
Abstract
Wild grapevine (Vitis vinifera subsp. sylvestris) is widely recognized as an important source of resistance or tolerance genes for diseases and environmental stresses. Recent studies revealed partial resistance to powdery mildew (Erysiphe necator, PM) in V. sylvestris from Central Asia. Here, we report resistance to PM of V. sylvestris collected from different regions of Croatia and in seedling populations established from in situ V. sylvestris accessions. Ninety-one in situ individuals and 67 V. sylvestris seedlings were evaluated for PM resistance according to OIV 455 descriptor. Three SSR markers (SC47-18, SC8-071-0014, and UDV-124) linked to PM resistance locus Ren1 were used to decipher allelic structure. Nine seedlings showed resistance in in vivo evaluations while leaf disk assays revealed three PM-resistant accessions. One V. vinifera cultivar used as a control for PM evaluations also showed high phenotypic resistance. Based on the presence of one or two resistance alleles that are linked to the Ren1 locus, 32 resistant seedlings and 41 resistant in situ genotypes were identified in the investigated set. Eight seedlings showed consistent phenotypic PM resistance, of which seven carried one or two alleles at the tested markers. This study provides the first evidence of PM resistance present within the eastern Adriatic V. sylvestris germplasm.
Similar content being viewed by others


Introduction
Powdery mildew (Erysiphe necator) is an economically important fungal disease of grapevine. It has been a continuous problem since its introduction from North America to Europe around 1845. In < 10 years, E. necator became a problem in vineyards throughout the Mediterranean1. Shortly after the onset of this disease in Europe, inorganic fungicides and sulfur were used to control powdery mildew (PM). There is a risk of the fungus developing resistance to fungicides2. Chemical protection is applied ~ 10 times during the growing season, making viticulture one of the largest fungicide consumers worldwide3. Fungicides can also cause undesirable characteristics, such as off-flavors in wine4. Modern researchers are developing revolutionary methods to control PM, including exposure of the fungus to UV light at night, when the fungal defense system ‘turns off’5. Another solution comes from the grapevine genome itself. Twelve loci from diverse grapevine species originating in North America, Central Asia and China carry genes for plant defense against PM6. Loci such as Ren4 and Ren6 (Resistance to Erysiphe necator) provide complete resistance, not allowing the fungus to proliferate7.
European grapevine (Vitis vinifera subsp. vinifera; hereafter called V. vinifera) is susceptible to powdery mildew, generally showing no resistance8. However, strong natural resistance to PM was first identified in the Central Asian V. vinifera cultivar (cv.) ‘Kishmish vatkana’9, which carries the major locus Ren1 on chromosome 13. The Ren1 locus has many advantages in breeding programs and mediates partial resistance to PM6, similar to loci Ren3 and Ren910.
Research on PM resistance was recently broadened by including the wild Eurasian grapevine (Vitis vinifera subsp. sylvestris Gmel Hegi; hereafter called V. sylvestris) into disease evaluations. V. sylvestris usually grows in habitats isolated from human impact. It is mostly susceptible to PM11,12, but considerably less affected by the disease than V. vinifera11,13 indicating some level of tolerance to PM11. V. sylvestris populations in northern Spain were susceptible to PM12, as were vines growing in six river basins in southern Spain14, with varying infection levels of different parasitic species in vines from the same location.
Recent identification of Ren1 in two V. sylvestris accessions from Central Asia revealed that PM resistance is located at the same genetic position on chromosome 13 as in cv. ’Kishmish vatkana’15. Additionally, a few dozen PM-resistant V. sylvestris accessions from Central Asia were identified as carriers of new resistance-linked alleles at five microsatellite (SSR) markers associated with the Ren1 locus7. The presence of Ren1 in V. sylvestris is very intriguing for the grapevine breeding community. The genetic resources of V. sylvestris in European populations represent important sources of genetic variability that are worth preserving. However, no research has investigated the presence of resistance-linked alleles at the Ren1 locus.
V. sylvestris has been referenced in Croatian literature historically16,17. Recently, Croatian V. sylvestris was systematically identified and characterized18,19,20 as belonging to five natural populations in humid Mediterranean forests and in sub-Mediterranean ecosystems19. Prospecting at natural sites is ongoing and recently two additional populations were identified (data not published). However, these studies did not evaluate resistance to fungal disease among the populations.
To bridge that gap, the present work (i) evaluates PM resistance of V. sylvestris accessions from the eastern Adriatic region (Croatia and Bosnia and Herzegovina) using simple sequence repeat (SSR) markers linked to the powdery mildew resistance locus Ren1, and (ii) performs a detailed phenotypic evaluation of visible powdery mildew symptoms on V. sylvestris seedlings. This is the first report of PM evaluations in V. sylvestris collected in Croatia and Bosnia and Herzegovina and identifies germplasm that could be used for grapevine breeding.
Results
Disease evaluations
The inventory of V. sylvestris in their natural habitats found very few vines with powdery mildew (PM) symptoms. Only ~ 7% of individuals developed PM symptoms, mostly from the Krka National Park population. Detailed evaluation for PM resistance continued on 67 V. sylvestris seedlings grown in pots in an ex-situ seedling collection.
Seedlings were previously established from seeds of five female individuals during inventory work aiming at gene conservation. Each seedling was represented by one biological replicate due to juvenile growth phase and lower plant vigour in a shaded collection.
Visual PM symptoms on each accession were rated according to the OIV 455 descriptor using a five-class scale (1–9), for both in vivo observations of the collection and in vitro leaf disk testing (Fig. 1).

Grapevine powdery mildew (E. necator) mycelium development on V. sylvestris leaves. Powdery mildew spores were inoculated onto the top (adaxial) surface of leaf disks of V. sylvestris seedlings. Each figure represents fungal growth on a single leaf disk of five different genotypes according to the OIV 455 scale from 1 to 9 (left to right), respectively. The PM-susceptible seedlings received a lower scale number, while resistant seedlings received a higher number.
Two-year in vivo evaluation of PM on entire seedling plants was possible for 62 and 57 seedlings out of 67 seedlings in 2018 and 2019, respectively (Fig. 2a). Failing accessions either dried up or were too small at the time of observation. Nearly half of the seedlings analyzed in 2018 (30 accessions) showed intermediate PM resistance (score 5). Twenty-two accessions were resistant: 15 resistant and seven very resistant (scores 7 and 9, respectively). Nine accessions were susceptible and one was very susceptible to PM (scores 3 and 1, respectively). Similar trends were observed in 2019. In both years, intermediate resistance was confirmed in 12 accessions, resistance in nine and susceptibility in four (Table 1). Three control accessions of V. vinifera cultivar ‘Plavac mali sivi’ showed resistance to high resistance, except that accession PMS22 had intermediate resistance in 2018 (Fig. 2a).

Resistance of V. sylvestris seedlings to grapevine powdery mildew (E. necator) based on OIV 455 descriptor five-class scale (1–9) and two phenotypic approaches: (a) In vivo evaluation in 2018 encompassed 62 accessions and 57 accessions in 2019. V. vinifera cv. ‘Plavac mali sivi’ (PMS) was used as control. (b) Leaf disk assay on a subset of 35 V. sylvestris accessions in 2018 and 2019. Three V. vinifera cvs. were used as controls. Each chart column in (a,b) shows the number of different accessions sharing the same OIV score (level of resistance) in the year of evaluation. Unless otherwise indicated, all accessions were represented by a single biological replicate.
A subset of 35 seedlings with the best fitness in 2018 and 2019 was chosen for the leaf disk assay. Three V. vinifera cvs. were used as controls (Fig. 2b). The majority of seedlings (20 accessions) in 2018 showed PM resistance: 18 were resistant and two very resistant. Intermediate resistance was found in 11 accessions and four were susceptible. In 2019, only five accessions of the same subset were resistant, 17 accessions had intermediate resistance, while 13 were susceptible. Three seedlings (SjCer14, SjCer23 and SjCer24) had leaf disk PM resistance in both years. Three seedlings had intermediate resistance and one was susceptible in both years (Table 1). Control cv. 'Plavac mali sivi' (PMS 39) showed intermediate resistance, 'Pošip bijeli' (PSP 01 and PSP B2) displayed very high resistance in 2018, but were susceptible and resistant, respectively, in 2019. 'Rukatac' (RKTC 02) was rated highly resistant in 2018 and very susceptible in 2019.
Genetic polymorphism at the Ren1 locus with reference to phenotypic disease evaluation
Genetic SSR data based on three markers: SC47-18, SC8-0071-14 and UDV124, were adapted to match alleles with previous studies7,15. Thirty-two out of 67 seedlings had the R-allele at one or two SSR markers (Table 1). R-allele 239 at marker SC47-18 was the most frequent (29 seedlings). Five seedlings had R-allele 212 at UDV124. Two seedlings had both 239 and 212 R-alleles.
Seedlings from the Cerovica population showed the greatest overall phenotypic resistance (mean value 6), while seedlings from the Gizdavac population had the least overall resistance (4.97), despite relatively high R-allele variability.
None of the seedlings showed full resistance, evaluated using two phenotypic approaches, over both years (Table 1). However, three seedlings were resistant when analyzed in vivo and in one year of leaf disk assay: SjPak13 (no R-alleles), SjCer7 (allele 212), and SjCer12 (allele 239). Three Cerovica seedlings, SjCer14, SjCer23, and SjCer24, showed resistance in leaf disk assays and in one year of in vivo. Seedling SjCer23 had two R-alleles (239 and 212), SjCer14 one (239), and SjCer24 had none of the R-alleles.
PM resistance of in situ V. sylvestris genotypes
Ninety-one V. sylvestris individuals from natural habitats (in situ) were analyzed at three SSR markers (Table 2). Forty-one individuals carried alleles associated with PM resistance. All these individuals had R-allele 239 at SC47-18 except one (Im4), which had R-allele 246 at the same marker. One accession (Luk8) contained both alleles: 239 and 246. The Paklenica population had the most individuals carrying R-alleles (13), followed by Psunj (12) and Lukovdol (8). Populations Imotski (4), Krka (2) and Gizdavac (1) had the fewest individuals with R-alleles.
None of the in situ-tested individuals had R-alleles at SC8-0071-014 and UDV124, despite polymorphisms at both markers (Table 2).
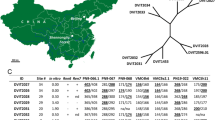
The set of 91 in situ-tested individuals was subjected to neighbor-joining (NJ) clustering and perceptual mapping (Principal Coordinate Analysis, PCoA) to visualize the similarity among individuals based on SSR allelic profiles (Fig. 3a,b). The NJ tree showed clusters of mixed individuals from different populations, only roughly outlining their geographical origin. However, individuals with R-alleles are clearly more abundant on the right side of the tree (red dots, Fig. 3a). Relative relationships between individuals based on PCoA showed the same pattern as NJ. They separated individuals with R- alleles on the right side of plot from individuals without R-alleles at Ren1 on the left side. PCoA projections of the first two dimensions accounted for 33.69% of the total molecular variation. PCoA revealed slight overlapping between the two groups (Fig. 3b).

Clustering of 91 wild grapevine individuals based on three SSR markers at the Ren1 locus. (a) Neighbor-joining dendrogram showing genetic relationships among individuals with bootstrap support value ≥ 75%. The samples with resistance alleles are marked with red dots. Analyses were conducted in MEGA721. (b) Principal coordinate (PCoA) projections of individuals in a 2-D plot based on genotypic distances conducted in GenAlex22.
Genetic diversity
Genetic diversity was calculated for the two sets of V. sylvestris: 91 in situ individuals and 67 seedlings. Three SSR markers were polymorphic in both sets (Table 3). The observed heterozygosity was lower than the expected heterozygosity, except at marker UDV124 and for the in situ set at marker SC47-18. The in situ set showed more heterozygosity at the tested markers than the seedling set. The allele frequency (AF) for resistance-related allele 212 at UDV124 was determined only in the seedling set with (AF = 0.04). At marker SC8-0071-014, no resistance-linked alleles were observed. The AF was lower than 0.3 for all alleles except for the 197 allele in seedlings, which was present in the entire Cerovica set (Table 1). The greatest abundance of R-alleles was found at marker SC47-18: R-allele 239 was found in the in situ and seedling sets at an AF of 0.25 and 0.26, respectively. An additional R-allele 246 was observed in the in situ set only, at a very low AF of 0.01.
Discussion
This study represents the first screening for powdery mildew resistance in eastern Adriatic V. sylvestris germplasm. The phenotypic focus of study was on the V. sylvestris seedlings in an ex situ collection. These were young potted plants evaluated for powdery mildew resistance under in vivo and in vitro conditions. The resistance level of seedlings varied from very susceptible to medium to highly resistant. The control cultivars in this study showed medium to very high resistance (OIV455 scores from 5 to 9), except cvs. ‘Rukatac’ and ‘Pošip bijeli’ in 2019. These white grape cultivars are considered susceptible to fungal diseases. None of the control cultivars had R-alleles at the three SSR markers, even though cv. ‘Plavac mali sivi’ is known to have good tolerance toward fungal infections. Control cultivars clearly expressed more robust vigour than V. sylvestris accessions. Twenty-two and 23 in vivo-evaluated seedlings showed high resistance to PM in 2018 and 2019, respectively. The majority of seedlings showed PM infection, indicating homogeneous field infections. Seedlings from the Cerovica population (Bosnia and Herzegovina; Fig. 4, Table 1) had the greatest average resistance and the most accessions with R-alleles (23 out of 27 accessions). Cerovica, which is part of the large Neretva population in Bosnia and Herzegovina, contained a considerable number of different and private alleles20 and possibly retained an important fraction of the historic V. sylvestris biodiversity. The seedlings from Paklenica and Gizdavac had intermediate resistance. The Paklenica population is one of the most-conserved V. sylvestris populations in Croatia and had the most R-alleles in its in situ accessions, while Gizdavac is one of the most vulnerable populations, close to urban centers and with visible human impact19,23. Seedlings from Gizdavac had greater allelic diversity of SSR markers than those from Paklenica (the least), even though the overall phenotypic resistance of Gizdavac seedlings was the lowest in this study.

Map of seven in situ Croatian V. sylvestris populations and the Cerovica population in Bosnia and Herzegovina.
During in vivo evaluation, nine seedlings were confirmed as resistant in both years. Of these nine, three were susceptible in the leaf disk assay. Seedling SjCer7 had the greatest leaf disk resistance and carried R-allele 212 at marker UDV124. This allele is associated with resistance in V. vinifera 6. On the other hand, four seedlings were susceptible both years in vivo. One had a single R-allele 212, two had a single R-allele 239 and one had no R-alleles.
PM resistance (score 7) in the seedling subset leaf disk assay was confirmed in both years for only three accessions from the Cerovica population. The resistant accessions SjCer14, carrying the single allele 239, and SjCer23, carrying two alleles (212 and 239), had the same phenotypic disease scores regardless of the evaluation method and were slightly more resistant than SjCer24, the resistant accession without R-alleles. Among the entire seedling set, only two accessions from Cerovica, SjCer1 and SjCer23, had R-alleles at two markers (UDV124 and SC47-18) and expressed PM resistance.
In general, disease evaluations are very subjective and vary among studies due to different environmental conditions, experimental set up, disease pressure, and pathogen population structure. The phenotypic evaluations conducted in this study differed to some extent from Riaz et al.7, in which a modified and inverted OIV scale was used. Similar modifications were reported in other studies8,10,24,25. The seedling plants in this study were very young at the time of phenotypic evaluations, which probably affected both the level of resistance and disease pressure to some extent. Young plants do not have excess vegetative growth, which allows more air circulation and potentially reduces infection severity. Nevertheless, the final outcomes were comparable. In this study, most seedlings showed intermediate resistance to PM, characterized by clearly visible, but localized and fragmented, sporulation with small patches of mycelium. However, in the 2018 in vitro analysis, most seedlings expressed resistance to PM. Mycelial growth was highly suppressed and conidiospores were not observed. The resistance of the seedling set under in vitro conditions varied greatly between the two seasons (14 and 57% of the accessions expressed resistance). Under in vivo conditions, unsprayed, naturally-infected plants showed more consistency between seasons, with the number of resistant seedlings varying between 35 and 40%. This variation might be because of using more mature leaves in the assay and variation in the PM inoculum, which was collected from the field. Field PM strains can vary from one year to another26, which might cause variations in resistance levels. In our samples, intermediate resistance was predominant. This differed from phenotyping results of Riaz et al.7, where the intermediate rating was not observed among the seedlings. This was probably due to more consistent in vitro assay infections and inoculation with a single PM strain collected from pure cultures7. In our study, randomly-collected, spontaneous field PM inoculum was used. Moreover, full parentage was known for only nine seedlings (23% of the seedling set), thus information on the phenotypic resistance to PM of the in situ parental lineages is entirely missing. In a previous study7, the F1 population of seedlings resulted from crosses between known resistant (V. sylvestris) and susceptible (V. vinifera) parents. Research on ‘omics’ provides additional and unique insight into the linking of phenotypes with genotype interactions and plant stress responses27. Comprehensive research on the transcriptomics of Central Asian accessions revealed varying levels of phenotypic resistance to PM, matched with various transcriptomic responses to E. necator among Central Asian accessions24.
The three Ren1-linked SSR markers on chromosome 13 that we analyzed in situ in Croatian V. sylvestris populations and in seedlings were highly polymorphic. The observed heterozygosity was lower than the expected heterozygosity, indicating lower genetic diversity among the studied sets at the three SSR markers. This is similar to what Riaz et al.7 found at four (of five) R-SSR loci for the Central Asian grape germplasm, pointing to inbreeding due to geographic isolation of the populations. Increased gene flow might be assumed, for instance, in seedlings from Gizdavac, where out of two individuals, only the male parent, Giz2, had the R-allele 239, which was not found in its progeny (SjGiz4). Eight seedlings from Gizdavac had various R-alleles that were most likely inherited from their unknown paternal parents. The Gizdavac population, near urban centers, differs from the protected Paklenica population, where most in situ individuals had an R allele. Only two progeny seedlings had an R-allele, 239, which they shared with their mother accession, Pak12. SNP Genotyping by Sequencing approach revealed significant and unexpectedly high segregation distortions from Mendelian ratios on chromosome 13 in the susceptible V. vinifera cv. ‘Glera’, making one end of its chromatide less-inherited in the offspring28.
In this study, none of the accessions had R-alleles (141 or 143) at the SC8-0071-14 locus. This locus was previously shown to be polymorphic and a significant Quantitative trait loci (QTL), explaining up to 96% of the variation. It was mapped together with the SC47-18 locus. In a previous study7, SC8-0071-014 R-alleles were detected in both resistant and susceptible accessions. Here, accessions carrying R-alleles at UDV124 and SC47-18 were either resistant or susceptible. Locus UDV124 flanks Ren1 on chromosome 13, while loci SC8-0071–014 and SC47-18 co-segregated with resistance (Ren1)29. Clustering by NJ and PCoA both separated individuals carrying R-alleles to the right side of the graphs (Fig. 3a,b). There was no clear phylogenetic separation of individuals by population of origin, although there was rough separation between northern and southern populations (Fig. 3a). PCoA more clearly visualized the overlapp between two groups. These results may indicate a common source of Ren1 loci in studied V. sylvestris. However, as was recently discussed for Caucasus V. vinifera cvs., high complexity at the chromosome 13 region and information gathered thus far does not permit conclusions as to whether studied genotypes from the Caucasus and Central Asia share the same resistance genes28.
This study provides insight into powdery mildew resistance of V. sylvestris accessions from the eastern Adriatic region. The observed phenotypic resistance of V. sylvestris individuals to powdery mildew was clear and consistent for some individuals, but showed some weakness in connecting phenotypic and genetic resistance in other individuals. The SC47-18 marker (R-allele 239) co-segregated with resistance at Ren1. It was the most dominant marker in our V. sylvestris set and there were no R-alleles at another co-segregating marker, SC8-0071-014. However, a trend with two new allelic combinations was found in the studied set. This has interest for a deeper investigation on functional PM resistance related to that marker and the Ren1 locus. No substantial connection was observed between individuals with greater phenotypic resistance and expected resistant genotypes. This result was in accordance with previous results, in which both resistant and susceptible genotypes had R-alleles. However, accessions with two R-alleles at two different SSR markers showed, on average, greater resistance to powdery mildew in this study. The presence of R-alleles in the eastern Adriatic V. sylvestris confirms these genetic resources as important sources of biodiversity. Next-generation sequencing technologies and other –omics methods would be beneficial to access more information on the nature of PM resistance in the eastern Adriatic region.
Methods
Plant material
A total of 158 unique V. sylvestris genotypes were analyzed in this study. The sample set consisted of 91 V. sylvestris genotypes from seven natural populations in Croatia (in situ) and 67 V. sylvestris seedlings established from seeds of five V. sylvestris plants from three natural populations: Paklenica and Gizdavac in Croatia and Cerovica in Bosnia-Herzegovina. All V. sylvestris individuals included in this study were identified morphologically and using 20 SSR markers through our previous inventorying of V. sylvestris in Croatia and neighboring countries18,20,30. Permission to collect and examine the plant species Vitis vinifera subsp. sylvestris within protected natural areas of Croatia has been granted by the Ministry of Economy and Sustainable Development of the Republic of Croatia (Permission No. UP/I-612-07/15-33/74). The voucher specimens were identified by Goran Zdunić and Katarina Lukšić and are deposited in the publicly accessible herbarium of the Institute for Adriatic Crops and Karst Reclamation, Split, Croatia.
The natural habitats of the studied species cover diverse geographic areas: the eastern coast of the Adriatic Sea, the mountainous area of the Dinaric Alps, and the mountainous area in the Croatian part of the Pannonian Basin. Supplementary Table S1 lists the analyzed genotypes, their geographical origins and their sampling locations. The seven natural V. sylvestris populations were from Imotski, Grab, Gizdavac, Krka, Paklenica, Lukovdol and Psunj (Fig. 4).
Disease evaluations
PM symptoms were first observed on wild individuals in situ during an inventory of V. sylvestris around bloom. The overall health status of each plant was evaluated. PM symptoms on green tissues, including leaves (adaxial surfaces), were noted: white to grayish coatings of fungal colonies, upward leaf curling, and leaves that were drying and falling off.
The in situ V. sylvestris plants were not subjected to leaf disk assays in this study due to their spatial dislocation and the difficulty of keeping the plant material fresh and ensuring uniform trial conditions.
PM evaluation of the 67 V. sylvestris seedlings was performed (with no biological replicates) on plants and via a leaf disk assay (only 35 seedlings) using the OIV 455 descriptor31. Each genotype was evaluated by visual inspection for signs of pathogens using a stereomicroscope and the OIV 455 scale: 1 = unlimited infection; complete or nearly complete attack of the leave surface, abundant mycelial growth 3 = vast attacked patches, some of which were limited, obvious mycelial growth and fungus fructification; 5 = attacked patches were frequent, but usually clearly limited; 7 = sparse, small and limited attacked patches; little mycelium and fungus fructification, 9 = greatly suppressed symptoms or none at all; no mycelium or visible fructification.
The seedlings were grown in pots in an outdoor shaded area and were not sprayed from the beginning of vegetation, before and during the PM evaluations. The disease evaluations were conducted during 2018 and 2019.
The subset of 35 out of 67 seedlings selected for the in vitro leaf disk assays included only individuals with the best health status and overall fitness.
The in vivo PM evaluations were performed on May 21st in 2018 and on May 13th in 2019, during the highest disease pressure in the field collection period. The observations encompassed 67 V. sylvestris seedlings and the cv. 'Plavac mali sivi' as a control with three accessions. The in vivo inspections were carried out by evaluating an approximately equal part of each plant and taking into consideration the overall plant health, after which a single OIV 455 score was assigned per genotype.
For the leaf disk analyses, besides 35 V. sylvestris samples (Table 1), four potted cvs. 'Plavac mali sivi' (1 replicate), 'Pošip bijeli' (2 replicates) and 'Rukatac' (1 replicate), were included as references. The pink cv. 'Plavac mali sivi' is generally known to be less susceptible to mildew, while the white cvs. 'Rukatac' and 'Pošip bijeli' are susceptible.
Leaf disk assays
The in vitro analyses were performed by placing leaf disks on 1% water-agar (Bacto™ Agar, BD, France) in plastic Petri dishes, as described8,32. The leaf disks were infected with E. necator using fresh conidia from naturally infected leaves of susceptible V. sylvestris seedlings grown in the collection. The spores were collected with a small soft brush and transferred onto each adaxial leaf surface. The fourth to sixth fully developed leaves of each genotype were sampled: two leaves (four leaf disks) were analyzed per sample. The samples were placed in a climate chamber at 25 °C and 65% relative humidity on a 16-h light, 8-h dark cycle. Seven to nine days post inoculation (dpi), leaves were evaluated for mycelial growth and conidiophore formation. Evaluations of the infections were performed using a CarlZeiss stereomicroscope (Microimaging, GmbH Germany), and detailed inspections together with photos were captured under 0.65 × magnification.
DNA extraction and marker analysis
DNA was extracted from the young leaves using the NucleoSpin Plant II kit (Macherey–Nagel, Düren, Germany). The extracted DNA was quantified and used at a working DNA concentration of 1 ng/μL.
Three microsatellite markers, SC47-18, SC8–0071–14 and UDV1246, that are associated with the Ren1 gene for PM resistance were analyzed. The SSR markers were multiplexed within one run. All forward primers were labeled on the 5' end with fluorescent dyes (NED, 6-FAM and VIC).
A KAPA Fast Multiplex PCR Kit (2x) (Kapa Biosystems, Wilmington, MA) was used to set up 5-µL volume reaction mixtures containing a master mix, 100 pmol of each primer, and ~ 1 ng template DNA. PCR amplification was performed using the following program: three minutes of initial denaturation at 95 °C, followed by 30 cycles of denaturation at 95 °C (15 s), annealing at 60 °C (30 s), and extension at 72 °C (30 s). A final extension was performed at 72 °C for seven min.
The amplified products were resolved using capillary electrophoresis on an ABI 3730xl Genetic Analyzer (Applied Biosystems) using GeneScan-LIZ 500 as an internal standard. The peaks were identified with GeneMapper 4.0 software (Applied Biosystems). The allele calling was harmonized with previously published SSR data7,15 by amplifying two identical V. sylvestris samples separately at the University of California at Davis and at the Laboratory of the IAC in Split. Then, the allele sizes were aligned among the other samples.
The trueness to type of the V. sylvestris set was analyzed and discussed in a previous study30.
Data analysis
The phenotypic data and corresponding statistical analyses were performed in Excel 2013. The disease scores were determined according to the OIV 455 scale: by calculating mode values from four leaf disk scores per genotype each year and by determining a direct, single OIV score per genotype for all in vivo evaluations each year.
Descriptive statistics of the SSR markers used in this study were calculated for the following indices: the number of different alleles per locus (Na), expected heterozygosity (He), observed heterozygosity (Ho) and allele frequency (AF). These statistics were obtained using the GenAlEx 6.5 package22. Principal coordinate analysis (PCoA) was performed to visualize relationships between individuals based on the Ren1 scoring data via covariance, standardized using the same program.
The SSR data for 91 in situ individuals were analyzed to compute their evolutionary history using the Neighbor-Joining method33 implemented in MEGA 7.0 software21. The bootstrap interior branch test was used to test the reliability of each interior branch on the tree34.
Data availability
All data generated or analyzed during this study are included in this published article (and its Supplementary Information files). Correspondence and requests for materials should be addressed to G.Z. The study complies with local and national regulations.
References
Pirrello, C. et al. Emergent Ascomycetes in viticulture: An interdisciplinary overview. Front. Plant Sci. 10, 1394 (2019).
Jones, L. et al. Adaptive genomic structural variation in the grape powdery mildew pathogen, Erysiphe necator. BMC Genomics 15, 1081 (2014).
European Commission. The Use of Plant Protection Products in the European Union Data 1992–2003. (European Union: Brussels, 2007).
Riaz, S., Tenscher, A., Pap, D., Romero, N. & Walker, M. A. Durable powdery mildew resistance in grapevines: Myth or reality. Acta Hortic. 1248, 595–600 (2019).
Suthaparan, A. et al. Suppression of powdery mildews by UV-B: Application frequency and timing, dose, reflectance, and automation. Plant Dis. 100, 1643–1650 (2016).
Dry, I., Riaz, S., Fuchs, M., Sosnowski, M. & Thomas, M. Scion breeding for resistance to biotic stresses. in The Grape Genome (eds. Cantu, D. & Walker, M. A.). 319–347. (Springer, 2019).
Riaz, S., Menéndez, C. M., Tenscher, A., Pap, D. & Walker, M. A. Genetic mapping and survey of powdery mildew resistance in the wild Central Asian ancestor of cultivated grapevines in Central Asia. Hortic. Res. 7, 1–15 (2020).
Zendler, D., Schneider, P., Toepfer, R. & Zyprian, E. Fine mapping of Ren3 reveals two loci mediating hypersensitive response against Erysiphe necator in grapevine. Euphytica 213, 68 (2017).
Hoffmann, S. et al. Resistance to Erysiphe necator in the grapevine ‘Kishmish vatkana’is controlled by a single locus through restriction of hyphal growth. Theor. Appl. Genet. 116, 427–438 (2008).
Zendler, D., Töpfer, R. & Zyprian, E. Confirmation and fine mapping of the resistance locus Ren9 from the grapevine cultivar ‘Regent’. Plants. 10, 24 (2021).
Ocete, R., López, M. Á., Gallardo, A. & Arnold, C. Comparative analysis of wild and cultivated grapevine (Vitis vinifera) in the Basque Region of Spain and France. Agric. Ecosyst. Environ. 123, 95–98 (2008).
Meléndez, E. et al. Evolution of wild and feral vines from the Ega river gallery forest (Basque country and Navarra, Spain) from 1995 to 2015. J. Int. Sci. Vigne Vin. 50, 65–75 (2016).
García, R.A.A. & Revilla, E. The current status of wild grapevine populations (Vitis vinifera ssp. sylvestris) in the Mediterranean basin. in The Mediterranean Genetic-Code-Grapevine and Olive (ed. Sladonja, B.). https://www.intechopen.com/books/the-mediterranean-genetic-code-grapevine-and-olive/the-current-status-of-wild-grapevine-populations-vitis-vinifera-ssp-sylvestris-in-the-mediterranean (2013).
Cantos, M. et al. Current distribution and characterization of the wild grapevine populations in Andalusia (Spain). C. R. Biol. 340, 164–177 (2017).
Riaz, S. et al. Identification of mildew resistance in wild and cultivated Central Asian grape germplasm. BMC Plant Biol. 13, 149 (2013).
Turković, Z. & Aničić-Bošnjak, Z. Vitis vinifera subspecies silvestris Gmel. Agron. Glas. 9, 657–670 (1953).
Turković, Z. & Sučević-Šafar, O. Analize Vitis silvestris. Agron. Glas. 11–12, 817–824 (1953).
Zdunić, G. et al. Genetic diversity of wild grapevine [Vitis vinifera L. subsp sylvestris (Gmel.) Hegi] in the Eastern Adriatic Region. Am. J. Enol. Vitic. 68, 252–257 (2017).
Zdunić, G. et al. Inventory and descriptions of wild grapevine (Vitis vinifera subsp. sylvestris) from Slovenia, Croatia and Bosnia and Herzegovina. Acta Hortic. 1248, 63–68 (2019).
Zdunić, G. et al. Genetic structure and relationships among wild and cultivated grapevines from Central Europe and part of the Western Balkan Peninsula. Genes 11, 962 (2020).
Kumar, S., Stecher, G. & Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33, 1870–1874 (2016).
Peakall, R. & Smouse, P. E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-An update. Bioinformatics 28, 2537–2539 (2012).
Butorac, L. et al. Assessment of wild grapevine (Vitis vinifera ssp. sylvestris) chlorotypes and accompanying woody species in the Eastern Adriatic region. PLoS ONE 13, e0199495 (2018).
Amrine, K. C. H. et al. Comparative transcriptomics of Central Asian Vitis vinifera accessions reveals distinct defense strategies against powdery mildew. Hortic. Res. 2, 15037 (2015).
Zyprian, E. et al. Quantitative trait loci affecting pathogen resistance and ripening of grapevines. Mol. Genet. Genomics. 291, 1573–1594 (2016).
Martinson, T. & Cadle-Davidson, L. The Phenotyping Bottleneck: How Grape Breeders Link Desired Traits to DNA Markers. https://cpb-us-e1.wpmucdn.com/blogs.cornell.edu/dist/c/7890/files/2019/01/The-Phenotying-Bottleneck-_-Wines-and-Vines-Dec-2018-1w6gc3o.pdf (2018).
Harfouche, A. L. et al. Accelerating climate resilient plant breeding by applying next-generation artificial intelligence. Trends Biotechnol. 37, 1217–1235 (2019).
Possamai, T. et al. Construction of a high-density genetic map and detection of a major QTL of resistance to powdery mildew (Erysiphe necator Sch.) in Caucasian grapes (Vitis vinifera L.). BMC Plant Biol. 21, 528 (2021).
Coleman, C. et al. The powdery mildew resistance gene REN1 co-segregates with an NBS-LRR gene cluster in two Central Asian grapevines. BMC Genet. 10, 89 (2009).
Lukšić, K. Morphological and genetic diversity of wild grape (Vitis vinifera subsp. sylvestris Gmel Hegi) in Croatia. Doctoral Thesis. https://urn.nsk.hr/urn:nbn:hr:204:069255 (2020).
O.I.V. 2nd Edition of the OIV Descriptor List for Grapevine Varieties and Vitis Species. OIV Website. https://www.oiv.int/en/technical-standards-and-documents/description-of-grape-varieties/oiv-descriptor-list-for-grape-varieties-and-vitis-species-2nd-edition (2007).
Staudt, G. Evaluation of resistance to grapevine powdery mildew (Uncinula necator [Schw.] Burr., anamorph Oidium tuckeri Berk.) in accessions of Vitis species. Vitis. 36, 151–154 (1997).
Saitou, N. & Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425 (1987).
Dopazo, J. Estimating errors and confidence intervals for branch lengths in phylogenetic trees by a bootstrap approach. J. Mol. Evol. 38, 300–304 (1994).
Acknowledgements
We gratefully acknowledge the funding support of the Croatian Science Foundation (grant: UIP-2014-09-9737 and “Young researchers’ career development—training new doctoral students”) and the European Regional Fund (grant KK.05.1.1.02.0010).
Author information
Authors and Affiliations
Contributions
K.L., G.Z. and I.P. designed research; K.L. carried out phenotyping and genotyping of germplasm; K.H., M.Ž.M. and A.M. performed research; E.M. and S.R. advised, reviewed and edited paper; K.L. and G.Z. wrote the paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lukšić, K., Zdunić, G., Hančević, K. et al. Identification of powdery mildew resistance in wild grapevine (Vitis vinifera subsp. sylvestris Gmel Hegi) from Croatia and Bosnia and Herzegovina. Sci Rep 12, 2128 (2022). https://doi.org/10.1038/s41598-022-06037-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-06037-6
- Springer Nature Limited