
Abstract
Subjective hyperosmia refers to a self-reported olfactory ability that is superior to a normal, intact sense of smell (normosmia), and is associated with olfactory emotional experience. The current study used voxel-based morphometry to investigate the gray matter volume (GMV) in people with self-rated hyperosmia (subjective hyperosmia, SH, N = 18) in comparison to people with self-rated normal olfaction (subjective normosmia, SN, N = 14). Participants’ olfactory function were assessed by the extensive olfactory test battery, the “Sniffin’ Sticks” test. Within the predicted brain regions (regions-of-interest analyses), the SH participants showed larger GMV of the left hippocampus as compared to SN participants (FWE corrected p < 0.05). Further, the whole-brain search indicated that SH had larger GMV of the bilateral hippocampus, the right hypothalamus, the left precuneus, and the left superior frontal gyrus as compared to the SN group. ROI analyses showed positive correlations between the left hippocampal GMV and odor threshold or discrimination scores across all participants. In addition, the whole-brain analysis suggested that the self-rated olfactory ability was positively associated with GMV in the cerebellum, superior frontal gyrus and the precentral gyrus among SH participants. In conclusion, the current results suggest that SH was associated with increased GMV in several brain regions that were previously shown to be involved in the processing of cognitive aspects of odors.
Similar content being viewed by others

Introduction
Olfactory perception exhibits large inter-individual variations. Hyperosmia refers to an olfactory ability that is superior to a normal, intact sense of smell (normosmia)1 which includes subjects who reach at least the 90th percentile of the standardized Sniffin’ Sticks test score of the group aged 21–30 years (41.5 points or higher)2. People with hyperosmia are identified from 11 to 70 years, and their frequency peaks (around 11%) in the twenties2. Overall, clinically relevant hyperosmia seems to be a rare condition, although it is sometimes observed for specific odorants3,4.
Multiple studies have investigated the consistency between self-ratings of olfactory ability and the scores derived from standardized olfactory tests. Several studies found that patients with olfactory dysfunction could report their ability to smell with a relatively high accuracy5,6,7, although one study with large patient sample size (n = 1227) found nearly one-third patients with complete smell loss (anosmia) judged their sense of smell as at least “average”8. Such correlation between the olfactory functions derived from self-ratings and standardized psychophysical measurements becomes more ambiguous for normosmic people, with several studies reporting negative results5,7,8,9,10. Knaapila et al.11 found that in healthy participants, the self-rated olfactory acuity is associated with odor-evoked annoyance, which is related to negative valence (unpleasantness) of odors. Similarly, Knaapila and Tuorila12 noted that participants who regarded their sense of smell as exceptionally good reported more negative odor-related experiences than sex- and age-matched controls. In addition, people who regard novel objects’ odor as an important reason for liking or disliking them may assume this as an indicator for their particularly acute sense of smell13. Besides, it has been reported that the ability of creating olfactory mental images (e.g. the vividness of olfactory imagery) predicted self-evaluated olfactory function in both healthy participants and patients with smell loss5. Therefore, the self-reported superior olfactory performance seems to be associated with hypersensitivity towards the emotional and affective processing of olfactory stimuli.
In terms of the neural substrates of sensory hypersensitivity, one pervasive assumption in the literature is that individuals who are subjectively hypersensitive have greater neural responses to sensory stimuli14. Brain imaging studies using functional magnetic resonance imaging (fMRI) have suggested a link between sensory hypersensitivity and increased activity in limbic regions (amygdala, hippocampus). For example, groups known to report subjectively high sensory sensitivity such as autism15,16, have a greater BOLD response to auditory, visual or tactile stimuli. For the anatomical features, neurotypical variation in subjective hyper-sensitivity to sounds has been shown to be linked to increased gray matter volume of the auditory cortex but also a number of emotional processing related limbic regions, such as the amygdala and hippocampus17. Self-reported pain sensitivity rather than pain threshold was associated with increased gray matter volume in bilateral parahippocampal gyrus, extending to the hippocampus18. There were only few studies on the neural basis for olfactory hypersensitivity in literature. Professionals with long-time olfactory experiences showed increased gray matter volume in the insula and the entorhinal cortex as part of the olfactory system19. In a recent study, Wabnegger et al.20 showed that participants with hyperosmia (determined by Sniffin’ Sticks test with combined odor threshold, discrimination and identification [TDI] score over 41) had increased gray matter volume of the hippocampus and anterior insula. However, whether differences in self-evaluated olfactory function also show a neuroanatomical correlate, reflected by local differences in gray matter volume, is unknown.
In the present study, we used voxel-based morphometry (VBM) to investigate the gray matter volumes in a group with subjective hyperosmia (SH), and compared them to a group with subjective normosmia (SN), who have a self-reported normal sense of smell. Given that the emotional eloquent areas share function with olfactory processing, especially the limbic brain structures (hippocampus and amygdala), the orbitofrontal cortex (OFC) and the insula21,22,23,24, we focused on those brain regions and tempted to hypothesize that an increased gray matter volume in SH compared to SN participants.
Methods and materials
Participants recruitment
Participants underwent two steps of screening. First, participants were interviewed on phone about their sense of smell (provided with three choices: worse than normal, normal; better than normal). On the test day which took place at least one week after the telephone interview, participants were asked again to perform self-ratings for their olfactory performance on a 9-point scale (9 = extremely well, 8 = very well; 7 = much better; 6 = moderately better; 5 = normal, 4 = moderately worse; 3 = much worse; 2 = very bad; 1 = no perception). The SH participants were those who reported their sense of smell as “better than normal” over the phone interview, and also rated their sense of smell better than others (score higher than 5) on the test day. Similarly, SN participants were those who reported their sense of smell as “normal” during the phone interview and rated their sense of smell as normal (score = 5) on the test days.
In total 14 SH and 18 SN participants were included in the study. The participants’ demographical information are shown in Table 1. Participants were also screened to be free of major nasal pathology, which was ascertained by nasal endoscopy and a detailed, standardized interview. Their right-handedness were ascertained by a handedness questionnaire25. Other criteria for exclusion were pregnancy, neurological or psychiatric diseases, ferromagnetic implants (e.g., pacemakers and cochlear implants), previous head injury, or claustrophobia, smoking, and diseases or medication use known to be associated with olfactory dysfunction. The study was approved by the Ethics Committee at the TU Dresden (Protocol # EK262082010).
Olfactory test
Olfactory function was assessed using the “Sniffin’ Sticks” test (Burghart GmbH, Wedel, Germany)26, which is one of the most widely used standardized tests of olfactory performance, based on pen-like odor dispensing devices. The test assesses individual’s odor threshold (sensitivity), odor discrimination and odor identification performances. In the olfactory threshold test, odours were presented in 16 triplets of pens, two containing an odourless solvent and the other containing the target odor (2-Phenylethanol, rose-like smell) at a certain dilution (16 dilutions). The participants were asked to indicate the pen with the rose-like smell. Sensitivity was determined using a single-staircase, triple-forced choice procedure. Two successive correct identifications or one incorrect identification triggered a reversal of the staircase, i.e., the next higher or the next lower concentration step was presented, respectively. Seven reversals had to be obtained (including the starting point), and the sensitivity was defined as the mean of the last four staircase reversals. In the odour discrimination test, 16 triplets of pens, with two containing the same odor and the third a different one, were presented to the participants. Participants were asked to identify which of the three pens smelled differently. In the odor identification test, 16 common odors had to be identified from a list of four descriptors (multiple forced-choice procedure). For odor presentation, the experimenter removed the cap of the pens and held the stick for about 3 s at approximately 1–2 cm in front of both nostrils of the participants. The interval between two triplet presentations was approximately 20 s. Three olfactory tests were performed, always in the same order: odour detection threshold, odour discrimination, and odour identification. The composite odor threshold, discrimination and identification score (TDI) range from 1 to 48 points. The “Sniffin Sticks” test were conducted after participants had rated on the 9-point scales regarding their subjective evaluation of olfactory performances. This was to exclude the influence of psychophysical olfactory testing on the attention towards olfactory function among participants9. The Sniffin’ Sticks test took place in a quiet and well-ventilated room of the MRI facilities at TU Dresden immediately before MRI.
Importance of Olfaction assessment
In addition to the Sniffin’ Sticks test, participants were asked about their individual significance of olfaction with the importance of olfaction (IOQ) questionnaire27. The 20-item IOQ consists of three subscales evaluating the emotions, memories and evaluations associated with odors (Association-scale), the degree of application of smells in daily life (Application-scale), and the importance of smells in decision making (Consequence-scale). Repectively, 6 items define 1 of 3 subscales (“application”, “association”, “consequence”). The 2 remaining items relate to the scale named “aggravation,”which captures the tendency to exaggerate the importance of olfaction. Participants answered each statement on a 4-point Likert scale (I totally agree, I mostly agree I mostly disagree, I totally disagree).The scale exhibits a good internal reliability (Cronbach’s α = 0.77) which capture the subjective awareness and attitude towards chemosensory stimuli, and is suitable for participants with normal or abnormal olfactory functions.
Structural brain image acquisition
High-resolution T1 anatomical brain images were acquired on a 3 T Siemens Sonata scanner (model Trio, Siemens, Erlangen, Germany) with a 32-channel phased-array head coil. For each subject, the axial T1 weighted images (in total 192 slices) were acquired using a 3-dimensional magnetization-prepared rapid acquisition gradient echo (MPRAGE) sequence with the following parameter: voxel size: 0.73 × 0.73 × 1.0 mm; repetition time: 2180 ms; echo time: 3.93 ms; flip angle: 15°, field of view: 384 mm.
Voxel-based morphometry
Voxel-based morphometry of T1 weighted images was performed using the CAT12 software (https://dbm.neuro.uni-jena.de/vbm/) implemented in SPM12 (Wellcome Centre of Imaging Neuroscience, Institute of Neurology, UCL, London, UK; https://www.fil.ion.ucl.ac.uk/spm) ran with MATLAB (version 2013a, The MathWorks, Natick, MA, USA). T1 images were first segmented into gray matter (GM), white matter (WM) and cerebrospinal fluid (CSF). The GM images were spatially normalized to a template in Montreal Neurological Institute (MNI) space using the high dimensional Diffeomorphic Anatomical Registration Through Exponentiated Lie Algebra (DARTEL)28. The quality control (homogeneity check) in CAT12 was applied to ensure high quality of the segmentation of GM from WM. Mean correlation of the GM segments (the unsmoothed segmentations that provide more anatomical details were used) across all participants were performed as a measure of image quality after pre-processing. This tool visualizes the correlation between the volumes using a boxplot and correlation matrices. Outliers (deviated greater than 2 standard deviations from the global mean) are indicated and were excluded from the sample before smoothing and statistical analyses. For both groups, no data was indicated as having a mean correlation more than 2 standard deviations (supplementary Figure S1). Finally, the normalized GM images were smoothed with a Gaussian kernel (full width at half maximum 8 mm). Automated data quality checks were performed as per the CAT12 toolbox. An absolute threshold masking value of 0.2 was applied to avoid possible edge effects between different tissue types28. The volumes of GM, WM and CSF of each participant were summed up to the total intracranial volume (TIV).
Gray matter volume (GMV) between the SH and SN groups was examined using the two-sample t-test model in SPM12, including sex, age, TDI score and TIV as covariates. The multiple regression approach implemented in SPM12 was used for correlation analyses between GMV and the following variables: Sniffin’ Sticks test scores, IOQ scores, and self-ratings of olfactory performances. Age, sex and TIV were included as covariates of interest in all the multiple regression models. Besides, for the correlation analysis between GMV and TDI score, self-ratings for olfactory performance was included as a covariate, in order to eliminate the group effect.
A voxel-intensity tests was conducted within the regions related to olfactory and emotional processing, including the amygdala, the orbitofrontal cortex (OFC), the insula and the hippocampus (Regions-of-interest [ROI] analyses). Anatomical masks of ROIs were defined from the WFU Pickatlas toolbox (ANSIR, Wake Forest University, Winston-Salem, NC, USA)29 based on the “automated anatomical labelling (AAL)” atlas30. The OFC mask was defined as follows: bilateral Frontal_Mid_Orb + bilateral Frontal_Inf_Orb + bilateral Front_Sup_Orb. Results were considered as significant with peak wise p < 0.05 family wise error corrected (pFWEcorrected < 0.05) after small volume correction across each individual ROI mask. All ROIs were tested simultaneously for the left and right hemispheres. We did not correct each individual ROI for multiple testing to minimize Type II errors.
In addition, the aforementioned analyses were also conducted on the whole-brain level. To control for multiple statistical testing, we maintained a cluster-level false-positive detection rate at p < 0.05 using a voxel-level threshold of p < 0.001 with a cluster extent (k) empirically determined by Monte Carlo simulations (n = 1000 iterations), by means of AlphaSim procedure31, implemented in the REST toolbox (https://www.restfmri.net/forum/REST_V1.7)32. With an initial uncorrected voxel-wise threshold of 0.001, the following cluster size was determined to achieve a cluster-level FWE corrected p < 0.05: k = 32 voxels (for comparison between groups), k = 23 voxels (for correlation between GMV and TDI score) and k = 24 voxels (for correlation between GMV and subjective olfactory ratings). Significant brain regions were labelled and reported with the AAL toolbox30.
Statistical analysis
Statistical analyses were performed by means of SPSS (Version 24.0, SPSS Inc. Chicago, IL, USA) and GraphPad Prism (Version 6, GraphPad Software Inc., La Jolla, Ca, USA). The two-sample t-test was used to compare the age between groups. Chi-square test was used to compare the sex distributions. One-way ANOVA was used to compare the subjective ratings for olfactory function, the Sniffin’ Sticks test scores, and the importance of olfaction subscale scores, with age and sex included as covariates of interest. Data are presented as mean ± standard deviations (SD) and results were considered as significant with p-values below 0.05.
Results
Demographics and olfactory function
Olfactory performances measured by the Sniffin’ Sticks test showed no significant difference between the SH and SN groups. Among the SH group, seven participants rated their sense of smell as “moderately better (score 6)”, four participants as “much better (score 7)”, and seven participants as “very well (score 8)”. The importance of smell did not differ between SH and SN groups (Table 1).
Group comparison of GMV
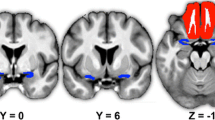
The ROI analyses showed significantly enhanced GMV in the left hippocampus (cluster peak coordinates − 30 − 17 − 23; T = 4.95; pFWEcorrected = 0.015, k = 131 voxels, Fig. 1) in SH group relative to the SN group, no significant result was observed for the SH < SN contrast. The whole-brain analysis further revealed significantly larger GMV of the right hippocampus, right hypothalamus, left superior frontal gyrus and left precuneus among the SH group compared to the SN group (corrected p < 0.05; Table 2 and Fig. 2). For the reversed contrast (SN > SH) revealed a cluster in the right cerebellum (Table 2 and Fig. 2).

Larger gray matter volume of the left anterior hippocampus (pFWEcorrected < 0.05) in subjective hyperosmia as compared to subjective normosmia groups. Activation is visualized on a template provided by MRIcron (www.nitrc.org/projects/mricron). The T-map threshold was set at puncorrected ≤ 0.001 for visualization purpose.

Whole-brain results (puncorrected < 0.001 and cluster size ≥ 32 voxels) of the gray matter volume (GMV) difference between subjective hyperosmia (SH) and subjective normosmia (SN) groups. Red clusters indicate larger GMV for SH > SN; Blue cluster indicates larger GMV for SN > SH. Activation of the significant cluster is visualized on a template provided by MRIcron (www.nitrc.org/projects/mricron).
Correlation between regional GMV and other variables
For all participants (N = 32), the ROI analyses found significant positive correlation between the GMV of the left hippocampus (peak coordinates − 32 − 12 − 23; T = 4.57; pFWEcorrected = 0.034, k = 51 voxels) and TDI scores. Extracted GMV from this significant cluster were found correlated to Sniffin’ Sticks score for odor threshold (r = 0.61, p < 0.001) and odor discrimination (r = 0.43, p = 0.020), but not for odor identification (r = − 0.07; p = 0.74) (Fig. 3). Whole-brain analyses showed positive correlation between the TDI score and the GMV of the left hippocampus the left cerebellum, and the right middle temporal gyrus (Table 3 and Fig. 4). Negative association between TDI score and GMV was observed for the right cerebellum and right superior frontal gyrus (Table 3 and Fig. 4).

Correlations between the left hippocampal gray matter volume (cluster peak coordinates − 32 − 12 − 23; T = 4.57; pFWEcorrected = 0.034, k = 51 voxels) and the Sniffin’ Sticks odor threshold and discrimination score. Activation of significant cluster is visualized on a template provided by MRIcron (www.nitrc.org/projects/mricron). The T-map was threshold was set at puncorrected ≤ 0.001 for visualization purpose.

Whole-brain results (puncorrected < 0.001 and cluster size ≥ 24 voxels) of the gray matter volume showing correlation with Sniffin’ Sticks test scores among the whole study sample (N = 32). Brain regions with positively correlations are depicted in red, and with negative correlations are depicted in blue. Activation of the significant cluster is visualized on a template provided by MRIcron (www.nitrc.org/projects/mricron).
For the SH participants (N = 18) and with the whole-brain analysis, the self-ratings for olfactory performance was positively associated with the GMV in the bilateral cerebellum, the right superior frontal gyrus, and the left precentral gyrus; and negatively associated with GMV of the right putamen and the right angular gyrus (Table 4 and Fig. 5). No significant cluster was observed with ROI analyses.

Whole-brain results (puncorrected < 0.001 and cluster size ≥ 24 voxels) of the gray matter volume showing correlation with subjective ratings of olfaction among the subjective hyperosmia group (N = 18). Brain regions with positively correlations are depicted in red, and with negative correlations are depicted in blue. Activation of the significant cluster is visualized on a template provided by MRIcron (www.nitrc.org/projects/mricron).
Discussion
The current study compared the regional gray matter volume (GMV) between subjective hyperosmia (SH) and subjective normosmia (SN) participants. Results revealed a larger left anterior hippocampal GMV in SH as compared to SN, although the “Sniffin’ Sticks” test scores were not different between the two groups. The hippocampus is a key region involved in a variety of olfactory processes, including odor discrimination33, odor familiarity processing34, episodic odor memory35 and odor-visual integration35,36. Following a coordinate-based segmentation for the hippocampus, the cluster showing increased GMV among SH participants from the current study was part of the anterior hippocampus (anterior to y = -21 mm in MNI space)37. There are substantial evidence showing that the subregions along the hippocampal anterior–posterior axis are associated with different functional specializations. The anterior part of the left hippocampal formation has been reported to specialize in emotional processing37,38,39, particularly the encoding of emotional memories40,41,42, as well as modality-independent memory retrieval43. Studies regarding the specific role of the anterior hippocampal subregions in olfactory process is limited. However, a coordinate search revealed that the anterior hippocampus is involved in cross-modal emotional processing of odor pleasantness44, as well as in olfactory-related emotional memory45, cross-modal recognition memory35,46.
It had been found that the self-rating of superior olfactory function correlated with higher degree of self-rated odor annoyance11, and a tendency to be influenced by odors in their liking (and not liking) for new stimuli than other participants47. Studies have shown the association between self-reported sensory sensitivity and structural change of the hippocampus. For example, self-reported pain sensitivity was positively associated with larger GMV in the hippocampus and parahippocampus, however no GMV changes was found in relation to the pressure pain threshold18. Neurotypical hyper-sensitivity to sounds, assessed with questionnaire, has been shown to be linked to increased GMV in the auditory cortex but also in limbic regions including the bilateral hippocampus17. Thus, a larger hippocampus may be associated with subjective sensory sensitivity in general. Historically, sensory sensitivity has been linked to a sensitive personality type that includes introversion and increased emotionality48. Hence, we may speculate that subjective sensory hypersensitivity is related to the ability to form the associations between sensory stimuli and emotional experiences, and the hippocampal GMV serve as a neural mediator in this association. However, the odor related annoyance or affective attitudes were not measured in our study, which limits further test on the functional role of the hippocampus. Besides, the cross-sectional design in those studies, makes it impossible to detect causal relationships. Perhaps, self-reported hyperosmic individuals are born with a predisposition for larger volumes of the hippocampus, leading them to be more prone to evaluate the emotional sensory stimuli. Inferring the functional meaning of local GMV alterations as detected by VBM is difficult although links between the GMV and task-related brain activation had been shown49. An intriguing question for future study is to investigate the hippocampal brain responses to emotional olfactory cues among SH people.
We also found a positive association between the GMV of the anterior hippocampus and odor sensitivity and discrimination scores. It is noted that the significant cluster was close to the one from the between-group comparison, suggesting the potential role of this part of the hippocampus in integrating olfactory perception, olfactory emotion and episodic olfactory memory retrieval. In an earlier study with 112 healthy participants, Smitka et al.50 found a significant correlation between the GMV of the right hippocampus and odor threshold. Recently, Wabnegger et al.20 showed that hyperosmic participants (“Sniffin’ Sticks” TDI score over 41) had larger GMV of the posterior subregion of the hippocampus (cluster peak coordinates posterior to y = − 21 mm in MNI space). Hence, both studies show positive associations between TDI score and GMV in the hippocampus. It was not clear from the Wabnegger’s paper which specific olfactory functions (e.g. odor threshold, discrimination and identification) was significantly correlated to the hippocampal GMV. It is possible that the anterior and posterior subregions of the hippocampus serve different roles in specific olfactory functions. The specific functions for the anterior–posterior hippocampal subregions could also explain the weak (although significant) correlation between olfactory functions and the whole right hippocampus GMV, and the null findings between olfaction and the left hippocampus GMV reported in the paper from Smitka et al.50.
Beyond the predicted regions of interest, the SH group also showed increased GMV in the left precuneus, left superior frontal gyrus and the right hypothalamus. Precuneus is involved in odor-evoked autobiographical memory, mental imagery or olfactory memory retrieval51,52. The hypothalamus, as a secondary olfactory area, receives projections from the primary olfactory cortex and appears to be involved in odor processing under emotional context35,53. However, there is lack of evidence supporting its role in subjective hyperosmia or general sensory sensitivity.
There are limitations to the current study. First, although the ROI analyses had controlled for multiple comparisons, it is necessary to validate the current research findings with a larger sample. Second, the variation in normalisation accuracy or smoothness during the VBM analyses is likely to result in statistical sensitivity varying over different brain regions, which is one major limitation for the VBM technique54. Thus, the absence of a statistically significant effect in any particular region does not prove that the region is unaffected. In addition, the null finding regarding the importance of olfaction between SH and SN groups may suggest the specificity of the questionnaire. Future study combining brain imaging techniques with psychometric assessment regarding the olfaction related affective responses (e.g. the affective impact of odor scale) or odor awareness (e.g. the odor awareness scale) should obtain deeper knowledge about the neurobehavioral basis of subjective hyperosmia.
In conclusion, the current study suggested that self-rated hyperosmia is associated with increased GMV in the anterior hippocampus. The GMV of this region also showed positive correlation to the odor threshold and odor discrimination scores. Future research should elaborate on the neural basis for emotional and perceptual olfactory processing.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. The study was approved by the Ethics Committee at the TU Dresden (Protocol # EK262082010).
Informed consent
Informed consent was obtained from all individual participants included in the study.
References
Hummel, T. et al. Position paper on olfactory dysfunction. Rhinology 56, 1–30 (2016).
Oleszkiewicz, A., Schriever, V. A., Croy, I., Hahner, A. & Hummel, T. Updated Sniffin’ Sticks normative data based on an extended sample of 9139 subjects. Eur. Arch. Otorhinolaryngol. 276, 719–728 (2019).
Menashe, I. et al. Genetic elucidation of human hyperosmia to isovaleric acid. PLoS Biol. 5, e284 (2007).
Gilbert, A. N. & Kemp, S. E. Odor perception phenotypes: multiple, specific hyperosmias to musks. Chem. Senses 21, 411–416 (1996).
Kollndorfer, K. et al. The inability to self-evaluate smell performance. How the vividness of mental images outweighs awareness of olfactory performance. Front Psychol. 6, 627 (2015).
Welge-Luessen, A., Hummel, T., Stojan, T. & Wolfensberger, M. What is the correlation between ratings and measures of olfactory function in patients with olfactory loss?. Am. J. Rhinol. 19, 567–571 (2005).
Nguyen, D. T., Nguyen-Thi, P. L. & Jankowski, R. How does measured olfactory function correlate with self-ratings of the sense of smell in patients with nasal polyposis?. Laryngoscope 122, 947–952 (2012).
Lotsch, J. & Hummel, T. Clinical usefulness of self-rated olfactory performance-a data science-based assessment of 6000 patients. Chem. Senses 44, 357–364 (2019).
Landis, B. N., Hummel, T., Hugentobler, M., Giger, R. & Lacroix, J. S. Ratings of overall olfactory function. Chem. Senses 28, 691–694 (2003).
Philpott, C. M., Wolstenholme, C. R., Goodenough, P. C., Clark, A. & Murty, G. E. Comparison of subjective perception with objective measurement of olfaction. Otolaryngol. Head Neck Surg. 134, 488–490 (2006).
Knaapila, A. et al. Self-ratings of olfactory function reflect odor annoyance rather than olfactory acuity. Laryngoscope 118, 2212–2217 (2008).
Knaapila, A. & Tuorila, H. Experiences of environmental odors among self-reported hyperosmics: a pilot study. J. Health Psychol. 19, 897–906 (2014).
Wrzesniewski, A., McCauley, C. & Rozin, P. Odor and affect: individual differences in the impact of odor on liking for places, things and people. Chem. Senses 24, 713–721 (1999).
Ward, J. Individual differences in sensory sensitivity: a synthesizing framework and evidence from normal variation and developmental conditions. Cogn. Neurosci. 10, 139–157 (2019).
Green, S. A. et al. Neurobiology of sensory overresponsivity in youth with autism spectrum disorders. JAMA Psychiatry 72, 778–786 (2015).
Green, S. A. et al. Overreactive brain responses to sensory stimuli in youth with autism spectrum disorders. J. Am. Acad. Child Adolesc. Psychiatry 52, 1158–1172 (2013).
Kliuchko, M. et al. Neuroanatomical substrate of noise sensitivity. Neuroimage 167, 309–315 (2018).
Ruscheweyh, R., Wersching, H., Kugel, H., Sundermann, B. & Teuber, A. Gray matter correlates of pressure pain thresholds and self-rated pain sensitivity: a voxel-based morphometry study. Pain 159, 1359–1365 (2018).
Banks, S. J. et al. Structural and functional MRI differences in master sommeliers: a pilot study on expertise in the brain. Front Hum. Neurosci. 10, 414 (2016).
Wabnegger, A., Schlintl, C., Hofler, C., Gremsl, A. & Schienle, A. Altered grey matter volume in “super smellers”. Brain Imaging Behav. 13, 1726–1732 (2019).
Gottfried, J. A. Central mechanisms of odour object perception. Nat. Rev. Neurosci. 11, 628–641 (2010).
Seubert, J., Freiherr, J., Djordjevic, J. & Lundstrom, J. N. Statistical localization of human olfactory cortex. Neuroimage 66, 333–342 (2013).
Fjaeldstad, A. et al. Brain fingerprints of olfaction: a novel structural method for assessing olfactory cortical networks in health and disease. Sci. Rep. 7, 42534 (2017).
Kadohisa, M. Effects of odor on emotion, with implications. Front Syst. Neurosci. 7, 66 (2013).
Oldfield, R. C. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9, 97–113 (1971).
Hummel, T., Sekinger, B., Wolf, S. R., Pauli, E. & Kobal, G. “Sniffin” Sticks’: Olfactory performance assessed by the combined testing of odor identification, odor discrimination and olfactory threshold. Chem. Senses 22, 39–52 (1997).
Croy, I., Buschhuter, D., Seo, H. S., Negoias, S. & Hummel, T. Individual significance of olfaction: development of a questionnaire. Eur. Arch. Otorhinolaryngol. 267, 67–71 (2010).
Ashburner, J. & Friston, K. J. Voxel-based morphometry–the methods. Neuroimage 11, 805–821 (2000).
Maldjian, J. A., Laurienti, P. J., Kraft, R. A. & Burdette, J. H. An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. Neuroimage 19, 1233–1239 (2003).
Tzourio-Mazoyer, N. et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 15, 273–289 (2002).
Forman, S. D. et al. Improved assessment of significant activation in functional magnetic resonance imaging (fMRI): use of a cluster-size threshold. Magn. Reson. Med. 33, 636–647 (1995).
Song, X. W. et al. REST: a toolkit for resting-state functional magnetic resonance imaging data processing. PLoS ONE 6, e25031 (2011).
Kareken, D. A., Mosnik, D. M., Doty, R. L., Dzemidzic, M. & Hutchins, G. D. Functional anatomy of human odor sensation, discrimination, and identification in health and aging. Neuropsychology 17, 482–495 (2003).
Plailly, J. et al. Involvement of right piriform cortex in olfactory familiarity judgments. Neuroimage 24, 1032–1041 (2005).
Gottfried, J. A., Smith, A. P., Rugg, M. D. & Dolan, R. J. Remembrance of odors past: human olfactory cortex in cross-modal recognition memory. Neuron 42, 687–695 (2004).
Gottfried, J. A. & Dolan, R. J. The nose smells what the eye sees: crossmodal visual facilitation of human olfactory perception. Neuron 39, 375–386 (2003).
Poppenk, J., Evensmoen, H. R., Moscovitch, M. & Nadel, L. Long-axis specialization of the human hippocampus. Trends Cogn. Sci. 17, 230–240 (2013).
Robinson, J. L. et al. Neurofunctional topography of the human hippocampus. Hum. Brain. Mapp. 36, 5018–5037 (2015).
Robinson, J. L., Salibi, N. & Deshpande, G. Functional connectivity of the left and right hippocampi: Evidence for functional lateralization along the long-axis using meta-analytic approaches and ultra-high field functional neuroimaging. Neuroimage 135, 64–78 (2016).
Murty, V. P., Ritchey, M., Adcock, R. A. & LaBar, K. S. fMRI studies of successful emotional memory encoding: a quantitative meta-analysis. Neuropsychologia 48, 3459–3469 (2010).
Dolcos, F., LaBar, K. S. & Cabeza, R. Interaction between the amygdala and the medial temporal lobe memory system predicts better memory for emotional events. Neuron 42, 855–863 (2004).
Richardson, M. P., Strange, B. A. & Dolan, R. J. Encoding of emotional memories depends on amygdala and hippocampus and their interactions. Nat. Neurosci. 7, 278–285 (2004).
Lehn, H., Kjonigsen, L. J., Kjelvik, G. & Haberg, A. K. Hippocampal involvement in retrieval of odor vs. object memories. Hippocampus 23, 122–128 (2013).
Bensafi, M. et al. Cross-modal integration of emotions in the chemical senses. Front. Hum. Neurosci. 7, 883 (2013).
Herz, R. S., Eliassen, J., Beland, S. & Souza, T. Neuroimaging evidence for the emotional potency of odor-evoked memory. Neuropsychologia 42, 371–378 (2004).
Ghio, M., Schulze, P., Suchan, B. & Bellebaum, C. Neural representations of novel objects associated with olfactory experience. Behav. Brain Res. 308, 143–151 (2016).
Knaapila, A., Raittola, A., Sandell, M. & Yang, B. Self-ratings of olfactory performance and odor annoyance are associated with the affective impact of odor, but not with smell test results. Perception 46, 352–365 (2017).
Aron, E. N. & Aron, A. Sensory-processing sensitivity and its relation to introversion and emotionality. J. Pers. Soc. Psychol. 73, 345–368 (1997).
Takeuchi, H. et al. Associations among imaging measures (2): the association between gray matter concentration and task-induced activation changes. Hum. Brain. Mapp. 35, 185–198 (2014).
Smitka, M. et al. Is there a correlation between hippocampus and amygdala volume and olfactory function in healthy subjects?. Neuroimage 59, 1052–1057 (2012).
Arshamian, A. et al. The functional neuroanatomy of odor evoked autobiographical memories cued by odors and words. Neuropsychologia 51, 123–131 (2013).
Zhang, S. & Li, C. S. Functional connectivity mapping of the human precuneus by resting state fMRI. Neuroimage 59, 3548–3562 (2012).
Royet, J. P. et al. Emotional responses to pleasant and unpleasant olfactory, visual, and auditory stimuli: a positron emission tomography study. J. Neurosci. 20, 7752–7759 (2000).
Ridgway, G. R. et al. Ten simple rules for reporting voxel-based morphometry studies. Neuroimage 40, 1429–1435 (2008).
Funding
Open Access funding enabled and organized by Projekt DEAL. The study was supported by the Hans Skouby Foundation and Aase and Ejnar Danielsens Foundation. The sponsors had no influence on the study design, data collection, writing of the manuscript, or the decision to publish.
Author information
Authors and Affiliations
Contributions
P.H.: Conceptualization, Data curation, Formal analysis, Writing - original draft. F.P.S.-S.: Data curation. A.F.: Conceptualization, Writing - review & editing. T.H.: Conceptualization, Funding acquisition, Writing - review & editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Han, P., Stiller-Stut, F.P., Fjaeldstad, A. et al. Greater hippocampal gray matter volume in subjective hyperosmia: a voxel-based morphometry study. Sci Rep 10, 18869 (2020). https://doi.org/10.1038/s41598-020-75898-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-75898-6
- Springer Nature Limited
This article is cited by
-
Post-COVID-19 conditions: a systematic review on advanced magnetic resonance neuroimaging findings
Neurological Sciences (2024)