
Abstract
High stability of fluctuation in physiological patterns across fixed time periods suggest healthy fractal complexity, while greater randomness in fluctuation patterns may indicate underlying disease processes. The importance of fractal stability in mid-life remains unexplored. We quantified fractal regulation patterns in 24-h accelerometer data and examined associations with cognitive function in midlife. Data from 5097 individuals (aged 46) from the 1970 British Cohort Study were analyzed. Participants wore thigh-mounted accelerometers for seven days and completed cognitive tests (verbal fluency, memory, processing speed; derived composite z-score). Detrended fluctuation analysis (DFA) was used to examine temporal correlations of acceleration magnitude across 25 time scales (range: 1 min–10 h). Linear regression examined associations between DFA scaling exponents (DFAe) and each standardised cognitive outcome. DFAe was normally distributed (mean ± SD: 0.90 ± 0.06; range: 0.72–1.25). In males, a 0.10 increase in DFAe was associated with a 0.30 (95% Confidence Interval: 0.14, 0.47) increase in composite cognitive z-score in unadjusted models; associations were strongest for verbal fluency (0.10 [0.04, 0.16]). Associations remained in fully-adjusted models for verbal fluency only (0.06 [0.00, 0.12]). There was no association between DFA and cognition in females. Greater fractal stability in men was associated with better cognitive function. This could indicate mechanisms through which fractal complexity may scale up to and contribute to cognitive clinical endpoints.
Similar content being viewed by others

Introduction
Self-affinity, or scale invariance, is a phenomenon where fluctuations across a short period of time resemble fluctuations observed over a longer-period of time1. In healthy individuals, self-affinity reflects the successful coordination of the neural and motor systems2. In unhealthy states, self-affinity declines and increased randomness is observed2. Self-affinity has been observed across various physiological outputs including heart rate3,4,5, respiration rate6,7, speech8, gait9,10,11 and neural activity12,13,14,15. Signal data from accelerometers can also provide insight into the complexity of activity fluctuations across the wear period16, beyond traditional measures of physical activity volume, intensity or duration17. Although acceleration signals from physical activity monitors were initially assumed to solely be due to random noise, exploration of fractal patterns in accelerometer-based data indicates that these fluctuations provide meaningful information into the intrinsic and complex fractal patterns of the individual.
Detrended fluctuation analysis (DFA) captures similarities in fluctuation patterns across time by deriving a scaling exponent to capture consistency of these fluctuations across different temporal scales (e.g. sequential ‘activity windows’ of 1 min, 30 min, 5 h, etc.). Scaling exponents that approach 1 indicate healthy fractal complexity18,19, whereas smaller scaling components highlight irregularity in the physiological integration of neural and muscular systems (e.g. 0.5 indicates no correlation). Fractal stability is generally lower in females and declines with increasing age20. Lower DFA scaling exponents are associated with higher risk of mortality, frailty and various neurodegenerative outcomes including dementia20,21,22,23,24. Early assessment of fractal stability at the individual level may capture physiological dysfunction before it scales up to be clinically visible, and therefore DFA exponents may be a useful biomarker to indicate underlying health dysregulation.
Direct evidence points to poor fractal complexity as an antecedent to cognitive decline20,21,22,23,24. Existing evidence suggests strong associations between the disturbance in the fractal complexity of individual motor movements and incidence and progression of dementia21,23,24,25,26,27,28. This is largely thought to be due to underlying neuromotor pathways28, where subtle fractal disruptions may be indicative of underlying cognitive impairment. Promisingly, fractal instability, even as an indicator of underlying disease processes, appears to be reversable, with engagement in increased physical activity demonstrating a positive impact in regaining healthy fractal complexity29,30,31. Yet, investigation of DFA scaling exponents in relation to health outcomes has focused solely on older adults, with no investigation in individuals younger than age 50, despite meaningful variation in DFA across the life course20. Therefore, our aim was to quantify fractal regulation patterns in 24-h accelerometer data in a midlife cohort at age 46 and examine associations with cognitive function.
Results
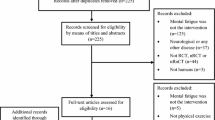
The birth cohort sample is described in Table 1, and the derivation of the study sample is provided in Fig. 1. Briefly, participants were aged 46–48 years during assessment, 53.5% (n = 2729) were female, and most of the sample were non-smokers (never smoker: 49.1%, ex-smoker: 32.9%) and either abstinent (10%) or non-problematic drinkers (67.5%). Nearly half of the sample (46%) was educated to A level (typically attained at age 18), with 26% reporting no formal qualifications and 28% having a degree or higher; similarly, nearly half the sample (46.7%) had a lower managerial/ intermediate occupation. Two thirds (65.8%) were married or had a civil partner. Mean body mass index (BMI) was 28.2 ± 5.5 kg/m2 and 5% of the sample were classified being severely hampered according to the European Union- Statistics on Income and Living Conditions (EU-SILC) disability definition. Males were more likely to be problem drinkers, have higher BMI, engage in more daily activity and have a professional/managerial occupation. Females were more likely to perform better on the cognitive tests, have formal qualifications, to be married or be widowed/separated, to have a disability and have longer sleep (see Supplementary Table 1). DFA exponents were normally distributed (Fig. 2), with a mean of 0.900 ± 0.646, median of 0.892 (Q1:0.855, Q3: 0.936) and ranged from 0.720 to 1.251.

Flowchart indicating the derivation of study sample.

Distribution of detrended fluctuation analysis (DFA) scaling exponent.
DFA exponents were slightly higher in males compared to females (0.903 ± 0.067 vs. 0.897 ± 0.062; see Table 1), in non-smokers compared to current smokers, in those with higher levels of educational attainment and those in higher occupational classes. Those who were divorced, widowed or separated had lower DFA exponents, compared to those who were currently or had never married (p = 0.10). There was a weak negative correlation between BMI and DFA exponents (r = − 0.07) and weak positive correlation of both sleep (r = 0.04) and total activity with DFA exponents (r = 0.10). There was no difference in DFA exponent amongst disability or alcohol consumption status. Supplementary Table 1 describes covariates and cognitive outcomes by sex.
Given clear interactions between sex and DFA (Fig. 3) with composite cognitive z-score, all models were stratified by sex. In females, there was no association between DFA and any cognitive outcome (Table 2, models i–iii). In males, higher DFA was consistently associated with higher cognitive performance. In the unadjusted model, a 0.1 increase in DFA exponent was associated with a 0.30 (95% Confidence Interval: 0.14, 0.47) increase in composite cognitive z-score. The association was attenuated by approximately two thirds after adjustment for education, self-reported health, disability, BMI, smoking, alcohol, sleep, occupational class and total daily PA time (model iii: 0.11 [− 0.05, 0.26]). When examining individual domains, associations were strongest for verbal fluency (Fig. 4). For example, a 0.1 increase in DFA exponent was associated with a 0.10 (0.04, 0.16) and 0.06 (0.00, 0.12) increase in verbal fluency z-scores in unadjusted and fully-adjusted models, respectfully (Table 2; models i and iii). This is equivalent to a 0.65 (0.27, 1.02) and 0.35 (− 0.02, 0.72) increase in the number of animals named (Supplementary Table 2). Although there was some evidence of an association between larger DFA exponent and each of processing speed (0.07 [0.01, 0.13]), immediate recall (0.08 [0.02, 0.14]) and delayed recall (0.05 [− 0.01, 0.11]), associations were fully attenuated in fully-adjusted models.

Associations between DFA coefficient and composite cognitive z-score, stratified by females (n = 2729) and males (n = 236).

Associations between DFA coefficient and (a) verbal fluency; (b) immediate recall; (c) delayed recall; (d) processing speed, stratified by females (n = 2729) and males (n = 2368).
Cognitive scores were higher in those with valid DFA data (mean composite cognitive z-score: 0.23 ± 2.66) compared to those who participated in the age 46 wave but did not wear an accelerometer (− 0.30 ± 2.74; up to n = 3484). This was true for verbal fluency (animals named: 24.0 ± 6.1 vs. 23.0 ± 6.3), immediate recall (6.7 ± 1.4 vs. 6.4 ± 1.5) and delayed recall (5.6 ± 1.8 vs. 5.3 ± 1.8) but not processing speed (letters searched: 346 ± 82 vs. 347 ± 89). Conversely, the analytical sample had lower DFA exponents (n = 5097, 0.900 ± 0.06) than those missing cognitive data on one or more test (n = 142 with DFA data, 0.912 ± 0.06).
Discussion
In a large population representative sample of adults in midlife, healthy fractal complexity (i.e. higher DFA scaling exponents indicating stability of fluctuation patterns across different time periods) was associated with better cognitive performance in males. Associations were strongest for executive function measures, with DFA exponent remaining positively associated with verbal fluency independent of socioeconomic measures, health behaviours, health status indicator and total PA levels. There were no associations in females across any cognitive measure. These novel findings indicate that meaningful decline in fractal complexity may begin to emerge well before clinical presentation, possibly decades earlier. Assessment of fractal complexity in midlife may be a clinically promising method to capture meaningful deterioration in neural motor coordination before it becomes clinically visible.
Despite increased recognition of the importance of midlife factors for dementia risk32,33, no other study has examined associations between fractal complexity and health outcomes prior to age 50, therefore comparison to existing evidence is limited. However, previous investigation in older adults has demonstrated robust associations between disrupted fractal complexity in motor activity and both the onset and progression of dementia21,23,24,25,26,27,28. In a US cohort study of > 1000 adults with no baseline dementia diagnosis (mean age: 79.7 ± 0.2), lower DFA exponents were strongly associated with increased risk of Alzheimer’s disease diagnosis and a faster decline in global cognitive function25. Although five distinct cognitive domains were assessed across 19 tests, authors did not explore how associations differed by cognition type. Further exploration in this cohort indicated that fractal degradation progressively worsened over time, even at pre-clinical stages of Alzheimer’s disease, with an accelerated decline after diagnosis21. Other studies have reported similar robust patterns of associations with clinical cognitive end-points23,24,26,27,28. Notably, one study reported opposing sex patterns to those observed above, suggesting that disrupted fractal complexity was associated with amyloid plaque pathology in females but not males24. Differing results in the present study may be due to the sample being older (mean age: 65.9 ± 8.3), having insufficient power (n = 61 males) or differences in ascertainment of neuropathological versus cognitive outcomes. The effect sizes presented in this study are smaller than those reported in studies on dementia or mortality outcomes20,23,24,26,27,28, which is not unexpected to the young age of the participant. Nonetheless, the robustness of associations, even after adjustment for key confounders, suggest that this physiological relationship emerges at a younger age than previously expected.
A central hypothesis explaining the link between reduced fractal activity regulation and cognitive impairment focuses on the neuroanatomical changes in the suprachiasmatic nucleus 34,35,36. Degeneration in the suprachiasmatic nucleus, the area of the brain responsible for behavioral and physiological circadian rhythms34, may explain why disruptions in fractal activity arise before cognitive impairments become clinically visible28. The suprachiasmatic nucleus is located in the anterior part of the hypothalamus, which plays an integral role in coordinating and maintaining homeostasis across many of the body’s systems37. Therefore, irregular fluctuations in motor activity might indicate underlying degeneration in this central area. This hypothesis is consistent with patterns from other pre-clinical measures of health. For example, a frailty index constructed of abnormal blood biomarkers captures damage at a subclinical level38,39 and can predict adverse health outcomes including mortality even in young, otherwise healthy adults40.
Crucially, the presence of this association at age 46 indicates that self-affinity may begin to break down earlier than previously recognized, which is consistent with a life course approach of accumulation of damage at the cellular, organ and system level41. This steady decline in DFA scaling has been previously observed in a US cohort of participants from age 6 to 8520. The lack of association of DFA scaling exponents with cognitive function in women is surprising, although there was some indication of an association in unadjusted verbal fluency models. We hypothesize that women may be able to withstand a greater degree of poor fractal complexity in midlife, before it impacts clinical outcomes. When considering the emergence of the male–female health-survival paradox in later life42, this has important implications. Women often exhibit poorer health than age-matched male counterparts, with greater levels of frailty43, healthcare use, disability44, cognitive impairment45,46, yet often have lower mortality risk and longer life expectation43,47. This may be due to a better intrinsic capacity to handle ill health and damage at the cellular and system level, which could explain why women had poorer DFA exponents than men, yet better scores on cognitive tests. Conversely, other hypotheses have suggested that sex difference in lifestyle behaviours, social roles or health-care seeking behaviours may explain why women appear to have poorer health than men42—for example, higher disability observed in this study-, yet better survival outcomes.
Associations were strongest for verbal fluency and weakest for both immediate and delayed recall. This was surprising, given the hypothalamus plays a central role in learning and memory37—and thus the memory recall tasks-, and reduced fractal activity regulation is hypothesized to result from impairments in the anterior hypothalamus34,35,36. Verbal fluency – measured as the number of animals participants could name within 1 min—may have been a more sensitive and demanding measure of fluid cognition, compared to the cruder 10-word immediate and delayed recall tasks. For example, the animal naming task relies upon several aspects of executive function including clustering and self-initiation (e.g. generating a series of animal names that make up a specific cluster such as bird type, fish type), task switching (e.g. switching to the next cluster after all names in a single species has been exhausted) and action inhibition (e.g. suppressing the tendency to repeat previous names or names that do not fit within the instructions)48,49. Therefore, it may more sensitively capture neuroanatomical changes in the hypothalamus.
Key strengths of this study include the large population representative and age-homogenous sample, ascertainment of motor activity using a continuous 7-day wear period with a thigh worn device, and assessment of several cognitive domains with tests that are commonly used in large epidemiological studies50,51. Limitations include the cross-sectional nature of the data, loss to follow-up of participants across the life course, and lower cognitive scores in those who did not wear an accelerometer. Due to the cross-sectional design, we are unable to rule out reverse causality, as it is possible that cognitive impairment may cause poor fractal stability given some of the neural mechanisms described above34,35,36,37. Additionally, there was no assessment of neuropathological outcomes, which could have provided further insight into mechanisms through which fractal complexity may influence cognitive function before impairment manifest clinically.
Our findings need replication in longitudinal studies examining associations throughout midlife to better understand the age (and threshold) at which fractal complexity can meaningfully identify those in need of early intervention. It is also important to better understand the neuropathological mechanisms through which the loss of self-affinity impacts cognitive performance, particularly to understand how it scales up to Alzheimer and dementia outcomes. The significance of this lies in the potential connection between increased exercise and improvements in fractal complexity29,30,31. Should longitudinal evidence reveal a meaningful relationship between fractal complexity and cognitive clinical end-points, increased physical activity may emerge as a promising avenue to influence cognitive health, with fractal complexity as a potential mediator. Another important area of exploration is to investigate how these associations may differ across key characteristics given DFA gradients across most covariates; the role of occupational physical activity may play an intriguing role given known negative effects of occupational physical activity on various health outcomes52. Finally, studies that can combine device-assessed physical activity measurements and neuroimaging are needed to examine the pathological characteristics of those who demonstrate poor fractal complexity in midlife. This could provide further insight into the common discordance between underlying neuropathology or genetic risk and clinical expression of cognitive impairment53,54.
In summary, we identified robust associations between poor fractal complexity and worse cognitive function in males in midlife. Considering the robust associations of fractal complexity with both the onset and progression of clinical cognitive outcomes (e.g. Alzheimer’s disease)21,25, early identification has enormous potential to identify those at early risk. In addition to increasingly accurate, accessible and detailed assessments of physical activity55, consumer wearables have seen the implementation of increasingly complex biotechnologies including photoplethysmography and single-lead electrocardiography56. Incorporation of fractal physiology into these wearables could one day be feasible by embedding the algorithms used in this study. This holds enormous promise for the future of personalized medicine and early intervention.
Materials and methods
Sample
The 1970 British Cohort Study is a birth cohort of over 17,000 individuals born in England, Scotland or Wales during a single week in April 197057; there were no further exclusion criteria. Participants have been followed up to ten times across life, including the most recent data collection at age 46–48 (over an 18-month period, hereafter referred to age 46 wave). At age 46, study members participated in a home visit which consisted of a self-completion questionnaire, a computer-assisted personal interview and a biomedical assessment including a battery of cognitive tests. Participants were subsequently invited to wear a thigh-mounted accelerometer (activPAL3 micro; PAL Technologies Ltd., Glasgow, United Kingdom) for a 7-day wear period. All participants provided informed consent and the most recent ethical approval was given by NRES Committee South East Coast- Brighton & Sussex (ref: 15/LO/1446). This research was conducted in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments. Any individuals who lacked capacity to consent were not included. Of the 8581 individuals who participated in the age 46 wave, 5097 had accelerometer and cognitive data and thus were included in the analytical sample (see Fig. 1 for detail on derivation of sample). Therefore, inclusion criteria were: (i) participated in age 46 data collection wave; (ii) provided valid accelerometer data (minimum of 3 weekdays and 1 weekend of wear time; where a valid day was 10 + h); (iii) had data on all four cognitive tests.
Device-measured DFA assessment
Participants were fitted with the activPAL3 micro device (frequency: 20 Hz) on the midline anterior part of the upper thigh during the home visit58. The device was waterproofed with an adhesive and participants were instructed to wear the device continuously for at least seven days, including while sleeping, bathing, swimming or participating in any activity. Participants were also instructed not to re-attach the device if it fell off accidentally or was removed; devices were returned to the research team via post. Partial data from the first day was removed, thus the seven-day collection period commenced at midnight on the day the device was fitted. The raw data acceleration signal from each 1 s window was processed to derive the signal vector magnitude measuring movement acceleration along three planes of movement (x, y, z axes); this yielded up to 60,480 data points per participant (60 s * 60 min * 24 h * 7 days) provided. To be included in analysis, participants must have 3 weekdays and 1 weekend day of wear time, where a valid day was 10 + h.
To derive the DFA scaling components, standard procedures were followed59. First, all non-wear and sleep time were removed from the continuous wear data. Twenty-five distinct time series window sizes were selected (range: 30 s–10 h); note the log of this range was taken to ensure non-linear equally spaced window sizes. Each acceleration signal value was mean centred, by subtracting the average acceleration signal across all valid wear time from each data point. Next, the signal was divided into non-overlapping windows of the chosen size (e.g. consecutive 30 s windows across the whole time series). A least squares line was fitted to the data in each window; for example, in the 10-h time series, a single slope value was produced for each 10 h window representing the trend in that window. The signal was ‘detrended’ by taking the root mean square fluctuation of the residuals from each window. This was repeated across all 25 window sizes. Finally, the log of the root mean square residual values was plotted against the log of the window size; the slope of this line gives the DFA scaling component. A linear relationship on a log–log plots suggests a power law relationship (i.e. fractal scaling)60. Therefore, a DFA scaling exponent of ~ 0.50 indicates no correlation in the fluctuations, while values > 0.5 indicate positive correlations, with increasing fractal complexity as DFA exponent surpasses 1.019,59.
Cognitive outcomes
Four cognitive tests were administered at age 46 to measure (1) immediate short-term memory (word memory); (2) verbal fluency (animal naming); (3) processing speed (letter search) and (4) delayed-recall (delayed word memory). These cognitive assessments have been used in many large-scale epidemiological studies and have been validated for use in population representative samples as sensitive measures of executive function, memory and are established predictors of increased dementia risk61,62,63,64,65,66,67.
First, participants were verbally presented with 10 words at a rate of one word every two seconds. They were then asked to immediately state all the words that they could remember. Immediate recall score was the total number of words they correctly recalled within two minutes. Next, verbal fluency was assessed using a task in which participants named as many animals as they could within one minute. Different breeds (e.g. dog, terrier, poodle) or gender/generation-specific names (e.g. bull, cow, calf) counted as distinct names, while redundancies (e.g. brown cow, black cow) and specific characters (e.g. Bambi) did not. Processing speed was assessed using a visual search speed task. Individuals were given a grid of numbers and were instructed to go line by line and cross our all the ‘P’s and ‘W’s as quickly and as accurately as possible in one minute. The score was the total number of letters searched. Finally, individuals were asked to verbally recall the ten words from the immediate recall task; delayed recall score was the total number of correct words they recalled within two minutes. A combined standardised cognitive score was derived by summing the z-scores of each individual task50.
Covariates
Covariates were a priori chosen based on established associations with fractal complexity and cognition21,22,25,68. Sex was coded as female or male. Current legal marital status was based on self-reported data at age 46 and existing information from previous sweeps and was categorized as ‘never married’, ‘married or civil partner’, and ‘divorced, widowed, separated or former civil partner’. Education was ascertained as the highest completed academic qualification recorded across any data collection sweep and categorized as: ‘none’, ‘up to A levels or diploma (typically attained at age 18)’ or ‘degree or higher. Employment types were coded using the National Statistics Socioeconomic Classifications of Occupations21 as: routine/Semi-routine employment, small employer/lower supervisory, lower managerial/ intermediate, high professional/managerial.
Smoking status was categorized as ‘never smoker’, ‘previous smoker’, ’occasional smoker (not daily)’ and ‘daily smoker’. Drinking status was categorized as ‘non-drinker’, ‘non-problem drinker’ or ‘problem drinker’, derived from five-item the Alcohol Use Disorders Identification Test69. BMI (kg/m2) was derived from study-nurse assessments of height and weight; where missing, self-reported height and weight values were used. Based on a series of questions about longstanding physical or mental health conditions or illnesses lasting or expected to last for 12 months or more, disability was categorized as ‘none’, ‘limited to some extent’ and severely hampered’ using the EU Statistics on Income and Living Conditions classification of disability. Finally, average daily total PA time (any ambulatory movement), sleep and wear time were determined using the validated ActiPASS software, which applies a decision tree model to classify activities based on the signal standard deviation and tilt angle70,71.
Statistical analyses
Descriptive characteristics provided mean ± SD and frequency (proportion) across all covariates and cognitive outcomes. We assessed normality of DFA scores and each cognitive outcome using Kolmogorov–Smirnov tests72. One-way ANOVAs and Pearson correlations were used to assess associations between DFA scores and all covariates. Chi-square tests and one-way ANOVAs were used to assess differences in covariates and cognitive outcomes between females and males. Interactions between sex and DFA coefficient were assessed, whereby analyses were stratified by sex if p < 0.05. We used linear regression to examine associations between DFA coefficients and each cognitive outcome in three models: (i) sex-adjusted (or stratified) models, (ii) model i + education, self-reported health, disability, BMI, smoking, alcohol and sleep-adjusted models and (iii) model ii + additionally adjusted for total PA. Following a missing at random assumption, covariate data were imputed using multiple imputation by chained equations in Stata V.17 (Stata, College Station, TX, USA). Missing data ranged from 0% (smoking, total PA time) to 1.8% (marital status); estimates from 25 imputed datasets were combined using Rubin’s rules73. Characteristics of individuals missing cognitive or DFA data were described and compared to the analytical sample.
Significance statement
This novel study demonstrated that increased fractal complexity is associated with worse cognitive function in midlife. To our knowledge, this is the first study to demonstrate that associations emerge before clinical onset of cognitive impairment and highlights the potential of fractal complexity as an early indicator of those at risk of cognitive decline.
Data availability
The BCS70 datasets are publicly available in the UK Data Archive repository: BCS70 https://discover.ukdataservice.ac.uk/series/?sn=200001.
References
Goldberger, A. L. et al. Fractal dynamics in physiology: Alterations with disease and aging. Proc. Natl. Acad. Sci. USA 99(Suppl 1), 2466–2472 (2002).
Schmitt, D. T. & Ivanov, P. Fractal scale-invariant and nonlinear properties of cardiac dynamics remain stable with advanced age: A new mechanistic picture of cardiac control in healthy elderly. Am. J. Physiol. Regul. Integr. Comp. Physiol. 293, R1923–R1937 (2007).
Sen, J. & McGill, D. Fractal analysis of heart rate variability as a predictor of mortality: A systematic review and meta-analysis. Chaos 28, 072101 (2018).
Chiang, J. Y. et al. Detrended fluctuation analysis of heart rate dynamics is an important prognostic factor in patients with end-stage renal disease receiving peritoneal dialysis. PLoS ONE 11, e0147282 (2016).
Meyer, M. & Stiedl, O. Self-affine fractal variability of human heartbeat interval dynamics in health and disease. Eur. J. Appl. Physiol. 90, 305–316 (2003).
Swapna, M. S., Renjini, A., Raj, V., Sreejyothi, S. & Sankararaman, S. Time series and fractal analyses of wheezing: A novel approach. Phys. Eng. Sci. Med. 43, 1339–1347 (2020).
Tanabe, N., Sato, S., Suki, B. & Hirai, T. Fractal analysis of lung structure in chronic obstructive pulmonary disease. Front. Physiol. 11, 603197 (2020).
Kello, C. T., Anderson, G. G., Holden, J. G. & Van Orden, G. C. The pervasiveness of 1/f scaling in speech reflects the metastable basis of cognition. Cogn. Sci. 32, 1217–1231 (2008).
Dutta, S., Ghosh, D. & Chatterjee, S. Multifractal detrended fluctuation analysis of human gait diseases. Front. Physiol. 4, 274 (2013).
Kamal, S. M. et al. Decoding of the relationship between human brain activity and walking paths. Technol. Health Care 28, 381–390 (2020).
Scafetta, N., Marchi, D. & West, B. J. Understanding the complexity of human gait dynamics. Chaos 19, 026108 (2009).
Paun, M. A., Nichita, M. V., Paun, V. A. & Paun, V. P. Fractal analysis in the quantification of medical imaging associated with multiple sclerosis pathology. Front. Biosci. (Landmark Ed) 27, 66 (2022).
Campbell, O. L. & Weber, A. M. Monofractal analysis of functional magnetic resonance imaging: An introductory review. Hum. Brain Mapp. 43, 2693–2706 (2022).
Favela, L. H., Coey, C. A., Griff, E. R. & Richardson, M. J. Fractal analysis reveals subclasses of neurons and suggests an explanation of their spontaneous activity. Neurosci. Lett. 626, 54–58 (2016).
Burr, R. L., Kirkness, C. J. & Mitchell, P. H. Detrended fluctuation analysis of intracranial pressure predicts outcome following traumatic brain injury. IEEE Trans. Biomed. Eng. 55, 2509–2518 (2008).
Hu, K. et al. Non-random fluctuations and multi-scale dynamics regulation of human activity. Physica A 337, 307–318 (2004).
Backes, A. et al. Advanced analytical methods to assess physical activity behavior using accelerometer time series: A scoping review. Scand. J. Med. Sci. Sports 32, 18–44 (2022).
Aybek, S. et al. Fractal temporal organisation of motricity is altered in major depression. Psychiatry Res. 200, 288–293 (2012).
Paraschiv-Ionescu, A., Buchser, E., Rutschmann, B. & Aminian, K. Nonlinear analysis of human physical activity patterns in health and disease. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 77, 021913 (2008).
Raichlen, D. A., Klimentidis, Y. C., Hsu, C. H. & Alexander, G. E. Fractal complexity of daily physical activity patterns differs with age over the life span and is associated with mortality in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 74, 1461–1467 (2019).
Li, P. et al. Interaction between the progression of Alzheimer’s disease and fractal degradation. Neurobiol. Aging 83, 21–30 (2019).
Li, P. et al. More random motor activity fluctuations predict incident frailty, disability, and mortality. Sci. Transl. Med. 11, eaax1977 (2019).
Hu, K. et al. Progression of dementia assessed by temporal correlations of physical activity: Results from a 3.5-year, longitudinal randomized controlled trial. Sci. Rep. 6, 27742 (2016).
Gao, L. et al. Fractal motor activity regulation and sex differences in preclinical Alzheimer’s disease pathology. Alzheimers Dement. (Amst.) 13, e12211 (2021).
Li, P. et al. Fractal regulation and incident Alzheimer’s disease in elderly individuals. Alzheimers Dement. 14, 1114–1125 (2018).
Huber, S. E. et al. Assessment of fractal characteristics of locomotor activity of geriatric in-patients with Alzheimer’s dementia. Front. Aging Neurosci. 11, 272 (2019).
Hu, K., Van Someren, E. J., Shea, S. A. & Scheer, F. A. Reduction of scale invariance of activity fluctuations with aging and Alzheimer’s disease: Involvement of the circadian pacemaker. Proc. Natl. Acad. Sci. USA 106, 2490–2494 (2009).
Hu, K., Harper, D. G., Shea, S. A., Stopa, E. G. & Scheer, F. A. Noninvasive fractal biomarker of clock neurotransmitter disturbance in humans with dementia. Sci. Rep. 3, 2229 (2013).
Hausdorff, J. M. et al. Etiology and modification of gait instability in older adults: A randomized controlled trial of exercise. J. Appl. Physiol. 1985(90), 2117–2129 (2001).
Tulppo, M. P. et al. Effects of exercise and passive head-up tilt on fractal and complexity properties of heart rate dynamics. Am. J. Physiol. Heart Circ. Physiol. 280, H1081-1087 (2001).
Soares-Miranda, L. et al. Benefits of achieving vigorous as well as moderate physical activity recommendations: Evidence from heart rate complexity and cardiac vagal modulation. J. Sports Sci. 29, 1011–1018 (2011).
Livingston, G. et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 396, 413–446 (2020).
Livingston, G. et al. Dementia prevention, intervention, and care. Lancet 390, 2673–2734 (2017).
Hofman, M. A. & Swaab, D. F. Living by the clock: The circadian pacemaker in older people. Ageing Res. Rev. 5, 33–51 (2006).
Farajnia, S. et al. Evidence for neuronal desynchrony in the aged suprachiasmatic nucleus clock. J. Neurosci. 32, 5891–5899 (2012).
Stopa, E. G. et al. Pathologic evaluation of the human suprachiasmatic nucleus in severe dementia. J. Neuropathol. Exp. Neurol. 58, 29–39 (1999).
Burdakov, D. & Peleg-Raibstein, D. The hypothalamus as a primary coordinator of memory updating. Physiol. Behav. 223, 112988 (2020).
Rockwood, K. et al. A frailty index based on deficit accumulation quantifies mortality risk in humans and in mice. Sci. Rep. 7, 43068 (2017).
Rockwood, R. & Howlett, S. Fifteen years of progress in understanding frailty and health in aging. BMC Med. 16, 1–4 (2018).
Blodgett, J., Theou, O., Howlett, S. & Rockwood, K. A frailty index from common clinical and laboratory tests predicts increased risk of death across the life course. Geroscience 39, 447–455 (2017).
Hanson, M. A. et al. Developmental aspects of a life course approach to healthy ageing. J. Physiol. 594, 2147–2160 (2016).
Oksuzyan, A., Juel, K., Vaupel, J. W. & Christensen, K. Men: Good health and high mortality. Sex differences in health and aging. Aging Clin. Exp. Res. 20, 91–102 (2008).
Gordon, E. H. et al. Sex differences in frailty: A systematic review and meta-analysis. Exp. Gerontol. 89, 30–40 (2017).
Lee, J., Meijer, E., Phillips, D. & Hu, P. Disability incidence rates for men and women in 23 countries: Evidence on health effects of gender inequality. J. Gerontol. A Biol. Sci. Med. Sci. 76, 328–338 (2021).
Laws, K. R., Irvine, K. & Gale, T. M. Sex differences in cognitive impairment in Alzheimer’s disease. World J. Psychiatry 6, 54–65 (2016).
Mielke, M. M. Sex and gender differences in Alzheimer’s disease dementia. Psychiatr. Times 35, 14–17 (2018).
Seifarth, J. E., McGowan, C. L. & Milne, K. J. Sex and life expectancy. Gend. Med. 9, 390–401 (2012).
Shao, Z., Janse, E., Visser, K. & Meyer, A. S. What do verbal fluency tasks measure? Predictors of verbal fluency performance in older adults. Front. Psychol. 5, 772 (2014).
Gustavson, D. E. et al. Integrating verbal fluency with executive functions: Evidence from twin studies in adolescence and middle age. J. Exp. Psychol. Gen. 148, 2104–2119 (2019).
Mitchell, J. J. et al. Exploring the associations of daily movement behaviours and mid-life cognition: A compositional analysis of the 1970 British Cohort Study. J. Epidemiol. Community Health 77, 189–195 (2023).
Sabia, S. et al. Physical activity, cognitive decline, and risk of dementia: 28 year follow-up of Whitehall II cohort study. BMJ 357, j2709 (2017).
Holtermann, A. et al. 24-Hour physical behavior balance for better health for all: “The sweet-spot hypothesis”. Sports Med. Open 7, 98 (2021).
Wallace, L. M. K. et al. Investigation of frailty as a moderator of the relationship between neuropathology and dementia in Alzheimer’s disease: A cross-sectional analysis of data from the Rush Memory and Aging Project. Lancet Neurol. 18, 177–184 (2019).
Ward, D. D., Ranson, J. M., Wallace, L. M. K., Llewellyn, D. J. & Rockwood, K. Frailty, lifestyle, genetics and dementia risk. J. Neurol. Neurosurg. Psychiatry 93, 343–350 (2022).
Dunn, J., Runge, R. & Snyder, M. Wearables and the medical revolution. Per. Med. 15, 429–448 (2018).
Basza, M. et al. An Apple Watch a day keeps the doctor away?. Cardiol. J. 28, 801–803 (2021).
Elliott, J. & Shepherd, P. Cohort profile: 1970 British Birth Cohort (BCS70). Int. J. Epidemiol. 35, 836–843 (2006).
Hamer, M. et al. Feasibility of measuring sedentary time using data from a thigh-worn accelerometer. Am. J. Epidemiol. 189, 963–971 (2020).
Hardstone, R. et al. Detrended fluctuation analysis: A scale-free view on neuronal oscillations. Front. Physiol. 3, 450 (2012).
Shalalfeh, L. & AlShalalfeh, A. Early warning signals for bearing failure using detrended fluctuation analysis. Appl. Sci. 10, 8489 (2020).
Davis, D. et al. Verbal memory and search speed in early midlife are associated with mortality over 25 years’ follow-up, independently of health status and early life factors: A British birth cohort study. Int. J. Epidemiol. 45, 1216–1225 (2016).
Steptoe, A., Breeze, E., Banks, J. & Nazroo, J. Cohort profile: The English longitudinal study of ageing. Int. J. Epidemiol. 42, 1640–1648 (2013).
Richards, M. et al. Lifetime affect and midlife cognitive function: Prospective birth cohort study. Br. J. Psychiatry 204, 194–199 (2014).
Sutin, A. R., Stephan, Y. & Terracciano, A. Verbal fluency and risk of dementia. Int. J. Geriatr. Psychiatry 34, 863–867 (2019).
Bowling, A., Pikhartova, J. & Dodgeon, B. Is mid-life social participation associated with cognitive function at age 50? Results from the British National Child Development Study (NCDS). BMC Psychol. 4, 58 (2016).
Cerami, C. et al. Clinical validity of delayed recall tests as a gateway biomarker for Alzheimer’s disease in the context of a structured 5-phase development framework. Neurobiol. Aging 52, 153–166 (2017).
Uttl, B. & Pilkenton-Taylor, C. Letter cancellation performance across the adult life span. Clin. Neuropsychol. 15, 521–530 (2001).
Richards, M. & Deary, I. J. A life course approach to cognitive capability. In A Life Course Approach to Healthy Ageing (eds Kuh, D. et al.) (Oxford Scholarship Online, 2014).
Babor, T. F. & Robaina, K. The Alcohol Use Disorders Identification Test (AUDIT): A review of graded severity algorithms and national adaptations. Int. J. Alcohol Drug Res. 5, 17–24 (2016).
Stemland, I. et al. Validity of the Acti4 method for detection of physical activity types in free-living settings: Comparison with video analysis. Ergonomics 58, 953–965 (2015).
Clark, B., Winker, E., Ahmadi, M. & Trost, S. Comparison of three algorithms using thigh-worn accelerometers for classifying sitting, standing, and stepping in free-living office workers. J. Meas. Phys. Behav. 4, 89–95 (2021).
Mishra, P. et al. Descriptive statistics and normality tests for statistical data. Ann. Card. Anaesth. 22, 67–72 (2019).
Rubin, D. B. Multiple Imputation for Nonresponse in Surveys (Wiley, 2004).
Funding
JMB and MA are supported through a British Heart Foundation Grant SP/F/20/150002. ES was funded by the NHMRC (Australia) through an Investigator Grant level 2.
Author information
Authors and Affiliations
Contributions
JMB and MH contributed to the conception and design of the study. MA derived and prepared DFA scalar exponents. JMB performed statistical analyses and wrote the first draft of the manuscript. All authors contributed to the article and approved the submitted version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Blodgett, J.M., Ahmadi, M., Stamatakis, E. et al. Fractal complexity of daily physical activity and cognitive function in a midlife cohort. Sci Rep 13, 20340 (2023). https://doi.org/10.1038/s41598-023-47200-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-47200-x
- Springer Nature Limited