
Abstract
Pregnancy represents a crucial period in which several exposures—and especially maternal diet—might shape children’s health. Thus, identifying how maternal dietary intakes early affect biological aging in children represents a public health mission. We aimed to assess the relationship between maternal intake of nutrients in early pregnancy and telomere length of cell-free circulating DNA (cfDNA) from amniotic fluid. We used data and samples from the ongoing prospective “Mamma & Bambino” study, which recruits mother–child pairs from Catania at the first prenatal visit. Maternal nutrient intakes were assessed using a Food Frequency Questionnaire, while relative telomere length of cfDNA was assessed by real-time polymerase chain reaction. Our analysis included 174 mother–child pairs. The intakes of iron, vitamin B1, and magnesium were positively correlated with relative telomere length (p-values < 0.05). However, only the intake of magnesium was positively associated with relative telomere length, after applying a linear regression model (β = 0.002; SE = 0.001; p = 0.024). Magnesium deficiency was negatively associated with relative telomere length after adjusting for the same covariates (β = −0.467; SE = 0.176; p = 0.009). To our knowledge, this is the first evidence of a positive relationship between maternal nutrient intake and telomere length of cfDNA. Further efforts are needed for deeply investigating the effect of maternal dietary intakes on telomere length, in order to develop effective public health strategies.
Similar content being viewed by others


Introduction
Human aging is defined as the dynamic process characterized by the recurrent adaptation to internal and external stressors during lifetime1, which results in a complex mosaic of the interaction between environmental, genetic and epigenetic events2,3. Despite this complexity, several molecular signatures have been proposed to reflect the aging process, also in the context of studying the risk for non-communicable diseases4. Among them, the following hallmarks of aging are well investigated: genomic instability5,6,7,8, telomere attrition9,10, epigenetic alterations11,12,13,14,15,16,17, mitochondrial dysfunction18,19, cellular senescence20,21,22,23, steam cell exhaustion24 and altered intercellular communication25,26. In vertebrates, telomeres are repetitive sequence of 5′-(TTAGGG)-3′ at the ends of each chromosome, which progressively shorten with cell division giving a partial estimation of the chronological age throughout lifetime27. In particular, telomere shortening has been associated with aging28 and age-related diseases, such as cardiovascular diseases29, cancer30, and neurological disorders31. The contemporary relationship of telomere length with environmental exposures and lifestyles have made it as an interesting molecular mechanisms in the epidemiological research on age-related diseases32.
It is also worth noting that telomere shortening is regulated by epigenetic mechanisms and that telomere length is influenced by DNA methyltransferases and histone methyltransferases33,34.
In the last decades, as done for DNA methylation and histone modification35,36,37,38, several studies have investigated the association of nutrients, foods, and dietary patterns with telomere length. As summarized by Freitas-Simoes and colleagues, the intake of antioxidants and the consumption of plant-derived foods help protect against telomere shortening, while the intake of saturated fats and the consumption of high-sugar and high-calorie products seem to be associated with shorter telomere length39. It is also important to note that understanding what maternal factors—and especially maternal dietary habits—might affect biological aging in children could be useful to identify simple strategies for preventing or delaying age-related diseases over the lifetime. In fact, the first 1000 days of life—from conception to two years of age—represents a crucial period in which maternal diet and other exposures might shape children’s health. For instance, some maternal factors, such as stress, smoking and exposure to pollutants, have been associated with shorter telomeres in cord blood40,41,42 and placenta43. In addition, a recent systematic review by Habibi and colleagues has sought to unravel how the diet of pregnant women affects telomere length in their offspring44. Although some studies reported potential positive (e.g., folate and caffeine) or negative (e.g., fat and sodium) associations with telomere length, the authors stated that the evidence is currently limited and often controversial44. Moreover, none of these studies has investigated the effect of maternal dietary factors on telomere length of cell-free circulating DNA (cfDNA) from amniotic fluid44. In particular, cfDNA in plasma and serum has been proposed as an innovative biomarker for prenatal diagnosis45,46. However, since amniotic fluid contains a greater amount of cfDNA than maternal serum47,48,49,50, there is a growing interest in investigating the relationship between maternal diet and telomere length of cfDNA from amniotic fluid. To achieve this goal, we used data and samples from the “Mamma & Bambino” cohort to evaluate the relationship between maternal intake of nutrients in early pregnancy and telomere length of cfDNA from amniotic fluid.
Results
Characteristics of the study population
The current analysis was conducted on 174 women recruited at a median gestational week of 14 (IQR = 5) and with a median age of 38 years (IQR = 4). Table 1 summarizes the characteristics of the study population. The majority of women (83.3%) reported a high educational level, having a secondary or tertiary education. More than half of them (54.0%), instead, were part-time or full-time employed. The median pre-gestational BMI was 22.8 kg/m2 (IQR = 4.6) and thus 21.3% women were overweight or obese before pregnancy. With respect to smoking status, nearly 80% of women were non-current smokers. Table 1 also reports the median intake of nutrients considered in the current analysis. Overall, women had a total energy intake of 1662 kcal (IQR = 634) and their dietary deficiencies varied from 17.8% for vitamin A to 98.3% for iron. About one in three women (35%) took multivitamin or multimineral supplements.
The relationship between relative telomere length and nutrient intakes
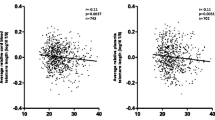
Overall, median relative telomere length was 0.73 (IQR = 1), while no differences were evident by maternal age, educational level, employment status, pre-gestational BMI, and smoking status (p-values > 0.05). Figure 1 reports the Spearman’s rank correlation coefficients between relative telomere length and nutrient intakes. Most of the relationships were not significant and only the intakes of magnesium, vitamin B1 and iron were positively but weakly correlated with relative telomere length (p-values < 0.05). However, only the correlation with magnesium remained significant after adjusting for multiple comparison. Figure 2 shows positive linear relationships of the intakes of magnesium, vitamin B1, and iron with relative telomere length. However, only the intake of magnesium was positively associated with relative telomere length (β = 0.002; SE = 0.001; p = 0.024), after applying a linear regression model which included maternal age, smoking, pre-gestational BMI, total energy intake, and supplement use. Figure 3 illustrates differences in relative telomere length according to nutrient deficiency. In particular, women with magnesium deficiency (73% of the total population) exhibited lower relative telomere length than those with adequate dietary intake (p = 0.005). Similarly, women with vitamin B1 deficiency (53% of the total population) showed lower values than those with adequate intake (p = 0.040). By contrast, no significant difference was evident according to iron deficiency (p = 0.240). After adjusting for covariates, however, only magnesium deficiency was negatively associated with relative telomere length (β = −0.467; SE = 0.176; p = 0.009).

Correlations of nutrient intakes with relative telomere length.

The relationships between relative telomere length and the intake of magnesium (A), vitamin B1 (B), and iron (C). Results are reported as β coefficient and SE.

Differences in relative telomere length according to deficiency of magnesium (A), vitamin B1 (B), and iron (C). P-values are based on the Mann Whitney U test.
Discussion
To our knowledge, this is the first evidence of a relationship between maternal nutrient intake and telomere length of cfDNA from amniotic fluid. In particular, we found positive but weak correlations of intakes of magnesium, vitamin B1 and iron with relative telomere length. While findings on vitamin B1 and iron were not confirmed by further statistical analyses, we demonstrated a positive association between magnesium intake and relative telomere length. The positive effect observed for magnesium remained significant after adjusting for covariates, which hence resulted in shorter telomeres in cfDNA from women with magnesium deficiency. It is worth noting, however, that there is a lack of studies investigating the relationship between magnesium intake and telomere length of cfDNA from amniotic fluid. Although some of our findings were consistent with previous studies, it is important to underline differences in sample types and methods used to perform molecular analyses, which discourage a proper comparison. In vitro and in vivo studies showed that long-term exposure to magnesium deficiency led to telomere shortening51,52. In a cross-sectional analysis of the Sister Study, instead, reported a positive association between magnesium intake and telomere length of leukocyte DNA from women who did not use multivitamin supplements53. From a biological point of view, magnesium is an important cofactor for the catalytic activity of enzymes implicated in DNA replication and repair54,55,56,57, and in RNA synthesis54. Magnesium deficiency is also often associated with oxidative stress51 and pro-inflammatory status58, which in turn might lead to telomere shortening.
As discussed above, the evidence remained scarce and inconclusive for other nutrients. In fact, our analysis suggested positive but weak correlations between relative telomere length of cfDNA and the intakes of vitamin B1 and iron. A previous cross-sectional study on Italian subjects failed in demonstrating a relationship between telomere length of leukocyte DNA and the intake of vitamin B159. Moreover, to the best of our knowledge, no studies were conducted during the periconceptional period. Regarding iron, results from the Sister Study did not demonstrate an association between its dietary intake and telomere length53, however, it has been suggested a negative effect of iron supplementation53,60. Indeed, iron is a prooxidant and its supplementation might increase the production of free radicals61, thus fostering an oxidative environment60. By contrast, the supplementations with multivitamins, which contained less iron than specific supplements, did not produce a negative effect53. The lack of any solid evidence in this field of research, therefore, encourages further efforts to understand the influence of maternal dietary factors on biological aging, as determined by telomere length. Furthermore, future studies investigating other biomarkers of aging could help resolve this question and to translate the answers into effective public health strategies.
The main strength of our study lay on the possibility of investigating the early effect of maternal nutrient intake on biological aging of their offspring, by analysing telomere length of cfDNA from amniotic fluid. In fact, compared to maternal serum, amniotic fluid contains cfDNA largely uncontaminated by maternal-derived nucleic acids. However, as the first study on this topic, our results should be confirmed by further analyses. Our work had also some limitations that should be considered when interpreting our results. Firstly, the limited sample size did not allow us to perform additional analyses for residual confounders. However, as showed by Habibi and colleagues, our study was, at the time of writing, one of the larger in its field. Secondly, the FFQ that we used for dietary assessment—although simple, time- and cost-efficient—did not preclude subjective assessment, measurement errors and inaccuracies62. Moreover, this tool did not consider any changes related to food cooking. To partially address these issues, the use of some biomarkers of validation (e.g., serum level of nutrients) could have made the evidence more solid. Thirdly, differences in analysed samples, methods, and kits used for DNA extraction and telomere length assessment did not allow a proper comparison with previous studies. Although amniotic fluid is considered a relatively pure fetal sample, a low proportion of cfDNA from placenta cannot be completely excluded63. Indeed, placenta could contribute to cfDNA in amniotic fluid, especially in women with placental abnormalities and preeclampsia. Accordingly, we excluded from our analysis all women with pregnancy complications. cfDNA is also inherently degraded and therefore DNA strands are highly fragmented if compared with cellular DNA from other samples. Moreover, the extraction kit used in our study was not specifically developed for cfDNA from amniotic fluid, even if it was appropriate for body fluids in general. Regarding telomere length assessment, the qPCR used in our study had higher assay variability than terminal restriction fragment analysis used by previous studies64. For these reasons, further validation analyses and comparative studies—using different samples, as well as alternative methods for DNA extraction and telomere length assessment—should be encouraged.
Finally, although we adjusted the analysis for some factors that could influence the observed relationship, we cannot completely exclude the effect of unmeasured confounders.
In conclusion, we found a positive association between maternal intake of magnesium and telomere length of cfDNA from amniotic fluid, while results on other micronutrients (i.e., vitamin B1 and iron) were marginally significant. This is the first evidence of an early effect of maternal magnesium intake on biological aging of offspring. Although our approach could help to understand molecular mechanisms underpinning the transgenerational effects of maternal diet on biological aging, further research is needed to identify strategies for preventing or delaying age-related diseases as early as pregnancy.
Methods
Study design
The “Mamma & Bambino” cohort is a prospective study that recruits pregnant women during the prenatal genetic counseling (at 4th–20th gestational week) with planned follow-up of their children at delivery and up to two years of age. Full details of the study design and procedures are described elsewhere65,66,67,68,69,70. In brief, the recruitment of women referring to the Azienda Ospedaliero Universitaria Policlinico “G. Rodolico—San Marco” (Catania, Italy) started in 2015 and the study is still ongoing. Mothers with multiple pregnancy, pre-existing autoimmune and/or chronic diseases, and complications, such as preeclampsia, gestational hypertension and diabetes, intrauterine fetal death, and congenital malformations, are excluded. Accordingly, the current analysis was conducted on 174 women (aged 15 to 48 years), who satisfied inclusion criteria reported above. The study is conducted in accordance with the Declaration of Helsinki and its protocol was approved by Ethics Committee of the “Azienda Ospedaliero Universitaria Policlinico-Vittorio Emanuele” and by the Ethics Committee “Catania 1” with the protocol numbers 47/2014/VE, 48/2015/EMPO, 186/2015/EMPO, 197/2016/EMPO, 213/2017/EMPO, 231/2018/EMPO, and 263/2019/EMPO. All participants or their legal guardian give a written informed consent to participate in the study. For the current analysis, we used data and samples from women who completed pregnancy and who provided an aliquot of amniotic fluid obtained through amniocentesis71.
Data collection
For each woman, information on socio-demographic characteristics and lifestyles are collected through structured questionnaires administered by trained epidemiologists. Full details on data collection and management are reported elsewhere65,66,67,68,69,70,71. In particular, dietary data are collected through a 95-item semiquantitative Food Frequency Questionnaire (FFQ) referred to one month before recruitment, as reported by previous studies on Sicilian women66,67,72,73,74,75,76,77,78. For each item, information on frequency of consumption and portions size are collected to calculate their daily dietary intake. Next, the intakes of calories, minerals (iron, calcium, magnesium, and zinc), fatty acids (saturated, monounsaturated, and polyunsaturated), and vitamins (A, B1, B6, C, D, and folate) are computed using the U.S. Department of Agriculture (USDA) Food Composition Database (http://ndb.nal.usda.gov/) adapted to typical Italian foods. Nutrient intakes are considered as continuous values or categorized according to the Recommended Dietary Allowance by the Food and Nutrition Board of the Institute of Medicine79. At recruitment, each woman reported her height and weight before pregnancy to compute the pre-pregnancy BMI, and hence all women were classified as underweight, normal weight, overweight or obese according to WHO criteria80.
DNA extraction and relative telomere length assessment
Different samples types—including maternal blood, amniotic fluid, cord blood and placenta—are collected from mother–child pairs included in the Mamma & Bambino cohort. In the current analysis, we used an aliquot of 1 ml amniotic fluid obtained from women who underwent amniocentesis, as previously reported71. In brief, after centrifugation at 12,500 g, the cfDNA was extracted using the QIAamp Blood Kit (Qiagen, Milan Italy), which is suitable to extract DNA from blood, plasma and serum, cultured cells, swabs, and body fluids. The entire procedure was automatically performed on the QIAcube instrument (Qiagen, Milan, Italy). Quantity and quality of cfDNA were assessed using the dsDNA HS Assay Kit (Thermo Fisher Scientific, Carlsbad, CA, USA) on the Qubit 3.0 Fluorometer and the NanoDrop 1000 spectrometer. Next, relative telomere length of cfDNA was evaluated real-time quantitative polymerase chain reaction (qPCR), using the Relative Human Telomere Length Quantification Assay Kit (ScienCell Research Laboratories, Carlsbad, CA, USA) on the QuantStudio 7 Flex Real-Time PCR System (Thermo Fisher Scientific, Carlsbad, CA, USA). Full details on the qPCR protocol are described elsewhere81. All reactions were run in duplicate and relative telomere length was expressed as telomere/single copy reference (T/S) ratio. The procedures described above were conducted according to the manufacturers' protocols, unless otherwise stated.
Statistical analysis
The characteristics of the study population were described using frequencies (percentage, %) for qualitative variables, and using median and interquartile range (IQR) due to the skewness of quantitative variables. The correlations between nutrient intakes and relative telomere length were tested using the Spearman's rank correlation test and adjusting for multiple comparisons with Bonferroni correction. For nutrients that showed a significant correlation (p < 0.05), we plotted their continuous value against relative telomere length, and then we compared relative telomere length between deficient and not deficient women using the Mann Whitney U test. Finally, we tested the association between nutrient intake (as continuous or categorical variable) and relative telomere length, adjusting for factors that could influence nutrient intake and/or telomere length (i.e., maternal age, smoking status, pregestational BMI, total daily energy intake, and supplement use). The adjusted association was reported as β coefficient and its standard error (SE). All the statistical analyses were performed using SPSS (version 26), all tests were two-sided and performed at a significance level α = 0.05.
Institutional review board statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of Azienda Ospedaliero-Universitaria Policlinico-Vittorio Emanuele” and Ethics Committee “Catania 1″ with the following protocol numbers: 47/2014/VE; 48/2015/EMPO; 186/2015/EMPO; 197/2016/EMPO; 213/2017/EMPO; 231/2018/EMPO; 263/2019/EMPO.
Informed consent
Informed consent was obtained from all subjects involved in the study.
Data availability
The datasets analyzed during the current study are available from the corresponding author on reasonable request.
References
Franceschi, C. et al. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 908, 244–254. https://doi.org/10.1111/j.1749-6632.2000.tb06651.x (2000).
Cevenini, E. et al. Human models of aging and longevity. Expert. Opin. Biol. Ther. 8, 1393–1405. https://doi.org/10.1517/14712598.8.9.1393 (2008).
Colloca, G., Santoro, M. & Gambassi, G. Age-related physiologic changes and perioperative management of elderly patients. Surg. Oncol. 19, 124–130. https://doi.org/10.1016/j.suronc.2009.11.011 (2010).
Xia, X., Chen, W., McDermott, J. & Han, J. J. Molecular and phenotypic biomarkers of aging. F1000Res 6, 860 (2017).
Zhang, L. et al. Single-cell whole-genome sequencing reveals the functional landscape of somatic mutations in B lymphocytes across the human lifespan. Proc. Natl. Acad. Sci. USA 116, 9014–9019. https://doi.org/10.1073/pnas.1902510116 (2019).
Franco, I. et al. Somatic mutagenesis in satellite cells associates with human skeletal muscle aging. Nat. Commun. 9, 800. https://doi.org/10.1038/s41467-018-03244-6 (2018).
Bae, T. et al. Different mutational rates and mechanisms in human cells at pregastrulation and neurogenesis. Science 359, 550–555. https://doi.org/10.1126/science.aan8690 (2018).
Lodato, M. A. et al. Aging and neurodegeneration are associated with increased mutations in single human neurons. Science 359, 555–559. https://doi.org/10.1126/science.aao4426 (2018).
Greider, C. W. Telomerase discovery: the excitement of putting together pieces of the puzzle (Nobel lecture). Angew Chem. Int. Ed. Engl. 49, 7422–7439. https://doi.org/10.1002/anie.201002408 (2010).
Vera, E., Bernardes de Jesus, B., Foronda, M., Flores, J. M. & Blasco, M. A. The rate of increase of short telomeres predicts longevity in mammals. Cell Rep. 2, 732–737. https://doi.org/10.1016/j.celrep.2012.08.023 (2012).
Hannum, G. et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 49, 359–367. https://doi.org/10.1016/j.molcel.2012.10.016 (2013).
Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 14, R115. https://doi.org/10.1186/gb-2013-14-10-r115 (2013).
Gensous, N. et al. The epigenetic landscape of age-related diseases: the geroscience perspective. Biogerontology 18, 549–559. https://doi.org/10.1007/s10522-017-9695-7 (2017).
Levine, M. E. et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging (Albany NY) 10, 573–591 (2018).
Chen, B. H. et al. DNA methylation-based measures of biological age: meta-analysis predicting time to death. Aging (Albany NY) 8, 1844–1865 (2016).
Degerman, S. et al. Maintained memory in aging is associated with young epigenetic age. Neurobiol. Aging 55, 167–171. https://doi.org/10.1016/j.neurobiolaging.2017.02.009 (2017).
Gale, C. R. et al. The epigenetic clock and objectively measured sedentary and walking behavior in older adults: the Lothian Birth Cohort 1936. Clin. Epigenet. 10, 4. https://doi.org/10.1186/s13148-017-0438-z (2018).
Zane, A. C. et al. Muscle strength mediates the relationship between mitochondrial energetics and walking performance. Aging Cell 16, 461–468. https://doi.org/10.1111/acel.12568 (2017).
Dikalov, S. I. & Harrison, D. G. Methods for detection of mitochondrial and cellular reactive oxygen species. Antioxid Redox Signal 20, 372–382. https://doi.org/10.1089/ars.2012.4886 (2014).
Childs, B. G. et al. Senescent cells: An emerging target for diseases of ageing. Nat. Rev. Drug Discov. 16, 718–735. https://doi.org/10.1038/nrd.2017.116 (2017).
Andriani, G. A. et al. Whole Chromosome Instability induces senescence and promotes SASP. Sci. Rep. 6, 35218. https://doi.org/10.1038/srep35218 (2016).
Baker, D. J. & Petersen, R. C. Cellular senescence in brain aging and neurodegenerative diseases: Evidence and perspectives. J. Clin. Invest. 128, 1208–1216. https://doi.org/10.1172/JCI95145 (2018).
Tanaka, T. et al. Plasma proteomic signature of age in healthy humans. Aging Cell 17, e12799. https://doi.org/10.1111/acel.12799 (2018).
Ren, R., Ocampo, A., Liu, G. H. & Izpisua Belmonte, J. C. Regulation of stem cell aging by metabolism and epigenetics. Cell Metab 26, 460–474. https://doi.org/10.1016/j.cmet.2017.07.019 (2017).
Bektas, A., Schurman, S. H., Sen, R. & Ferrucci, L. Aging, inflammation and the environment. Exp. Gerontol. 105, 10–18. https://doi.org/10.1016/j.exger.2017.12.015 (2018).
Fabbri, E. et al. Aging and the burden of multimorbidity: Associations with inflammatory and anabolic hormonal biomarkers. J. Gerontol. A Biol. Sci. Med. Sci. 70, 63–70. https://doi.org/10.1093/gerona/glu127 (2015).
Blackburn, E. H., Epel, E. S. & Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 350, 1193–1198. https://doi.org/10.1126/science.aab3389 (2015).
Kimura, M. et al. Telomere length and mortality: A study of leukocytes in elderly Danish twins. Am. J. Epidemiol. 167, 799–806. https://doi.org/10.1093/aje/kwm380 (2008).
Njajou, O. T. et al. Shorter telomeres are associated with obesity and weight gain in the elderly. Int. J. Obes. (Lond.) 36, 1176–1179. https://doi.org/10.1038/ijo.2011.196 (2012).
Mundstock, E. et al. Effect of obesity on telomere length: Systematic review and meta-analysis. Obesity (Silver Spring) 23, 2165–2174. https://doi.org/10.1002/oby.21183 (2015).
Eitan, E., Hutchison, E. R. & Mattson, M. P. Telomere shortening in neurological disorders: an abundance of unanswered questions. Trends Neurosci. 37, 256–263. https://doi.org/10.1016/j.tins.2014.02.010 (2014).
Gorenjak, V., Akbar, S., Stathopoulou, M. G. & Visvikis-Siest, S. The future of telomere length in personalized medicine. Front. Biosci. (Landmark Ed) 23, 1628–1654. https://doi.org/10.2741/4664 (2018).
García-Cao, M., O’Sullivan, R., Peters, A. H., Jenuwein, T. & Blasco, M. A. Epigenetic regulation of telomere length in mammalian cells by the Suv39h1 and Suv39h2 histone methyltransferases. Nat. Genet. 36, 94–99. https://doi.org/10.1038/ng1278 (2004).
Gonzalo, S. et al. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat. Cell Biol. 8, 416–424. https://doi.org/10.1038/ncb1386 (2006).
Maugeri, A. & Barchitta, M. How dietary factors affect DNA methylation: Lesson from epidemiological studies. Medicina 56, 1. https://doi.org/10.3390/medicina56080374 (2020).
Zhang, Y. & Kutateladze, T. G. Diet and the epigenome. Nat. Commun. 9, 3375. https://doi.org/10.1038/s41467-018-05778-1 (2018).
Kadayifci, F. Z., Zheng, S. & Pan, Y. X. Molecular Mechanisms Underlying the Link between Diet and DNA Methylation. Int. J. Mol. Sci. 19, 1. https://doi.org/10.3390/ijms19124055 (2018).
Choi, S. W., Claycombe, K. J., Martinez, J. A., Friso, S. & Schalinske, K. L. Nutritional epigenomics: A portal to disease prevention. Adv. Nutr. 4, 530–532. https://doi.org/10.3945/an.113.004168 (2013).
Freitas-Simoes, T. M., Ros, E. & Sala-Vila, A. Nutrients, foods, dietary patterns and telomere length: Update of epidemiological studies and randomized trials. Metabolism 65, 406–415. https://doi.org/10.1016/j.metabol.2015.11.004 (2016).
Gorenjak, V., Petrelis, A. M., Stathopoulou, M. G. & Visvikis-Siest, S. Telomere length determinants in childhood. Clin. Chem. Lab. Med. 58, 162–177. https://doi.org/10.1515/cclm-2019-0235 (2020).
Liu, B. et al. Prenatal second-hand smoke exposure and newborn telomere length. Pediatr. Res. 87, 1081–1085. https://doi.org/10.1038/s41390-019-0594-2 (2020).
Salihu, H. M. et al. Association between maternal-perceived psychological stress and fetal telomere length. South Med. J. 109, 767–772 (2016).
Martens, D. S. et al. Prenatal air pollution and newborns’ predisposition to accelerated biological aging. JAMA Pediatr. 171, 1160–1167. https://doi.org/10.1001/jamapediatrics.2017.3024 (2017).
Habibi, N. et al. Maternal diet and offspring telomere length: a systematic review. Nutr. Rev. 79, 148–159. https://doi.org/10.1093/nutrit/nuaa097 (2021).
Bianchi, D. W. Circulating fetal DNA: its origin and diagnostic potential—A review. Placenta 25(Suppl A), S93–S101. https://doi.org/10.1016/j.placenta.2004.01.005 (2004).
Lo, Y. M. Recent advances in fetal nucleic acids in maternal plasma. J. Histochem. Cytochem. 53, 293–296. https://doi.org/10.1369/jhc.4R6362.2005 (2005).
Grisaru-Granovsky, S. et al. Population-based trends in mortality and neonatal morbidities among singleton, very preterm, very low birth weight infants over 16 years. Early Hum. Dev. 90, 821–827. https://doi.org/10.1016/j.earlhumdev.2014.08.009 (2014).
Hug, L., Alexander, M., You, D., Alkema, L. & Estimation, U. I.-a. G. f. C. M. National, regional, and global levels and trends in neonatal mortality between 1990 and 2017, with scenario-based projections to 2030: a systematic analysis. Lancet Glob. Health 7, e710-e720 (2019). https://doi.org/10.1016/S2214-109X(19)30163-9
Moutquin, J. M. Classification and heterogeneity of preterm birth. BJOG 110(Suppl 20), 30–33 (2003).
Goldenberg, R. L., Culhane, J. F., Iams, J. D. & Romero, R. Epidemiology and causes of preterm birth. Lancet 371, 75–84. https://doi.org/10.1016/S0140-6736(08)60074-4 (2008).
Martin, H. et al. Effects of long-term dietary intake of magnesium on oxidative stress, apoptosis and ageing in rat liver. Magnes Res. 21, 124–130 (2008).
Killilea, D. W. & Ames, B. N. Magnesium deficiency accelerates cellular senescence in cultured human fibroblasts. Proc. Natl. Acad. Sci. USA 105, 5768–5773. https://doi.org/10.1073/pnas.0712401105 (2008).
Xu, Q. et al. Multivitamin use and telomere length in women. Am. J. Clin. Nutr. 89, 1857–1863. https://doi.org/10.3945/ajcn.2008.26986 (2009).
Hartwig, A. Role of magnesium in genomic stability. Mutat. Res. 475, 113–121. https://doi.org/10.1016/s0027-5107(01)00074-4 (2001).
Batra, V. K. et al. Magnesium-induced assembly of a complete DNA polymerase catalytic complex. Structure 14, 757–766. https://doi.org/10.1016/j.str.2006.01.011 (2006).
Sirover, M. A. & Loeb, L. A. Metal activation of DNA synthesis. Biochem. Biophys. Res. Commun. 70, 812–817. https://doi.org/10.1016/0006-291x(76)90664-1 (1976).
Mazia, D. The particulate organization of the chromosome. Proc. Natl. Acad. Sci. USA 40, 521–527. https://doi.org/10.1073/pnas.40.6.521 (1954).
Guerrero-Romero, F. & Rodríguez-Morán, M. Relationship between serum magnesium levels and C-reactive protein concentration, in non-diabetic, non-hypertensive obese subjects. Int. J. Obes. Relat. Metab. Disord. 26, 469–474. https://doi.org/10.1038/sj.ijo.0801954 (2002).
Marcon, F. et al. Diet-related telomere shortening and chromosome stability. Mutagenesis 27, 49–57. https://doi.org/10.1093/mutage/ger056 (2012).
Aviv, A. Leukocyte telomere length: The telomere tale continues. Am. J. Clin. Nutr. 89, 1721–1722. https://doi.org/10.3945/ajcn.2009.27807 (2009).
Lund, E. K., Wharf, S. G., Fairweather-Tait, S. J. & Johnson, I. T. Oral ferrous sulfate supplements increase the free radical-generating capacity of feces from healthy volunteers. Am. J. Clin. Nutr. 69, 250–255. https://doi.org/10.1093/ajcn/69.2.250 (1999).
Mozaffarian, D., Rosenberg, I. & Uauy, R. History of modern nutrition science-implications for current research, dietary guidelines, and food policy. BMJ 361, k2392. https://doi.org/10.1136/bmj.k2392 (2018).
Lun, F. M. et al. Epigenetic analysis of RASSF1A gene in cell-free DNA in amniotic fluid. Clin. Chem. 53, 796–798. https://doi.org/10.1373/clinchem.2006.084350 (2007).
Aviv, A. et al. Impartial comparative analysis of measurement of leukocyte telomere length/DNA content by Southern blots and qPCR. Nucl. Acids Res. 39, e134. https://doi.org/10.1093/nar/gkr634 (2011).
Barchitta, M. et al. Dietary folate intake and folic acid supplements among pregnant women from Southern Italy: Evidence from the “Mamma & Bambino” Cohort. Int. J. Environ. Res. Public Health 17, 1. https://doi.org/10.3390/ijerph17020638 (2020).
Maugeri, A. et al. The impact of social determinants and lifestyles on dietary patterns during pregnancy: Evidence from the “Mamma & Bambino” study. Ann. Ig 31, 1 (2019).
Maugeri, A. et al. Maternal dietary patterns are associated with pre-pregnancy body mass index and gestational weight gain: Results from the “Mamma & Bambino” cohort. Nutrients 11, 1. https://doi.org/10.3390/nu11061308 (2019).
Barchitta, M. et al. Single nucleotide polymorphisms in vitamin D receptor gene affect birth weight and the risk of preterm birth: Results from the “Mamma & Bambino” cohort and a meta-analysis. Nutrients 10, 1. https://doi.org/10.3390/nu10091172 (2018).
Barchitta, M. et al. Vaccination status of mothers and children from the ‘Mamma & Bambino’ cohort. Vaccines 9, 168 (2021).
Lio, M. S. et al. The impact of socio-demographic factors on breastfeeding: Findings from the “Mamma & Bambino” cohort. Medicina (Kaunas) 57, 1. https://doi.org/10.3390/medicina57020103 (2021).
Maugeri, A. et al. The effect of alcohol on telomere length: A systematic review of epidemiological evidence and a pilot study during pregnancy. Int. J. Environ. Res. Public Health 18, 1. https://doi.org/10.3390/ijerph18095038 (2021).
Maugeri, A. et al. Adherence to the Mediterranean diet partially mediates socioeconomic differences in leukocyte LINE-1 methylation: evidence from a cross-sectional study in Italian women. Sci. Rep. 10, 14360. https://doi.org/10.1038/s41598-020-71352-9 (2020).
Barchitta, M. et al. The association of dietary patterns with high-risk human papillomavirus infection and cervical cancer: A cross-sectional study in Italy. Nutrients 10, 1. https://doi.org/10.3390/nu10040469 (2018).
Maugeri, A. et al. Determinants of adherence to the mediterranean diet: Findings from a cross-sectional study in women from Southern Italy. Int. J. Environ. Res. Public Health 16, 1. https://doi.org/10.3390/ijerph16162963 (2019).
Barchitta, M. et al. Dietary antioxidant intake and human papillomavirus Infection: evidence from a cross-sectional study in Italy. Nutrients 12, 1. https://doi.org/10.3390/nu12051384 (2020).
Barchitta, M. et al. Dietary patterns are associated with leukocyte LINE-1 methylation in women: A cross-sectional study in Southern Italy. Nutrients 11, 1. https://doi.org/10.3390/nu11081843 (2019).
Agodi, A. et al. Low fruit consumption and folate deficiency are associated with LINE-1 hypomethylation in women of a cancer-free population. Genes Nutr. 10, 480. https://doi.org/10.1007/s12263-015-0480-4 (2015).
Barchitta, M. et al. Mediterranean Diet and Particulate Matter Exposure Are Associated With LINE-1 Methylation: Results From a Cross-Sectional Study in Women. Front. Genet. 9, 514. https://doi.org/10.3389/fgene.2018.00514 (2018).
Institute of Medicine Standing Committee on the Scientific Evaluation of Dietary Reference Intakes and its Panel on Folate, O. B. V. a. C. in Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline (National Academies Press (US) National Academy of Sciences., 1998).
Eveleth, P. B. et al. Uses and interpretation of anthropometry in the elderly for the assessment of physical status. Report to the Nutrition Unit of the World Health Organization: the Expert Subcommittee on the Use and Interpretation of Anthropometry in the Elderly. J Nutr Health Aging 2, 5–17 (1998).
Maugeri, A. et al. The relationship between telomere length and gestational weight gain: Findings from the Mamma & Bambino cohort. Biomedicines 10, 67 (2022).
Acknowledgements
We are grateful to all women who gave their consent to participate in the study.
Funding
The research “La coorte Mamma & Bambino: un approccio Multisettoriale Alla salute Materno-Infantile Mediante valutazione dell’Esposoma nelle Donne, MAMI-MED” was funded by the University of Catania, Italy, Department of Medical and Surgical Science and Advanced Technologies “GF Ingrassia” (Programma ricerca di ateneo UNICT 2020–22 linea 2, PIAno di inCEntivi per la RIcerca di Ateneo 2020/2022). This work was also supported by the “Fondi di Ateneo 2020-2022, Università di Catania, Linea Open Access”.
Author information
Authors and Affiliations
Contributions
Conceptualization, R.M.S.L., A.M., A.A. and M.B.; software, R.M.S.L. and A.M.; formal analysis, R.M.S.L., A.M., M.C.L.R., G.G., M.P., A.C., M.A.T.C. and M.B.; resources, A.A.; data curation, R.M.S.L., A.M., M.C.L.R. and M.B.; writing—original draft preparation, R.M.S.L. and A.M.; writing—review and editing, all the Authors; visualization, R.M.S.L. and A.M.; supervision, A.A. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Magnano San Lio, R., Maugeri, A., La Rosa, M.C. et al. Nutrient intakes and telomere length of cell-free circulating DNA from amniotic fluid: findings from the Mamma & Bambino cohort. Sci Rep 12, 11671 (2022). https://doi.org/10.1038/s41598-022-15370-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-15370-9
- Springer Nature Limited