
Abstract
Vegetation structures and dynamics are the result of interactions between abiotic and biotic factors in an ecosystem. The present study was designed to investigate vegetation structure and species diversity along various environmental variables in the Yakhtangay Hills of the Hindu-Himalayan Mountain Pakistan, by using multivariate statistical analysis. Quadrat quantitative method was used for the sampling of vegetation. PC-ORD version 5 software was used to classify the vegetation into different plants communities using cluster analysis. The results of regression analysis among various edaphic variables shows that soil organic matter, total dissolved solids, electrical conductivity, CaCO3 and moisture contents shows a significant positive correlation with species abundance, while the soil pH has inverse relationship with plant species abundance. Similarly, species richness increases with increase in soil organic matter, CaCO3 and moisture contents, while decrease with increase in soil pH, total dissolved solids and electrical conductivity (p < 0.05). The vegetation was classified into four major plant communities and their respective indicators were identified using indicator species analysis. Indicator species analysis reflects the indicators of the study area are mostly the indicators to the Himalayan or moist temperate ecosystem. These indicators could be considered for micro-habitat conservation and respective ecosystem management plans not only in the study area but also in other region with similar sort of environmental conditions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
Environmental gradients are those components of the environment which influence vegetation composition and structure either directly or indirectly1,2. Species that have significant environmental tolerance developed a distinct plant associations with unique floristic and structural attributes3. Vegetation of an area can be classified or categorized into different communities/associations based on various similarities and differences under the influence of different environmental gradients4. Floristic diversity is usually dependent upon environmental factors including topography, soil and climate. Various plants life characteristics such as leaf and life forms (biological spectra), phenological attributes, further mirrors the existence of ecological and natural surrounding5. Whereas, vegetation is the expression of environment inside the specified locality within a specified period, therefore it needs to be properly studied along with it is surrounding both at species and at community level6. The term vegetation indicates the life of plant in a specific region, while flora represents plant composition. Analyzing the characteristics, classification, relationship and distribution of plant communities and the effort to describe the diversity of species inside the plant communities is termed as phytosociology. The primary aim of the phytosociology is describing the vegetation structure and classification by a meaningful way through quantitative method7,8.
The limitation of resources inside the environment affect the plant community properties, structure, species distribution, and ecosystem functions like productivity and nutrient circulation. Though till now a couples of factors has been recognized and a few studies have investigated a single plant or a combined response of a community to such limitations9. Due to the lack of these information which may be concerned to many treatments required to evaluate such responses to both single resource restrictions and their interrelationship with other factors. Each species inside a community might be limited due to the availability or lack of different resources, making community level and individuals plant responses in species-rich communities difficult to interpret10. Despite these challenges, such insight is crucial to understand how individual’s species and communities are affected by natural and anthropogenic disturbances11. Thus, it is crucial to understand the vegetation distribution pattern and influencing factors at different scales12,13. It is very important to understand the vegetation-environment relationship to check the effect of changes in environment14. It has been observed by various plant ecologist that not only at local or regional scale, the diversity of plants may be influenced by abiotic conditions, but also depend on other ecosystem processes, such as biotic interaction and seed dispersal limitations15,16. The limiting factors inside any biome, limit the total number and diversity of the species. While other environmental gradients, particularly topography, elevation, snow cover, soil moisture and pH, have been analyzed and concluded that these are correlated with plant species richness at both regional and local levels17,18,19.
Among the abiotic gradients, topography contributing to the physiognomic differentiation for vegetation and thus play a significant role in the heterogeneity of habitat20,21,22,23 and consequently changing the structure of plant community and their species composition. An extensive variation in mountain landscape variation regarding to the soil age, erosion rates, topography, hydrology, among other factors has effects the structure, composition and ultimately function of the ecosystem24,25,26,27. Landscape matrices are also molded by various topographic element, such as altitude, aspect, and slope28. Therefore, it is important to investigate the vegetation at multiscale rather than through a single scale, to fulfill different needs for resource management. Altitudinal gradient also affect vegetation structure and species composition of different ecosystem and various studies indicated that plants diversity is highest at intermediate elevation, like in tropical rain forests29,30,31,32. Moreover, a humped (unimodal) curve between species diversity, and the increase in altitude was observed at Himalayan Mountain23.
Among the edaphic factors, the soil nutrients play an important role influencing certain plants parameters such as tree height, basal area, canopy cover and consequently influencing the structure of plant communities33,34. Several studies have been carried out to check the relationship between the change in species richness and the availability of soil gradients35,36. Generally lower species richness was observed at lower nutrient level, that increases with the intermediate level of the nutrients while gradually decline at high nutrient level making a typical “humped back curve” in response to the nutrient gradient.
This pattern has been found by a numbers of plants ecologist37,38. Soil pH is one of the basic environmental gradients suggested to be an important driver of the number of vascular species at the local scale (fine-scale species richness, alpha diversity)39. It has been suggested that on acidic pH (pH < 4), the number of species could be constrained by high phytotoxicity 40,41,42,43 and limit nutrition availability. It has been observed that on high-pH soils (pH > 7), phosphorus and iron become limitedly available nutrients because of their decrease in solubility and thus affecting on species diversity of an area. Slope facing to poles are wetland cool, with rich organic matter and deeper soil and characterized by mesic vegetation, while the slope facing toward equator are usually hot and xeric type with low soil nutrients and thus having severe erosion, subsequently they are supporting xeric type of vegetation44,45. It has a common observation that sunny slope retains less moisture compared to shady slope, because of greater solar radiation and high evaporation. Therefore, the vegetation of the sunny slope such as grasses must be xeric type, drought tolerating and radiation resistant. On the other hand, shade loving plant such as forbs are dominant on shady slopes. Soil moisture availability is a key driver for both biological and chemical processes and one of the important limiting factors responsible for the productivity of plant46,47. Owing to the response, vegetation type and structure and type can influence soil water dynamics and soil characteristics including soil hydraulic characteristics48,49,50. Taking water as a source of nutrients if measured generate a similar humped shaped curve with vegetation. Usually, plant species richness shows a positive logarithmic relationship with rainfall51,52. The geological conditions, seawater, and domestic activities (domestic, agricultural, and industrial effluents) are the prime source of the particulate matters53. Due to the rapid changes in landscape, climate and geo-climate history mountain ecosystem possess diverse biological communities all over the world.
Therefore, this study was designed to analyze vegetation of the Yakhtangay hill, Hindu-Himalayan series of Pakistan in relation to various physiographic and edaphic variables. It is hypothesized that slope, altitude, and aspect of the mountain plays a significant role in vegetation structure and species diversity. We aim to analyze plants assemblage i.e., communities’ formation under the influence of environmental variables including edaphic and climatic variables on species diversity i.e., species abundance and richness by using recently developed analytical techniques. This study will answer how to classify and identify indicator species of particular habitat or community.
Materials and methods
Study area
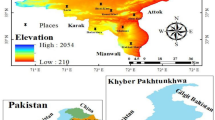
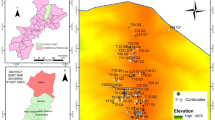
District Shangla is located at western extremities of the Himalayan and eastern Hindu Kush Mountain series generally termed as Hindu-Himalayas, in the Khyber Pakhtunkhwa province of Pakistan. Topographically, it consists of narrow valleys and high mountains covered by dense forest at elevation range from 1300 to 3000 masl54. The Yekhtangay hill is one of such high mountains famous tourist spots, located at 34°-31' to 33°-08' north latitude and 72°-33' to 73°-01 south longitude at about 2200 masl. Temperature of the study area observed severe cold in winter and moderate in summer. The area is covered mostly by snow, usually from November to April that makes winter season more prolonged than summers. Mountains of the study area possess diverse moist temperate forest. The inhabitants in the foothill of Yakhtangay hills are dependent on the forest resources as source of earning, agricultural tools and for fuel purposes. Therefore, the forest of the area is under severe anthropogenic pressure. The soil of the area is mostly clay suitable for the growth of diverse vegetation (Fig. 1, Table 1; Supplementary Table S1)55.

GIS generated map of the study area representing the sampling localities56.
Data collection
The plants data of summer and spring were collected during the months of July–August 2020 and April-May 2021, respectively. Transect and quadrat quantitative methods were applied for the sampling of vegetation57. Permission was taken from the local forest and wildlife department for carrying out plants collection activities in the forest. Quadrats were placed systematically along each transect at 200 m intervals using Global Positioning System (GPS). Quadrats having size of 20 × 20 m, 10 × 10 m and 2 × 2 m were taken place for tree, shrubs and herb species, respectively58. Quantitative data for each plant species were recorded to measure the phytosociological attributes including total frequency, density and cover. Plant specimens were collected and identified with help of Flora of Pakistan and expert taxonomist59. The dried specimens were preserved in the Herbarium of Pakistan, Quaid -i-Azam University, Islamabad, Pakistan. The biological spectra such as leaf form and life from were determined for each plant species using Raunkiaer60 classification.
Phytosociological attributes/quantitative characteristics of the vegetation
The quantitative characteristics of vegetation such as density, relative density, abundance, cover, relative cover, frequency, relative frequency, and importance value index were calculated by using the following various formulas, followed by61,62.
Topographic and climatic information
The topographic and climatic factors such as altitude, aspect, slope angle, availability of sunlight, moisture and soil characteristics greatly influence vegetation structure, species composition and community structure in one or other way. Topographic gradients of the area such as altitude, elevation, height, slope angle, aspect were measured through the GPS essential application. The altimeter was used to record the altitude data, while a compass was used to check north, south, east, and west aspect of the mountain.
Soil analysis
Soil samples were collected from 1 to 15 cm depth with the help a digger. About 20–25 g of soil samples from three different spots of each quadrat were collected and homogenized into one sample. A specific tag number was labeled according to the specific quadrat63. Various analysis of the soil were performed including soil moisture contents, texture, organic matter, calcium carbonates, Electrical Conductivity (EC), Total Dissolved Solids (TDS) and soil pH using different standard protocols48,64,65.
Statistical data analysis
PC-ORD (version 5) software was used to classify the plant species and stations/quadrat into potential plant communities based on their similarities and differences66. Indicator species were identified using Indicator Species Analysis (ISA). While CANOCO software (version 4.5) was used for canonical correspondence analysis to investigate the relationship between vegetation and environmental variables67,68,69. Regression analysis was used for the assessment of the relationship between species richness/abundance along with various environmental variables using R Software (https://cran.r-project.org/bin/windows/base/)70.
Results
Floristic composition and biological spectra of the vegetation in the study area
A total of 114 plant species belong to 45 families were recorded from the studied region. Rosaceae was recorded as dominant family followed by Poaceae and Asteraceae (Supplementary Table S2). The dominant species were Pinus wallichiana (IVI = 2367), Viburnum grandiflorum (IVI = 2118), Buxus wallichiana (IVI = 1697) and Bistorta amplexicaulis (IVI = 450) (Supplementary Table S3). While among the leaf form, microphylls was the dominant (24 species that constituents about 33% of the total plants), followed by leptophylls (24%) and nanophyll (16%). Hemicryptophytes and nanophanerophytes were the dominant life forms that constituents about 52% of the total plant species. While therophytes constituents about 20% followed by mesophanerophytes (8%) in the region.
Impact of edaphic factors on plant species abundance
Impact of edaphic factors on plant species were analyzed to check whether these variables are significantly representing the vegetation abundance of the area or not. The data of edaphic variables and species abundance were analyzed in” R” software version 4.0.4 through regression analysis. The result showed that the soil organic matter, TDS, EC, CaCO3 and moisture contents have a linear relationship (i.e., positive correlation) with species abundance (with degree of correlation, R = 0.16, R = 0.33, R = 0.21, R = 0.20, R = 0.47, respectively) at significant level (p value < 0.05). While the relationship between soil pH and plant species abundance represent a negative correlation with R = − 0.19 and p value 0.001 (Fig. 2).

Regression analysis between soil variables and plant species abundance. Impact of soil (A) TDS (B) EC (C) pH (D) OM (E) CaCO3 and (F) moisture contents with species abundance.
Impact of edaphic factors on species richness
The regression analysis between species richness and edaphic factors shows that soil pH, TDS and EC are negative correlated with species richness having beta values i.e., R = − 0.65, R = − 0.46 and R = − 0.78, respectively at p value < 0.05 significance level. The relationship between soil organic matter (R = 0.89), calcium carbonates (R = 0.78) and moisture contents (R = 0.84) were positively correlated with species richness at, p-value < 0.05 (Fig. 3).

Regression analysis between soil variables and plant species richness. Impact of soil (A) EC (B) TDS (C) pH (D) OM (E) CaCO3 (F) moisture contents with species richness . Here, OM organic matter, EC electrical conductivity, TDS total dissolved solvents.
Species area curve
We used the PC-ORD version 5 software to find that either the quadrat size was adequate or not through abundance data combined with Sorenson distance value in the form of species area curve and compositional area curve61. The result shows that the quadrat number 10 shows maximum numbers of plant species appearing and are continued up to quadrat number 40, then declines till the last quadrat with minimum number of species illustrating the adequacy of the sample size (Fig. 4).

Species area curves representing relationship between occurring of new plant species along the sampling quadrats through compositional area curves.
Formation of plant communities through cluster analysis (CA)
The result of cluster analysis classified all the quadrat into 4 major plant communities (Fig. 5). The detail description of each plant community are as follows:

Cluster analysis classified all the stations quadrats into 4 major communities.
Pinus wallichiana-Berberis lycium-Impatiens brachycentra plants community
The community name is based on the topmost indicator species. This community was observed at an elevation range of 1666–2882 m. The topmost indicator species of this community were Pinus wallichiana, Berberis lyceum and Impatiens brachycentra, one each from tree, shrubs and herbs, respectively (Fig. 6). The identified indicator species were the indicators of low altitude, soil pH, silt contents, and high percentage of calcium carbonates, clay, EC, soil moisture, organic matter, TDS and slope angle, at significant level (p ≤ 0.005).
Abies pindrow, Rhus javanica, Quercus dilatata were the dominant, while Prunus persica, Diospyros lotus, Pistacia chinensis were the rare tree species of this community. The dominant shrub species having high IVI of this community were Viburnum grandiflorum, Berberis lyceum, while the rare shrubs were Rubus ulmifolius, Rubus occidentalis, Zanthoxylum armatum,. The rare herb species of this community were Dryopteris ramosa, Bistorta amplexicaulis, Arisaema flavum, while Eulaliopsis binata, Solidago virga-aurea and Galinsoga parviflora were the rare herb species.
Edaphic and environmental gradients of this community ranges as, pH = 4.91–6.8, EC = 43–687 (µS), TDS = 44–428 (ppm), CaCO3 = 4.04–16.12%, OM = 0.85–9.85%, moisture contents = 2–32%, silt=14–52%, sand = 2–52%, clay = 10–78%, slope = 113–168° (Supplementary Table S1).

Indicator species biplot for (a) Pinus wallichiana, (b) Berberis lyceum and (c) Impatiens brachycentra top 3 indicators of first plant community in relation to various environmental variables.
Quercus incana-Buxus wallichiana-Agrostis gigantea (QBA) community
This community was observed at an elevation range of 1824–2258 m. The topmost indicator species of this community were Quercus incana, Boxus wallichiana and Agrostis gigantea (Fig. 7). The indicator species observed in this community were clustered around acidic pH, low silt, clay contents, calcium carbonates, moderate amount of organic matter, high level of TDS, EC, moisture contents and high slope angle at significant level (p ≤ 0.005). The values of edaphic and environmental gradients of this community ranges as, pH = 3.9–7.8, EC = 23–487 (µS), TDS = 41–288 (ppm), CaCO3 = 5.36–22.13%, OM = 3.10–12.1%, moisture contents = 7–39%, silt = 18–58%, sand = 3–56%, clay = 11–48%, slope = 110–1156°.

Indicator species biplot for (a) Quercus incana, (b) Boxus wallichiana and (c) Agrostis gigantea, top 3 indicator plants of 2nd plants community after CCA of Canoco software.
Abies pindow-Rubus fruticosus-Poa annua (ARP) community
The indicator species of this community were Abies pindrow, Rubus fruticocus and Poa annua (Fig. 8). The elevation range of this community was 2246–2824 m. The indicator species observed in this community have a significant positive relation with altitude, EC, less amount of CaCO3, moderate amount of clay, silt contents, moisture, neutral soil pH and less slope angle, while it was observed that these indicators were negatively correlated with the percentage of organic matter and TDS (p ≤ 0.005).
The dominant tree species having higher IVI were Rhus javanica and Quercus dilatata, while the tree species having lower IVI included Cedrus deodara, Pistacia chinensis subsp. integerrima, Taxus baccata. The abundant shrub species of this community having higher IVI were Viburnum grandiflorum, Parrotiopsis jacquemontiana while the shrub species were Crataegus songarica, Berberis asiatica, Isodonrugosus, Viburnum cotinifolium. The dominant herb species of this community were Urtica dioca, Carex cardiolepis, while the rare herb species of this community were Achyranthes aspera, Tussilago farfara, and Adiantum venustum. Edaphic and environmental gradients of this community ranges as, pH = 4.7–7.6, EC = 72–380 (µS), TDS = 51–239 (ppm), CacO3 = 6.36–19.13%, OM = 3.81–10.1%, moisture contents = 5–29%, silt = 16–54%, sand = 6–36%, clay = 13–67%, slope = 118–164° (Supplementary Table S1).

Indicator species biplots for (a) Abies pindrow, (b) Rubus fruticocus and (c) Poa annua in relation to various environmental factors.
Taxus baccata-Rubus ellipticus-Potentilla atrorubens (TRP) community
The indicator species of this community were Taxus baccata, Rubus ellipticus and Potentella atrorubens ranging at the altitude of 2200–3028 m (Fig. 9). The indicator species observed in this community are clustered in relation to the lower altitude, less amount of calcium carbonate, clay contents, slope angle, TDS, higher pH, silt contents, moderate moisture and EC at p ≤ 0.005.
The dominant tree species of this community was Abies pindrow, while the rare tree species having lower IVI were Cedrus deodara and Quercus dilatata. The dominant shrub species of this community was Viburnum grandiflorum, while the rare shrub species was Isodonrugosus. The dominant herb species of this community were Bistorta amplexicaulis, Muhlenbergia duthieana, Dryopteris ramosa, while the rare herb species of this community were Diplazium esculentum, Tussilago farfara, and Dryopteris marginali.
The edaphic and environmental gradients of this community ranges as, pH = 5.1–7.2, EC = 53–403 (µS)s, TDS = 49–269 (ppm), CaCO3 = 2–18%, OM = 1.7–9.65%, moisture contents = 1–8%, silt = 23–68%, sand = 8–61%, clay = 10–41%, slope = 109–146° (Supplementary Table S1).

Indicator species biplots for (a) Taxus baccata, (b) Rubus ellipticus and (c) Potentella atrorubens top three indicators of 4th plants community.
Canonical Correspondence Analysis (CCA)/species ordinations under the influence of environmental gradients
All the environmental variables including altitude, slope, aspect and edaphic variables i.e., organic matter, pH, CaCO3, sand, silt and slay contents, soil moisture, EC and TDS significantly affecting the vegetation characteristics such as species abundance, distribution, and evenness in the studied area (p ≤ 0.001). Each border or cross in ordination bi-plot representing plant species in relation with environmental gradients, and the distance between them shows the differences index. The direction and length of arrows directed environmental variables in the CCA biplot, while the triangle shape indicated plant species based upon the relationship with environmental variables (Fig. 10). The length of the arrow indicated the dimension between different variables and degree of correlation between them. The variables occupying on the same axis shows direct relationship, while those on the opposite direction indicated an inverse relationship.
The first quadrant of CCA bi-plot shows that the plants including Adiantum emarginatum, Achyranthes aspera, Quercus dilatata, Arisaema flavum, Buxus wallichiana were primarily associated with soil pH and silt contents. Plant species i.e., Debregea siasaeneb, Quercus baloot, Diospyros lotus, Rubus ulmifolius, Arisaema jacquemontii etc. were mostly associated with EC, TDS, soil moisture and clay natured soil in the 2nd quadrant of canonical correspondence analysis. The plant species including Berberis vulgaris, Dryopteris marginalis, Pinus wallichiana, Urtica dioca, Poa annua etc were clustered around organic matter, calcium carbonate and slope angle at 3rd quadrant . The 4th quadrant of the CCA bi-plot shows the species that are associated with altitude of the mountain and sandy nature of the soil. These species were Viburnum grandiflorum, Abies pindrow, Rhus javanica, Taraxacum officinale, and Zanthoxylum armatum etc (Table 2; Fig. 10).

Canonical correspondence analysis (CCA) of 114 plant species in relation to measured environmental variables.
Discussion
The vegetation structure, dynamics and distribution pattern are significantly influenced by environmental factors within and among ecosystems71. The present study reported 114 plant species, with maximum number of Rosaceae family, followed by Poaceae and Asteraceae. The above mentioned families are widely distributed across the globe having broad ecological amplitude that makes the possibility of their distribution particularly in microhabitats of temperate forest. These findings can be compared with the studies of Himalaya’s mountain where Rosaceae and Asteraceae were found as most dominant families72. The predominance of family Rosaceae may also be due to its central distribution from temperate to subtropical of the northern hemisphere. Our results are also comparable with the studies of Ilyas et al. & Amjad et al.73,74 reporting Poaceae and Ateraceae as the dominant families in Kabal Valley Swat, Valley and subtropical forest of Kotli Azad Kashmir, respectively. It was further observed that these families have characteristics similarities with the vegetation communities of Malakand, Hazara, Kashmir, and Gilgit Baltistan, Pakistan, where Asteraceae, Lamiaceae, Rosaceae, and Poaceae were reported as the uppermost prominent groups53,75,76.
The current study reported that the vegetation at higher altitudes contains typical moist temperate forest species, including Pinus wallichiana, Abies pindrow, Viburnum grandiflorum and Bistorta amplexicaule, which are in close harmony with the assemblage reported in the moist temperate Himalaya77,78. Among these Pinus walllichianais the dominant tree in the region under consideration with highest IVI ( 1047), which corroborates with the findings of74,79 from western Himalayan region. The topography and altitude have negative relationship with the vegetation species richness and abundance, which tends to decrease along the altitude. Similarly Kharkwal et al.80, also reported the same pattern of species abundance and richness from Kuman, India. Our results are also in line with the findings of Bano et al.58, they also concluded that there was a decrease in numbers and diversity of species along the altitude in Beer hills, district Haripur, Khyber Pakhtunkhwa, Pakistan. The members of the family Poaceae were most common on the east slope compared to west slope due to more isolation, sandy nature of soil and less moisture contents, which provide a semi-xeric condition to the vegetation. Some xeric types of grasses like Eulaliopsis binata and Muhlenbergia duthieana were recorded as well on the same slope. The western slope of the mountain was dominated by some members of Gymnosperms such as Pinus, Abies, Taxus and moist temperate indicator species such as Viburnum cotinifolium, Viburnum grandiflorum, Berberis lycium, Dryopteris ramosa, Urtica dioca etc. These species are supported by the soil moist conditions, less availability of sunlight and more clay contents in soil. Various studies from semiarid region reported significant influence of microclimatic condition (such as air, soil temperature, evapotranspiration, wind speed along with edaphic factors (organic matter, soil moisture, soil depth and soil texture) on vegetation that subsequently results in distinct vegetation types inhabiting on different slope)81.
The attributes of plant species such as life form, leaf form (biological spectra), phenological trait mirrors the prevailing status of ecological and natural environmental condition of the region. The life form of a plant species is the reflection of adaptation to micro and macro climatic conditions and used frequently in vegetation assessment 82. In the current study, the dominant life form was hemicryptophytes (26%) followed by nanophanerophytes (26%) therophytes (20%) and mesophanerophytes (8%). These present findings show similarities with Shimwell83 work as they reported hemicryptophytes as a dominant life form and considered as an indicator of the moist and cold temperate vegetation. A study was also carried out by Malik84 in Ganga Chotti and Badhori hills at Bagh district Azad Jammu Kashmir and reported that hemicryptophytes is the dominant life form in western Himalayas. Among the leaf form microphyll (33%) was the dominant most followed by leptophyll (24%) and mesophyll (16%). Our findings correlates with the studies of Khan et al.85 where they reported microphyll was dominant life form followed by leptophyll in moist temperate forest of Thandiani, Abbottabad, Khyber Pakhtunkhwa. Species with comparatively smaller leaves symbolize dry and cold climate, while larger leaves species occurs in warmer and moist climate. In current study microphyll were evident at higher altitude whereas leptophyll at lower elevation. Small leaves have been linked to cold or hot desert condition, which is an adaptive feature in preserving soil moisture. When the root is sensitive to low temperatures, which causes a decrease in water absorption from the soil, moisture retention is essential86. The large number of microphylls suggests a cool, sub-alpine, meadow climate with little moisture and nutrient absorption by the roots.
The total plant species in the study area were classified into 4 major plant communities through PC-ORD version 5. Bano et al.58 also identified 4 major plants communities in their study from Beer hills along the Indus River in district Haripur, Pakistan using the same statistical tool. Iqbal et al.87 conducted a study on Biha Valley, of Hindukush Mountain, District Swat, Pakistan and identified 7 different plant communities, using Two-way way cluster analysis classification. A phytosociological study was carried out by Ahmed77 in Himalayan forests of Pakistan and identified twenty-four different plant communities. The difference in number of the communities formed in any region might be due prevailing environmental conditions in those habitats.
The first plant community established at an elevation range of 1666–2882 m in the current study. The dominant species of this community were Pinus wallichiana, Berberis lyceum, and Impatiens brachycentra. It was influenced by environmental factors that ranges pH = 4.91–6.8, EC = 43–687 (µS), TDS = 44–428 (ppm), CaCO3 = 4.04–16.12%, OM = 0.85–9.85%, moisture contents = 2–32%, 14–52%, sand = 2–52, Clay = 10–78%, Slope = 113–168°. The dominant species of this community was influenced by EC, TDS, and clay content. Community 2 occurs at an elevation range of 1824-2258 m and dominated by Quercus incana, Boxus wallichiana and Agrostis gigantea. The dominant species have significant relationship with EC, TDS, Sand, pH, CaCO3, OM, clay content, sand and silt. Community 3 established at an elevation range of 2246–2824 m and dominated by Abies pindrow, Rubus fruticocus and Poa annua. The dominant species have strong relationship with soil pH, sand, altitude, CacO3 and OM. Community 4 established at the elevation range of 2200–3028 m and dominated by Taxus baccata, Rubus ellipticus and Potentella atrorubens. The dominant species of this community have strong relationship with pH, sand and altitude. Our finding of classification plant communities and identification of indicator species corroborates with various research studies in the adjacent mountainous75,79,88,89,90,91. Species richness and abundance is significantly influenced by the environmental variables including OM, TDS, OM, soil moisture and pH. Species abundance is positively correlated with environmental variable (OM, TDS, EC, Soil moisture) except soil pH. The direct correlation of quantitative characteristics of biodiversity with soil salinity, TDS and EC also reported by Canfora et al. & Liu et al.92,93. The moisture content significantly increases species richness and abundance, which are in line with the studied of Niu et al. 94 where species richness decreases by 3.56% with 10 degree decreases in the soil moisture. So, we can say that decreasing moistures also decreases in species richness.
We used CCA analysis to further confirm the significant relationship of different environmental variable (such as slope, topography, elevation, soil EC, TDS, moisture contents, soil texture, pH, CaCO3, OM etc.) with vegetation structure. The CCA bi-plot shows that most of the species influenced by EC, TDS, and CaCO3, slope, soil moisture contents and clay, silt, pH contents of the soil. These findings shows similarity with the study conducted, in Suez region of Egypt, that different physiographic and soil variables (such as latitude, longitude altitude pH, TDS, CaCO3, silt and sand) contributes to the distribution of vegetation pattern. The regression analysis among various environmental variables with species richness and abundance clearly revealed that the effect of a single environmental variables such as soil EC, TDS, CaCO3, pH and moisture contents upon species abundance is steady, it means that the abundance of a species is likely affected by a specific environmental variable. While the richness of plant species affected by different variables in different ways due to various in demands and needs of the species. Similarly, Nadal-Romero et al.95, also conducted research and analyzed environmental factors (i.e., soil properties and aspect) and species diversity. Based upon these results from different regions of the world they concluded that not only species diversity is totally dependent on soil characteristics, but also on topography as well81. Slope and aspect are also known to affect the diversity and density of plant communities.
The studies reveal that Pinus wallichiana and Viburnum grandiflorum, are also the indicator species of moist temperate forest87. Furthermore, remaining species are somewhat native or exotic to the moist temperate forest ecosystem. While among the herbs and shrubs Buxus wallichiana (IVI = 568) and Dryopteris ramosa (IVI = 165) respectively were also founded abundant in the area. A similar study was also carried out by Haq et al.58, where they find out herbaceous vegetation as a dominant form in Beer Hills district Haripur, Khyber Pakhtunkhwa in the Himalayan range of Pakistan.
Conclusion
It is concluded that the Yakhtangay Hills in the western Hindu-Himalayan series has a considerably rich phytodiversity. Soil organic matter, TDS, EC, CaCO3 and moisture contents showed a linear correlation (i.e. positive correlation) with species abundance, while inverse relationship was recorded between pH and plant species abundance. Similarly, a significant positive correlation exist between species richness and soil organic matter, calcium carbonate and moisture contents while significant inverse correlation exist between species richness and soil pH, TDS and EC in the studied mountainous region. Pinus wallichiana, Viburnum grandiflorum, Berberis lycium, and Dryopteris ramosa were recorded as an indicators of moist temperate ecosystem. CCA ordination analysis showed that all the measured edaphic and climatic variables have significant impact on the distribution pattern and composition of plant species in the studied region. Statistical tool and techniques used in the current study can be used for the classification of plant communities, identification of indicator plants, impact of environmental gradient on vegetation complexity and dynamics in any micro or macro habitat of the world. The dominant and rare species of the Yakhtangay Hills in the District Shangla can be declared a site for conservation priority as a micro-habitat representative of the Hindu Kush Himalayas ecotonal space.
Data availability
All data generated or analyzed during this study are included in this published article (and its Supplementary Information files).
References
Khan, M. et al. Plant species and communities assessment in interaction with edaphic and topographic factors; an ecological study of the mount Eelum District Swat Pakistan. Saudi J. Biol. Sci. 24(4), 778–786 (2017).
Ur Rahman, A. et al. Impact of multiple environmental factors on species abundance in various forest layers using an integrative modeling approach. Global Ecol. Conserv. 29, e01712 (2021).
Arneth, A., Uncertain future for vegetation cover. Nature 524(7563), 44–45.
Goldsmith, F., Description and analysis of vegetation. Methods Plant Ecol. (1976).
Rahman, I. U. et al. First insights into the floristic diversity, biological spectra and phenology of Manoor Valley Pakistan. Pak. J. Bot 50(3), 1113–1124 (2018).
Khan, S.M., Plant communities and vegetation ecosystem services in the Naran Valley, Western Himalaya, 2012, University of Leicester.
Haq, F., Ahmad, H. & Iqbal, Z. Vegetation description and phytoclimatic gradients of subtropical forests of Nandiar Khuwar catchment District Battagram. Pak. J. Bot 47(4), 1399–1405 (2015).
Iqbal, M. et al. A novel approach to phytosociological classification of weeds flora of an agro-ecological system through Cluster, two way cluster and indicator species analyses. Ecol. Ind. 84, 590–606 (2018).
Shaw, M. R. et al. Grassland responses to global environmental changes suppressed by elevated CO2. Science 298(5600), 1987–1990 (2002).
Drenovsky, R.E., Effects of mineral nutrient deficiencies on plant performance in the desert shrubs Chrysothamnus nauseosus ssp. consimilis and Sarcobatus vermiculatus2002: University of California, Davis.
Iqbal, M. et al. Vegetation classification of the Margalla Foothills, Islamabad under the influence of edaphic factors and anthropogenic activities using modern ecological tools. Pak. J. Bot 53(5), 1831–1843 (2021).
Bai, Y. et al. Landscape-level dynamics of grassland-forest transitions in British Columbia. J. Range Manag. 57(1), 66–75 (2004).
Zhao, T. et al. Retrievals of soil moisture and vegetation optical depth using a multi-channel collaborative algorithm. Remote Sens. Environ. 257, 112321 (2021).
Austin, M., Chapter 2: Vegetation and environment: discontinuities and continuities. IN VAN DER MAAREL, E.(Ed.) Végétation ecology. Etats‐Unis, 2005, Blackwell Publishing.
Peña-Claros, M. et al. Soil effects on forest structure and diversity in a moist and a dry tropical forest. Biotropica 44(3), 276–283 (2012).
Miao, R. et al. Effects of long-term grazing exclusion on plant and soil properties vary with position in dune systems in the Horqin Sandy Land. CATENA 209, 105860 (2022).
Abbas, Z. et al. Plant communities and anthropo-natural threats in the Shigar valley,(Central Karakorum) Baltistan-Pakistan. Pak. J. Bot. 52, 987–994 (2020).
Anwar, S., et al., Plant diversity and communities pattern with special emphasis on the indicator species of a dry temperate forest: A case study from Liakot area of the Hindu Kush mountains, Pakistan. Trop. Ecol. 1–16 (2022).
Mumshad, M. et al. Phyto-ecological studies and distribution pattern of plant species and communities of Dhirkot, Azad Jammu and Kashmir, Pakistan. PLoS ONE 16(10), e0257493 (2021).
Baldeck, C. A. et al. Soil resources and topography shape local tree community structure in tropical forests. Proc. R. Soc. B Biol. Sci. 280(1753), 20122532 (2013).
Guerra, T. N. F. et al. Influence of edge and topography on the vegetation in an Atlantic Forest remnant in northeastern Brazil. J. For. Res. 18(2), 200–208 (2013).
Townsend, A. R., Asner, G. P. & Cleveland, C. C. The biogeochemical heterogeneity of tropical forests. Trends Ecol. Evol. 23(8), 424–431 (2008).
Becknell, J. M. & Powers, J. S. Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest. Can. J. For. Res. 44(6), 604–613 (2014).
Geri, F., Rocchini, D. & Chiarucci, A. Landscape metrics and topographical determinants of large-scale forest dynamics in a Mediterranean landscape. Landsc. Urban Plan. 95(1–2), 46–53 (2010).
Lomolino, M. V. Elevation gradients of species-density: historical and prospective views. Glob. Ecol. Biogeogr. 10(1), 3–13 (2001).
Zhang, K. et al. An integrated flood risk assessment approach based on coupled hydrological-hydraulic modeling and bottom-up hazard vulnerability analysis. Environ. Model. Softw. 148, 105279 (2022).
Liu, Y. et al. A hybrid runoff generation modelling framework based on spatial combination of three runoff generation schemes for semi-humid and semi-arid watersheds. J. Hydrol. 590, 125440 (2020).
Mir, A. Y. et al. Ethnopharmacology and phenology of high-altitude medicinal plants in Kashmir Northern Himalaya. Ethnobot. Res. Appl. 22, 1–15 (2021).
Vetaas, O. R. & Grytnes, J. A. Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal. Glob. Ecol. Biogeogr. 11(4), 291–301 (2002).
Li, W. et al. Fine root biomass and morphology in a temperate forest are influenced more by the nitrogen treatment approach than the rate. Ecol. Ind. 130, 108031 (2021).
Su, N. et al. Landscape context determines soil fungal diversity in a fragmented habitat. CATENA 213, 106163 (2022).
Yang, Y., et al., Nitrogen fertilization weakens the linkage between soil carbon and microbial diversity: a global meta‐analysis. Global Change Biol. (2022).
Ahmad, Z. et al. Weed species composition and distribution pattern in the maize crop under the influence of edaphic factors and farming practices: A case study from Mardan Pakistan. Saudi J. Biol. Sci. 23(6), 741–748 (2016).
Rahman, A. U. et al. Ecological assessment of plant communities and associated edaphic and topographic variables in the Peochar Valley of the Hindu Kush mountains. Mt. Res. Dev. 36(3), 332–341 (2016).
Ashton, P. S. A contribution of rain forest research to evolutionary theory. Ann. Mo. Bot. Gard. 64(4), 694–705 (1977).
Yang, Y. et al. Negative effects of multiple global change factors on soil microbial diversity. Soil Biol. Biochem. 156, 108229 (2021).
Pärtel, M. Local plant diversity patterns and evolutionary history at the regional scale. Ecology 83(9), 2361–2366 (2002).
Taylor, D.R., Aarssen, L.W., & Loehle, C. On the relationship between r/K selection and environmental carrying capacity: A new habitat templet for plant life history strategies. Oikos 239–250 (1990).
Knapp, A. K. et al. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science 298(5601), 2202–2205 (2002).
Zscheischler, J. et al. Short-term favorable weather conditions are an important control of interannual variability in carbon and water fluxes. J. Geophys. Res. Biogeosci. 121(8), 2186–2198 (2016).
Gao, C. et al. Simulation and design of joint distribution of rainfall and tide level in Wuchengxiyu Region China. Urban Clim. 40, 101005 (2021).
Wang, S. et al. Exploring the utility of radar and satellite-sensed precipitation and their dynamic bias correction for integrated prediction of flood and landslide hazards. J. Hydrol. 603, 126964 (2021).
Zhang, K. et al. The sensitivity of North American terrestrial carbon fluxes to spatial and temporal variation in soil moisture: An analysis using radar-derived estimates of root-zone soil moisture. J. Geophys. Res. Biogeosci. 124(11), 3208–3231 (2019).
Yang, Y. et al. Increasing contribution of microbial residues to soil organic carbon in grassland restoration chronosequence. Soil Biol. Biochem. 170, 108688 (2022).
Li, J. et al. Differential mechanisms drive species loss under artificial shade and fertilization in the Alpine Meadow of the Tibetan Plateau. Front. Plant Sci. 13, 832473–832473 (2022).
Fischer, C. et al. How do earthworms, soil texture and plant composition affect infiltration along an experimental plant diversity gradient in grassland?. PLoS ONE 9(6), e98987 (2014).
Zhao, T. et al. Soil moisture experiment in the Luan River supporting new satellite mission opportunities. Remote Sens. Environ. 240, 111680 (2020).
Marandi, A., Polikarpus, M. & Jõeleht, A. A new approach for describing the relationship between electrical conductivity and major anion concentration in natural waters. Appl. Geochem. 38, 103–109 (2013).
Xu, J. et al. Modeling of coupled transfer of water, heat and solute in saline loess considering sodium sulfate crystallization. Cold Reg. Sci. Technol. 189, 103335 (2021).
Chen, X. et al. Spatiotemporal characteristics and attribution of dry/wet conditions in the Weihe River Basin within a typical monsoon transition zone of East Asia over the recent 547 years. Environ. Model. Softw. 143, 105116 (2021).
Ali, G., Siddique, S. & Suliman, M. Effect of canopy cover on natural regeneration of pinus wallichiana in moist temperate forest of Yakh Tangay, District Shangla Swat Pakistan. FUUAST J. Biol. 8(2), 193–201 (2018).
Zhang, K. et al. Characteristics and influencing factors of rainfall-induced landslide and debris flow hazards in Shaanxi Province, China. Nat. Hazard. 19(1), 93–105 (2019).
Khan, W. et al. Vegetation mapping and multivariate approach to indicator species of a forest ecosystem: A case study from the Thandiani sub Forests Division (TsFD) in the Western Himalayas. Ecol. Ind. 71, 336–351 (2016).
Iqbal, J. & Ahmed, M. Vegetation description of some pine forests of Shangla district of Khyber Pakhtunkhwa Pakistan: A preliminary study. FUUAST J. Biol. 4(1), 83–88 (2014).
Sparrow, B. D. et al. A vegetation and soil survey method for surveillance monitoring of rangeland environments. Front. Ecol. Evol. 8, 157 (2020).
Esri, R., ArcGIS desktop: release 10. Environmental Systems Research Institute, CA (2011).
Salzer, D., & Willoughby, J. Standardize this! The futility of attempting to apply a standard quadrat size and shape to rare plant monitoring. in Proceedings of the symposium of the North Coast Chapter of the California Native Plant Society: the ecology and management of rare plants of northwestern California. Arcata, CA. Sacramento, CA: The California Native Plant Society (2004).
Bano, S. et al. Eco-Floristic studies of native plants of the Beer Hills along the Indus River in the districts Haripur and Abbottabad Pakistan. Saudi J. Biol. Sci. 25(4), 801–810 (2018).
Perveen, A. & Qaiser, M. Pollen flora of Pakistan–XXXI Betulaceae. Pak. J. Bot. 31, 243–246 (1999).
Raunkiaer, C., The life forms of plants and statistical plant geography; being the collected papers of C. Raunkiaer. The life forms of plants and statistical plant geography; being the collected papers of C. Raunkiaer. (1934).
Hussain, S.S., Pakistan manual of plant ecology1984: National Book Foundation.
Kamran, S. et al. The role of graveyards in species conservation and beta diversity: A vegetation appraisal of sacred habitats from Bannu Pakistan. J. For. Res. 31(4), 1147–1158 (2020).
Manan, F. et al. Environmental determinants of plant associations and evaluation of the conservation status of Parrotiopsis jacquemontiana in Dir, the Hindu Kush Range of Mountains. Trop. Ecol. 61(4), 509–526 (2020).
Tfaily, M. M. et al. Sequential extraction protocol for organic matter from soils and sediments using high resolution mass spectrometry. Anal. Chim. Acta 972, 54–61 (2017).
Chaney, R., Slonim, S., & Slonim, S. Determination of calcium carbonate content in soils, in Geotechnical properties, behavior, and performance of calcareous soils1982, ASTM International.
McCune, B., & Mefford, M. PC-ORD, Multivariate analysis of ecological data, Version 5 for Windows edition. MjM Software Design, Gleneden Beach, Oregon USA (2005).
Lepš, J., & Šmilauer, P. Multivariate analysis of ecological data using CANOCO2003: Cambridge university press.
Xie, W. et al. A novel hybrid method for landslide susceptibility mapping-based geodetector and machine learning cluster: A case of Xiaojin county, China. ISPRS Int. J. Geo Inf. 10(2), 93 (2021).
Li, L., Lei, Y. & Pan, D. Economic and environmental evaluation of coal production in China and policy implications. Nat. Hazards 77(2), 1125–1141 (2015).
Team, R.C., R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. http://www.R-project.org/ (2013).
Anwar, S. et al. Floristic composition and ecological gradient analyses of the Liakot Forests in the Kalam region of District Swat Pakistan. J. For. Res. 30(4), 1407–1416 (2019).
Haq, S. M. et al. Exploring and understanding the floristic richness, life-form, leaf-size spectra and phenology of plants in protected forests: A case study of Dachigam National Park in Himalaya Asia. Acta Ecol. Sin. 41(5), 479–490 (2021).
Ilyas, M. et al. A Preliminary checklist of the vascular flora of Kabal Valley, Swat Pakistan. Pak. J. Bot 45(2), 605–615 (2013).
Amjad, M. S. et al. Floristic composition, biological spectrum and phenological pattern of vegetation in the subtropical forest of Kotli District, AJK Pakistan. Pure Appl. Biol. (PAB) 6(2), 426–447 (2017).
Shaheen, H. et al. Species diversity, community structure, and distribution patterns in western Himalayan alpine pastures of Kashmir Pakistan. Mount. Res. Dev. 31(2), 153–159 (2011).
Abbas, Z. et al. Ethnobotany of the balti community, tormik valley, karakorum range, baltistan, pakistan. J. Ethnobiol. Ethnomed. 12(1), 1–16 (2016).
Ahmed, M. et al. Phytosociology and structure of Himalayan forests from different climatic zones of Pakistan. Pak. J. Bot. 38(2), 361 (2006).
Shehzadi, S. et al. Floristic compositions along an 18-Km long transect in Ayubia National Park District Abbottabad Pakistan. Pak. J. Bot. 41(5), 2115–2127 (2009).
Khan, W., et al., Life forms, leaf size spectra and diversity indices of plant species grown in the Thandiani forests, district Abbottabad, Khyber Pakhtunkhwa, Pakistan. Saudi J. Biol. Sci.
Kharkwal, G. et al. Phytodiversity and growth form in relation to altitudinal gradient in the Central Himalayan (Kumaun) region of India. Curr. Sci. 1, 873–878 (2005).
Bennie, J. et al. Slope, aspect and climate: Spatially explicit and implicit models of topographic microclimate in chalk grassland. Ecol. Model. 216(1), 47–59 (2008).
Choudhary, K. & Nama, K. S. Phyto-diversity of Mukundara hills national park of Kota district, Rajasthan India. Adv. Appl. Sci. Res. 5(1), 18–23 (2014).
Shimwell, D.W., Description and classification of vegetation (1971).
Malik, Z.H., Comparative study of vegetation of GungaChotti and Bedori Hills, Distric Bagh, Azad Jammu and Kashmir with special reference to range conditions, 2005, University of Peshawar, Pakistan.
Khan, W. et al. Life forms, leaf size spectra, regeneration capacity and diversity of plant species grown in the Thandiani forests, district Abbottabad, Khyber Pakhtunkhwa Pakistan. Saudi J. Biol. Sci. 25(1), 94–100 (2018).
Grytnes, J. A. & Vetaas, O. R. Species richness and altitude: A comparison between null models and interpolated plant species richness along the Himalayan altitudinal gradient Nepal. Am. Nat. 159(3), 294–304 (2002).
Majid, A., Khan, M. & Calixto, E. Ecological assessment of plant communities along the edaphic and topographic gradients of biha valley, District Swat Pakistan. Appl. Ecol. Environ. Res. 16(5), 5611–5631 (2018).
Khan, S.M., et al., Vegetation dynamics in the Western Himalayas, diversity indices and climate change. Sci. Tech. Dev. 31(3), 232–243 (2012).
Khan, S. M. et al. Identifying plant species and communities across environmental gradients in the Western Himalayas: Method development and conservation use. Eco. Inform. 14, 99–103 (2013).
Shaheen, H. & Shinwari, Z. K. Phyto diversity and endemic richness of Karambar lake vegetation from Chitral Hindukush-Himalayas. Pak. J. Bot 44(1), 17–21 (2012).
Wana, D., Plant communities and diversity along altitudinal gradients from Lake Abaya to Chencha Highlands, 2002, MA Thesis, School of Graduate Studies, Addis Ababa University. Addis Ababa.
Canfora, L. et al. Is soil microbial diversity affected by soil and groundwater salinity? Evidences from a coastal system in central Italy. Environ. Monit. Assess. 189(7), 1–15 (2017).
Liu, S., et al., The distribution characteristics and human health risks of high-fluorine groundwater in coastal plain: A case study in Southern Laizhou Bay, China. Front. Environ. Sci. 568 (2022).
Niu, Y. et al. Vegetation distribution along mountain environmental gradient predicts shifts in plant community response to climate change in alpine meadow on the Tibetan Plateau. Sci. Total Environ. 650, 505–514 (2019).
Nadal-Romero, E. et al. Effects of slope angle and aspect on plant cover and species richness in a humid Mediterranean badland. Earth Surf. Proc. Land. 39(13), 1705–1716 (2014).
Acknowledgements
We would like to thank King Abdullah University of Science and Technology (KAUST) for financial support in the process of publications. URF QAU is acknowledged for support in the experimental part. National Center for Physics (NCP), QAU and especially, Umar Nawaz is acknowledged for helping in some of the soil analyses.
Author information
Authors and Affiliations
Contributions
All authors contributed for the study conceptualization and design. H.U. performed conceptualization, field data, data analysis and wrote the first draft of the manuscript and makes changes accordingly. S.M.K. performed in conceptualization, interpretation, overall supervision, and finalization of the draft. Z.U. helps in the identification of plants specimens with the help of Flora of Pakistan and world floras. Z.A. performed the analysis of data through CANOCO, PCORD and R-software and designing methodology. S.J. performed in revision, editing and data curation. M.J. and I.A. help in the financial support for paying the APC fee, while H.B. help in data collection. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ullah, H., Khan, S.M., jaremko, M. et al. Vegetation assessments under the influence of environmental variables from the Yakhtangay Hill of the Hindu-Himalayan range, North Western Pakistan. Sci Rep 12, 20973 (2022). https://doi.org/10.1038/s41598-022-21097-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-21097-4
- Springer Nature Limited
This article is cited by
-
Eco-systematic assessment of the spring herbaceous vegetation under edaphic and topographic effects
Scientific Reports (2024)
-
Multivariate analysis of the summer herbaceous vegetation and environmental factors of the sub-tropical region
Scientific Reports (2024)
-
Similarity between soil seed bank and standing vegetation and their relationship with soil and topographical characteristics in a riparian zone
Community Ecology (2024)
-
Vegetation diversity pattern during spring season in relation to topographic and edaphic variables in sub-tropical zone
Botanical Studies (2023)