
Abstract
During weaning, piglets experience concurrent social, physical, and nutritional stressors. Consequently, piglets often have poor feed intake and display increased oral manipulative behaviours post-weaning, indicative of compromised welfare. Black soldier fly larvae (BSFL) possess many attractive properties for pigs and could therefore function as effective edible enrichment, potentially alleviating weaning stress by facilitating exploration and promoting feed intake. In this study, pairs of piglets received a small amount of either live BSFL or wood shavings (8 pens/treatment) scattered throughout the pen twice a day for 11 days after weaning. Home-pen behaviour was scored by instantaneous scan sampling on day 2, 5 and 8, and behavioural responses to a novel environment and novel object were scored on day 10/11. Performance-related parameters were observed regularly. Larvae provisioning increased floor-directed exploration and decreased object-directed exploration, pig-directed oral manipulation, fighting and eating of pellets, and reduced neophobia towards a novel object. Pellet intake was significantly decreased by BSFL provisioning during day 4–11 post-weaning, although feed and net energy intake including BSFL never differed between treatments. BSFL provisioning did not influence piglet growth, feed efficiency, energy efficiency, and faecal consistency. To conclude, live BSFL provisioning positively affected post-weaning piglet behaviour while maintaining performance.
Similar content being viewed by others


Introduction
On commercial pig farms, piglets are often weaned abruptly around 3–4 weeks of age. This process involves several stressors, including separation from the sow, transitioning from a mainly liquid (milk) diet to a completely solid diet, and usually relocation to a new environment and mixing with unfamiliar piglets. Piglets are generally neophobic towards unfamiliar feed items1, and early and abrupt weaning often results in decreased feed consumption post-weaning. Decreased feed consumption can reduce growth and intestinal integrity, and increase disease susceptibility, reflected by the high incidence of post-weaning diarrhoea (as reviewed by2,3). Furthermore, abruptly weaned piglets frequently display increased levels of aggression and pig- and pen-directed oral manipulation behaviour, especially when housed in barren environments4,5. Early and abrupt weaning can severely impact the welfare of piglets, and may have lifelong detrimental effects on pig performance and intestinal health6,7.
One approach to ease the weaning transition would be to implement systems that allow for more gradual weaning, such as group farrowing systems with a prolonged lactation period8, though this requires large-scale adjustments to current husbandry management. Alternatively, providing environmental enrichment post-weaning can benefit piglet welfare, as its presence may facilitate enrichment oriented exploration, reducing the motivation to explore and manipulate pen mates4. Furthermore, previous studies have indicated that enriched-housed piglets often respond less neophobic towards novel objects9 and novel feed items10 than barren-housed piglets. Hemsworth et al. attributed reduced neophobia to regular confrontation with novel items which likely habituates pigs to novelty11. Additionally, stressful situations were found to increase food neophobia in mammals such as sheep and humans12,13. For pigs, it was found that environmental enrichment can reduce stress14, and as such enrichment could lessen feed neophobia. Reduced feed neophobia may result in increased feed intake10 and consequently better performance post-weaning. It has been shown, for example, that providing hanging ropes and tyres to piglets during several weeks after weaning improved piglet growth and reduced fearful responses towards a person15, and providing hanging ropes and wood shavings also improved piglet growth and increased exploration behaviour16. Other studies demonstrated that providing peat (for 5 weeks post-weaning) or providing increased space combined with straw, peat and branches (for 2 weeks post-weaning) reduced pig-directed oral manipulation behaviour, and increased exploration behaviour and growth4,17,18.
Edible items (e.g. straw) are very effective as enrichment, as they provide a consumable reward and thereby reinforce exploration behaviour19. Exploring and eating edible enrichment allow piglets to develop feeding-related behaviours such as chewing, adapting them to consuming solid feeds20, while simultaneously benefitting gut functioning21. Furthermore, providing edible enrichment creates dietary diversity, which has been shown to increase feed intake before weaning and may have a similar effect after weaning22,23. Durán et al. found that newly weaned piglets actively interact with and consume whey-based edible enrichment24. Also, providing straw post-weaning increased exploration behaviour and reduced pig- and pen-directed oral manipulation25,26. These studies highlight the potential of using edible enrichment post-weaning to ease the weaning transition, though this has not been widely studied.
Living insect larvae, which are a natural feed source of wild boar27, have the potential to function as effective edible enrichment. One species suitable for consumption by commercial pigs is the black soldier fly. Black soldier fly larvae (BSFL) are rich in fat, calcium, and protein with an appropriate amino acid profile for pigs28,29,30. Their palatability and high moisture content (55–65%)28,29,31 might stimulate intake by newly weaned piglets that are accustomed to a liquid milk diet, and their consistency could allow larvae to function as a transitory feed between milk and concentrate. BSFL are essentially free of carbohydrates28, therefore digestion is not limited by low secretion of starch-degrading enzymes in recently weaned piglets. In addition, BSFL possess many characteristics of effective enrichments. The larvae are not only edible, but also odorous, destructible, and manipulable, and these properties are highly interesting for pigs and facilitate exploration behaviour32,33. The ability of live larvae to crawl to different places could decrease the predictability of interacting with the larvae, and decreased predictability is known to prolong interest in enrichment34. Moreover, piglets seem to prefer interacting with larger sized feed items. Previous studies found that piglets prefer interacting with 5-mm diameter feed pellets over smaller 1.8-mm diameter pellets35, and they prefer interacting with 12-mm diameter pellets over 2-mm diameter pellets36. BSFL can grow up to 6 mm wide and up to 15–27 mm long28,37,38, and are thus substantially larger than most commonly provided pelleted feeds. Like other enrichment items, the presence of larvae might habituate pigs to novelty and/or have stress-reducing effects on pigs, potentially reducing neophobia.
Altogether, the properties of live BSFL suggest that they could function as effective edible enrichment post-weaning. This study aims to explore this potential by determining the effect of live BSFL provisioning post-weaning on piglet behaviour, fearfulness and neophobia, growth, feed and energy intake, feed and energy efficiency, and faecal consistency. It is expected that live BSFL provisioning will promote behaviours that are indicative of good welfare, e.g. exploration and play behaviours, and reduce aggression and pig- and pen-directed oral manipulation behaviour. Furthermore, providing live BSFL is expected to reduce neophobia and improve the performance and faecal consistency of post-weaning piglets.
Methods
The experimental protocol was approved by the Animal Care and Use committee of Wageningen University & Research under project licence number AVD1040020187184. The protocol was in accordance with the Dutch animal experimentation law and complies with European Directive 2010/63/EU. The experiment was carried out at the animal experiment facilities of Wageningen University & Research (Wageningen, The Netherlands). The ARRIVE guidelines for reporting animal experiments were taken into account in this study39.
Animals, housing and management
A total of 32 female, tail docked piglets (Pietrain x TN70) from 15 litters (range parity sows: 1–5) were weaned at 24.8 ± 0.8 days of age (day 0) and transported to the experimental facility. Pairs of unfamiliar piglets were selected based on body weight, to achieve a similar average weight (7.4 ± 0.2 kg) per pen. Pens were distributed over two rooms, balanced for treatment. Pairs of pigs were housed in pens of 2.85 × 1.20 m equipped with a feed trough (12 × 50 cm with three feeding places), one drinking nipple and one rubber ball on a chain. Feed and water were provided ad libitum. Flooring of all pens was a rubber mat covered by a 5 cm layer of wood shavings. Pigs were fed the creep feed they had also received during the suckling period (Speenkruimel Fit, ABZ Diervoeding) until day 2 after weaning. This feed was then mixed with the weaner feed (Speen Havic Top, AgruniekRijnvallei) for one day, after which only the weaner feed was provided. As feed consumption was initially very low, in all pens a spoonful of sugar was added when the weaner diet was first provided to ease the feed transition. In the experimental rooms a radio and lights were on from 07:00 to 19:00 h, and between 19:00 and 07:00 h the lights were dimmed, and the radio was off. Room temperature was 27 °C on arrival and was gradually decreased to 24 °C on day 11 post-weaning (the end of the experiment).
Experimental design
Pens were assigned to either the control (CON) or the black soldier fly larvae (LAR) treatment (8 pens/treatment). Piglets were provided with wood shavings (CON) or live BSFL (LAR) twice a day, at 09:00 and 13:00 h, from day 1 after weaning, by scattering the item in the pen on the wood shavings bedding. Wood shavings were provided to the CON treatment to create a similar disturbance in all pens. LAR pens received either 75 g (day 1–4) or 150 g (day 5–11) of live larvae twice a day, and CON pens always received a similar volume of wood shavings (approximately 100 and 200 ml, respectively). Live, 14-day-old BSFL were supplied weekly (by Bestico B.V., Berkel en Rodenrijs, The Netherlands) and stored at 12 °C. From approximately 30 min before provisioning, larvae were stored at room temperature to increase larvae activity.
Measurements
Home-pen behaviour
At arrival and on day 7 of the experiment, each piglet was marked with a colour stripe (stock marker spray) on their back for individual identification. The behaviour of all pigs was observed by 1-min interval instantaneous scan sampling on day 2, 5 and 8 post-weaning according to the ethogram in Table 1. Each of these 3 days had seven observation periods, starting at 07:15, 09:00, 10:15, 11:30, 13:00, 14:15 and 17:00 h. The periods starting at 09:00, 10:15, 13:00 and 14:15 h were observed live, where two observers observed 8 experimental pens (4 CON and 4 LAR) in one room each, switching pens every hour. Before observing, the observers were trained, and inter-observer reliability was regarded “almost perfect” (Cohen’s kappa > 0.8)40. The remaining observation periods were observed afterwards from video by one person (one of the two observers). All observations were done using the program Observer 14.2 (Noldus Information Technology B.V., Wageningen, The Netherlands).
Novel environment and novel object test
To determine pig responses to novelty, all experimental pigs, except one pig from the LAR treatment which was lame on the day of testing, were subjected to a Novel Environment Test and subsequent Novel Object Test. Tests were performed on day 10 and 11 post-weaning between 09:00 and 15:00 h. Half of the pigs were tested on 1 day and the other half on the other day, with the order being balanced for treatment and room. Prior to testing, pigs were individually caught, placed in a closed cart, and transported to an experimental room nearby. Here, the pig was placed in a start box alongside the experimental area. The door of the start box was opened within 10 s, giving the pig access to an unfamiliar area of 5.3 × 5.3 m with a rubber floor, surrounded by 1 m high hardwood walls. The novel environment test started when the pig had entered the area and the door was closed behind it. After 5 min, the novel object test was started by lowering an unfamiliar object (a metal bucket) from the ceiling in the centre of the area. The bucket was placed on the floor, which resulted in a noise. The rope attached to the bucket was held loosely. After five more minutes the test ended, and the pig was transported back to the home pen. During the tests, two observers were in the same room but out of sight of the pig, viewing the pig on a screen. One observer scored the behavioural and postural states, and the other observer scored behavioural events, according to the ethogram in Table 2. Scoring was done on a tablet with the program Observer 14.2 (Noldus Information Technology B.V., Wageningen, The Netherlands). Between tested pigs, faeces and urine were removed and the experimental area was cleaned with water and soap and dried with towels.
Performance
Piglets were individually weighed directly after arrival at the experimental facilities, and on day 1, 4, 7 and 11 post-weaning. Pellet intake per pen was determined by weighing leftovers on day 1, 2, 4, 7 and 11. Regular observations of the bedding indicated that live larvae were almost absent within a few hours after provisioning, therefore complete consumption of the larvae was assumed. Prior to the experiment, three samples of BSFL were analysed for their dry matter (DM), crude protein, crude fat, calcium, and phosphorus content, and based on this the net energy content (MJ NE/kg DM) of the larvae was calculated (Supplementary Table S1). The net energy content of the pelleted feed (MJ NE/kg DM) was calculated using the energy values provided by the feed manufacturers. The DM intake per pen excluding and including larvae were calculated and analysed separately, and based on the total DM intake including larvae the net energy intake (MJ NE), feed efficiency (weight gain (g)/dry matter intake (g)), and energy efficiency (weight gain (g)/net energy intake (MJ NE)) were calculated to account for the large difference in the net energy content between pellets and BSFL.
Faecal consistency and dry matter content
Every day at 08:30 h the faecal consistency of individual pigs was scored by two observers. Faecal consistency scores as visible around the anus of the pig at that time were based on the definition by Pedersen and Toft41. Score 1 represents firm faeces, score 2 soft but shaped faeces, score 3 loose faeces and score 4 water thin faeces. On day 4 and 7 faecal samples were collected during the weighing procedure, either by allowing the piglet to defecate by itself, or by stimulating defecation through gentle anal manipulation with a cotton swab for at most 10 s. The faecal samples were weighed before and after drying at 70 °C for 24 h and based on this the dry matter percentage was calculated.
Statistical analyses
Data processing
The feeder in one CON pen had fallen over on the last experimental day and this data was excluded from analysis. Home-pen behaviours were summed per piglet per day and expressed as the percentage of observed scans. To limit the number of variables, a factor analysis was conducted on data from the novel environment test and the novel object test (e.g.42). Behavioural states and postures were expressed as a percentage of time, and behavioural events were expressed as absolute frequencies. During the Novel Environment and Novel Object Test the behaviours “drawing back”, “defecating”, and “urinating”, and the postures “sitting” and “lying” were very rare, and they were not included in the factor analysis. The behaviours “high-pitched vocalizations” and “low-pitched vocalizations” were combined into “vocalizations” for the factor analyses. The latency (s) to explore the novel object was calculated and included in the factor analysis. Prior to analysis, a Pearson’s correlation test performed on all variables showed that the variable “standing” was strongly negatively correlated to “moving” in the novel object test (r = − 0.80), therefore “standing” was excluded from the factor analysis. For the novel environment test, the variables “freezing” and “moving” each contained one outlier (based on the Grubb’s test), and these outliers were excluded from analysis. The variables “freezing” (for the novel environment test and the novel object test) and “exploring object” (for the novel object test) were arcsine square root transformed, and “latency to explore object” was logarithmically transformed for normalization.
Data analysis
All data were analysed using the statistical software SAS 9.4 (SAS Institute Inc., Cary, NC, USA). General linear (mixed) model residuals were checked for normality.
The proportions of time spent on each observed behaviour in the home pen were analysed in generalised linear mixed models (GLIMMIX in SAS) with a binomial distribution, logit link function, and additional multiplicative over-dispersion parameter. Treatment, room, and day were included as fixed effects. The interaction between day and treatment was removed from the final models as this effect was never significant. Furthermore, the models included a random effect of pen nested within treatment and room, and a repeated effect of test day with pig as subject, using a homogenous first-order autoregressive covariance structure.
The variables obtained from the novel environment test and the novel object test were put in a factor analysis with orthogonal varimax rotation, and factors with an eigenvalue above one were retained. This resulted in one factor for the novel environment test and one factor for the novel object test (Table 3), explaining 63% and 72% of the variation, respectively. The scores of each pig for each factor were analysed with a linear mixed model (MIXED in SAS) including a fixed effect of treatment, room and test day and a random effect of pen nested in treatment and room.
Body weight and average daily gain were analysed in a linear mixed model including a fixed effect of treatment and room, a random effect of pen nested in treatment and room, and weight directly after weaning as a covariate. Dry matter intake excluding and including BSFL, net energy intake, feed efficiency, and energy efficiency were analysed for treatment effects on pen level in general linear models. The proportion of faecal dry matter was analysed per day in a similar model as the home-pen behaviours but without a repeated effect.
P-values below 0.05 were considered statistically significant. Significant fixed effects were further analysed using differences in least square means with a Tukey’s HSD correction. Data are presented as pen means ± SEM unless stated otherwise.
Results
Home-pen behaviour
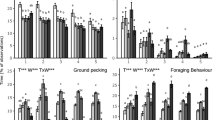
Averaged over the three time points, LAR piglets spent less time on pig-directed oral manipulation (F(1,13) = 24.27, p < 0.001), fighting (F(1,13) = 6.92, p = 0.02), exploring objects (F(1,13) = 17.89, p < 0.001) and eating feed (F(1,13) = 23.89, p < 0.001), and they spent more time on exploring the floor (F(1,13) = 29.14, p < 0.001) than the CON piglets. The time spent inactive (F(1,13) = 0.02, p = 0.89), social playing (F(1,13) = 0.39, p = 0.54), and non-social playing (F(1,13) = 0.09, p = 0.77) did not differ between treatments. Considering the effect of day, the time spent on pig-direct oral manipulation varied over time (F(1,78) = 5.13, p = 0.01), with the lowest levels occurring on day 5 as compared to day 2 and 8 after weaning. The time spent on exploring objects decreased over time (F(1,78) = 9.33, p < 0.001) and the time spent on social play (F(1,78) = 18.18, p < 0.001) and eating feed (F(1,78) = 7.93, p < 0.001) increased over time. The time spent fighting (F(1,78) = 2.43, p = 0.09), non-social playing (F(1,78) = 0.00, p = 1.00), exploring the floor (F(1,78) = 1.90, p = 0.16) and being inactive (F(1,78) = 0.45, p = 0.64) were not influenced by day (Fig. 1).

Behaviours observed in the home pen. Behavioural time budgets expressed as pen means ± SEM of pigs receiving wood shavings (CON) or black soldier fly larvae (LAR) twice a day. Effects of Treatment (T) and Day (D) are indicated as ***p < 0.001, **p < 0.01, *p < 0.05 and ns if p > 0.05. The Treatment x Day interaction was never significant and is therefore not shown.
Novel environment and novel object test
The one retained factor of the novel environment test had high positive loadings for standing, freezing and vocalizations, and high negative loadings for exploring the environment and moving, therefore this factor will be designated as “fearful” (Table 3). Treatment did not influence the pigs’ scores for this factor (CON: 0.02 ± 0.20, LAR: 0.03 ± 0.28, F(1,13) = 0.02, p = 0.90).
The one retained factor of the novel object test had high positive loadings for freezing and the latency to explore the object, and high negative loadings on time spent moving and exploring the object, and this factor will be designated as “neophobic” (Table 3). Here, treatment influenced piglet scores, where CON piglets scored higher on the “neophobic” factor than LAR piglets (CON: 0.43 ± 0.27, LAR: − 0.39 ± 0.25, F(1,13) = 5.30, p = 0.04).
Performance
Performance parameters measured over the experimental period are shown in Table 4. Treatment did not affect average daily gain (F(1,13) = 0.07, p = 0.79) and final weight (F(1,13) = 0.07, p = 0.79) of the piglets, and it also did not influence their feed efficiency (based on dry matter intake including BSFL, F(1,12) = 2.64, p = 0.13) and energy efficiency (based on dry matter intake including BSFL, F(1,12) = 0.71, p = 0.42)). Dry matter intake excluding BSFL over the whole experimental period was lower in LAR piglets than in CON piglets (F(1,12) = 6.35, p = 0.03). Detailed analysis of the separate periods for which feed intake was determined showed that dry matter intake excluding BSFL was lower in LAR piglets than in CON piglets during day 4–7 (F(1,13) = 5.57, p = 0.03) and during day 7–11 (F(1,12 = 10.18, p = 0.01, Supplementary Table S2). Treatment did not influence the dry matter intake including BSFL (F(1,12) = 0.06, p = 0.82) or the total energy intake including BSFL (F(1,12) = 2.31, p = 0.15).
Faecal consistency and dry matter content
Faecal consistency scores of 3 and 4 were rare (< 2%), and as only these scores indicate diarrhoea, the faecal consistency scores were not further analysed. The faecal dry matter content on day 4 and 7 was not influenced by treatment (Day 4, CON: 31.6 ± 1.7%, LAR: 28.7 ± 1.4%, F(1,13) = 1.14, p = 0.30, Day 7, CON: 28.6 ± 3.4%, LAR: 27.7 ± 1.7%, F(1,13) = 0.02, p = 0.89).
Discussion
In this study, we investigated the potential of using live black soldier fly larvae (BSFL) as edible enrichment for piglets post-weaning. Providing a small amount of live BSFL twice a day increased floor-directed exploration, while it decreased object-directed exploration, pig-directed oral manipulation, fighting, and time spent eating pellets. In addition, a factor analysis suggests that LAR piglets were less neophobic, i.e. fearful of a novel object, than CON piglets when this object is introduced to them in an unfamiliar environment. Daily consumption of a small amount of live BSFL reduced pellet intake, but total feed and net energy intake (including BSFL), average daily gain, feed efficiency, energy efficiency, and faecal consistency were not affected by live BSFL provisioning.
In line with our expectations, live BSFL provisioning redirected exploration and manipulation behaviours away from objects and conspecifics and towards the floor where the larvae were provided, while simultaneously decreasing aggression. These effects occurred over the whole day despite the larvae being only transiently present each day and indicate that temporary access to live BSFL can benefit piglet welfare. In the ethogram applied in the current study, pig-directed oral manipulation included nosing of pen mates, which does not necessarily negatively impact piglet welfare. However, under barren (i.e. without bedding and with limited space) commercial conditions, pig-directed oral manipulation often accumulates into damaging behaviours that are harmful to pigs, such as tail biting4,17,43. Additionally, post-weaning manipulation behaviours and aggression tend to be more common in barren commercial conditions compared to our experimental setting17,44,45. Therefore, the observed welfare benefits associated with live BSFL provisioning are likely amplified in commercial pig husbandry conditions, and this merits investigation in future studies.
As live BSFL possess many features interesting for pigs32,33, they likely provided ample exploration opportunities to satisfy the piglet’s exploratory needs. By promoting exploration, the motivation to perform other behaviours such as oral manipulation behaviours directed at pen mates may be decreased. Moreover, providing enrichment can have stress-reducing effects on pigs14, and as such, the presence of larvae may have attenuated the piglet’s weaning stress. This could have shifted their behavioural repertoire towards more favourable behaviours and reduced aggressive behaviours. In line with our results, increased exploration and reduced harmful social behaviour were also found in studies using peat, straw, or a complex environment with increased space and a multitude of enrichment materials4,17,25. However, while these studies also observed increased play behaviour in enriched pigs, live BSFL provisioning did not affect social and non-social play. Possibly, the elevated level of exploratory behaviour direct towards the BSFL reduced the motivation to perform other behaviours including (social) play. Also, in the current study piglets were housed in relatively small groups with 2 piglets per pen as compared to 4–11 piglets per pen in the previous studies, and facilitation of social play was therefore relatively low. On the other hand, all CON and LAR piglets in the current study had access to bedding, which can already promote play as compared with barren environments46. The influence of the experimental conditions may have outweighed any additional effects of live BSFL provisioning on play behaviour, and differential effects on play behaviour may be found under commercial conditions.
Besides studying home-pen behaviour, behavioural responses to novel situations were determined and analysed with a factor analysis. It can be speculated that piglets with reduced neophobic responses can better cope with novel situations such as changes to their environment. The factor resulting from the analysis of the novel object test data had high positive loadings on the time spent freezing and the latency to explore the unfamiliar object, and high negative loadings on time spent moving and exploring the object. Previous studies indicated that relatively slower and lesser exploration of a (novel) object and being less active in unfamiliar situations can be associated with increased fearfulness and/or anxiety47,48,49, and an alert standing posture, i.e. freezing, is often exhibited in response to aversive situations50. Donald et al. found a factor with similar behavioural components and loadings for piglets participating in an open field test in which novel objects were present, and concluded that this factor mainly reflected the degree of neophobia, as the fear-related behaviours occurred in response to novelty51. Likewise, the factor found in our study is assumed to reflect the level of neophobia, and as LAR piglets scored lower on this factor compared to CON piglets, LAR piglets are assumed to be less neophobic. The transient presence of live larvae is therefore sufficient to decrease neophobia. This may indicate that the feedback provided by tasting and eating larvae enhances the positive associations with novelty enough to compensate for the occasional absence of larvae. In line with our results, long-term access to live BSFL decreased broiler fearfulness, as shown by a decreased amount of time spent in tonic immobility52. Similarly, previous studies found that housing pigs with marginally increased space and more toys elevated exploration of a novel object53, and providing hanging ropes together with hanging rubber tyre tubes reduced the latency to approach a person15.
It should be noted that reduced neophobic responses by piglets that were given larvae were only observed in the novel object test, and not in the preceding novel environment test. The factors retained in this test had, similarly to the novel object test, a high negative loading for moving and a high positive loading on freezing. Additionally, this factor had a high negative loading for time spent on exploring the environment, and high positive loadings for standing and the frequency of vocalizations. In a previous study, pigs which had received fear-reducing drugs spent more time on exploring their environment and less time on vocalizing compared to saline-treated pigs51, indicating that these behaviours are also related to fearfulness. Taken together, the factor retained from the novel environment test is assumed to reflect general fearfulness and/or anxiety. The observation that scores for this factor were not affected by treatment likely relates to the stimulus-specific responses of piglets towards novelty. This was demonstrated by Hemsworth et al. who found that piglets that received objects in their home pen were more willing to explore a novel object compared to piglets that had received human contact in their home pen11. The absence of any observed behaviour difference in the novel environment test could therefore be due to the absence of novel items in this test. A novel object test was previously found to cause more fear-related behaviour and higher serotonergic responses compared to a novel environment test49, and the lower fear response prompted by a novel environment might not be sufficient to observe any treatment effects on fearfulness. While our results demonstrate the potential of live BSFL provisioning to reduce neophobia in piglets, further research is required to assess whether also general fearfulness is affected.
Some studies have found a link between increased exploration behaviour and increased feed intake pre-weaning54 and post-weaning55. As live BSFL provisioning increased exploratory behaviours, it was expected to also increase feed intake, and potentially increase the time spent eating pellets. In contrast, LAR piglets spent less time eating pellets than CON piglets, and pellet consumption during day 4–11 was reduced in LAR piglets. The main reason that pellet consumption was reduced by live BSFL provisioning appears to be that consumption of live BSFL sufficiently fulfilled the pigs’ motivation to eat. In a previous study in which a range of dietary options were available pre-weaning, consumption of the more palatable feed items was preferred22, and the high palatability of BSFL likely caused a similar preference and high consumption. Including BSFL meal in pig diets has previously had positive56 or neutral57,58 effects on pig growth, while the impact of consuming whole, live BSFL on piglet performance has, to our knowledge, not been studied. The absence of any effect of whole live BSFL consumption on total dry matter intake, net energy intake, feed efficiency, energy efficiency and average daily gain illustrates the nutritional equivalence of BSFL to feed on a dry matter basis in the current experimental setting. This finding highlights the potential nutritional value of BSFL for pigs, as suggested previously28,31. It is important to note that availability of BSFL in the current study was restricted to a fixed daily portion, and ad libitum provisioning could differentially influence piglet performance. Immediately post-weaning, when solid feed intake is generally low2, ad libitum BSFL provisioning might boost piglet performance by increasing nutrient intake. This, in turn, likely supports good intestinal integrity and growth59, warranting future investigation. Furthermore, when a substantial part of the diet is replaced with larvae, it will be necessary to adjust the nutrient composition of the regular feed to assure adequate pig performance. Optimizing a pig diet including larvae requires further research into the complete nutrient composition and digestibility of BSFL.
The piglets’ faecal dry matter content was not affected by live BSFL consumption. The occurrence of diarrhoea, indicated by a faecal consistency score of 3 or 441, was very low in both treatment groups. This could be the results of the wood shavings present in all pens, which are known to reduce the number of days pigs have diarrhoea and benefit the overall faecal consistency score60. Under commercial conditions, diarrhoea is more common, for example due to the reduced feed intake and increased stress around weaning (as reviewed by59). Investigating live BSFL provisioning in this setting is recommended to determine their effect on post-weaning diarrhoea.
To conclude, post-weaning live BSFL provisioning had beneficial effects on piglet behaviour, by facilitating exploration behaviours and reducing the need to orally manipulate objects and pen mates. The presence of larvae also reduced neophobic responses towards a novel object. The performance of piglets that consumed a small amount of larvae was maintained, as their total feed and net energy intake, feed efficiency, energy efficiency, and average daily gain did not differ from that of control piglets. Under barren commercial conditions, larvae provisioning has the potential to further benefit piglet welfare, though this remains to be confirmed by future studies. These studies should also consider appropriate larvae provisioning methods under commercial conditions, as scattering larvae on slatted floors is impractical.
References
Rozin, P. & Vollmecke, T. A. Food likes and dislikes. Annu. Rev. Nutr. 6, 433–456 (1986).
Dong, G. Z. & Pluske, J. R. The low feed intake in newly-weaned pigs: problems and possible solutions. Asian-Austral. J. Anim. Sci. 20, 440–452 (2007).
Campbell, J. M., Crenshaw, J. D. & Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 4, 14 (2013).
Oostindjer, M., van den Brand, H., Kemp, B. & Bolhuis, J. E. Effects of environmental enrichment and loose housing of lactating sows on piglet behaviour before and after weaning. Appl. Anim. Behav. Sci. 134, 31–41 (2011).
Colson, V., Orgeur, P., Foury, A. & Mormède, P. Consequences of weaning piglets at 21 and 28 days on growth, behaviour and hormonal responses. Appl. Anim. Behav. Sci. 98, 70–88 (2006).
Moeser, A. J., Pohl, C. S. & Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 3, 313–321 (2017).
Cabrera, R. A. et al. Impact of lactation length and piglet weaning weight on long-term growth and viability of progeny1,2. J. Anim. Sci. 88, 2265–2276 (2010).
van Nieuwamerongen, S. E., Soede, N. M., van der Peet-Schwering, C. M. C., Kemp, B. & Bolhuis, J. E. Gradual weaning during an extended lactation period improves performance and behavior of pigs raised in a multi-suckling system. Appl. Anim. Behav. Sci. 194, 24–35 (2017).
Beattie, V. E., O’Connell, N. E., Kilpatrick, D. J. & Moss, B. W. Influence of environmental enrichment on welfare-related behavioural and physiological parameters in growing pigs. Anim. Sci. 70, 443–450 (2000).
Oostindjer, M., Muñoz, J. M., Van den Brand, H., Kemp, B. & Bolhuis, J. E. Maternal presence and environmental enrichment affect food neophobia of piglets. Biol. Lett. 7, 19–22 (2011).
Hemsworth, P. H., Price, E. O. & Borgwardt, R. Behavioural responses of domestic pigs and cattle to humans and novel stimuli. Appl. Anim. Behav. Sci. 50, 43–56 (1996).
Burritt, E. A. & Provenza, F. D. Effect of an unfamiliar location on the consumption of novel and familiar foods by sheep. Appl. Anim. Behav. Sci. 54, 317–325 (1997).
Pliner, P., Eng, A. & Krishnan, K. The effects of fear and hunger on food neophobia in humans. Appetite 25, 77–87 (1995).
Munsterhjelm, C. et al. Environmental enrichment in early life affects cortisol patterns in growing pigs. Animal 4, 242–249 (2010).
Rodarte, L. F., Ducoing, A., Galindo, F., Romano, M. C. & Valdez, R. A. The effect of environmental manipulation on behavior, salivary cortisol, and growth of piglets weaned at 14 days of age. J. Appl. Anim. Welf. Sci. 7, 171–179 (2004).
Oliveira, R. F. et al. Environmental enrichment improves the performance and behavior of piglets in the nursery phase. Arq. Bras. Med. Veterinária e Zootec. 68, 415–421 (2016).
Vanheukelom, V., Driessen, B., Maenhout, D. & Geers, R. Peat as environmental enrichment for piglets: The effect on behaviour, skin lesions and production results. Appl. Anim. Behav. Sci. 134, 42–47 (2011).
Oostindjer, M. et al. Effects of environmental enrichment and loose housing of lactating sows on piglet performance before and after weaning. J. Anim. Sci. 88, 3554–3562 (2010).
Van de Weerd, H. A., Docking, C. M., Day, J. E. L., Breuer, K. & Edwards, S. A. Effects of species-relevant environmental enrichment on the behaviour and productivity of finishing pigs. Appl. Anim. Behav. Sci. 99, 230–247 (2005).
Cox, L. N. & Cooper, J. J. Observations on the pre- and post-weaning behaviour of piglets reared in commercial indoor and outdoor environments. Anim. Sci. 72, 75–86 (2001).
Pluske, J. R. et al. Microbial diversity in the large intestine of pigs born and reared in different environments. Livest. Sci. 108, 113–116 (2007).
Middelkoop, A., van Marwijk, M. A., Kemp, B. & Bolhuis, J. E. Pigs like it varied; Feeding behavior and pre- and post-weaning performance of piglets exposed to dietary diversity and feed hidden in substrate during lactation. Front. Vet. Sci. 6, 408 (2019).
Middelkoop, A. et al. Dietary diversity affects feeding behaviour of suckling piglets. Appl. Anim. Behav. Sci. 205, 151–158 (2018).
Durán, E., Churio, O., Lagos, J., Tadich, T. & Valenzuela, C. Development of edible environmental enrichment objects for weaned pigs. J. Vet. Behav. 34, 7–12 (2019).
Bolhuis, J. E., Schouten, W. G. P., Schrama, J. W. & Wiegant, V. M. Behavioural development of pigs with different coping characteristics in barren and substrate-enriched housing conditions. Appl. Anim. Behav. Sci. 93, 213–228 (2005).
Kelly, H. R. C., Bruce, J. M., English, P. R., Fowler, V. R. & Edwards, S. A. Behaviour of 3-week weaned pigs in Straw-Flow, deep straw and flatdeck housing systems. Appl. Anim. Behav. Sci. 68, 269–280 (2000).
Ballari, S. A. & Barrios-García, M. N. A review of wild boar Sus scrofa diet and factors affecting food selection in native and introduced ranges. Mamm. Rev. 44, 124–134 (2014).
Makkar, H. P. S., Tran, G., Heuzé, V. & Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 197, 1–33 (2014).
Spranghers, T. et al. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 97, 2594–2600 (2017).
Barragan-Fonseca, K. B., Dicke, M. & van Loon, J. J. A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed a review. J. Insects Food Feed 3, 105–120 (2017).
Veldkamp, T. & Bosch, G. Insects—A protein rich feed ingredient in pig and poultry diets. Anim. Front. 5, 45–50 (2015).
Van de Weerd, H. A., Docking, C. M., Day, J. E. L., Avery, P. J. & Edwards, S. A. A systematic approach towards developing environmental enrichment for pigs. Appl. Anim. Behav. Sci. 84, 101–118 (2003).
Studnitz, M., Jensen, M. B. & Pedersen, L. J. Why do pigs root and in what will they root?: A review on the exploratory behaviour of pigs in relation to environmental enrichment. Appl. Anim. Behav. Sci. 107, 183–197 (2007).
van de Weerd, H. & Ison, S. Providing effective environmental enrichment to pigs: How far have we come?. Animals 9, 254 (2019).
Edge, H. L., Dalby, J. A., Rowlinson, P. & Varley, M. A. The effect of pellet diameter on the performance of young pigs. Livest. Prod. Sci. 97, 203–209 (2005).
van den Brand, H. et al. Effects of pellet diameter during and after lactation on feed intake of piglets pre- and postweaning. J. Anim. Sci. 92, 4145–4153 (2014).
Jucker, C., Erba, D., Leonardi, M. G., Lupi, D. & Savoldelli, S. Assessment of vegetable and fruit substrates as potential rearing media for Hermetia illucens (Diptera: Stratiomyidae) larvae. Environ. Entomol. 46, 1415–1423 (2017).
Tinder, A. C., Puckett, R. T., Turner, N. D., Cammack, J. A. & Tomberlin, J. K. Bioconversion of sorghum and cowpea by black soldier fly (Hermetia illucens (L.)) larvae for alternative protein production. J. Insects Food Feed 3, 121–130 (2017).
du Sert, N. P. et al. Reporting animal research: Explanation and elaboration for the arrive guidelines 2.0. PLoS Biol. 18, e3000411 (2020).
Landis, J. R. & Koch, G. G. The measurement of observer agreement for categorical data. Biometrics 33, 159–174 (1977).
Pedersen, K. S. & Toft, N. Intra- and inter-observer agreement when using a descriptive classification scale for clinical assessment of faecal consistency in growing pigs. Prev. Vet. Med. 98, 288–291 (2011).
Reimert, I., Rodenburg, T. B., Ursinus, W. W., Kemp, B. & Bolhuis, J. E. Responses to novel situations of female and castrated male pigs with divergent social breeding values and different backtest classifications in barren and straw-enriched housing. Appl. Anim. Behav. Sci. 151, 24–35 (2014).
Zupan, M., Janczak, A. M., Framstad, T. & Zanella, A. J. The effect of biting tails and having tails bitten in pigs. Physiol. Behav. 106, 638–644 (2012).
Weary, D. M., Jasper, J. & Hötzel, M. J. Understanding weaning distress. Appl. Anim. Behav. Sci. 110, 24–41 (2008).
Day, J. E. L., Van de Weerd, H. A. & Edwards, S. A. The effect of varying lengths of straw bedding on the behaviour of growing pigs. Appl. Anim. Behav. Sci. 109, 249–260 (2008).
Tuyttens, F. A. M. The importance of straw for pig and cattle welfare: A review. Appl. Anim. Behav. Sci. 92, 261–282 (2005).
Brown, J. A. et al. Reliability of temperament tests on finishing pigs in group-housing and comparison to social tests. Appl. Anim. Behav. Sci. 118, 28–35 (2009).
Dalmau, A., Fabrega, E. & Velarde, A. Fear assessment in pigs exposed to a novel object test. Appl. Anim. Behav. Sci. 117, 173–180 (2009).
Ursinus, W. W. et al. Relations between peripheral and brain serotonin measures and behavioural responses in a novelty test in pigs. Physiol. Behav. 118, 88–96 (2013).
Reimert, I., Bolhuis, J. E., Kemp, B. & Rodenburg, T. B. Indicators of positive and negative emotions and emotional contagion in pigs. Physiol. Behav. 109, 42–50 (2013).
Donald, R. D., Healy, S. D., Lawrence, A. B. & Rutherford, K. M. D. Emotionality in growing pigs: Is the open field a valid test?. Physiol. Behav. 104, 906–913 (2011).
Ipema, A. F., Bokkers, E. A. M., Gerrits, W. J. J., Kemp, B. & Bolhuis, J. E. Long-term access to live black soldier fly larvae (Hermetia illucens) stimulates activity and reduces fearfulness of broilers, without affecting health. Sci. Rep. 10, 1–13 (2020).
Tönepöhl, B. et al. Effect of marginal environmental and social enrichment during rearing on pigs’ reactions to novelty, conspecifics and handling. Appl. Anim. Behav. Sci. 140, 137–145 (2012).
Kuller, W. I., Tobias, T. J. & van Nes, A. Creep feed intake in unweaned piglets is increased by exploration stimulating feeder. Livest. Sci. 129, 228–231 (2010).
Middelkoop, A., Costermans, N., Kemp, B. & Bolhuis, J. E. Feed intake of the sow and playful creep feeding of piglets influence piglet behaviour and performance before and after weaning. Sci. Rep. 9, 16140 (2019).
Yu, M. et al. Evaluation of full-fat Hermetia illucens larvae meal as a fishmeal replacement for weanling piglets: Effects on the growth performance, apparent nutrient digestibility, blood parameters and gut morphology. Anim. Feed Sci. Technol. 264, 114431 (2020).
Chia, S. Y. et al. Effect of dietary replacement of fishmeal by insect meal on growth performance, blood profiles and economics of growing pigs in Kenya. Animals 9, 705 (2019).
Spranghers, T. et al. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Anim. Feed Sci. Technol. 235, 33–42 (2018).
Heo, J. M. et al. Gastrointestinal health and function in weaned pigs: A review of feeding strategies to control post-weaning diarrhoea without using in-feed antimicrobial compounds. J. Anim. Physiol. Anim. Nutr. (Berl.) 97, 207–237 (2013).
Munsterhjelm, C. et al. Experience of moderate bedding affects behaviour of growing pigs. Appl. Anim. Behav. Sci. 118, 42–53 (2009).
Acknowledgements
This study is part of the research project ‘Assessing the potential of insects to reduce the environmental impact of livestock production and improve livestock health and welfare in concert’, with project number 15567. The project is financed by the Netherlands Organization for Scientific Research (NWO), HatchTech, ForFarmers and Bestico. The authors thank Yilin Bian, Manon van Marwijk, and other colleagues of the Adaptation Physiology group of Wageningen University for their assistance with the experiment. Also, the help of the personnel of the experimental facilities (CARUS) is gratefully acknowledged.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: A.I., E.B., W.G., B.K. and J.E.B. Performed research: A.I. Analysed data: A.I. Wrote the paper: A.I. Contributed to the critical appraisal of the paper and approved the final version: A.I., E.B., W.G., B.K. and J.E.B.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ipema, A.F., Bokkers, E.A.M., Gerrits, W.J.J. et al. Providing live black soldier fly larvae (Hermetia illucens) improves welfare while maintaining performance of piglets post-weaning. Sci Rep 11, 7371 (2021). https://doi.org/10.1038/s41598-021-86765-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-86765-3
- Springer Nature Limited
This article is cited by
-
Future opportunities for products derived from black soldier fly (BSF) treatment as animal feed and fertilizer - A systematic review
Environment, Development and Sustainability (2024)
-
Principles for the responsible use of farmed insects as livestock feed
Nature Food (2022)