
Abstract
In this contribution, we investigated the role of plants in the prehistoric community of Casale del Dolce (Anagni, FR, central Italy), through microparticles recovered from dental calculus. The finding of a great amount of pollen types, even in form of compact lumps, could indicate use of natural substances, such as honeybee products and/or conifer resins. This plant-microremain record also suggested environmental implications relative to the Neolithic and Chalcolithic period. Additionally, the stability of the tartar microenvironment had preserved starches and other microparticles, such as one epidermal trichome, a sporangium, and fragments of plant tissue, rarely detected in ancient dental calculus. The detection of secondary metabolites in the ancient matrix confirmed the familiarity of this community with plant resources. All these data supply various interesting food for thought and expand the knowledge about the potential of dental calculus in archaeological and archaeobotanical fields with a special focus on palaeoecology.
Similar content being viewed by others

Introduction
Prehistorical and archaeological context
The spread of Neolithic culture in the Italian peninsula started from VI millennium B.C.1. The Neolithic transition began with populations which probably had come from the East. The first Neolithic cultural pattern arrived in the southern regions of Italy and moved northwards spreading along two sides: the Tyrrhenian and Adriatic coastlines. The impact of this phenomenon determined the onset of different bio-cultural innovations on both sides. While the Adriatic communities rapidly absorbed these new models, the Tyrrhenian ones (with reference to central Italy and specifically Latium) persisted for a long time in a condition still very similar to that of the last hunter-gatherers of the upper Palaeolithic. This is confirmed by the presence of so-called aceramic sites, populated by individuals morphologically very similar to the cro-magnonoid types, as in the case of Cisterna di Latina2. The advent of the Eneolithic culture in these territories produced a real cultural and biological revolution. In fact, the permeation of Gaudo's culture determined the introduction of a wider range of socio-economic innovations (e.g., changes in settlement structures, cultural assemblages, and mortuary practices), anatomical modernity, and further refinement of the encephalization process, the brachicrania3,4. In this regard, the site of Casale del Dolce (185 m.a.s.l., Anagni, Latium, central Italy) would represent the transition from the Neolithic to the Eneolithic period, with radiocarbon calibrated dates at 4.927 and 4.867 BP5, in which Gaudo’s culture is witnessed but brachycephalicalization not yet.
The archaeological context is characterized by inhabited and productive structures distributed on a series of terraces overlooking the Sacco River. The economic regime of this prehistoric community, settled in the valley from the Neolithic both for the fertile nature of the soil and for the relative proximity to the water source, would seem to be developed gradually out of a spectrum of well-established Neolithic subsistence practices.
Stable isotope signatures of several Neolithic and Eneolithic communities from central Italy would suggest a general sedentary lifestyle and a subsistence economy based on the procurement of local resources, with a general inclination to consume C3 plants and C3 consumer backbone resources. Interestingly, the same biomolecular approach has demonstrated that the individuals of Casale del Dolce could have a diet with a greater intake of carbohydrates compared to the others6,7. Additionally, more than 500 anthracological and carpological remains (e.g., Carpinus sp., Quercus sp., Pomoideae, Corylus sp.) were detected in the investigated site and analysed5,8. All this evidence would indicate that the Tyrrhenian community of Casale del Dolce was mainly devoted to agricultural practices and maximized the exploitation of the local environment, which had promoted its stabilization in the area. To corroborate this supposition, dental calculus analysis was performed on the human skeletal series of Casale del Dolce, addressing relevant aspects of the exploitation of plant and natural resources.
Ancient plant diversity and dental calculus
Palaeobotany and archaeobotany are very informative topics of research which are characterized by interdisciplinary studies. In particular, the second one should be considered a valuable link between science and humanities. The scientific investigation on all types of plant remains, preserved both in natural and anthropogenic sediments, contributes to the comprehension of human–environment interaction and cultural diversity of our ancestors, in terms of evolution of phyto-associations, ecology, human impact, land exploitation, plant use, diet, and trades9,10,11,12,13.
So far, the study of plant and animal macroremains, the characterisation of organic residue in pottery, the analysis of stable isotopes, and the investigation of dental microwear have represented the most common methods to understand ancient habits. Nowadays, many research groups have exploited the great informative potential contained in ancient dental calculus to open a window on past lifeways. The recovery of microparticles trapped inside this matrix has been employing to infer new awareness about the impact of plants on different prehistoric and historical communities14,15,16,17,18,19,20,21,22,23. The interpretation of such type of results is still quite problematic and one of the major concerns occurs in establishing the authenticity of ancient particles, especially starch24,25. Due to the multifactorial aetiology of tartar and the individual variability of its growth, a linear correlation between plant consumption and presence of plant microremains in a calculus deposit does not exist. Additionally, how, when, and where different natural microdebris (i.e., from plants, minerals, and animals) have been trapped into the calculus remain quite challenging to assess26. As reported in literature27, several environmental particles, such as pollen, plant fibres, micro-charcoals, soot, soil particles, and dirt, grit and dust settled on food, can be also embedded in this matrix as a consequence of inhalation or accidental ingestion. Unambiguous interpretative scenarios are arduous to be defined for some types of microparticles, such as pollen and trichomes; thus, it is necessary to consider and critically explore all possible pathways of inclusion for them, beyond inhalation hypothesis. Published data mainly have focused on identifiable and generally diagnostic microparticles, such as starch granules. However, ancient calculus has revealed the capacity to preserve other microremains, such as trichomes, which would represent a direct link with the environment surrounding the individuals. For example, pollen can be considered as a bioindicator of plant diversity.
In this respect, our contribution aims at reconstructing the dietary ecology of the Neo-Eneolithic community of Casale del Dolce (Fig. 1). It also uses ancient dental calculus to deduce palaeoenvironmental implications, through the micro-botanical findings embedded in this archaeological repository.

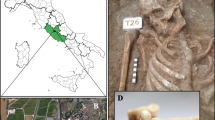
Archaeological site and masticatory apparatus of CDD7. Location of Casale del Dolce archaeological site (A): images produced using Google Earth Pro, V 7.3.3.7786 (June 25, 2019) Casale del Dolce (Latium, central Italy), 41° 41′ 47.43″ N, 13° 07′ 43.24″ E, Eye alt 184 m; Rome (Latium, Italy), 41° 54′ 10.02″ N, 12° 29′ 46.92″ E, Eye alt 47 m; Image Landsat/Copernicus, Data: SIO, NOAA, U.S. Navy, NGA, GEBCO, https://www.google.com/earth/ [Accessed September, 2020]; skeletal remains of CDD7 (image captured by Prof. Mauro Rubini) (B).
Results and discussion
Morphological analysis
Food preparation or processing of plant material involve multiple activities and all of them can potentially leave micro-traces in the tartar, together with the environmental components.
Eleven dental calculi showed plant record: starches, pollen grains, one trichome, one sporangium, and tissue fragments (Table 1).
Plant hair
Trichomes are epidermal outgrowths characterized by different structure and function. Although plant hairs are some of the most common findings in the overall particulate matter carried by air (as pollen grains), in literature, only very few examples of trichomes in ancient contexts have been reported28,29,30. Trichome identification is not a common area in dental calculus research since they do not have a diagnostic morphology. For this reason, the identification of such type of microdebris must be based on realistic criteria, also in accordance with the geographical and historical context, providing all possible interpretative scenarios. The detection of trichomes in ancient tartar may disclose other lines of evidence than nutrition, representing a reliable archaeological environmental proof31.
One plant hair was identified in CDD1 sample (Table 1). This remain (Fig. 2L) falls into the general class of dendritic trichomes and its peculiar morphology has been more specifically termed a candelabrum or abietiform32. The overall structure corresponded to non-glandular and pluricellular trichomes with a central uniseriate axis and whorls of unicellular rays emerging at the joints of the axis. Usually, 4 radii from each node occurred perpendicular to the central axis. As exhaustively reported in literature, dendritic trichomes are known in ferns, different groups of modern monocots and basal eudicots, such as Scrophulariaceae and Platanaceae. Although dendritic, trichomes of ferns and monocots were excluded. Indeed, the first ones possess single secondary branches that alternatingly arise at an angle of 70°–120° with respect to the main axis along a single plane33, while the second ones show morphological features and appearance different from the ancient debris34,35. Candelabrum-like trichomes have been usually detected in Verbascum L. and Platanus L. species36. For this work, an experimental reference collection of trichomes from these plants was created (Supplementary Information 1). The general aspect of mullein trichomes appears to be capitate, bigger, more elongated, and slenderer than the microremain found in tartar sample. In addition, these trichomes seem to possess a pair of secondary elements per side or single secondary branches, which depart from the nodes, only rarely perpendicular to the central axis37,38,39. Thanks to the well-preserved morphology, the ancient candelabrum hair was interpreted as a Platanus sp. foliar trichome based on literature40 and our experimental reference, although mullein cannot be totally excluded. Dimensions, distance between nodes and the number of tapering secondary branches attached to the central axis of the microremain were like those of all plane species documented in literature40,41,42.

Plant microremains identified by light microscopy in dental calculus samples. Some of the images captured by optic microscopy were shown. Aggregate of Triticeae starch granules and relative polarized image (A); Fabaceae starch granule and relative polarized image (B); Pinaceae pollen grain (C); aggregate of Triticeae starch granules and relative polarized image (D); Cupressaceae pollen grain (E); Poaceae spontaneous group pollen (F); polyhedral starches of morphotype II (G,H); fragments of plant tissues (I–K); dendritic hair (L). The scale bar indicates 15 µm [45 µm for panel (L)]. Small flecks of calculus still attached to microremains can be observed in some panels.
This finding leads to consider some paleoenvironmental implications. Fossil pollen analysis has demonstrated that, during the Plio-Pleistocene, Platanaceae were present in the Upper Valdarno (Italy)43. For the Holocene, likely as a consequence of Pleistocene glaciations, fragmentary and scarce evidence of plane tree have been found in Spain and French Mediterranean coast; no record of Platanus sp. has hitherto been found in Italy44,45. This thermophylous taxon has reappeared later as an ornamental tree, providing shade, during Roman times46. As we applied rigorous decontamination protocols, the evidence of this ancient trichome, probably accidentally inhaled by CDD1, may testify the presence of Platanus sp. and humid environments in central Italy during the Neo-Chalcolithic period.
Starch granules
More than 70 starches were retrieved from calculus samples (Table 1). Some of them were found in an extraordinary state of preservation, likely due to intentional ingestion and/or accidental inhalation during the processing of starchy foods. These grains were clustered in three different morphological types, based on the morphometric parameters (i.e., shape, size, presence of lamellae and hilum, aggregation level, and other secondary features) evidenced by literature. They were described using the International Code for Starch Nomenclature47,48.
Morphotype I These starches were consistent with those of Triticeae Dumort. tribe and occurred in almost all samples, as the most copious group (Table 1; Fig. 2A,D). Some grains were still lodged together. The morphotype was characterised by a bimodal distribution, or rather co-presence of large and small granules. Occasionally, the morphology was not completely intact, probably due to chewing as well as grinding and/or cooking procedures. These starch grains were similar to those occurring in caryopses of cereals, such as Hordeum sp. L. and Triticum sp. L. In particular, the diagnostic starches were oval to sub-round in 2D shape (15–43 µm in length; 10–35 µm in width). They had a central and distinct hilum and, sometimes, no visible lamellae. The small granules (≤ 10 μm in diameter) were spherical in shape with a central hilum. Knowledge about the Neo-Eneolithic period in central Italy is characterized by discontinuous data. The archaeobotanical dataset available for Latium is still limited49 but information about cultivated and wild-collected plants from Casale del Dolce site exists. In fact, the carpological analysis previously conducted50 has identified several caryopses of barley and wheat, supporting our results. The recovery of these starch grains, in almost all samples, suggested that the use of cereals was common and probably frequent for Casale del Dolce people, even if it is quite difficult to correlate presence of plant remains in calculus and quantity of consumed food26. The hypothesis of cereal consumption for this community has been also proposed by stable isotope data. Isotopic values would suggest a subsistence economy based on a great intake of carbohydrates and a lifestyle characterized by a progressive agricultural exploitation, even more evident than other Eneolithic sites of central Italy6,51. Lastly, Triticeae starches have been also found in dental calculus from Grotta dello Scoglietto (southern Tuscany), for the same pre-historical period52.
Morphotype II A low number of starch granules with faceted shape, perpendicular extinction cross and, sometimes, evident central fissures was recovered from dental calculus (Table 1; Fig. 2G,H). The morphology appeared oval to polygon (2D) with centric hilum and fissures radiating from it. The most frequent size distribution length was 14–25 μm in length and 13–17 μm in width. This type of grains exists in seeds of grasses belonging to the Andropogoneae Dumort. and Paniceae R. Br. tribes, as shown in the modern reference material19. Since an overlap in size and shape occurs among starches of species related to these tribes, the identification of these plant remains is arduous at a lower taxonomic level. Sorghum sp. Moench (sorghum), Setaria sp. P. Beauv. (foxtail millet) and Panicum sp. L. (millet) can be considered as potential candidates. Unfortunately, no phytolith, which would have helped us in distinguishing between the species of Paniceae53, was detected. In addition, the lack of an isotopic signal specific for this type of consumption and the absence of relative carpological remains for the archaeological site of Casale del Dolce might be due to a limited usage of these plants. In fact, although several species of these genera were diffused in Italy, little is known about their employment. The archaeobotanical evidence of millets (i.e., Panicum sp. and Setaria sp.) from the Late Neolithic period has been discussed; however, their cultivation is certain during the Bronze and Iron Ages52,54,55. Recently, Accelerator Mass Spectrometry-datings of prehistoric charred broomcorn millet grains has pinpointed the earliest occurrence of Panicum miliaceum L. in Europe at the middle of the 2nd millennium BCE (Middle/Late Bronze Age)56.
Morphotype III Only one grain contributed to the third type of starch (Table 1; Fig. 2B). It appeared to be consistent with the Fabaceae family, probably Vicieae (Bronn) DC. tribe (e.g., vetches) for its oval to elongated (irregular) shape and kidney-like. The hilum was obscured and sunken, while the lamellae were not fully visible. The size was 42 μm in length and 30 μm in width. Data about pulses are scarce for this period. In northern Italy, a high variety of pulses was already present in the Neolithic57,58 but this starch grain would seem to be one of the few and unique evidence of consumption in central and southern Italy. As this finding refers to a single individual, certainly, it is not expected to provide an exhaustive image of the use of pulses for the period and region but its presence, together with the carpological remains of Fabaceae49,50, could attest plant protein consumption.
A single starch granule was not classified because missing diagnostic and distinguishable characteristics. Probably modification events, such as grinding process, cooking procedure in water and/or chewing, and exposure to alfa-amylase, altered its shape.
Pollen grains
Four calculus samples showed the presence of different pollen types (Table 1). In total, 49 grains were found. Three of them were detected in CDD2, 4, and 9 (Fig. 2C,EF), while the remaining ones (46), both in single and in aggregate form, were retrieved from only one individual (CDD7) (e.g., in Fig. 3). All palynomorphs were identified according to morphometric parameters described in literature and evidenced in the Palynological Database59 and the names of the pollen types refer to literature60,61,62.

Plant micro-remains detected by morphological analysis in the dental calculus of CDD7 sample. Representative images obtained by light microscopy analysis were shown. Aggregates of pollen and spores (A,B); Pinaceae and Cupressaceae pollen grains (C); Brassicaceae pollen grain (D); Pinaceae pollen grains (E,F); Cupressaceae pollen (G); Quercus deciduous pollen (H); Alchemilla type pollen (I); sporangium of Monylophyta (J). The scale bar indicates 15 µm. Small flecks of calculus still attached to microparticles can be observed in some panels.
In this paragraph we describe the pollen grains found in CDD2, 4, and 9 samples.
The ancient microremain embedded in sample CDD2 was apolar and medium in size (63 µm in diameter), showing a morphology which typically occurs in Poaceae63,64. The stenopalynous nature of such type of pollen (that is, uniform monoporate) makes its systematic identification difficult. Although a low taxonomic determination limits paleoecological inferences, the evidence of Poaceae pollen is usually interpreted as indicative of open grasslands65.
One ancient palynomorph displayed morphological traits consistent with Pinaceae (sample CDD4). It appeared as a bisaccate monad with an elliptic corpus and medium reticulation on bladders59,66,67. Including sacci, the dimension was 56 µm in equatorial view.
A non-saccate Cupressaceae-type pollen, instead, was found in sample CDD9. It appeared spherical (with polar and equatorial axes of 30 µm) and inaperturate at light microscope; the protoplast exhibited itself star-like. Pollen grains produced by several species of Cupressaceae are considered morphologically uniform68. Since prehistoric times, Gymnosperm wood has been widely used as raw material and firewood, while needles, nuts and inner bark represented the edible parts of these trees69. Noteworthy is that the resins of these plants, possessing adhesive qualities and antibacterial properties, might have been also appreciated by Neanderthal14. Cupressaceae pollen grain is generally scarce in ancient sediments and one of the most underrepresented palynomorph in archaeological context. Several archaeobotanical studies have demonstrated the use of Juniperus L. species in the Mediterranean basin since the Holocene. In particular, the use of them as a source of aromatic foliage and resins employed for medicinal purposes, wood as fuel and for construction of dwellings, and fresh or dried berries as food has been proposed70,71,72. Sporadic fossil discoveries of Cupressus sp. L, instead, are rather sparse in the Mediterranean area, although some ancient record has been registered in Italy during the Quaternary73. Thus, the investigated plant microdebris testify the presence of Cupressaceae and provide additional evidence about the possible existence of evergreen Mediterranean forests, during the Neo-Chalcolithic period, in the Sacco River Valley.
Pollen grains in CDD7
CDD7 specimen (Fig. 1B), an adult male affected by severe malocclusion, preserved an interesting set of microparticles at microscopic analysis; therefore, we decided to report and discuss separately the data obtained from his calculus.
Eleven pollen grains out of 46 were not distinguishable due to the lack of diagnostic characteristics. The remaining 35 were found (singly, in pairs, or aggregates; Table 1, Fig. 3) in good or excellent state of conservation. The latter appeared as clusters of Pinaceae pollen (Gymnosperm) and other palynomorphs, including spores. Examples are shown in panels A and B of Fig. 3.
Two Cupressaceae, ten Pinaceae and one Poaceae pollen, presenting the same morphological features described in the previous paragraph, were also found in this sample (e.g., see Fig. 3C,E,F,G).
In addition, pollen grains from four herbaceous plants, namely Cyperaceae, Urticaceae, Trifolium, and Alchemilla species, and from the arboreal genus Corylus L. were detected and aredescribed below. Although pollen morphological variation within Cyperoideae subfamily is notable, one ancient microremain, possessing a pear-shape and a scabrate sculpture on its surface, appeared belonging to the genus Carex74,75. In equatorial view it was triangular and the polar axis length was 41 µm. A second pollen grain was recognised as Urticaceae-type; it exhibited spheroidal shape (equatorial diameter 23 μm) and scabrate ornamentation. This morphology occurs both in Parietaria sp. and Urtica sp. pollen grains59,62 and it is very difficult to distinguish them by optical microscope, especially if degraded. The shape of a third ancient monad, attributed to Trifolium-type (Fabaceae), was subprolate in equatorial view (46 μm) with scabrate ornamentation76. The Alchemilla-type (Rosaceae) microremain (26 μm equatorial view, Fig. 3I) was radially symmetrical, elliptic and prolate in shape77. Finally, another pollen type was found and attributable to Corylus sp. L. (Betulaceae). It was oval in equatorial view (19 μm), smooth, and tripolar with deep oncusis in each pore78.
Seven pollen grains were single, prolate, isopolar, and elliptic in equatorial view (polar axis 19–25 µm long). They were tricolpate, with long and narrow colpi. Pores were at times indistinct. Pollen of the different species of Fagaceae shows a high variability in form, size, sculpturing; for this reason, most of them overlap in morphology. The ancient palynomorphs in exam were closely similar to a Quercus-type (examples in Fig. 3A,H)79,80.
The last 10 grains showed a morphology (3-colpate, reticulate and subprolate) ascribable to Brassicaceae pollen grains (example in Fig. 3D). This is a stenopalynous family in which pollen varies among the genera but rarely in the species under the same genus81,82.
Intriguingly, pollen findings in sample CDD7 were numerous and deriving also from insect-pollinated plants (e.g., Brassicaceae). This evidence appeared like a honey palynospectrum. This type of assemblage has been never registered in dental calculus deposits and, especially for the aggregates, the hypothesis of accidental inhalation seems implausible. Precisely, the presence of aggregates induced us to reflect upon a common origin of the whole pollen record. However, for single granules, to date, the supposition of aspiration cannot be completely excluded, due to the multiple pathways of inclusion of such type of microparticles27. The high pollen variety could be explained by the presence of residues of natural matrices, as well as honey or beehive products (e.g., wax, propolis), in the calculus sample. To support our hypothesis, we prepared a reference collection based on modern multifloral honey samples (Supplementary Information 1, panel E–J).
Archaeological finds of bee products are quite rare83,84,85,86,87,88. Since the end of the upper Palaeolithic, honey has been employed as sweetener, while beeswax for technological, ritual, cosmetic and medicinal applications89,90. Regarding the latter, Bernardini et al.91 have found fascinating traces of a filling with beeswax, highlighting Neolithic dentistry procedures. It is important to recall that bees may also visit non-nectariferous plants (e.g., Poaceae, Betulaceae like Corylus sp.) for collecting pollen as protein source. Moreover, Pinaceae (Pinus sp. L. and Abies sp. Mill.) and Fagaceae (Fagus sp. L. and Quercus sp. L.), among others, emit sweet secretions and may be classified as honeydew producers88. Therefore, it is not unlikely to discover pollen grains of pine, hazel, oak, and cereals mixed with melliferous taxa. In fact, similarly, Carboni et al.92 have observed a lump of pollen inside an Eneolithic vessel, suggesting the use of a fermented honey-based drink, the mead, for ritual purposes.
According to all this evidence, the pollen record detected in the present ancient calculus could be likely interpreted as direct honey consumption and/or remain of food or beverage including honey as natural sweetener. However, the use of conifer resins as antimicrobial or flavouring agents, mixed to honey or alone, cannot be excluded, together with the hypothesis of inhalation of bisaccate pollen from the immediate environment.
Unfortunately, for the investigated site, no evidence supporting the previous hypotheses exists. Nevertheless, it is possible that the individuals from Casale del Dolce practised bee-keeping culture near woodland pastures, although this interpretation cannot be definitive.
Currently, pollen spectra from beehive products are used to deduce plant biodiversity of the areas visited by insects for nectar collection93,94. Bearing in mind this indication and the typical habitats of the identified plant taxa, some ecological implications were inferred. A thermophilic broad-leaved forest mainly made up of conifers (such as Pinus) and several deciduous trees (such as Quercus and Corylus), together with wet grasslands (Cyperaceae, Urticaceae, Alchemilla sp.), was outlined by pollen analysis. This hypothesis would seem consistent with Coubray’s work50, who has identified the wood charcoals found in the archaeological site of Casale del Dolce as Carpinus L., Quercus, Maloideae, Cornus L., Corylus, Ulmus L., Fraxinus L., and Acer L. remains. In addition, palynological analyses performed in the same region95,96,97 have detected similar vegetational elements.
Other plant microremains
We detected an unusual range of microparticles, that is, fragments of plant tissues and a sporangium, rarely documented in human dental calculus investigations (Table 1)69,98,99,100.
Among the first, one microparticle was made up of plant cells associated to a scalariform xylem vessel (Fig. 2I), while another debris showed wood cells with simple pits (Fig. 2J). A brown-yellowish fragment was also photographed (Fig. 2K). As reported in literature99, no evidence of charring or burning may be attributed to this type of darkening colouring but, if so, it would suggest an involuntary inhalation of ash particles from trees or shrubs used for fire. Thus, this type of microremain could derive from both non-edible and edible plants. In general, all these fragments retrieved from calculus might be the result of some activities, such as chewing of fresh plant organs, food and/or other uses of bark, oral hygiene procedures with woody dental picks, and/or use of teeth as a third hand99,101,102.
The second type of uncommon microparticle, found in sample CDD7 (Table 1), appeared morphologically like a sporangium, probably from Monylophyta (Fig. 3J). It was brownish in colour and ovoid in shape. This type of microremain has never been observed in so ancient human dental calculus. A more specific taxonomical identification is very complex and would be risky, since at palaeobotanical and/or archaeological level there is no evidence to support this finding. However, considering that sporangia are typically attached to the abaxial surface of the leaf and that airborne dispersal capability of fern spores into stronger wind currents is rare and improbable100,103, the recovery of the whole sporangium allowed us to hypothesize a voluntary use of fern leaves.
Biochemical analysis
GC–MS approach revealed the presence of organic compounds derived from the matter ingested and/or inhaled by the individuals. However, the potential of the biomolecular approach on dental calculus is still highly challenging and the capacity to trace the origin of some molecules is still difficult, due to the multifactorial dental calculus’s aetiology31,104.
In Supplementary Information 2 (SI2), the molecules detected in each sample were listed and clustered in chemical classes. The chromatographic profiles were dominated by a series of C6 to C30 n-alkenes and n-alkanes, not reported in SI2 because ubiquitous and not taxonomically specific. They could probably come from degradation of oral bacteria or consumed food, representing, for instance, fragments of unsaturated or saturated lipids14,105,106,107.
The typology of residues accumulated in dental calculus and their adsorption capacity determine the lipid profile of this matrix, considering that different foods naturally possess variable lipid composition. For this reason, it is difficult to associate fatty acids to specific dietary sources. The presence of fatty acids (e.g., odd, short, and long chains), ubiquitous components of organic matter, could be considered indicator for consumption of animal fats or plant oils (e.g., oil-rich seeds and fruits)104,108,109,110,111,112,113. Long-chained polyunsaturated fatty acid derivatives (PUFAs; e.g., eicosapentaenoic acid, EPA), abundant in dried fruits114, were detected in some samples. Polyunsaturated omega-3 fatty acids have been rarely identified in archaeological contexts115, due to their highly inclination to oxidative alteration116. However, dental calculus has shown itself conservative for this type of molecules31. The consumption of aquatic organisms cannot be excluded, being rich of PUFAs114 and considering the proximity of the ancient settlement to the Sacco River.
Monoterpene derivatives, non-specific compounds with volatile nature, retrieved from some samples, such as citronellol, menthol and pinanol (commonly found in leaves, fruit, and bark of a wide range of plant species), could generically indicate the ingestion of plant materials or waxes109.
In CDD5 calculus, azulene and coumarin derivatives were also recovered. These secondary metabolites usually occur in species belonging to Apiaceae, Asteraceae, and Rutaceae families, well known medicinal plants possessing a wide range of biological activities117,118. As suggested by Hardy et al.14, the plant species rich in such type of bitter-tasting compounds might have been ingested for self-medication.
Two alkaloids were found: trigonelline and hordenine, respectively, in CDD4 and CDD7 specimens. The first one, whose accumulation takes place in various plant species (i.e., Achillea sp. L.) and especially in Fabaceae seeds (e.g., Trigonella sp. L., Trifolium sp. L., and Medicago sp. L.)119,120, might represents a further proof for consumption of pulses.
Hordenine, which naturally occurs in certain grasses, like cereals (e.g., barley, millet, and sorghum)121, could demonstrate the ingestion of starchy material, as already testified by the detection of a Triticeae starch granule in the same calculus flakes and the recovery of caryopses at the site50.
Conclusions
To date, the complex correlation between ancient dental calculus and environmental flora biodiversity has not been fully understood and exploited yet. Thus, the main goal of the current study was the expansion of this issue. The high pollen variety observed in our pre-historic calculus samples pushes forward this field of research, allowing to hypothesize that the identified species were of local origin. These data would permit us to deduce some floristic-vegetational implications during the Neolithic and Chalcolithic period in central Italy. The simultaneous presence of traces of Poaceae and Fabaceae outlines a very important combination from a nutritional point of view, since cereals, rich in carbohydrates, are complementary to legumes, rich in proteins, and together creating a balanced and complete diet. Moreover, combining morphological approach with analytical chemistry, we provided an extensive body of evidence about the strong human–environment relationship of Casale del Dolce site. Although restricted to a limited series of samples, dental calculus analysis allowed us to confirm that these prehistoric individuals were comfortable with plants and able to exploit natural matrixes, offering the opportunity to maximize the biographical detail of this ancient population. To date, dental calculus represents one of the best archaeo-anthropological tools to highlight the value of plants in human evolution and culture, although it is still difficult to provide a univocal interpretation for some microremains embedded in it. The application of Next Generation Sequencing analysis on the ancient plant DNA extracted from this matrix could open up new and interesting interpretative scenarios.
Methods
Calculus sampling and analytical procedures
Twelve Neo-Eneolithic human skeletal remains (radiocarbon calibrated dated at 4.927 year. BP., Beta Analytic), hosted and preserved at the storerooms of the Villa Adriana and Villa d' Este (VA.VE.) (located in the Archaeological area of Santuario di Ercole Vincitore, Tivoli, Rome), were examined for dental calculus. Light deposits were detected above the gingival margin and removed from tooth enamel by an autoclaved dental pick. The mineralized flakes were collected separately on an aluminium foil and placed in sterilized micro-centrifuge tubes. The samples were then transferred to the Department of Biology of the University of Rome ‘Tor Vergata’ (Italy) for analysis.
Meticulous sterilisation and decontamination procedures were performed following our lab standard methods31. Here, laboratory contamination checks were regularly carried out on all workspaces, and supplies. Fresh disposable consumables (e.g., centrifuge tubes) and instruments (e.g., glassware, microscope slides, cover slip, metal tools) were autoclaved for 2 h, immediately prior to use. The absence of plant micro-residues in laboratory reagents and materials was also monitored. This intensive cleaning regime was applied with the same accuracy already reported in literature20,31,122,123.
Before proceeding with the preparation of dental calculus sample, the soil still adhering to the external part of the mineralized plaque was gently removed using a stereomicroscope (Leica ZOOM 2000, Leica, Buffalo, NY, USA) and a fine sterile acupuncture needle, under a sterile vertical laminar flow hood31. Afterward, calculus was treated by UV light for 10 min, immersed in 2% sodium hydroxide for 15 min, washed twice with sterilised water and dried at 37 °C. Preceding the cleaning procedure, six randomly selected dental calculi were washed by sterile water, which was examined by optical microscopy, to confirm the efficacy of the method. No microdebris was detected by light microscopy in the samples after decontamination.
To obtain the highest amount of information from these samples, the protocols used in our lab20,31,122,123 were applied, after some minor modification. For each individual, the mineralized plaque retrieved from different teeth was combined into a single tube where, after the decontamination procedures, 0.5 mL of 0.2 M hydrochloric acid was added and left to act for 24 h. Once solubilized, the sample was incubated with 0.5 mL of hexane, in agitation, for two hours. After centrifugation at 11,000g for 5 min, the supernatant fraction was recovered, dried out and, later, subjected to gas-chromatographic mass-spectrometry (henceforth GC–MS) analysis (see details below). The pellet resulting from the decalcification step was used to extract microparticles and perform the morphological analysis by optical microscopy (henceforth OM). In detail, this pellet was washed three times with ultrapure water and mounted on a glass slide in a water-glycerol solution (1:1, v/v). The sample was observed through an optical microscope (ZEISS Axio Observer 7, Zeiss, Jena, Germany) equipped with polarized filters and Zen imaging software 2.6, operating at different magnifications. The recovered microdebris were identified on morphometric features, in comparison to literature and our modern reference materials19,48,124. The starches were described using the International Code for Starch Nomenclature47.
GC–MS analysis was performed using a QP2010 system (Shimadzu, Kyoto, Japan), in triplicate on each dental calculus sample. The pellet obtained from the hexane fraction was resuspended in 60 µL of hexane and derivatized with 40 µL of Methyl-8-Reagent (Thermo Scientific, Bellefonte, PA, USA). Two microliters of extract were injected into the instrument at the temperature of 280 °C, in splitless modality. The sample was separated by an SH-Rtx-5MS capillary column (Shimadzu; length 30 m × diameter 0.25 mm × thickness 0.25 μm). The carrier gas was helium, employed at a constant flow of 1 mL/min. The temperature gradient was set as follows: 60 °C for 5 min, 150 °C for 5 min (reached at a rate of 6 °C/min), 250 °C for 5 min (reached at a rate of 6 °C/min), and 300 °C for 5 min (reached at a rate of 6 °C/min); to obtain a better resolution. An electron impact of 70 eV (scanning from 100 to 700 m/z) was used for the ionization (ion source temperature 230 °C; interface temperature 320 °C; solvent cut time 6 min). The detected molecules were identified by comparing their mass spectra with those registered in the software database NIST (National Institute of Standards and Technology) Library 14 and on-line support125. Scientific food databases and literature data were consulted for reconstructing food categories and plant species.
Experimental reference
Some of the microdebris (i.e., trichomes, pollen grains) found in the dental calculus were compared to a reference collection composed of plant microremains extracted from modern species and honeys. The reference collection was prepared in a separated laboratory. Trichomes of Verbascum sinuatum L. and Platanus sp. L. were collected with a drop of ultrapure water, mounted as previously described for calculus samples, and observed at light microscope. Pollen residue from honey was obtained by resuspending the bee product (5 g) in ultrapure sterilized water (15 mL) and centrifuging it at 7830g for 10 min. This procedure was repeated twice, and the final pellet was dissolved in fuchsine and analysed by OM.
Abbreviations
- GC–MS:
-
Gas chromatography mass spectrometry
- OM:
-
Optic microscopy
References
Radi, G. & Pannocchia, C. P. The beginning of the Neolithic era in Central Italy. Quat. Int. 470, 270–284 (2018).
Rubini, M. Considerazioni preliminari sui rinvenimenti antropo-logici di Cisterna di Latina. Problematiche di popolamento umano durante il neolitico nel territorio veliterno. Museo e Territorio. Velletri, Atti della I e II Giornata degli Studi “Il territorio veliterno nell’antichità”, 23–30 (2003).
Mallegni, F. & Rubini, M. Recupero dei materiali scheletrici umani in archeologia (CISU, 1994).
Rubini, M., Zaio, P. & Mogliazza, S. L’Eneolitico nell’Italia centro-meridionale In Il fenomeno antropologico della facies del Gaudo. IIPP, XLIII Riunione Scientifica—L’età del rame in Italia 409-414 (2011).
Zarattini, A. & Petrassi, L. Casale del Dolce. Ambiente, economia e cultura di una comunità preistorica della Valle del Sacco (TAV Treno Alta Velocità, 1997).
Martinez-Labarga, C. et al. Palaeodiet in Central and Southern Italy from Upper Palaeolithic to Eneolithic. In Biological and Cultural Heritage of the Central-Southern Italian Population Through 30 Thousand Years: EPIC (Eredità della Popolazione dell’Italia Centro-meridionale) (eds Rickards, O. & Sarti, L.) 137–152 (UniversItalia, 2016).
De Angelis, F. et al. Exploring mobility in Italian Neolithic and Copper Age communities. Sci. Rep. 11, 1–14 (2011).
Manfredini, A. & Muntoni, I. Gli spazi del vivere: funzioni e cronologia delle strutture d’abitato dell’insediamento neolitico di Casale del Dolce (Anagni—FR). Atti Riunione Scientifica IIPP 35, 187–198 (2003).
Henry, A. G., Brooks, A. S. & Piperno, D. R. Plant foods and the dietary ecology of Neanderthals and early modern humans. J. Hum. Evol. 69, 44–54 (2014).
Morales, J. et al. The introduction of South-Western Asian domesticated plants in North-Western Africa: An archaeobotanical contribution from Neolithic Morocco. Quat. Int. 412, 96–109 (2016).
Cristiani, E. et al. Dental calculus and isotopes provide direct evidence of fish and plant consumption in Mesolithic Mediterranean. Sci. Rep. 8, 8147 (2018).
Goude, G. et al. New insights on Neolithic food and mobility patterns in Mediterranean coastal populations. Am. J. Phys. Anthropol. 173, 218–235 (2020).
Mercuri, A. M. et al. From influence to impact: The multifunctional land use in Mediterranean prehistory emerging from palynology of archaeological sites (8.0–2.8 ka BP). Holocene 29, 830–846 (2019).
Hardy, K. et al. Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus. Naturwissenschaften 99, 617–626 (2012).
Cristiani, E., Radini, A., Edinborough, M. & Borić, D. Dental calculus reveals Mesolithic foragers in the Balkans consumed domesticated plant foods. PNAS 113, 10298–10303 (2016).
Hendy, J. et al. Proteomic evidence of dietary sources in ancient dental calculus. Proc. R. Soc. B 285, 20180977 (2018).
Juhola, T., Henry, A. G., Kirkinen, T., Laakkonen, J. & Väliranta, M. Phytoliths, parasites, fibers, and feathers from dental calculus and sediment from Iron Age Luistari cemetery, Finland. Quat. Sci. Rev. 222, 105888 (2019).
Schrader, S. Examining diet and foodways via human remains In Activity, Diet and Social Practice 127–164 (Springer, 2019).
Henry, A. G. Handbook for the Analysis of Micro-Particles in Archaeological Samples (ed. Herny, A. G.) 1-304; https://doi.org/10.1007/978-3-030-42622-4 (Springer, 2020).
Gismondi, A. et al. Back to the roots: Dental calculus analysis of the first documented case of coeliac disease. Archaeol. Anthropol. Sci. 12, 1–10 (2020).
Sawafuji, R., Saso, A., Suda, W., Hattori, M. & Ueda, S. Ancient DNA analysis of food remains in human dental calculus from the Edo period, Japan. PLoS ONE 15, e0226654 (2020).
Scott, A. et al. Exotic foods reveal contact between South Asia and the Near East during the second millennium BCE. PNAS https://doi.org/10.1073/pnas.2014956117 (2020).
Tromp, M. et al. Exploitation and utilization of tropical rainforests indicated in dental calculus of ancient Oceanic Lapita culture colonists. Nat. Hum. Behav. 4, 489–495 (2020).
Crowther, A., Haslam, M., Oakden, N., Walde, D. & Mercader, J. Documenting contamination in ancient starch laboratories. J. Archaeol. Sci. 49, 90–104 (2014).
Soto, M. et al. Structural characterization and decontamination of dental calculus for ancient starch research. Archaeol. Anthrop. Sci. 11, 4847–48721 (2019).
Leonard, C., Vashro, L., O’Connell, J. F. & Henry, A. G. Plant microremains in dental calculus as a record of plant consumption: A test with Twe forager-horticulturalists. J. Archaeol. Sci. Rep. 2, 449–457 (2015).
Radini, A., Nikita, E., Buckley, S., Copeland, L. & Hardy, K. Beyond food: The multiple pathways for inclusion of materials into ancient dental calculus. Am. J. Phys. Anthropol. 162, 71–83 (2017).
Krings, M., Taylor, T. N. & Taylor, E. L. Structural diversity and spatial arrangement of trichomes in a Carboniferous seed fern In Vistas in Palaeobotany and Plant Morphology: Evolutionary and Environmental Perspectives. (ed. Pant Memorial, D. D.) 61–69 (UP Offset, 2004).
Erdei, B. & Wilde, V. The Eocene flora of Tatabánya “marl-mine” (N Hungary). Palaeobiodivers. Palaeoenviron. 100, 1–17 (2020).
Lippi, M. M. et al. Plant remains in an Etruscan-Roman well at Cetamura del Chianti, Italy. Archaeol. Anthropol. Sci. 12, 1–18 (2020).
D’Agostino, A. et al. Investigating Plant Micro-Remains Embedded in Dental Calculus of the Phoenician Inhabitants of Motya (Sicily, Italy). Plants 9, 1395 (2020).
Radford, A. E., Dickison, W. C., Massey, J. R. & Bell, C. R. Vascular Plant Systematics (Harper & Row, 1974).
Pérez-de la Fuente, R. et al. Early evolution and ecology of camouflage in insects. PNAS 109, 21414–21419 (2012).
Simpson, M. G. Phylogeny and classification of the Haemodoraceae. Ann. Missouri Bot. Gard. 77, 722–784 (1990).
Zomlefer, W. B., Whitten, W. M., Williams, N. H. & Judd, W. S. An overview of Veratrum sp (Liliales: Melanthiaceae) and an infrageneric phylogeny based on ITS sequence data. Syst. Bot. 28, 250–269 (2003).
Dickison, W. C. Trichomes. In Integrative Plant Anatomy (ed. Dickison, W. C.) (Academic Press, 2000).
Grabias, B., Świątek, L. & Świętosławski, J. The morphology of hairs in Verbascum L. species. Acta Soc. Bot. Pol. 60, 191–208 (1991).
Alan, S., Saltan, F. Z., Gktrk, R. S. & Skmen, M. Taxonomical properties of three Verbascum L. species and their antioxidant activities. As. J. Chem. 21, 5438 (2009).
Cornara, L. et al. Smart drugs: Green shuttle or real drug?. Int. J. Legal Med. 127, 1109–1123 (2013).
Corujo, V. L. F., Basilio, M. A. & Galati, B. G. Analysis of digestive tract content of the larvae of Polybia scutellaris (White) (Hymenoptera, Vespidae). Rev. Bras. Entomol. 55, 241–246 (2011).
Nixon, K. C. & Poole, J. M. Revision of the Mexican and Guatemalan species of Platanus (Platanaceae). Lundellia 6, 103–137 (2003).
Carpenter, R. J., Hill, R. S. & Jordan, G. J. Leaf cuticular morphology links Platanaceae and Proteaceae. Int. J. Plant Sci. 166, 843–855 (2005).
Bertini, A., Magi, M., Mazza, P. P. & Fauquette, S. Impact of short-term climatic events on latest Pliocene land settings and communities in Central Italy (Upper Valdarno basin). Quat. Int. 225, 92–105 (2010).
Bottema, S. The Holocene history of walnut, sweetchestnut, manna-ash and plane tree in the Eastern Mediterranean. Pallas 52, 35–59 (2000).
Rinaldi, R. et al. The influence of a relict distribution on genetic structure and variation in the Mediterranean tree, Platanus orientalis. AoB Plants 11, plz002 (2019).
Yilmaz, H., Akkemik, Ü. & Karagöz, Ş. Identification of plant figures on stone statues and sarcophaguses and their symbols: The Hellenistic and roman periods of the eastern Mediterranean basin in the Istanbul Archaeology Museum. Mediterr. Archaeol. Archaeom. 13, 135–145 (2013).
ICSN. The international code for starch nomenclature http://fossilfarm.org/ICSN/Code.html (2011). Accessed 15 June 2020.
Gismondi, A. et al. Starch granules: A data collection of 40 food species. Plant Biosyst. 153, 273–279 (2019).
Anzidei, A. P., et al. L’abitato eneolitico di Osteria del Curato-via Cinquefrondi: nuovi dati sulle facies archeologiche di Laterza e Ortucchio nel territorio di Roma In Atti della XL Riunione Scientifica dell’Instituto Italiano di Preistoria e Protostoria. Strategie di insediamento fra Lazio e Campania in età preistorica e protostorica, 477–508 (2007).
Coubray, S. Analisi preliminare dei macroresti vegetali. In Casale del Dolce: Ambiente Economia e Cultura di una comunità preistorica della Valle del Sacco (eds Zarattini, A. & Petrassi, L.) 273–281 (Roma, 1997).
Cianfanelli, A., et al. Valutazione delle abitudini alimentari di popolazioni eneolitiche dell'Italia centro-meridionale In Preistoria del Cibo. 50ma Riunione Scientifica dell’Istituto Italiano di Preistoria e protostoria. della Toscana, Soprintendenza Archeologia (2015).
Lippi, M. M., Pisaneschi, L., Sarti, L., Lari, M. & Moggi-Cecchi, J. Insights into the Copper-Bronze Age diet in central Italy: Plant microremains in dental calculus from Grotta dello Scoglietto (Southern Tuscany, Italy). J. Archaeol. Sci. Rep. 15, 30–39 (2017).
Lu, H. et al. Phytoliths analysis for the discrimination of foxtail millet (Setaria italica) and common millet (Panicum miliaceum). PLoS ONE 4, e4448 (2009).
Tafuri, M. A., Craig, O. E. & Canci, A. Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy. Am. J. Phys. Anthropol. 139, 146–153 (2009).
Ganzarolli, G., Alexander, M., Arnau, A. C. & Craig, O. E. Direct evidence from lipid residue analysis for the routine consumption of millet in Early Medieval Italy. J. Archaeol. Sci. 96, 124–130 (2018).
Filipović, D. et al. New AMS 14 C dates track the arrival and spread of broomcorn millet cultivation and agricultural change in prehistoric Europe. Sci. Rep. 10, 1–18 (2020).
Rottoli, M. & Castiglioni, E. Prehistory of plant growing and collecting in northern Italy, based on seed remains from the early Neolithic to the Chalcolithic (c. 5600–2100 cal BC). Veg. Hist. Archaeobot. 18, 91–103 (2009).
Marchesini, M., et al., Ambiente e agricoltura tra VI e IV millennio a.C. In Emilia Romagna. Atti della XLV Riunione Scientifica Istituto Italiano di Preistoria e Protostoria, Modena 26–31 ottobre 2010, Studi di Preistoria e Protostoria 3, Preistoriae Protostoria dell’Emilia Romagna 2016, 159–168 (2017).
PalDat. A palynological database (2000 onwards) https://www.paldat.org/ (2019). Accessed 19 June 2020.
Berglund, B. E. & Ralska-Jasiewiczowa, M. Pollen analysis and pollen diagrams. In Handbook of Holocene Palaeoecology and Palaeohydrology (ed. Berglund, B. E.) 455–484 (Wiley, 1986).
Faegri, K. & Iversen, J. Textbook of Pollen analysis, 4th ed. (eds. Faegri, K. et al.) (John Wiley and Sons-Chichester, 1989).
Moore, P. D., Webb, J. A. & Collinson, M. E. Pollen Analysis, 2nd ed. (Blackwell Scientific Publications, 1991).
Perveen, A. A contribution to the pollen morphology of family Gramineae. World Appl. Sci. J. 1, 60–65 (2006).
Perveen, A. N. J. U. M. & Qaiser, M. U. H. A. M. M. A. D. Pollen flora of Pakistan-LXIX. Poaceae. Pak. J. Bot. 44, 747–756 (2012).
Radaeski, J. N., Bauermann, S. G. & Pereira, A. B. Poaceae pollen from Southern Brazil: Distinguishing grasslands (campos) from forests by analyzing a diverse range of Poaceae species. Front. Plant Sci. 7, 1833 (2016).
Grímsson, F. & Zetter, R. Combined LM and SEM study of the middle Miocene (Sarmatian) palynoflora from the Lavanttal Basin, Austria: Part II. Pinophyta (Cupressaceae, Pinaceae and Sciadopityaceae). Grana 50, 262–310 (2011).
Martin, A. C. & Harvey, W. J. The Global Pollen Project: A new tool for pollen identification and the dissemination of physical reference collections. Methods Ecol. Evol. 8, 892–897 (2017).
Mohanty, R. P., Buchheim, M. A., Portman, R. & Levetin, E. Molecular and ultrastructural detection of plastids in Juniperus (Cupressaceae) pollen. Phytologia 98, 298–310 (2016).
Radini, A. et al. Neanderthals, trees and dental calculus: New evidence from El Sidrón. Antiquity 90, 290–301 (2016).
Borojević, K., Forenbaher, S., Kaiser, T. & Berna, F. Plant use at Grapčeva cave and in the eastern Adriatic Neolithic. J. Field Archaeol. 33, 279–303 (2008).
Martin, L., Jacomet, S. & Thiebault, S. Plant economy during the Neolithic in a mountain context: the case of “Le Chenet des Pierres” in the French Alps (Bozel-Savoie, France). Veg. Hist. Archaeobot. 17, 113–122 (2008).
Moser, D., Di Pasquale, G., Scarciglia, F. & Nelle, O. Holocene mountain forest changes in central Mediterranean: Soil charcoal data from the Sila Massif (Calabria, southern Italy). Quat. Int. 457, 113–130 (2017).
Bagnoli, F. et al. Is Cupressus sempervirens native in Italy? An answer from genetic and palaeobotanical data. Mol. Ecol. 18, 2276–2286 (2009).
Nagels, A. et al. Palynological diversity and major evolutionary trends in Cyperaceae. Plant Syst. Evol. 277, 117 (2009).
Wronska-Pilarek, D., Janyszek, M. & Jagodzinski, A. M. Pollen morphology of selected Central European species from subgenera Vignea and Carex (Carex, Cyperaceae) and its relation to taxonomy. Bot. J. Linn. Soc. 164, 422–439 (2010).
Koçyiğit, M., Keskin, M. & Daştan, T. Pollen morphology of some Trifolium species, which are favorite plants of honeybees in Istanbul. J. Fac. Pharm. Istanbul 43, 85–94 (2013).
Faghir, M. B., Attar, F., Shavvon, R. S. & Mehrmanesh, A. Pollen morphology of the genus Alchemilla L. (Rosaceae) in Iran. Turk. J. Bot. 39, 267–279 (2015).
Nikolaieva, N., Brindza, J., Garkava, K. & Ostrovsky, R. Pollen features of hazelnut (Corylus avellana L.) from different habitats. Mod. Phytomorphol. 6, 53–58 (2014).
Denk, T. & Tekleva, M. V. Pollen morphology and ultrastructure of Quercus with focus on Group Ilex (= Quercus Subgenus Heterobalanus (Oerst.) Menitsky): Implications for oak systematics and evolution. Grana 53, 255–282 (2014).
Grímsson, F. et al. Fagaceae pollen from the early Cenozoic of West Greenland: Revisiting Engler’s and Chaney’s Arcto-Tertiary hypotheses. Plant Syst. Evol. 301, 809–832 (2015).
Erdtman, G. Pollen Morphology and Plant Taxonomy: Angiosperms (Leiden,The Netherlands: Brill Archive, 1986).
Khalik, K. A., Maesen, L. J. G., Kopman, W. J. M. & Berg, R. G. Numerical taxonomic study of some tribes of Brassicaceae from Egypt. Plant Syst. Evol. 233, 207–221 (2002).
Rösch, M. Pollen analysis of the contents of excavated vessels—direct archaeobotanical evidence of beverages. Veg. Hist. Archaeobot. 14, 179–188 (2005).
Kvavadze, E. V. The use of fossilized honey for paleoecological reconstruction: A palynological study of archaeological material from Georgia. Paleontol. J. 40, 595–603 (2006).
Kvavadze, E., Gambashidze, I., Mindiashvili, G. & Gogochuri, G. The first find in southern Georgia of fossil honey from the Bronze Age, based on palynological data. Veg. Hist. Archaeobot. 16, 399–404 (2007).
Cristiani, E., Pedrotti, A. & Gialanella, S. Tradition and innovation between the Mesolithic and early Neolithic in the Adige Valley (northeast Italy). New data from a functional analysis of trapezes from the Gaban rock-shelter. Documenta Praehistorica 36, 191–205 (2009).
Roffet-Salque, M. et al. Widespread exploitation of the honeybee by early Neolithic farmers. Nature 527, 226–230 (2015).
Chichinadze, M., Kvavadze, E., Martkoplishvili, I. & Kacharava, D. Palynological evidence for the use of honey in funerary rites during the Classical Period at the Vani. Quat. Int. 507, 24–33 (2019).
Belli, R. et al. Red painted stones from Riparo Dalmeri: a methodological approach to palaeoclimatic reconstruction In Atti del IV Congresso Nazionale AIAR, Pisa, 1–3 Febbraio 2006 (ed. D’Amico, C.) 15–24 (Pàtron Editore Bologna, 2007).
Mayyas, A. S., Al-Qudah, M. A., Douglas, K. A. & Al-Ajlouny, F. K. Beeswax preserved in archaeological ceramics: Function and use. Ann. Fac. Arts. 40, 343–371 (2012).
Bernardini, F. et al. Beeswax as dental filling on a Neolithic human tooth. PLoS ONE 7, e44904 (2012).
Carboni, G. et al. Inebriarsi per l’aldilà: Bevande alcoliche nelle necropoli di facies Rinaldone e Gaudo dell’area romana In Atti della L Riunione Scientifica dell’Istituto Italiano di Preistoria e Protostoria (2015).
Satta, A. et al. Combination of beehive matrices analysis and ant biodiversity to study heavy metal pollution impact in a post-mining area (Sardinia, Italy). Environ. Sci. Pollut. Res. Int. 19, 3977–3988 (2012).
Di Marco, G. et al. Geographical, botanical and chemical profile of monofloral Italian honeys as food quality guarantee and territory brand. Plant Biosyst. 151, 450–463 (2017).
Bellotti, P. et al. Late Quaternary landscape evolution of the Tiber River delta plain (Central Italy): New evidence from pollen data, biostratigraphy and 14C dating. Zeitschrift fur Geomorphologie 51, 505–534 (2007).
Magri, D. & Sadori, L. Late Pleistocene and Holocene pollen stratigraphy at Lago di Vico, central Italy. Veg. Hist. Archaeobot. 8, 247–260 (1999).
Sadori, L., Jahns, S. & Peyron, O. Mid-Holocene vegetation history of the central Mediterranean. Holocene 21, 117–129 (2011).
Power, R. C., Salazar-García, D. C., Straus, L. G., Morales, M. R. G. & Henry, A. G. Microremains from El Mirón Cave human dental calculus suggest a mixed plant–animal subsistence economy during the Magdalenian in Northern Iberia. J. Archaeol. Sci. 60, 39–46 (2015).
Cummings, L. S., Yost, C. & Sołtysiak, A. Plant microfossils in human dental calculus from Nemrik 9, a Pre-Pottery Neolithic site in Northern Iraq. Archaeol. Anthropol. Sci. 10, 883–891 (2018).
Fiorin, E., Sáez, L. & Malgosa, A. Ferns as healing plants in medieval Mallorca, Spain? Evidence from human dental calculus. Int. J. Osteoarchaeol. 29, 82–90 (2019).
Minozzi, S., Manzi, G., Ricci, F., di Lernia, S. & Borgognini Tarli, S. M. Nonalimentary tooth use in prehistory: An example from early Holocene in Central Sahara (Uan Muhuggiag, Tadrart Acacus, Libya). Am. J. Phys. Anthropol. 120, 225–232 (2003).
Hardy, K. et al. Diet and environment 1.2 million years ago revealed through analysis of dental calculus from Europe’s oldest hominin at Sima del Elefante, Spain. Sci. Nat. 104, 1–5. https://doi.org/10.1007/s00114-016-1420-x (2016).
Conant, D. S. A radioisotope technique to measure spore dispersal of the tree fern Cyathea arborea Sm. Pollen Spores 20, 583–593 (1978).
Buckley, S., Usai, D., Jakob, T., Radini, A. & Hardy, K. Dental calculus reveals unique insights into food items, cooking and plant processing in prehistoric central Sudan. PLoS ONE 9, e100808 (2014).
Eglinton, G., Gonzalez, A. G., Hamilton, R. J. & Raphael, R. A. Hydrocarbon constituents of the wax coatings of plant leaves: A taxonomic survey. Phytochemistry 1, 89–102 (1962).
Buckley, S. A., Stott, A. W. & Evershed, R. P. Studies of organic residues from ancient Egyptian mummies using high temperature-gas chromatography-mass spectrometry and sequential thermal desorption-gas chromatography-mass spectrometry and pyrolysis-gas chromatography-mass spectrometry. Analyst. 124, 443–452 (1999).
Luong, S., Tocheri, M. W., Sutikna, T., Saptomo, E. W. & Roberts, R. G. Incorporating terpenes, monoterpenoids and alkanes into multiresidue organic biomarker analysis of archaeological stone artefacts from Liang Bua (Flores, Indonesia). J. Archaeol. Sci. Rep. 19, 189–199 (2018).
Evershed, R. P. Chemical composition of a bog body adipocere. Archaeometry 34, 253–265 (1992).
Eerkens, J. The preservation and identification of Piñon resins by GC-MS in pottery from the Western Great Basin. Archaeometry 44, 95–105 (2002).
Buonasera, T. Investigating the presence of ancient absorbed organic residues in groundstone using GC–MS and other analytical techniques: A residue study of several prehistoric milling tools from central California. J. Archaeol. Sci. 34, 1379–1390 (2007).
Baeten, J., Jervis, B., De Vos, D. & Waelkens, M. Molecular evidence for the mixing of meat, fish and vegetables in Anglo-Saxon coarseware from Hamwic, UK. Archaeometry 55, 1150–1174 (2013).
Kanthilatha, N. et al. Identification of preserved fatty acids in archaeological floor sediments from prehistoric sites at Ban NonWat and Nong Hua Raet in northeast Thailand using gas chromatography. J. Archaeol. Sci. 46, 353–362 (2014).
Luong, S. et al. Development and application of a comprehensive analytical workflow for the quantification of non-volatile low molecular weight lipids on archaeological stone tools. Anal. Met. 9, 4349–4362 (2017).
Swanson, D., Block, R. & Mousa, S. A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 3, 1–7 (2012).
Reber, E. A. & Hart, J. P. Pine resins and pottery sealing: Analysis of absorbed and visible pottery residues from central New York State. Archaeometry 50, 999–1017 (2008).
Garnier, N., Bernal-Casasola, D., Driard, C. & Pinto, I. V. Looking for ancient fish products through invisible biomolecular residues in the roman production vats from the atlantic. Coast J. Marit. Archaeol. 13, 285–328 (2018).
McKay, D. L. & Blumberg, J. B. A review of the bioactivity and potential health benefits of chamomile tea (Matricaria recutita L.). Phytother. Res. 20, 519–530 (2006).
Gliszczyńska, A. & Brodelius, P. E. Sesquiterpene coumarins. Phytochem. Rev. 11, 77–96 (2012).
Matsui, A., Yin, Y., Yamanaka, K., Iwasaki, M. & Ashihara, H. Metabolic fate of nicotinamide in higher plants. Physiol. Plant. 131, 191–200 (2007).
Al-Ezzy, R. M. & Al Anee, R. S. Ibrahim, assessments of immunological activity of Achillea millefolium methanolic extract on albino male mice. J. Pharm. Pharmacol. 6, 563–569 (2018).
Balmer, D., Flors, V., Glauser, G. & Mauch-Mani, B. Metabolomics of cereals under biotic stress: Current knowledge and techniques. Front. Plant Sci. 4, 82 (2013).
Gismondi, A. et al. Archaeobotanical record from dental calculus of a Roman individual affected by bilateral temporo-mandibular joint ankylosis. Quat. Int. https://doi.org/10.1016/j.quaint.2020.11.017 (2020).
D’Agostino, A. et al. Lifestyle of a Roman Imperial community: Ethnobotanical evidence from dental calculus of the Ager Curensis inhabitants. J. Ethnobiol. Ethnomed. 15, 62 (2019).
Gismondi, A. et al. Dental calculus reveals diet habits and medicinal plant use in the Early Medieval Italian population of Colonna. J. Archaeol. Sci. Rep. 20, 556–564 (2018).
NIST. https://www.sisweb.com/software/ms/nist.html (2017). Accessed 16 June 2020.
Acknowledgements
This research is part of the PhD thesis in Evolutionary Biology and Ecology of Dr. Alessia D’Agostino. The authors thank Dr. Andrea Verolino (Earth Observatory of Singapore, Singapore) who provided support with Google Earth Pro and revised the English version of the text.
Author information
Authors and Affiliations
Contributions
Conceptualization, A.G., A.D.A. and M.R.; Methodology, A.D.A. and G.D.M.; Formal Analysis, A.D.A. and G.D.M.; Investigation, A.D.A., S.M., E.R. and G.D.M.; Validation, A.G. and A.D.A.; Data Curation, A.D.A. and A.G.; Writing—Original Draft Preparation, A.D.A., M.R. and A.G.; Writing—Review & Editing, A.D.A., M.R., G.D.M., S.M., E.R., A.C. and A.G.; Supervision, A.C. and A.G.; Resources, M.R., A.G. and A.C.; Funding Acquisition, A.C. and A.G.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
D’Agostino, A., Di Marco, G., Rubini, M. et al. Environmental implications and evidence of natural products from dental calculi of a Neolithic–Chalcolithic community (central Italy). Sci Rep 11, 10665 (2021). https://doi.org/10.1038/s41598-021-89999-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-89999-3
- Springer Nature Limited
This article is cited by
-
Applications of some advanced sequencing, analytical, and computational approaches in medicinal plant research: a review
Molecular Biology Reports (2024)
-
Exploring prehistoric plant use by molecular analyses of Neolithic grave goods
Vegetation History and Archaeobotany (2023)
-
Neolithic dental calculi provide evidence for environmental proxies and consumption of wild edible fruits and herbs in central Apennines
Communications Biology (2022)
-
Dental calculus - oral health, forensic studies and archaeology: a review
British Dental Journal (2022)