal environmental variability in studies of ocean change biology. Glob. Change Biol. 26, 54–67 (2020).
Gerhard, M. et al. Environmental variability in aquatic ecosystems: Avenues for future multifactorial experiments. Limnol. Oceanogr. Lett. 8, 247–266 (2023).
Colwell, R. K. Predictability, constancy, and contingency of periodic phenomena. Ecology 55, 1148–1153 (1974).
Marshall, D. J. & Burgess, S. C. Deconstructing environmental predictability: Seasonality, environmental colour and the biogeography of marine life histories. Ecol. Lett. 18, 174–181 (2015).
Vasseur, D. A. & Yodzis, P. The color of environmental noise. Ecology 85, 1146–1152 (2004).
Fernández-Martínez, M. et al. The consecutive disparity index, D: A measure of temporal variability in ecological studies. Ecosphere 9, e02527 (2018).
Guttal, V. & Jayaprakash, C. Changing skewness: An early warning signal of regime shifts in ecosystems. Ecol. Lett. 11, 450–460 (2008).
Pansch, C. et al. Heat waves and their significance for a temperate benthic community: A near-natural experimental approach. Glob. Change Biol. 24, 4357–4367 (2018).
Harvey, B. P., Marshall, K. E., Harley, C. D. G. & Russell, B. D. Predicting responses to marine heatwaves using functional traits. Trends Ecol. Evol. 37, 20 (2021).
Jackson, M. C., Pawar, S. & Woodward, G. The temporal dynamics of multiple stressor effects: From individuals to ecosystems. Trends Ecol. Evol. 36, 402–410 (2021).
Bennett, S. et al. Thermal performance of seaweeds and seagrasses across a regional climate gradient. Front. Mar. Sci. 9, 1–11 (2022).
Aitken, S. N., Yeaman, S., Holliday, J. A., Wang, T. & Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 1, 95–111 (2008).
Schiel, D. R. & Foster, M. S. The population biology of large brown seaweeds: Ecological consequences of multiphase life histories in dynamic coastal environments. Annu. Rev. Ecol. Evol. Syst. 37, 343–372 (2006).
Nicastro, K. R. et al. Shift happens: Trailing edge contraction associated with recent warming trends threatens a distinct genetic lineage in the marine macroalga Fucus vesiculosus. BMC Biol. 11, 6 (2013).
Casado-Amezúa, P. et al. Distributional shifts of canopy-forming seaweeds from the Atlantic coast of Southern Europe. Biodivers. Conserv. 28, 1151–1172 (2019).
Duarte, L. et al. Recent and historical range shifts of two canopy-forming seaweeds in North Spain and the link with trends in sea surface temperature. Acta Oecol. 51, 1–10 (2013).
Straub, S. C. et al. Persistence of seaweed forests in the anthropocene will depend on warming and marine heatwave profiles. J. Phycol. 58, 22–35 (2022).
Schlegel, R. W., Darmaraki, S., Benthuysen, J. A., Filbee-Dexter, K. & Oliver, E. C. J. Marine cold-spells. Prog. Oceanogr. 198, 102684 (2021).
Fernández, C. & Anadón, R. . La. cornisa cantabrica: Un escenario de cambios de distribución de comunidades intermareales. Algas 39, 30–32 (2008).
Müller, R., Laepple, T., Bartsch, I. & Wiencke, C. Impact of Oceanic Warming on the Distribution of Seaweeds in Polar and Cold-Temperate Waters (2009).
Voerman, S. E., Llera, E. & Rico, J. M. Climate driven changes in subtidal kelp forest communities in NW Spain. Mar. Environ. Res. 90, 119–127 (2013).
Piñeiro-Corbeira, C., Barreiro, R. & Cremades, J. Decadal changes in the distribution of common intertidal seaweeds in Galicia (NW Iberia). Mar. Environ. Res. 113, 106–115 (2016).
Muguerza, N. et al. A spatially-modelled snapshot of future marine macroalgal assemblages in southern Europe: Towards a broader Mediterranean region? Mar. Environ. Res. 176, 105592 (2022).
Fernández, C. The retreat of large brown seaweeds on the north coast of Spain: The case of Saccorhiza polyschides. Eur. J. Phycol. 46, 352–360 (2011).
Lüning, K. & Dieck, I. tom. The distribution and evolution of the Laminariales: North Pacific—Atlantic relationships. In Evolutionary Biogeography of the Marine Algae of the North Atlantic (eds Garbary, D. J. & South, G. R.) 187–204 (Springer, 1990).
van den Hoek, C. The distribution of benthic marine algae in relation to the temperature regulation of their life histories. Biol. J. Linnean Soc. 18, 81–144 (1982).
Breeman, A. M. Relative importance of temperature and other factors in determining geographic boundaries of seaweeds: Experimental and phenological evidence. Helgoländer Meeresuntersuchungen 42, 199–241 (1988).
Barrientos, S., Barreiro, R., Cremades, J. & Piñeiro-Corbeira, C. Setting the basis for a long-term monitoring network of intertidal seaweed assemblages in northwest Spain. Mar. Environ. Res. 160, 105039 (2020).
Viejo, R. M., Des, M. & Gutiérrez, D. Drivers of the fine-scale distribution of a canopy-forming seaweed at the southern edge of its range. Mar. Ecol. Prog. Ser. 727, 91–109 (2024).
Viejo, R. M., Martínez, B., Arrontes, J., Astudillo, C. & Hernández, L. Reproductive patterns in central and marginal populations of a large brown seaweed: Drastic changes at the southern range limit. Ecography 34, 75–84 (2011).
Duarte, L. & Viejo, R. M. Environmental and phenotypic heterogeneity of populations at the trailing range-edge of the habitat-forming macroalga Fucus serratus. Mar. Environ. Res. 136, 16–26 (2018).
Zardi, G. I. et al. Closer to the rear edge: Ecology and genetic diversity down the core-edge gradient of a marine macroalga. Ecosphere 6, 1–25 (2015).
de la Fernández González, M. C. Estudios estructurales y dinámica del fitobentos intermareal (facies rocosa) de la región de Cabo Peñas, con especial atención a la biología de Saccorhiza polyschides (Le Jol.) Batt. (1980).
Hereward, H. F. R., King, N. G. & Smale, D. A. Intra-annual variability in responses of a canopy forming kelp to cumulative low tide heat stress: Implications for populations at the trailing range edge. J. Phycol. 56, 146–158 (2020).
Fraga, F. Upwelling off the Galician coast, northwest Spain. Coast. Upwell. 1, 176–182 (1981).
Letelier, J., Pizarro, O. & Nuñez, S. Seasonal variability of coastal upwelling and the upwelling front off central Chile. J. Geophys. Res. Oceans 114, 1 (2009).
Salois, S. L. et al. Coastal upwelling generates cryptic temperature refugia. Sci. Rep. 12, 19313 (2022).
Pérez, F. F. et al. Plankton response to weakening of the Iberian coastal upwelling. Glob. Change Biol. 16, 1258–1267 (2010).
Sydeman, W. J. et al. Climate change and wind intensification in coastal upwelling ecosystems. Science 345, 77–80 (2014).
Otero, P., Cabrero, Á., Alonso-Pérez, F., Gago, J. & Nogueira, E. Temperature and salinity trends in the northern limit of the Canary current upwelling system. Sci. Total Environ. 901, 165791 (2023).
Sousa, M. C. et al. NW Iberian Peninsula coastal upwelling future weakening: Competition between wind intensification and surface heating. Sci. Total Environ. 703, 134808 (2020).
Rees, T. K. A note on the longevity of certain species of the Fucaceae. Ann. Bot. 46, 1062–1064 (1932).
Malm, T., Kautsky, L. & Engkvist, R. Reproduction, Recruitment and Geographical Distribution of Fucus serratus L. in the Baltic Sea (2001).
Lüning, K. Temperature tolerance and biogeography of seaweeds: The marine algal flora of Helgoland (North Sea) as an example. Helgoländer Meeresuntersuchungen 38, 305–317 (1984).
McCook, L. J. & Chapman, A. R. O. Community succession following massive ice-scour on an exposed rocky shore: Effects of Fucus canopy algae and of mussels during late succession. J. Exp. Mar. Biol. Ecol. 154, 137–169 (1991).
Biskup, S., Bertocci, I., Arenas, F. & Tuya, F. Functional responses of juvenile kelps, Laminaria ochroleuca and Saccorhiza polyschides, to increasing temperatures. Aquat. Bot. 113, 117–122 (2014).
Pereira, T. R. et al. Temperature effects on the microscopic haploid stage development of Laminaria ochroleuca and Sacchoriza polyschides, kelps with contrasting life histories. Cahiers Biol. Mar. 52, 395 (2011).
Cavanaugh, K. C., Reed, D. C., Bell, T. W., Castorani, M. C. N. & Beas-Luna, R. Spatial variability in the resistance and resilience of giant kelp in southern and Baja California to a multiyear heatwave. Front. Mar. Sci. 6, 413 (2019).
Schlegel, R. W., Oliver, E. C. J., Wernberg, T. & Smit, A. J. Nearshore and offshore co-occurrence of marine heatwaves and cold-spells. Prog. Oceanogr. 151, 189–205 (2017).
Izquierdo, P., Rico, J. M., Taboada, F. G., González-Gil, R. & Arrontes, J. Characterization of marine heatwaves in the Cantabrian Sea, SW Bay of Biscay. Estuar. Coast Shelf Sci. 274, 107923 (2022).
Fernández, C. Current status and multidecadal biogeographical changes in rocky intertidal algal assemblages: The northern Spanish coast. Estuar. Coast Shelf Sci. 171, 35–40 (2016).
Martins, G. M. et al. Direct and indirect effects of climate change squeeze the local distribution of a habitat-forming seaweed. Mar. Ecol. Prog. Ser. 626, 43–52 (2019).
Chabrerie, A. & Arenas, F. What if the upwelling weakens? Effects of rising temperature and nutrient depletion on coastal assemblages. Oecologia 1–17 (2024).
Moss, B. Apical meristems and growth control in Himanthalia elongata (SF Gray). N. Phytol. 68, 387–397 (1969).
Martínez, B., Viejo, R. M., Carreño, F. & Aranda, S. C. Habitat distribution models for intertidal seaweeds: Responses to climatic and non-climatic drivers. J. Biogeogr. 39, 1877–1890 (2012).
Huang, B. et al. Improvements of the daily optimum interpolation sea surface temperature (DOISST) version 2.1. J. Clim. 34, 2923–2939 (2021).
Schlegel, R. W. & Smit, A. J. heatwaveR: A central algorithm for the detection of heatwaves and cold-spells. J. Open Source Softw. 3, 821 (2018).
Barneche, D. & Burgess, S. envPred: Calculate Environmental Predictability for Even and Uneven Time Series Data. Preprint at https://github.com/dbarneche/envPred (2023).
Griffin-Nolan, R. J., Slette, I. J. & Knapp, A. K. Deconstructing precipitation variability: Rainfall event size and timing uniquely alter ecosystem dynamics. J. Ecol. 109, 3356–3369 (2021).
Chan, K.-S. & Ripley, B. TSA: Time Series Analysis. R Package Version 1.3 (2020).
Martínez, B., Arenas, F., Trilla, A., Viejo, R. M. & Carreño, F. Combining physiological threshold knowledge to species distribution models is key to improving forecasts of the future niche for macroalgae. Glob. Change Biol. 21, 1422–1433 (2015).
Tom Dieck, I. North Pacific and North Atlantic digitate Laminaria species (Phaeophyta): Hybridization experiments and temperature responses. Phycologia 31, 147–163 (1992).
Tom Dieck, I. Temperature tolerance and survival in darkness of kelp gametophytes (Laminariales, Phaeophyta): Ecological and biogeographical implications. Mar. Ecol. Prog. Ser. 100, 253 (1993).
Lüning, K. Seaweeds: Their Environment, Biogeography, and Ecophysiology (Wiley, 1991).
Barbosa, A. M. fuzzySim: Applying fuzzy logic to binary similarity indices in ecology. Methods Ecol. Evol. 6, 853–858 (2015).
Chevan, A. & Sutherland, M. Hierarchical partitioning. Am. Stat. 45, 90–96 (1991).
Mac Nally, R. Hierarchical partitioning as an interpretative tool in multivariate inference. Austral. Ecol. 21, 224–228 (1996).
Mac Nally, R. & Walsh, C. J. Hierarchical partitioning public-domain software. Biodivers. Conserv. 13, 659 (2004).
Warnes, G. R. et al. gplots: Various R Programming Tools for Plotting Data. Preprint at https://CRAN.R-project.org/package=gplots (2022).
Oksanen, J. et al. vegan: Community Ecology Package. Preprint at https://CRAN.R-project.org/package=vegan (2022).
Acknowledgements
This research has received funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie Grant Agreement No 894941 to RMC (MARHOT project).
Author information
Authors and Affiliations
Contributions
RMC: Conceptualization-Lead, Data curation-Equal, Formal analysis-Lead, Funding acquisition-Lead, Investigation-Lead, Methodology-Lead, Project administration-Lead, Visualization-Lead, Writing—original draft-Lead, Writing—review & editing-Lead. BDCM: Conceptualization-Equal, Data curation-Equal, Funding acquisition-Supporting, Writing—review & editing-Supporting. RMV: Conceptualization-Equal, Data curation-Equal, Formal analysis-Supporting, Investigation-Equal, Methodology-Supporting, Writing—original draft-Equal, Writing—review & editing-Equal.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chefaoui, R.M., Martínez, B.DC. & Viejo, R.M. Temporal variability of sea surface temperature affects marine macrophytes range retractions as well as gradual warming. Sci Rep 14, 14206 (2024). https://doi.org/10.1038/s41598-024-64745-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-64745-7
- Springer Nature Limited