
Abstract
Mediterranean pastures are experiencing strong changes in management, involving shifts from sheep to cattle-based livestock systems. The impacts of such shifts on biodiversity are still poorly understood. Here, we sought to contrast the grazing regime, vegetation structure, bird species richness and abundance, between sheep and cattle grazed parcels, to understand the mechanisms through which management decisions impact farmland birds. During spring 2019, we characterized livestock management, bird populations and sward structure in 23 cattle and 27 sheep grazed parcels. We used a Structural Equation Model to infer the direct and indirect effects of sheep and cattle grazing on birds. Although no effects were found on overall species richness, there were species-specific responses to sheep and cattle grazed systems. Grazing pressure (variable integrating stocking rate and the number of days in the parcel) had negative impacts on the prevalence/abundance of Zitting Cisticola, Corn Bunting and Little Bustard, either directly or indirectly, through the effects of grazing pressure on vegetation height. Animal density and vegetation cover had direct positive effects in Galerida spp. and Common Quail, respectively. Zitting Cisticola and Little Bustard also showed a direct response to livestock type. Our study emphasizes the importance of grazing pressure as a driver of negative impacts for bird populations in Mediterranean grasslands. Since the ongoing transition from sheep to cattle-based systems involves increases in stocking rate, and therefore potentially higher grazing pressure, we propose a policy change to cap the maximum allowed grazing pressure. At the landscape scale, a mix of sheep and cattle grazed fields would be beneficial for maintaining bird diversity.
Similar content being viewed by others

Introduction
The way grasslands are managed impacts their biodiversity and the potential for provisioning ecosystem service1,2. Understanding the mechanisms linking grassland management actions to biodiversity outcomes is therefore key to understand the impacts of existing or planned policies, and associated farmers’ decisions3.
Mediterranean grasslands are a stronghold for several farmland bird species of conservation concern4,5,6. The management of these farming systems has been changing in the last decades due to incentives from the European Union (EU) Common Agricultural Policy (CAP), which have been promoting the replacement of traditional dry cereal-based systems, including crop rotations and fallow land, by livestock-based systems associated to an increasing amount of permanent pastures6,7,8. The suitability of these pastures to farmland birds depends on management decisions including the livestock type, grazing pressure, timing of hay harvesting (when existing), or the application of fertilizers9,10. These will in turn impact on key drivers of bird populations, including vegetation structure, food resources, or disturbance levels1,9,11.
In the Iberian Peninsula, because of the persistence of CAP subsidies coupled to cattle (but less to sheep) production, and a highly subsidized beef-cattle production10, there has been a shift from sheep-based to cattle-based systems in these permanent pastures7,10,12. Previous studies have suggested that this management change is likely to differentially affect grassland birds through changes in sward structure and therefore habitat suitability for birds10,13, as well as increases in bird nest predation and trampling risk14.
Our overall aim was to contrast the implications and mechanisms of the cattle versus sheep management impacts on farmland bird populations in permanent pastures of the Iberian Peninsula. We focused on a High Nature Value region in Southern Portugal where the transition from traditional crop-based systems, which included sheep grazing in stubble fields and fallow land, towards livestock-based, mostly cattle, systems in permanent pastures has been occurring7,10. Previous studies in the area addressed the impact of field and landscape variables on bird populations, including grazing regimes, but were mostly focused on fallow parcels managed under a traditional farming system13 and did not address the mechanisms through which grazing impacted on birds. We sought to establish a connection between livestock management and bird populations through the characterization of the occurrence and density of breeding birds, vegetation structure and grazing pressure in a series of fields managed either for sheep or cattle. Subsequently, using a modelling approach, we explored the potential direct and indirect effects of livestock on birds’ density and occurrence. Direct effects of livestock management on birds were expected to occur either by differences in livestock type, resulting in behavioural differences with impacts on disturbance, trampling or nest predation9,14. Indirect effects were expected through impacts on vegetation, leading to changes in habitat quality and food resources that ultimately influence birds’ occurrence and density. Our main questions were: (a) how grazing regimes (animal density and duration of grazing) and vegetation structure differed between sheep versus cattle systems?; (b) how bird species richness, occurrence and density varied across livestock systems?; and, (c) what were the direct and indirect effects of livestock type on bird populations?
Methods
Study area and parcel selection
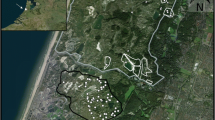
The study was conducted in Castro Verde Special Protection Area (SPA), located in southern Portugal (Fig. 1). The climate is Mediterranean, with hot summers (30–35 °C on average in July) and mild winters (averaging 5–8 °C in January), and over 75% of annual rainfall (500–600 mm) concentrated in October–March. The landscape is flat or gently undulating (100–300 m), mainly dominated by open areas used for rainfed pastures (ca. 60%) and annual crops (ca. 25%), and to a less extent by open woodlands (ca. 7%)15.

(a) Location of the study area within the Castro Verde Special Protected Area (SPA), southern Portugal. (b) Distribution of the 27 sheep (dark grey polygons) and 23 cattle (light grey polygons) grazing parcels and (c) Sampling scheme applied to each parcel surveyed. Bird counts were done at the centroid of the parcel (white dot) whereas vegetation sampling was performed at the indicated 10 points (black dots). The area covered with pastures and annual crops (derived from CORINE land cover 2018—https://land.copernicus.eu/pan-european/corine-land-cover/clc2018) is shown in yellow. The map was done using the version 3.10.0 of QGIS—https://qgis.org/en/site/index.html.
Since 1995, part of the study area has benefited from a CAP agri-environment aiming to protect the traditional farming system16. This scheme provides financial support to farmers for agricultural practices considered favourable to conservation, including the traditional rotation of cereals and fallows, the maintenance of low stocking rates (usually related with sheep grazing systems), and sowing of crops benefiting grassland birds16. However, in recent years the traditional farming system has been declining, with many farmers converting to specialized livestock systems, mainly, cattle grazing systems, with an increase of stocking rates7,15.
Parcel selection started by identifying grasslands grazed by either sheep or cattle, based on parcel-level statistical information from 2010 provided by the Portuguese Ministry of Agriculture7. To minimize potentially confounding effects of adjacent land uses (edge effects) and other non-crop elements within parcels on bird assemblages, we excluded parcels less than 100 m from shrubland or forested areas, with shrub and tree cover > 5% and with a minimum size of 10 ha. In January 2019 we visited 100 pre-selected parcels which were grazed by either sheep or cattle in 2010 in order to confirm the parcel land use in the agricultural year of 2018/2019, aiming to sample a balanced proportion of 50 sheep and cattle grazed parcels. Additional livestock information for the agricultural year of 2018/2019 was obtained during systematic visits to targeted parcels (see “Grazing Regime” section from Methods). We ended up with 23 cattle parcels and 27 sheep parcels (Fig. 1).
Bird and vegetation data
Breeding birds were sampled twice in each parcel during 7–16 April and 1–15 May 2019 respectively, always by the same observer (R.F.R). This was done to take into account species-specific breeding phenology in the area (early and late breeders)17 and minimize bias due to other factors (like weather or disturbance). Sampling was conducted using standardized 10 min point counts18 carried out at the central point of the parcel (Fig. 1). As the open terrain allowed for high visibility, a large detection radius was used, and all birds detected within 100 m of the central point were identified and counted. This radius is roughly similar to the one previously used for characterizing bird populations in the region19. All counts were carried out in the first four hours after sunrise and in the last two hours before sunset, with none in heavy or persistent rain, or in strong wind conditions. To estimate bird species richness and occurrences in each parcel, we pooled the data from the two counts. Species-level analyses focused on the six most common species, which occurred in > 30% of the parcels (see Supplementary Table S1). In addition to presence/absence, we also estimated population densities, using the count which yielded the highest estimate of density for each species (assuming this is the best indicator of population density, given the potential phenology and detectability biases above mentioned). Bird densities were based on the number of males simultaneously detected and expressed as breeding pairs/10 ha or males/10 ha (in the case of Little Bustard Tetrax tetrax and Common Quail Coturnix coturnix). Categorization to the genus level was made for the Crested and Thekla larks (Galerida cristata and G. theklae) due to difficulties in correctly identifying all individuals of these two very similar species in the field.
Vegetation height and cover were measured once in each parcel, between April 22 and May 6. Vegetation height was estimated in a set of ten 3 m radius plots defined inside the 100 m buffer (Fig. 1). In each plot, ten measurements of vegetation height were taken at random locations, for a total of 100 measurements per parcel. Vegetation height was measured using a 50 cm ruler and was defined as the highest point of vegetation projection within 3 cm of the ruler20. All values were estimated to the nearest half centimeter. When no vegetation was present (bare soil, soil litter, rocks or animal dung) the height was set to zero (0) but these measurements were not considered to estimate the mean height of the sward. Vegetation cover was measured inside a 50 × 50 cm quadrat placed at each of the ten grid points, by visual estimation to the nearest 5% of the percentage of the quadrat area covered by vegetation21 (Fig. 1). Vegetation height and cover measurements were averaged within each parcel.
Grazing regime
The number and type of livestock in each parcel as well as the extent of the grazing period since the start of the year (2019) were gathered from interviews (Supplementary Information S1) to land managers during 1–15 May 2019. This information was further validated, and corrected in a few cases, through field checks during regular visits (made at two-week intervals) to the parcels (see “Bird and vegetation data” section from Methods). Three grazing regime indicators were estimated for the whole period (January–May 2019): livestock type (either sheep or cattle), animal density, and grazing pressure. The animal density in each parcel was calculated as the average density (animals per hectare) of any species (regardless of being sheep or cattle) that grazed the parcel during the 5-months period. Stocking Rate translated animal density into livestock unit (LU) per hectare (LU/ha), between January and May, according to the following criteria: one adult bovine = 1 LU; bovine aged < 6 months = 0.4 LU; one adult sheep = 0.15 LU22. Using LUs allows the comparison of densities across livestock types after correcting for their relative feeding requirements23. Grazing Pressure was estimated as the Stocking Rate times the number of days a number of Livestock Units (LU) spent in a plot (LU/ha × number of days)24. The area used for these estimations corresponded to the available area where animals could freely roam, which in many cases was larger than the sampled parcel area, which was often not delimited by fences. The number of days in the parcel was collected mainly from the interviews. However, in some cases the extent of grazing period was expressed qualitatively and thus had to be inferred, from common expressions according to the following criteria: ‘few’ = 5 days, ‘some’ = 10 days, ‘a fortnight’ = 15 days, ‘many’ = 20 days, ‘almost all month’ = 25 days25.
Data analysis
Five explanatory variables describing grazing regime and sward structure (Table 1) were used as predictors of bird species richness, occurrence and abundance at parcel level. The correlation and multicollinearity between them were tested and all presented values of r < 0.70 and of variance inflation factor (VIF) smaller than 326.
Univariate differences between sheep and cattle parcels, both for bird response variables (species richness, density and occurrence) and for the explanatory variables, were tested using Generalized Linear Models (GLMs). We used a Gaussian error distribution and an identity link for quantitative variables, and a binomial error distribution and a logit-link function for occurrence data26.
A Structural Equation Modelling (SEM) approach was then used to investigate how grazing regime directly and indirectly affects the occurrence and density of birds. SEM are probabilistic models that hypothesize a causal network with multiple variables that can appear as both predictor and response variables27, allowing to look at both direct and indirect effects. We performed a confirmatory-exploratory path analysis28 in the form of a piecewise SEM conducted in the R software29, using the package “piecewiseSEM”27. In piecewise SEM the network is translated as a set of linear equations which can be evaluated individually, using R230. The goodness-of-fit of the entire model was quantified by a directed separation test (“d-separation test”), which tests the assumption that all variables are conditionally independent, i.e. that there are no missing relationships among unconnected variables27,30.
We started by building a theoretical model of our system (Fig. 2) based on previous literature and knowledge about birds and grasslands (See Supplementary Information S2 for more details on model construction). In short, the model states that the impacts of grazing regime on birds can occur: (A) indirectly, via the impacts of grazing pressure and potentially associated (non-measured) management decisions (e.g. fertilizer use or pasture improvement) on vegetation structure (vegetation height and cover); (B) directly, through the effect livestock-specific (sheep or cattle) behavior (trampling patterns, impacts of feeding mode on food resources for birds, potential egg predation) on birds; or, (C) directly through the disturbance impacts of animal density, expressed as number of herbivores spread over the area, irrespective of livestock type, on birds (Fig. 2). We considered all paths as significant if they had a p-value < 0.1. This threshold was used assuming it could indicate the existence of an effect, even if not significant at the traditional 0.05 level given the relatively low sample sizes. Other authors have used a similar approach in a SEM context (e.g. Sanz-Pérez et al.11). Moreover, we used the d-separation test from piecewise SEM output to evaluate our theoretical model and identify eventual significant paths not considered initially.

Theoretical model of the confirmatory-exploratory path analysis, where (A) represents the paths of the indirect effect of livestock type via impacts on vegetation structure; (B) represents the livestock type specific direct effects; (C) represents the direct effect of animal density through disturbance impacts on birds. For detailed information of model construction, see Supplementary Information S2.
After obtaining the final model for species richness and for the occurrence and density of each species, we estimated the standardized model parameters (expressed as mean ± standard error; SE) of causal effects. Effect estimates were used to calculate the strengths of direct and indirect effects between variables in the system. Indirect effects were described as a predictor variable (P1) having an effect on the response variable (R) through a simultaneous response and predictor variable (P2), P1 → P2 → R31. All statistical analyses were performed within “R” software environment, version 4.0.229.
Results
Grazing regime and vegetation structure
Animal density was significantly higher (GLM, p < 0.01) in sheep (4.8 ± 1.39 animals/ha) than cattle (0.8 ± 0.24 animals/ha) parcels (Fig. 3a). Grazing pressure was not significantly different between sheep and cattle parcels (GLM, p = 0.22) in spite of the trend for higher values in the latter (Fig. 3b). There were also no significant differences between livestock types regarding vegetation height (GLM, p = 0.61) and cover (GLM, p = 0.62) (Fig. 3c,d).

Comparison between sheep (dark grey) and cattle (light grey) grazed parcels for (a) Grazing pressure (LU * grazing days/ha); (b) Animal density (animals/ha); (c) Vegetation height (cm); (d) Vegetation cover (%). Values represent average and 95% confidence intervals (shown as vertical black lines). *Highlights significant differences.
Bird species richness and abundance patterns
A total of 25 bird species were observed in the studied parcels, with frequencies of occurrence ranging from a single parcel to all parcels (Supplementary Table S1). Ca. 70% of the species occurred in both livestock systems, and species registered in just one system type had low prevalence (see Supplementary Table S1 for details). The most prevalent species were Corn Bunting Emberiza calandra (100% of parcels), Galerida spp. (82%), Calandra Lark Melanocorypha calandra (82%), Common Quail (58%), Zitting Cisticola Cisticola juncidis (50%) and Little Bustard (46%). These six species occurred in both livestock systems and accounted for 86% of all the registers. Another eleven species were present in both cattle and sheep parcels (details in Supplementary Table S1).
Species richness was very similar between sheep (\(\overline{x }\) = 5.4 ± 0.45) and cattle (\(\overline{x }\) = 5.6 ± 0.53) parcels (GLM, p = 0.70) (Fig. 4a). There was a higher prevalence and abundance of Zitting Cisticola in sheep pastures (GLMdensity, p = 0.01; occurence, p < 0.01) and of Little Bustard in cattle parcels (GLMdensity, p = 0.03; occurence, p < 0.01) (Fig. 4b,c), while no significant univariate effects of livestock type were found for the other species.

Comparison between sheep (dark grey) and cattle (light grey) grazed parcels for (a) Species richness (average number of species in each parcel); (b) Occurrence of bird species (% occurrence); (c) Average density of bird species (pairs/10 ha or males/10 ha). 95% confidence intervals shown as vertical black lines. *Highlights significant differences.
Modelling the effects of grazing regime on birds
SEM results (Table 2) confirmed the higher animal densities in sheep parcels (β = − 0.5, p < 0.01). Also, we found a negative effect of grazing pressure on vegetation height (β = − 0.4, p = 0.01). As for response variables, there were no significant effects on species richness (Supplementary Fig. S2l), but there were several significant effects on individual species occurrences or abundances (Table 3).
Direct effects of livestock type included the positive association of Zitting Cisticola with sheep (βdensity = − 0.5, p < 0.01; βocurrence = − 0.6, p < 0.01) (Table 3 and Supplementary Fig. S2a,b), and the positive association of Little Bustard with cattle (βdensity = 0.5, p < 0.01; βocurrence = 0.6, p < 0.01) (Table 3 and Supplementary Fig. S2j,k). Animal density showed a positive effect on Galerida spp. density (β = 0.4, p = 0.05), but no significant effects on other species (Table 3; Supplementary Fig. S2f,g).
The only significant effect of vegetation cover was a positive relation with the prevalence of Common Quail (βocurrence = 0.4, p = 0.08) (Table 3 and Supplementary Fig. S2d). Half the species were influenced by vegetation height (Table 2), with positive effects on Zitting Cisticola (βdensity = 0.5, p < 0.01; βocurrence = 0.5, p = 0.02) and Corn Bunting (βdensity = 0.5, p < 0.01) (Table 3 and Supplementary Fig. S2e), and negative on the density of Calandra Lark (βdensity = − 0.3, p = 0.07) (Table 3 and Supplementary Fig. S2h). The Little Bustard was the only species directly affected (negatively) by increasing grazing pressure (βdensity = − 0.3, p = 0.04) (Table 3 and Supplementary Fig. S2j,k).
Discussion
Our results showed that the shift from sheep to cattle grazing systems in Mediterranean grasslands did not have major impacts on overall breeding bird species richness. However, there were some species-specific responses influenced by the type of livestock and animal density (both impacting stocking rates and grazing pressure) which were dependent on the livestock system. By analysing the direct and indirect paths through which management decisions are expected to impact on bird populations, we found that grazing pressure was a key driver of the observed responses, which has implications for policy recommendations.
Changes in grazing regime and vegetation structure
Our results corroborate previous findings that the transition from sheep to cattle grazing systems have clear implications for stocking rates8,12. However, an impact on grazing pressure was not observed. Although animal density was higher in sheep than in cattle parcels, grazing pressure tended to be similar, as the cattle feeding requirements are higher than that of sheep due to their larger size and associated nutritional requirements22.
Contrary to expectations, no direct effects of livestock type were observed for vegetation height and cover. Sheep and cattle have distinct diets and feeding behaviours32,33, with the former usually leading to shorter and uniform swards, and the latter usually promoting structural heterogeneity, with patches of lower and taller vegetation33,34. However, the fact that shorter swards were associated to higher grazing pressure, and the latter was tendentially higher in cattle parcels, might have minimized expected differences in vegetation structure.
Effects of grazing regime on birds
Overall, species richness was similar in sheep and cattle grazed parcels, with all but the least prevalent species occurring in both parcel types. This was expected, as both parcel types shared the same habitat and 68% of the species identified. It also suggests that the studied grazing regime changes are not likely to drastically change farmland bird assemblages at the regional level. Previous studies also did not find differences in bird species richness associated with different livestock management in grasslands10.
A direct effect of livestock type on birds was registered in 36% of the models (Fig. 5). Little Bustards’ positive relation with cattle grazed parcels may be related to food resources availability, mainly beetles and other invertebrates, which previous studies reported to be more abundant in cattle grazed fields35, and in more heterogeneous fields favouring the occurrence of both males and females36. Little Bustard preference for cattle pastures (Fig. 4) was also reported by Reino et al.13 for grazed fallow fields in the same region. Zitting Cisticola showed an opposite trend, with a positive response towards sheep grazed parcels. This finding is hard to explain as the species prefers taller swards20, which were not directly associated to any specific type of grazer. Other non-measured features might explain this result, such as the likely existence of more cereal fields, a preferred habitat for the species, in the vicinity of sheep parcels (because sheep are associated to the traditional system including cereals), the amount of hedges and tree lines or grazing rotations, previously found to influence the species’ abundance10.

Summary of SEM results for the six bird species density and occurrence, in a total of 11 SEMs. Thickness of the paths is proportional to the number of times that path was significant (< 0.1), regardless of the direction (positive or negative) of the effect.
Previous studies also reported a preference of Calandra Lark for cattle parcels10, a tendency (albeit non-significant) shown in our study. In contrast with our results, previous studies found associations of Corn Bunting to livestock type, although with contrasting patterns, some revealing preference for cattle13 and other for sheep10.
Positive effects of increasing vegetation height were observed for Zitting Cisticola and Corn Bunting. Both species are usually associated with cereal fields37 and tend to select fields with higher vegetation and a low proportion of bare ground20,38. This preference can be related to breeding and nesting sites availability. In contrast, Calandra Lark showed a negative relation to vegetation height, as previously documented for this species favouring bare soils20,39.
Higher grazing pressure was detrimental to Zitting Cisticola, Corn Bunting and Little Bustard, either direct or indirectly (through vegetation height). Grazing pressure is a key factor influencing species occurrence and breeding success in grassland birds, as it determines not only vegetation structure that provides cover and food availability, but also disturbance levels, which is particularly important during spring because of nest trampling10,14. The Little Bustard was directly influenced by grazing pressure, which is in line with previous studies showing its sensitivity to high grazing pressure, possibly because it increases disturbance and exposure to predation during the breeding season36. Moreover, male Little Bustards have very specific requirements of vegetation structure during the breeding season, preferring intermediate vegetation height (between 20 and 30 cm), which simultaneously provide concealment against threats and visibility for courtship40,41. As such, high grazing pressure likely prevents the development of a suitable sward structure for the species.
Other significant effects included a positive relation between vegetation cover and Common Quail occurrence, which is in line with previous studies suggesting that this species prefers dense and tall swards such as cereal fields37. Animal density had a positive effect on Galerida spp. density. The fact that we grouped two distinct species of Galerida larks (Crested and Thekla larks) can be a confounding factor while interpreting the results, since each species has different habitat requirements37. However, the former, usually associated with flat and human-disturbed areas, is much more abundant than the latter, which is more associated with shrublands in hilly areas42. This can help explain the positive effect of animal density (likely associated to higher disturbance) on Galerida spp.37.
Conclusions
Our study suggests that grazing pressure is the main path through which grazing regime has detrimental effects on several bird species in Mediterranean grasslands. Although we did not find a significant effect of livestock type on grazing pressure in our sampled parcels, other studies provide evidence that the ongoing transition from sheep to cattle systems in the region is reflected in a large increase in stocking rates with likely implications for grazing pressure8. This trend is caused by existing CAP subsidies keeping cattle payments partially or fully coupled7, meaning that financial support for farmers is proportional to the number of cattle they have. We therefore propose that this policy should be revised, decoupling the subsidies and implementing an area-based payment system limiting the maximum allowed grazing pressure.
Our results also suggest that at the landscape level, maintaining a mix of sheep and cattle grazed fields could be beneficial for maintaining bird diversity, since some species are more associated with a particular grazing regime.
Data availability
All supplementary information can be downloaded from the journal’s website.
References
Kipling, R. P. et al. Key challenges and priorities for modelling European grasslands under climate change. Sci. Total Environ. 566, 851–864 (2016).
Sollenberger, L. E., Kohmann, M. M., Dubeux, J. C. B. & Silveira, M. L. Grassland management affects delivery of regulating and supporting ecosystem services. Crop Sci. 59(2), 441–459 (2019).
Santos, J. L. et al. A farming systems approach to link agricultural policies to biodiversity and ecosystem services. Front. Ecol. Environ. https://doi.org/10.1002/fee.2292 (2020).
Suárez, F., Naveso, M. A. & De Juana, E. Farming in the drylands of Spain: birds of the pseudosteppes. In: Farming and Birds in Europe. The Common Agricultural Policy and its Implications for Bird Conservation 297–330 (Academic Press, 1997).
Delgado, A. & Moreira, F. Bird assemblages of an Iberian cereal steppe. Agric. Ecosyst. Environ. 78, 65–76 (2000).
Traba, J. & Morales, M. B. The decline of farmland birds in Spain is strongly associated to the loss of fallowland. Sci. Rep. 9, 1–6 (2019).
Ribeiro, P. F. et al. Modelling farming system dynamics in high nature value farmland under policy change. Agric. Ecosyst. Environ. 183, 138–144 (2014).
Marques, A. T. et al. Changes in grassland management and linear infrastructures associated to the decline of an endangered bird population. Sci. Rep. 10, 1–11 (2020).
Vickery, J. A. et al. The management of lowland neutral grasslands in Britain: Effects of agricultural practices on birds and their food resources. J. Appl. Ecol. 38, 647–664 (2001).
Faria, N. & Morales, M. B. Livestock species and grazing rotational patterns modulate grassland bird assemblages in Mediterranean drylands. Agric. Ecosyst. Environ. 295, 106893 (2020).
Sanz-Pérez, A. et al. Fallow management increases habitat suitability for endangered steppe bird species through changes in vegetation structure. J. Appl. Ecol. 56, 2166–2175 (2019).
Faria, N. Predicting agronomical and ecological effects of shifting from sheep to cattle grazing in highly dynamic Mediterranean dry grasslands. Land. Degrad. Dev. 30, 300–314 (2019).
Reino, L. et al. Effects of changed grazing regimes and habitat fragmentation on Mediterranean grassland birds. Agric. Ecosyst. Environ. 138, 27–34 (2010).
Beja, P. et al. Predators and livestock reduce bird nest survival in intensive Mediterranean farmland. Eur. J. Wildl. Res. 60, 249–258 (2014).
Ribeiro, P. F. et al. An applied farming systems approach to infer conservation-relevant agricultural practices for agri-environment policy design. Land Use Policy 58, 165–172 (2016).
Santana, J. et al. Mixed effects of long-term conservation investment in Natura 2000 farmland. Conserv. Lett. 7, 467–477 (2014).
Sutherland, W. J., Newton, I. & Green, R. Bird ecology and conservation: a handbook of techniques (Vol. 1) (OUP Oxford, 2004).
Fuller, R. J. & Langslow, D. R. Estimating numbers of birds by point counts: How long should counts last?. Bird Study 31, 195–202 (1984).
Moreira, F. et al. Population trends in the steppe birds of Castro Verde in the period 2006–2011: Consequences of a drought event and land use changes?. Airo 22, 79–89 (2012).
Moreira, F. Relationships between vegetation structure and breeding bird densities in fallow cereal steppes in Castro Verde, Portugal. Bird Study 46, 309–318 (1999).
Kent, M. & Coker, P. Vegetation description and analysis: A pratical approach (Wiley, 1992).
DRE. Anexo 2: Tabela de conversão em cabeças normais (CN). Diário da República, 1.ª série, 41: Portaria n.º 57 http://data.dre.pt/eli/port/57/(2015)/02/27/p/dre/pt/html (2015).
FAO. Guidelines for the Preparation of Livestock Sector Reviews (Animal Production and Health Guidelines. No. 5. Rome, 2011).
Gonçalves, P., Alcobia, S., Simões, L. & Santos-Reis, M. Effects of management options on mammal richness in a Mediterranean agro-silvo-pastoral system. Agrofor. Syst. 85, 383–395 (2012).
Redding, D. W. et al. Location-level processes drive the establishment of alien bird populations worldwide. Nature 571, 103–106 (2019).
Zuur, A. F., Leno, E. N., Walker, N. J., Saveliev, A. A. & Smith, G. M. Mixed Effects Models and Extensions in Ecology with R (Springer, 2009).
Lefcheck, J. S. piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 7, 573–579 (2016).
Grace, J. B. Structural Equation Modelling and Natural Systems (Cambridge University Press, 2006).
R Core Team. R: A language and environment for statistical computing (R Foundation for Statistical Computing, Vienna, 2020)
Shipley, B. Cause and Correlation in Biology: A User’s Guide to Path Analysis, Structural Equations and Causal Inference with R (Cambridge University Press, 2016).
Stenegren, M. et al. Piecewise structural equation model (SEM) disentangles the environmental conditions favoring diatom diazotroph associations (DDAs) in the western tropical North Atlantic (WTNA). Front. Microbiol. 8, 810 (2017).
Eldridge, D. J., Delgado-Baquerizo, M., Travers, S. K., Val, J. & Oliver, I. Do grazing intensity and herbivore type affect soil health? Insights from a semi-arid productivity gradient. J. Appl. Ecol. 54, 976–985 (2017).
Faria, N., Peco, B. & Carmona, C. P. Effects of haying on vegetation communities, taxonomic diversity and sward properties in Mediterranean dry grasslands: A preliminary assessment. Agric. Ecosyst. Environ. 251, 48–58 (2018).
Eldridge, D. J., Poore, A. G., Ruiz-Colmenero, M., Letnic, M. & Soliveres, S. Ecosystem structure, function, and composition in rangelands are negatively affected by livestock grazing. Ecol. Appl. 26, 1273–1283 (2016).
Traba, J., Morales, M. B., de la Morena, E. L. G., Delgado, M. & Krištn, A. Selection of breeding territory by little bustard (Tetrax tetrax) males in central Spain: the role of arthropod availability. Ecol. Res. 3, 615–622 (2008).
Faria, N., Rabaça, J. E. & Morales, M. B. The importance of grazing regime in the provision of breeding habitat for grassland birds: the case of the endangered little bustard (Tetrax tetrax). J. Nat. Conserv. 20, 211–218 (2012).
Leitão, P. J., Moreira, F. & Osborne, P. E. Breeding habitat selection by steppe birds in Castro Verde: A remote sensing and advanced statistics approach. Ardeola 57, 93–116 (2010).
Moreira, F. et al. Spatial distribution patterns, habitat correlates and population estimates of steppe birds in Castro Verde. Airo. 17, 5–30 (2007).
Morgado, R. et al. Calandra lark habitat selection: strong fragmentation effects in a grassland specialist. Acta Oecol. 36, 63–73 (2010).
Silva, J. P., Palmeirim, J. M. & Moreira, F. Higher breeding densities of the threatened little bustard Tetrax tetrax occur in larger grassland fields: Implications for conservation. Biol. Conserv. 143, 2553–2558 (2010).
Morales, M. B., Traba, J., Delgado, M. P. & de la Morena, L. E. Sexual differences in microhabitat selection of breeding little bustards Tetrax tetrax: Ecological segregation based on vegetation structure. Acta Oecol. 34, 345–353 (2008).
Manrique, J. & Yanes, M. Thekla Lark Galerida theklae. In: Birds in Europe: their conservation status. 362–363 (BirdLife Conservation Series No. 3. BirdLife International, 1994).
Acknowledgements
We would like to thank all the farmers who kindly granted us access to their land for the purpose of this study and participated in the interviews as well as to Associação de Agricultores do Campo Branco (AACB) for the support during field work. This research results from project FARSYD (POCI-01-0145-FEDER-016664-PTDC/AAG-REC/5007/2014), supported by Norte Portugal Regional Operational Program (NORTE 2020), under the PORTUGAL 2020 Partnership Agreement, through the European Regional Development Fund (ERDF), and by national funds through FCT—Portuguese Science Foundation as well as from the project IF/01053/2015. R.F.R. and F.M. were financed by FCT—Fundação para a Ciência e a Tecnologia, I.P., through the project IF/01053/2015. J.A.D. was financed by FCT—Fundação para a Ciência e a Tecnologia, I.P., through the project FARSYD. J.S. was funded by Fundo Europeu de Desenvolvimento Regional through the Operational Competitiveness Factors Program “COMPETE” and by National Funds through the Foundation for Science and Technology (FCT) through the projects FARSYD (POCI-01-0145-FEDER-016664-PTDC/AAG-REC/5007/2014) and ALIENTRADE (PTDC/BIA-ECO/30931/2017-POCI-01-0145-FEDER-030931). L.R. acknowledges support from Portuguese National Funds through FCT, I.P., under the programme of “Stimulus of Scientific Employment—Individual Support” within the contract “CEECIND/00445/2017”. P.B. was funded by EDP Biodiversity Chair. A.L. and J.P.S. were supported by national funds through FCT—Fundação para a Ciência e a Tecnologia, I.P., in the context of the Transitory Norm-DL57/2016/CP1440/CT0001 and DL57/2016/CP1440/CT0021, respectively. We thank two anonymous reviewers for their constructive suggestions to a previous version of the manuscript.
Author information
Authors and Affiliations
Contributions
R.F.R., J.S., J.P.S. and F.M. conceptualised and designed the study. R.F.R. and J.A.D. collected and curated the data and conducted the data analysis. F.M. assisted in the data analysis and helped R.F.R. and J.A.D. writing the manuscript. J.S., J.P.S., L.R., S.S., P.B., A.L. and F.M. critically reviewed and revised the manuscript. All authors approved the final manuscript as submitted.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ramos, R.F., Diogo, J.A., Santana, J. et al. Impacts of sheep versus cattle livestock systems on birds of Mediterranean grasslands. Sci Rep 11, 10827 (2021). https://doi.org/10.1038/s41598-021-89975-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-89975-x
- Springer Nature Limited