
Abstract
A. hypochondriacus leaves contained ample phytopigments including betalain, anthocyanin, β-xanthin, β-cyanin, and bioactive phytochemicals of interest in the industry of food. We have been evaluating the possibility of utilizing phytopigments of amaranth and bioactive constituents for making drinks. Therefore, we evaluated bioactive phytopigments and compounds including the potentiality of antioxidants in A. hypochondriacus leaves. A. hypochondriacus leaves have abundant protein, carbohydrates, and dietary fiber. We found considerable levels of inorganic minerals including magnesium, calcium, potassium (3.88, 3.01, 8.56 mg g−1), zinc, manganese, copper, iron (16.23, 15.51, 2.26, 20.57 µg g−1), chlorophyll b, chlorophyll ab chlorophyll a (271.08, 905.21, 636.87 μg g−1), scavenging capacity of radicals (DPPH, ABTS+) (33.46, 62.92 TEAC μg g−1 DW), total polyphenols (29.34 GAE μg g−1 FW), β-xanthin, betalain, β-cyanin (584.71, 1,121.93, 537.21 ng g−1), total flavonoids (170.97 RE μg g−1 DW), vitamin C, β-carotene, carotenoids (184.77, 82.34, 105.08 mg 100 g−1) in A. hypochondriacus leaves. The genotypes AHC6, AHC4, AHC11, AHC5, and AHC10 had a good scavenging capacity of radicals. Polyphenols, phytopigments, flavonoids, and β-carotene of A. hypochondriacus had potential antioxidant activity. Extracted juice of A. hypochondriacus can be an ample source of phytopigments and compounds for detoxification of reactive oxygen species (ROS) and attaining nutritional and antioxidant sufficiency.
Similar content being viewed by others


Introduction
In the globe, 795 million people are affected by a continuous deficit in calories for the scarcity of sufficient foods1. Around 2 billion people are affected by the deficiency of vitamins or minerals2. The main source of energy in the human diet is the consumption of staple foods regularly even though these are deficient in micronutrients, such as iodine, iron, zinc, pro-vitamin A, vitamin E, vitamin C, and carotenoids3. As a result, continuous consumption of staple foods in the human diet, consequently resulting in hidden hunger2. Hence, we can ensure a balanced and healthy diet by consuming vegetables and fruits as the occurrence of minerals and vitamins along with staple foods. Furthermore, we protect our health and reduce the risk of cancer, cardiovascular disease, and many chronic diseases by feeding fruits and vegetables. Bioactive constituents, such as nutrients, phenolics, pigments, flavonoids, and vitamins contribute to many health benefits4,5,6.
Amaranthus genus has a C4 photosynthetic pathway that is distributed widely in America, Africa, Australia, Asia, and Europe. Among the seventy species, seventeen are cultivated as edible leafy vegetables and three are cultivated as grain amaranths7. Amaranthus contains thirteen times more ascorbic acid, twenty times more calcium, eighteen times more pro-vitamin A, and seven times more Fe in comparison with lettuce8. It contains protein with essential amino acids including methionine and lysine, carotenoids, dietary fiber, vitamin C, minerals9,10,11,12,13,14,15. It has abundant phytopigments including betalain, anthocyanin, β-cyanin, carotenoids, β-xanthin, and chlorophylls16,17, bioactive compounds including flavonoids, vitamin C, phenolic acids, carotenes18. The phytopigments and bioactive constituents quenching ROS and had a magnificent contribution to the industry of food19,20,21. Amaranth is a unique source of betalain, β-xanthin, and β-cyanin. Amaranths have multipurpose uses and widely acclimated vegetables to different abiotic stresses including drought22,23,24,25 and salinity26,27,28.
The underutilized leafy vegetables, A. hypochondriacus originates from North America, possibly by the hybridization of wild A. powellii (North American) and A. cruentus (cultivated)29. Now, A. hypochondriacus is extensively cultivated throughout the world including tropical, subtropical and temperate climates. It is used as vegetables, grains, and ornamental plants. It has great diversity and phenotypic plasticity30. Mild flavored juvenile leaves and edible fleshy stems of A. hypochondriacus are popularly used as leafy vegetables in India, Bangladesh, Asia, and Africa due to nutritive values and taste. A. hypochondriacus contains tannin and has astringent flavor. Traditionally, it is used internally for the treatment of diarrhea, excessive menstruation, gargle to soothe inflammation of the pharynx, enhance the healing of ulcerated mouths and externally for nosebleeds and wounds. Yellow, red, and green natural pigments can be used as colorants in foods and medicines29.
Currently, researchers and consumers are very much interested in natural antioxidants of vegetables. Natural antioxidants of amaranth include phytopigments (carotenoids, chlorophyll, betaxanthin, and betacyanin), phenolics, ascorbic acids, and flavonoids19,20. These natural bioactive constituents protect us from neurodegenerative diseases, cancer, cardiovascular diseases, atherosclerosis, cataracts, emphysema, arthritis, and retinopathy5,31,32. These products containing antioxidant properties have a substantial interest to consumers. Many medicinal plants containing phenolics, vitamins C, carotenoids, flavonoids and other non-nutrient constituents are potentially used as an antioxidant with a protective capacity33.
Recently, we have been evaluating the possibility of utilizing phytopigments of amaranth containing abundant natural β-xanthin, betalain, β-cyanin, and bioactive compounds of interest in the industry of food16,17. It is the first attempt to evaluate nutrients, bioactive components and scavenging capacity of radicals in A. hypochondriacus. Hence, in the current investigation was carried out to evaluate the occurrence of bioactive constituents, and scavenging capacity of radicals of A. hypochondriacus in detail and study the possibility of A. hypochondriacus genotypes for making drinks containing high antioxidant constituents and antioxidant activity.
Results and discussion
Proximate compositions
Table 1 represents the proximate compositions of A. hypochondriacus genotypes. The genotype AHC11 (885.75 g kg−1 FW) exhibited the highest content of moisture, whereas the lowest moisture content was obtained from AHC4 (817.85 g kg−1 FW). The content of moisture ranged from 817.85 to 885.75 g kg−1 FW. As lower moisture contents ensured higher dry matter, the genotype AHC4, AHC8, AHC1, and AHC7 had 16–19% dry matter could be selected for dry matter contents. The content of moisture of A. hypochondriacus leaves directly associated with the maturity of the plant. Our obtained results were corroborated with the findings of sweet potato leaves34. The remarkable variations in protein content were noticed in A. hypochondriacus leaves. The highest protein content was noticed in the genotype AHC2 (61.62 g kg−1) followed by that of AHC10, AHC1, AHC6, and AHC11, whereas the lowest protein content was recorded in the genotype AHC8 (47.41 g kg−1). Across all genotypes, five genotypes exhibited better protein content over their average value. The genotypes AHC2, AHC10, AHC1, AHC6, and AHC11 had relatively high protein contents for a leafy vegetable. A. hypochondriacus provides important roles as a protein source for vegetarians and many people in low-income countries. A. hypochondriacus (54.18 g kg−1) had much higher protein content in comparison with A. tricolor of our earlier study12. AHC2 and AHC7 exhibited the highest fat content (4.44, 4.27 g kg−1 FW) showing the order of AHC2 = AHC7 ˃ AHC10 ˃ AHC5. The genotype AHC3 exhibited the lowest fat content (2.43 g kg−1 FW) which was statistically similar to AHC6, AHC8, AHC1, AHC9, AHC4, and AHC11 with an average of 3.09 g kg−1 FW. The fat content in the current investigation agreed to the fat content of sweet potato leaves by Sun et al.34. It is known that fat influences the cell function, covering the organs and upholding the temperature of the body. Fats play a vital role in digestion, absorption, and transport of vitamins A, E, K, and D that are soluble in fats.
The highest carbohydrates contents (70.74 g kg−1 FW) were recorded in AHC4 followed by that of AHC8 and AHC7, whereas AHC11 exhibited the lowest carbohydrates contents (1.95 g kg−1 FW) including an average of 46.90 g kg−1 FW. The highest energy (58.47 kcal 100 g−1) was found in genotype AHC4, followed by that of AHC8, AHC1, AHC6, and AHC7, while the genotype AHC5 and AHC3 (37.26, 38.15 kcal 100 g−1) showed the lowest energy including an average of 45.74 kcal 100 g−1 FW. AHC4 (56.22 g kg−1 FW) exhibited the highest ash content followed by that of AHC11, AHC8, and AHC1, while AHC9, AHC5, and AT17 had the lowest ash content (36.44, 36.46, 37.28 g kg−1 FW, respectively) including a mean value of 42.85 g kg−1 FW.
The remarkable variations were noticed in the content of the digestive fiber of 11 A. hypochondriacus studied. The highest dietary fiber was noticed in AHC2 and AHC6 (10.27, and 10.03 g 100 g−1) followed by AHC4, while AHC11 had the lowest dietary fiber contents (7.26 g 100 g−1) which was statistically similar to the genotype AHC8 and AHC7 with an average of 8.73 g 100 g−1. For mitigation of palatability, food digestibility, and remedy of constipation, dietary fiber has a crucial role9. In this study; we observed that A. hypochondriacus has abundant protein, carbohydrates, dietary fiber, and moisture. The moisture contents observed in A. hypochondriacus leaves was greater than our previous studies of red morph amaranth35, weedy amaranth36, green morph amaranth37, stem amaranth38, while the moisture contents of A. hypochondriacus leaves were corroborated with our previous studies of A. blitum39. In contrast, the protein contents observed in A. hypochondriacus leaves were greater than our previous studies of red morph amaranth35, A. viridis weedy amaranth36, green morph amaranth37, stem amaranth38, and A. blitum39, while the protein contents of A. hypochondriacus leaves were corroborated with our previous studies of A. spinosus weedy amaranth36. However, The carbohydrates contents observed in A. hypochondriacus leaves were greater than our previous studies of A. spinosus weedy amaranth36, while the carbohydrates contents of A. hypochondriacus leaves were lower than red morph amaranth35, green morph amaranth37, A. viridis weedy amaranth36, stem amaranth38, and A. blitum39. The digestible fiber obtained from A. hypochondriacus leaves were greater than our previous studies of red morph amaranth35, green morph amaranth37, stem amaranth38, and A. blitum39, while digestible fiber obtained from A. hypochondriacus leaves were lower than our previous studies of weedy amaranth36.
Macro and microelements compositions
The macro and microelements composition of A. hypochondriacus is presented in Table 2. In this study, the K content ranged from 5.19 mg g−1 to 8.56 mg g−1 FW. The lowest K content was recorded in the genotypes AHC3, AHC6, AHC1, AHC9, AHC4, AHC11, AHC7, AHC2, and AHC10, while the highest K content was exhibited in the genotype AHC8 followed by that of AHC5, with a grand mean value of 5.74 mg g−1 FW. Two genotypes had higher K contents than the mean K content. The highest Ca content was noted in the genotype AHC8 (3.01 mg g−1), followed by that of AHC2, whereas, the lowest Ca content was reported in the genotypes AHC4 (1.89 mg g−1) with an average of 2.43 mg g−1 FW. Six accessions exhibited high Ca contents than the corresponding mean. The highest magnesium content was recorded in the genotype AHC8 (3.88 mg g−1) and the lowest magnesium content was observed in AHC4 (3.20 mg g−1), including an average of 3.41 mg g−1 FW. In the current investigation, Mg content had not varied considerably among the genotypes (3.20 to 3.88 mg g−1 FW). Our results showed that A. hypochondriacus genotypes (FW basis) had remarkable Ca (3.01 mg g−1), K (8.56 mg g−1), and Mg (3.88 mg g−1) which was more pronounced than Mg, Ca, and K content of different amaranth species including A. hypochondriacus of Jimenez-Aguiar and Grusak40. Jimenez-Aguir and Grusak40 observed that different amaranth species including A. hypochondriacus had abundant Mg, Ca, and K in comparison with spider flower, kale, black nightshade, and spinach. K content of A. hypochondriacus leaves was greater than the K content of our previous studies of green morph amaranth37, while K content obtained from A. hypochondriacus leaves was lower than the K content of our previous studies of weedy amaranth36. Ca content observed in the current investigation were corroborated with green morph amaranth37 and weedy amaranth36. Mg obtained from A. hypochondriacus leaves were greater than the Mg content of our previous studies of green morph amaranth37 and A. spinosus36, while Mg obtained from A. hypochondriacus leaves were corroborated with the Mg content of our previous studies of A. viridis36.
The iron content of A tricolor had the significant and pronounced variations regarding genotypes (7.48 µg g−1 FW in AHC7 to 20.57 µg g−1 FW in AHC10). High iron content was recorded in the genotypes AHC10, AHC5, AHC8, and AHC6. On the contrary, the lowest content of iron was recorded in the genotype AHC3, AHC1, AHC9, and AHC4, including an average of 12.75 µg g−1 FW. Four accessions exhibited higher iron contents than their corresponding average value. The highest manganese content was noted in the genotype AHC8 (15.51 µg g−1 FW), while the lowest manganese content was noted in the genotype AHC10 (5.77 µg g−1 FW) with an average of 9.58 µg g−1 FW. The genotypes AHC5, AHC8, AHC3, and AHC1 had high manganese content. The genotypes differed remarkably in copper content (0.90–2.26 µg g−1 FW). The highest copper content was noted in AHC9 and AHC5 (2.26 and 2.11 µg g−1 FW) followed by AHC10. Four accessions exhibited greater Cu contents than the mean value. The significant and remarkable variations were noted in the zinc content of the studied genotypes (6.40 µg g−1 FW (AHC10) to 16.23 µg g−1 FW (AHC5). Four genotypes showed better zinc contents than the corresponding mean (9.98 µg g−1 FW). Iron and zinc content of A. hypochondriacus was greater than the cassava leaves41 and beach pea42. We observed considerable Mn (15.51 µg g−1), Fe (20.57 µg g−1), Zn (16.23 µg g−1), and Cu (2.26 µg g−1) in A. hypochondriacus (fresh weight basis). Similarly, Jimenez-Aguiar and Grusak40 noted abundant Cu, Fe, Zn, and Mn (FW basis) in various amaranth species with A. hypochondriacus. They reported higher Cu, Fe, Mn, and Zn in different amaranth species including A. hypochondriacus than spider flower, spinach, black nightshade, and kale. Mn content noticed in the current investigation was much lower than our previous studies of green morph amaranth37 and A. spinosus weedy amaranth36, while it corroborated with our previous findings of A. viridis weedy amaranth36. Fe content observed in the current investigation were much greater than our previous studies of green morph amaranth37 and lower than our previous studies of weedy amaranth36. Cu and Zn contents observed in the current investigation was corroborated with our previous studies of green morph amaranth37, while Cu and Zn contents obtained from A. hypochondriacus leaves were lower than our previous studies of weedy amaranth36.
Composition of phytopigments
Table 3 represents the composition of phytopigments of A. hypochondriacus genotypes studied. The chlorophyll a content (131.07–636.87 μg g−1 FW) exhibited prominent variations among genotypes. The highest content of chlorophyll a (636.87 μg g−1 FW) was noted in the genotype AHC4, while the genotype AHC9 exhibited the lowest content of chlorophyll a (131.07 μg g−1 FW). The high content of chlorophyll a was noted in the genotypes AHC2 and AHC11. Six genotypes had a higher content of chlorophyll a than the mean value. Similar to chlorophyll a, chlorophyll b content also had significant and progressive variations among 11 A. hypochondriacus genotypes studied (71.27–271.08 μg g−1 FW). The highest chlorophyll b content (271.08 μg g−1 FW) was recorded in AHC7, followed by that of AHC4 and AHC11. Conversely, the lowest chlorophyll b content was noticed in the genotype AHC9 (71.27 μg g−1 FW). The content of chlorophyll ab showed significant and remarkable variations (193.98 to 905.21 μg g−1 FW). The genotypes AHC4, AHC11, AHC5, and AHC6 exhibited high chlorophyll ab content, whereas, the genotype AHC8 exhibited the lowest chlorophyll ab content (193.98 μg g−1 FW). Six accessions had greater chlorophyll ab contents than the mean value. Our studied A. hypochondriacus genotypes had considerable chlorophyll ab (905.21 μg g−1 FW), chlorophyll a (636.87 μg g−1 FW), and chlorophyll b (271.08 μg g−1 FW) contents which were greater than the content of chlorophylls of red and green amaranth of Khanam and Oba43. Chlorophyll a, chlorophyll ab, and chlorophyll b content in the present study were much greater than chlorophyll a, chlorophyll ab, and chlorophyll b content of green morph amaranth37, red morph amaranth35, stem amaranth38, and weedy amaranth36 of our earlier study, while these photosynthetic pigments content of this study were lower corroborated with our earlier studies of A. blitum39.
The highest content of β-cyanin was noted in the genotype AHC6 (537.21 ng g−1 FW) followed by that of AHC11 and AHC5, with an average value of 384.23 ng g−1 FW. On the contrary, the lowest β-cyanin content was observed in the genotype AHC9 (233.87 ng g−1 FW). The β-xanthin content had significant and considerable variations regarding genotypes which ranged from 181.90 to 584.71 ng g−1 FW. The genotype AHC6 exerted the highest β-xanthin content (584.71 ng g−1 FW) and the genotypes AHC11, AHC5, and AHC10 had high β-xanthin content. In contrast, the lowest β-xanthin content was recorded in the genotype AHC8 (181.90 ng g−1 FW). Six accessions exerted greater β-xanthin contents than the mean value. There were pronounced variations in the betalain content of the genotypes studied. The highest betalain content was observed in the genotype AHC6 (1,121.93 ng g−1 FW) and ranged from 367.42 to 1,121.93 ng g−1 FW. The high betalain content was noted in the genotype AHC11, AHC5, and AHC10. Whereas, the lowest content of betalain was recorded in the genotype AHC8 (367.42 ng g−1 FW). Seven accessions exerted greater betalain contents than the mean value. The highest carotenoid content was found in the genotype AHC10 (105.08 mg 100 g−1) and ranged from 32.77 mg 100 g−1 to 105.08 mg 100 g−1. The high carotenoids were recorded in the genotype AHC2, AHC1, and AHC3. Five accessions exerted greater carotenoids than the mean value. The considerable β-xanthin (584.71 ng g−1 FW), chlorophyll a (636.87 μg g−1 FW), chlorophyll ab (905.21 μg g−1 FW), chlorophyll b (271.08 μg g−1 FW), β-cyanin (537.21 ng g−1 FW), betalain (1,121.93 ng g−1 FW), and carotenoids (105.08 mg 100 g−1) were recorded in A. hypochondriacus, which were fully corroborative to the findings of Khanam and Oba43 of green and red amaranth. Betalain, betacyanin, and betaxanthin content in the current investigation were much pronounced than betalain, betacyanin, and betaxanthin content of red morph amaranth35, green morph amaranth37, stem amaranth38, and weedy amaranth36, while the content of these color pigments of this study was corroborated with A. blitum39 of our earlier studies. Total carotenoids content in the current investigation was much greater than the total carotenoids content of green morph amaranth37, A. spinosus weedy amaranth36 and total carotenoids content of this study were corroborated with A. viridis weedy amaranth36, while total carotenoids content in the current investigation was lower than the total carotenoids content of red morph amaranth35, stem amaranth38 and A. blitum39 of our earlier studies.
Bioactive components and radical scavenging capacity
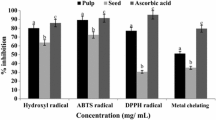
The total polyphenols (TP), vitamin C, β-carotene, total antioxidant activity (TA), and total flavonoids (TF) contents of A. hypochondriacus are presented in Table 4. The highest content of β-carotene was recorded in the genotype AHC2 (82.34 mg 100 g−1), while the genotype AHC4 exhibited the lowest β-carotene content (48.33 mg 100 g−1 FW) with an average of 58.26 mg 100 g−1 FW. Five accessions had greater β-carotene contents than the mean value. The content of β-carotene was high in the genotypes AHC3, AHC5, and AHC8. The vitamin C exhibited pronounced variations regarding genotypes, which ranged from 11.97 mg 100 g−1 in the genotype AHC3 to 184.77 mg 100 g−1 in the genotype AHC11 with an average of 84.44 mg 100 g−1 FW. Six accessions had higher vitamin C contents than the grand mean value. High vitamin C content was recorded in the genotypes AHC1, AHC2, AHC9, and AHC4. The total polyphenols (TP) ranged from 13.23 GAE μg g−1 FW (AHC8) to 29.34 GAE μg g−1 FW (AHC11) with an average TP content of 24.04 GAE μg g−1 FW. High TP was noted in the genotypes AHC4, AHC2, AHC1, and AHC6. Six accessions exerted greater TP contents than the mean TP content. The pronounced variability was recorded in TF content regarding genotypes, which ranged from 106.80 RE μg g−1 DW ( AHC8) to 170.97 RE μg g−1 DW) ( AHC9). The average value of TF content was 147.26 RE μg g−1 DW. The highest TF content was recorded in the genotype AHC9 with the order of AHC9 ˃ AHC4 = AHC5 ˃ AHC10 = AHC1 ˃ AHC2 = AHC6. Six accessions had higher TF content than the mean TF content. The TA (DPPH) content was high and variability was not pronounced, which ranged from 16.27 Trolox equivalent antioxidant capacity (TEAC) μg g−1 DW (AHC8) to 33.46 TEAC μg g−1 DW (AHC6). The highest TA (DPPH) content was recorded in the genotypes AHC6, AHC4, AHC11, AHC5, and AHC10 (DPPH). Conversely, the lowest TA (DPPH) content was found in the genotype AHC8 with a mean TA (DPPH) value of 30.52 TEAC μg g−1 DW. Eight accessions exerted much greater TA (DPPH) contents than the mean TA (DPPH) content. Similar to TA (DPPH), TA (ABTS+) showed a similar trend, which validated the measurement of the quenching capacity of radicals in two methods. The TA (ABTS+) was high and variations were not pronounced, which ranged from 28.54 TEAC μg g−1 DW (AHC8) to 62.92 TEAC μg g−1 DW (AHC6). The highest TA (ABTS+) was noted in the genotypes AHC6, AHC4 AHC11, AHC5, and AHC10. Conversely, the lowest TA (ABTS+) was recorded in the genotype AHC8 with an average of 56.88 TEAC mg g−1 DW. Nine genotypes exhibited much higher TA (ABTS+) contents than the mean TA (ABTS+) value. We observed considerable vitamin C and β-carotene (184.77 and 82.34 mg 100 g−1) in the accessions of A. hypochondriacus, which were greater than the findings of red amaranth of our previous studies10. The TP (29.34 GAE μg g−1 FW) recorded in this investigation was much greater than the findings of green and red amaranth of Khanam et al.44. Our obtained TF (170.97 RE μg g−1 DW), TA (DPPH) (33.46 TEAC μg g−1 DW), and TA (ABTS+) (62.92 TEAC μg g−1 DW) in A. hypochondriacus were corroborated with the findings of red amaranth of Khanam et al.44, while the results of green amaranth of Khanam et al.44 had much lower values compared to our results. The beta-carotene obtained from A. hypochondriacus leafy vegetables was greater than our previous study of A. spinosus weedy amaranth36 and corroborated with beta-carotene of A. viridis weedy amaranth36, while it was lower than our previous study of red morph amaranth35, stem amaranth38, and A. blitum39. The ascorbic acid obtained from A. hypochondriacus leafy vegetables was greater than our previous studies of green morph amaranth37, A. spinosus weedy amaranth36, stem amaranth38 and lower than red morph amaranth35, A. viridis weedy amaranth36, and A. blitum39. TP content of the present study was greater than red morph amaranth35 and A. spinosus weedy amaranth36, while it showed lower results than A. viridis weedy amaranth36. Total flavonoids recorded in A. hypochondriacus leaves were greater than our previous studies of red morph amaranth35, green morph amaranth37 stem amaranth38 and A. blitum39, while TF content of A. hypochondriacus leaves was lower than of our earlier studies of weedy amaranth36. The antioxidant capacity in DPPH and antioxidant capacity in ABTS+ obtained from A. hypochondriacus leaves were greater than our previous study of red morph amaranth35, green morph amaranth37, weedy amaranth36, stem amaranth38, and A. blitum39. The genotypes AHC6, AHC4, AHC11, AHC5, and AHC10 had high phenolics, vitamins, pigments, and antioxidants including high TA. We can utilize these five genotypes as high-yielding varieties containing high antioxidant profiles. We could conclude that A. hypochondriacus leaves have abundant phenolics, pigments, flavonoids, vitamins, and antioxidants that offered enormous prospects for nourishing the vitamin and antioxidant scarce people and detail pharmacological study.
Analysis of correlation coefficient
The correlation of TA (DPPH), β-carotene, phytopigments, vitamin C, TF, TA (ABTS+), and TP of A. hypochondriacus are presented in Table 5. The above-mentioned correlation coefficients exhibited interesting results. All phytopigments exerted positive and significant interrelationships with TA (DPPH), TF, TA (ABTS+), and TP. It indicated that the increase in chlorophylls, carotenoids, β-xanthin, betalain, and β-cyanin content was directly associated with the increment of TA (ABTS+), TP, TA (DPPH), and TF. Similarly, vitamin C had positive and insignificant correlations with TA, TF, and TP, whereas it exerted negative and insignificant correlations with all phytopigments. In the earlier works of our research group in A. tricolor9,11,12,16; in red morph amaranth35, in green morph amaranth37 and Jiménez-Aguilar and Grusak40 in 15 amaranth species observed the similar results which were corroborative to our present study. TA (ABTS+), TF, TA (DPPH), TPC, and β-carotene showed a positive and significant correlation among each other. It indicated that TF, TP, and β-carotene had strong quenching capacity of radicals. Similarly, positive and significant associations versus TA (DPPH) and TA (ABTS+) ensured the confirmation of the quenching capacity of radicals of A. hypochondriacus by two methods. All TP, phytopigments, TF, and β-carotene had strong quenching capacity of radicals as these antioxidant phytochemicals showed significant correlations with TA (DPPH) and TA (ABTS+). In this study, we observed that TP, phytopigments, TF, and β-carotene of A. hypochondriacus had strong quenching capacity of radicals.
In conclusions, It is the first report on A. hypochondriacus leaves. We grew eleven A. hypochondriacus genotypes to evaluate nutraceuticals, bioactive components, and quenching capacity of radicals. A. hypochondriacus leaves contained ample iron, K, zinc, Ca, copper, Mg, manganese, TF, protein, TP, dietary fiber, phytopigments, β-carotene, TA, vitamin C, carbohydrates, and antioxidants. Thus we can make drinks from A. hypochondriacus containing plentiful nutraceuticals, flavonoids, β-carotene, phytopigments, phenolics, vitamin C, and antioxidants to combat hidden hunger and achieve nutritional and antioxidant sufficiency. Based on correlation coefficients, all bioactive compounds of A. hypochondriacus leaves exhibited strong antioxidant activity. Hence, A. hypochondriacus leaves with staple foods (e.g., rice, wheat, maize) could significantly contribute to reducing the occurrence of hidden hunger in the globe. The genotypes AHC6, AHC4, AHC11, AHC5, and AHC10 can be utilized for making drinks.
Methods
Experimental materials
Eleven A. hypochondriacus genotypes were evaluated in this experiment.
Design and layout
We executed the experiment in three replicates following a completely randomized block design (RCBD) at Bangabandhu Sheikh Mujibur Rahman Agricultural University. Each genotype was grown in 1 m2 experimental plot following 20 cm and 5 cm distance between rows and plants, respectively.
Intercultural practices
Recommended fertilizer doses, such as triple super phosphate, murate of potash, gypsum, and urea, at the rate of 100, 150, 30, and 200 kg/ha, respectively were applied45. Cultural practices were maintained appropriately45. Compost (10 ton/ha) was applied during preparation of lands10. For maintaining the exact spacing of plants in a row, proper thinning was executed. Weeds of experimental plots were regularly removed through proper weeding and hoeing. We provide regular irrigation in the experimental plots for maintaining the proper growth of vegetable amaranth. We collected the leaf samples at 30 days old plant.
Solvent and reagents
Solvent: Acetone, hexane, and methanol. Reagents: dithiothreitol (DTT), cesium chloride, HClO4, HNO3, H2SO4, ascorbic acid, standard compounds of pure Trolox (6-hydroxy-2, 5, 7, 8-tetramethyl-chroman-2-carboxylic acid), Folin-Ciocalteu reagent, gallic acid, DPPH, rutin, ABTS+, 2, 2-dipyridyl, aluminum chloride hexahydrate, potassium acetate, sodium carbonate, and potassium persulfate.
Estimation of proximate composition
AOAC method22 was followed to estimate the ash, moisture, crude fat, fiber, crude protein contents, and gross energy. The nitrogen was calculated following the Micro-Kjeldahl method. Finally, nitrogen was multiplied by 6.25 to measure crude protein (AOAC method 976.05). The total moisture, crude protein, ash, and crude fat (%) was subtracted from 100 for calculating carbohydrate (g kg−1 FW).
Estimation of mineral composition
A. hypochondriacus leaf samples were dried in an oven at 70 °C for 24 h. Dried samples were ground in a mill. We determined calcium, potassium, magnesium, iron, manganese, copper, zinc, from powdered leaves following nitric-perchloric acid digestion method22. For this digestion, in the presence of carborundum beads 40 ml HClO4 (70%), 400 ml HNO3 (65%), and 10 ml H2SO4 (96%) were added to 0.5 g dried leaf sample. After digestion, the ascorbic acid method was followed to measure P through dilution of the solution appropriately in triplicate. We added ascorbic acid and antimony to the yellow-colored complex solution for converting it to a blue-colored phosphomolybdenum complex. Sarker and Oba22 method was followed to read the absorbance by atomic absorption spectrophotometry (AAS) (Hitachi, Tokyo, Japan) at wavelengths of 285.2 nm (magnesium), 76 6.5 nm (potassium), 248.3 nm (iron), 422.7 nm (calcium), 279.5 nm (manganese), 213.9 nm (zinc), 324.8 nm (copper).
Determination of chlorophylls and carotenoids
Chlorophyll ab, chlorophyll b, carotenoids, and chlorophyll a were calculated by extracting the leaves in acetone (80%)22,46. A spectrophotometer (Hitachi, U-1800, Tokyo, Japan) was used to measure the absorbance at 646 nm for chlorophyll b and 663 nm for chlorophyll a, and 470 nm for carotenoids, respectively.
The formulae were given below:
Chlorophyll a (μg/mL) = Ca = 12.21 A663 − 2.81 A646.
Chlorophyll b (μg/mL) = Cb = 20.13 A646 − 5.03 A663.
Carotenoids (μg/mL) = (1000A470 – 3.27 Ca – 104 Cb/229.
Where A646 = absorbance at a wavelength of 646 nm; A663 = absorbance at a wavelength of 663 nm; A470 = absorbance at a wavelength of 470 nm.
Finally, chlorophylls were calculated as micrograms per gram and carotenoids milligrams per 100 g of fresh weight, respectively.
Betacyanins and betaxanthins content measurement
The leaves were extracted in 80% methyl alcohol containing 50 mM ascorbate to measure betacyanins and betaxanthins according to the method of Sarker and Oba22. A spectrophotometer (Hitachi, U-1800, Tokyo, Japan) was used to measure the absorbance at 540 nm for betacyanins and 475 nm for betaxanthins, respectively. The data were calculated as nanograms betanin equivalent per gram of fresh weight for betacyanins and nanograms indicaxanthin equivalent per gram of fresh weight for betaxanthins.
Estimation of beta-carotene
For the estimation of beta-carotene, we followed our previously described method22. Exactly 500 mg of fresh leaf sample was added with 10 ml of 80% acetone and ground thoroughly in a mortar and pestle. The extract was centrifuged at 10,000 × g for 3–4 min. The final volume was marked up to 20 ml after removing the supernatant in a volumetric flask. A Hitachi (U-1800, Tokyo, Japan) spectrophotometer at 510 nm and 480 nm, respectively was set to take the absorbance. Data were expressed as milligrams beta-carotene per 100 g of fresh weight.
Beta-carotene = 7.6 (Abs. at 480) − 1.49 (Abs. at 510) × Final volume/ (1,000 × fresh weight of leaf).
Estimation of ascorbic acid
A Hitachi spectrophotometer (U-1800, Tokyo, Japan) was utilized to estimate ascorbic acid (AsA) and dehydroascorbic acid (DHA) from the fresh leaves. Dithiothreitol (DTT) was used for the sample pre-incubation and reduction of dehydroascorbic acid into ascorbic acid. Ascorbic acid reduced ferric ion to ferrous ion. Reduced ferrous ion forms complexes with 2, 2-dipyridyl22. We read the absorbance of Fe2+ complexes with 2, 2-dipyridyl at 525 nm for estimation of vitamin C through the spectrophotometric (Hitachi, U-1800, Tokyo, Japan). We calculated vitamin C in milligrams per 100 g of fresh weight.
Estimation of total polyphenols
Extraction of total polyphenols was carried out according to Jiménez-Aguilar and Grusak40 using 25 mg of sample in 2.5 mL of 1.2 M HCl containing methanol (90%) at 90 °C for 2 h in a water bath. With readjusting the volume (2.5 mL), the leaf extract was centrifuged at 7,500 rpm for 20 min. The leaf extracts (100 µL) were added to the Folin–Ciocalteau reagent (2 N, 50 µL). After 5 min, 2 N Na2CO3 (400 µL) and water (1 mL) was added. The leaf extracts were incubated for 90 min at 37 °C. Finally, it was removed to a microplate (flat bottom). In a microplate reader, the absorbance was detected at 740 nm. We estimated the results in equivalent to gallic acid (GAE) standard µg g−1 of FW. Note that results could be slightly overestimated because these samples contained ascorbic acid47.
Estimation of total flavonoids
Total flavonoids were extracted and quantified according to the method described by Jiménez-Aguilar and Grusak40. Samples (100 mg) were mixed with 5 mL methanol (50%) in water and placed for 1 h with ultrasound. The leaf extracts were centrifuged for 10 min at 13,000 g (4 °C). The supernatants were then recovered. Flavonoid extracts (400 µL) were homogenized with water (500 µL), 5% NaNO2 (60 µL), 10% AlCl3 (140 µL). After 10 min, 1 mM NaOH (400 µL) was added. The leaf extracts were incubated for 10 min at a normal temperature. Finally, it was removed to a flat bottom microplate. The absorbance was read at 500 nm in a microplate reader. Results are expressed in µg of rutin equivalents (RE) per gram of sample DW.
Radical quenching capacity assay
Thirty days old leaves were harvested. For the antioxidant capacity assay, the leaves were dried in the air in a shade. 40 ml aqueous methanol (90%) was utilized to extract grounded dried leaves (1 g) from each cultivar in a capped bottle (100 ml). A Thomastant T-N22S (Thomas Kagaku Co. Ltd., Japan) shaking water bath was utilized to extract leaf samples for 1 h. Exactly 0.45 µm filter (MILLEX-HV syringe filter, Millipore Corporation, Bedford, MA, USA) was used to filter the homogenized mixture. After centrifugation for 15 min at 10,000 × g, the antioxidant capacity was estimated from the filtered extract.
Diphenyl-picrylhydrazyl (DPPH) radical degradation method was used to estimate the antioxidant activity. We added 1 ml DPPH solution (250 µM) to 10 µl extract (in triplicate) in a test tube. After adding 4 ml distilled water the extract was placed in the dark for 30 min. A Hitachi U1800 spectrophotometer (Hitachi, Tokyo, Japan) was used to measure the absorbance at 517 nm. Method of Khanam et al.44 was followed for ABTS+ assay. To prepare two stock solutions separately ABTS+ solution of 7.4 mM and potassium persulfate of 2.6 mM were used. We mixed both solutions in equal proportion to prepare the working solution at room temperature. The working solution was allowed to react in the dark for 12 h. One hundred fifty μl extract was added to 2.85 ml of ABTS+ solution and allowed to react in the dark for 2 h. For the preparation of the solution, one ml of ABTS+ solution was mixed with sixty ml of methanol. A Hitachi spectrophotometer (U1800, Tokyo, Japan) was utilized to take the absorbance against methanol at 734 nm. The inhibition (%) of DPPH and ABTS+ corresponding with control was used to determine antioxidant capacity using the equation as follows:
Antioxidant activity (%) = (Abs. blank − Abs. sample/Abs. blank) × 100.
Where, Abs. blank is the absorbance of the control reaction [10 µl methanol for TAC (DPPH), 150 μl methanol for TAC (ABTS+) instead of leaf extract] and Abs. sample is the absorbance of the test compound. Trolox was used as the reference standard, and the results were expressed as μg Trolox equivalent g−1 DW.
Statistical analysis
The replication mean was obtained by averaging the replication-wise sample data. We performed the analysis of variance (ANOVA) using Statistix 8 software. The mean separation was performed by Tukey’s HSD test at a probability level of 1%. We reported the results as the mean ± SD.
Ethical statement
The lab and field experiment in this study was carried out following guidelines and recommendations of “Biosafety Guidelines of Bangladesh” published by the Ministry of Environment and Forest, Government of the People’s Republic of Bangladesh (2005).
Data availability
The data used in this manuscript will be available to the public.
References
FAO, IFAD, & WFP. The state of food security in the world 2015. Meeting the 2015 International Hunger Targets: Taking Stock of Uneven Progress (2015). Retrieved January 3, 2019. https://www.fao.org/3/a-i4646e.pdf.
Von Grebmer, K., Saltzman, A., Birol, E., Wiesmann, D., Prasai, N., Yin, S., Yohannes, Y., Menon, P., Thompson, J., & Sonntag, A. 2014 Global Hunger Index: The Challenge of Hidden Hunger. Welthungerhilfe, International Food Policy Research Institute, and Concern Worldwide, Bonn, Washington, D.C., and Dublin (2014).
Afari-Sefa, V., Tenkouano, A., Ojiewo, C. O., Keatinge, J. D. H. & Hughes, J. D. A. Vegetable breeding in Africa: constraints, complexity, and contributions toward achieving food and nutritional security. Food Secur. 4, 115–127 (2011).
Grosso, G. et al. Effects of vitamin C on health: a review of evidence. Front. Biosci. 18, 1017–1029 (2013).
Isabelle, M. et al. Antioxidant activity and profiles of common fruits in Singapore. Food Chem. 123, 77–84 (2010).
Randhawa, M. A., Khan, A. A., Javed, M. S. & Sajid, M. W. Green leafy vegetables: a health-promoting source. In Handbook of Fertility (ed. Watson, R. R.) 205–220 (Academic Press, San Diego, 2015).
Jensen, A. Chlorophylls & carotenoids. In Handbook of Physiological Methods and Biochemical Methods (eds Hellebust, J. A. & Craigie, J. S.) 5–70 (Cambridge University Press, Cambridge, 1978).
Guillet D. Grain amaranthus, history, and nutrition. Kokopelli Seed Foundation [Online] (2004). https://www.kokopelli-seed-foundation.com/amaranths.htm. Accessed March 10th 2020.
Sarker, U., Islam, M. T., Rabbani, M. G. & Oba, S. Genotypic variability for nutrient, antioxidant, yield and yield contributing traits in vegetable amaranth. J. Food Agri. Environ. 12, 168–174 (2014).
Sarker, U., Islam, M. T., Rabbani, M. G. & Oba, S. Variability, heritability and genetic association in vegetable amaranth (A. tricolor). Span. J. Agric. Res. 13, 1–8. https://doi.org/10.5424/sjar/2015132-6843 (2015).
Sarker, U., Islam, M. T., Rabbani, M. G. & Oba, S. Genotype variability in composition of antioxidant vitamins and minerals in vegetable amaranth. Genetika 47, 85–96 (2015).
Sarker, U., Islam, M. T., Rabbani, M. G. & Oba, S. Genetic variation and interrelationship among antioxidant, quality and agronomic traits in vegetable amaranth. Turk. J. Agric. For. 40, 526–535 (2016).
Sarker, U., Islam, M. T., Rabbani, M. G. & Oba, S. Genotypic diversity in vegetable amaranth for antioxidant, nutrient and agronomic traits. Indian J. Genet. Pl. Br. 77, 173–176 (2017).
Shukla, S. et al. Mineral profile and variability in vegetable amaranth (Amaranthus tricolor). Pl. Food. Hum. Nutri. 61, 23–28 (2006).
Chakrabarty, T., Sarker, U., Hasan, M. & Rahman, M. M. Variability in mineral compositions, yield, and yield contributing traits of stem amaranth (Amaranthus lividus). Genetika 50, 995–1010 (2018).
Sarker, U., Islam, M. T., Rabbani, M. G. & Oba, S. Variability in total antioxidant capacity, antioxidant leaf pigments and foliage yield of vegetable amaranth. J. Integr. Agric. 17, 1145–1153 (2018).
Sarker, U., Islam, M. T., Rabbani, M. G. & Oba, S. Antioxidant leaf pigments and variability in vegetable amaranth. Genetika 50, 209–220 (2018).
Sarker, U., Islam, M. T., Rabbani, M. G. & Oba, S. Phenotypic divergence in vegetable amaranth for total antioxidant capacity, antioxidant profile, dietary fiber, nutritional and agronomic traits. Acta Agric. Scand. Sect. B Soil Plant Sci. 68, 67–76 (2018).
Repo-Carrasco-Valencia, R., Hellstrom, J. K., Pihlava, J. M. & Mattila, P. H. Flavonoids and other phenolic compounds in Andean indigenous grains: quinoa (Chenopodium quinoa), kaniwa (Chenopodium pallidicaule) and kiwicha (Amaranthus caudatus). Food Chem. 120, 128–133 (2010).
Venskutonis, P. R. & Kraujalis, P. Nutritional components of amaranth seeds and vegetables: a review on composition, properties, and uses. Compr. Rev. Food Sci. Food Saf. 12, 381–412 (2013).
Cai, Y., Sun, M. & Corke, H. Antioxidant activity of betalains from plants of the Amaranthaceae. J. Agril. Food Chem. 51, 2288–2294 (2003).
Sarker, U. & Oba, S. Response of nutrients, minerals, antioxidant leaf pigments, vitamins, polyphenol, flavonoid and antioxidant activity in selected vegetable amaranth under four soil water content. Food Chem. 252, 72–83 (2018).
Sarker, U. & Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 18, 258. https://doi.org/10.1186/s12870-018-1484-1 (2018).
Sarker, U. & Oba, S. Drought stress effects on growth, ros markers, compatible solutes, phenolics, flavonoids and antioxidant activity in A. tricolor. Appl. Biochem. Biotech. https://doi.org/10.1007/s12010-018-2784-5 (2018).
Sarker, U. & Oba, S. Catalase, superoxide dismutase, and ascorbate-glutathione cycle enzymes confer drought tolerance of A. tricolor. Sci. Rep. 8, 16496. https://doi.org/10.1038/s41598-018-34944-0 (2018).
Sarker, U. & Oba, S. Salinity stress enhances color parameters, bioactive leaf pigments, vitamins, polyphenols, flavonoids and antioxidant activity in selected Amaranthus leafy vegetables. J. Sci. Food Agric. 99, 2275–2284. https://doi.org/10.1002/jsfa.9423 (2019).
Sarker, U., Islam, M. T. & Oba, S. Salinity stress accelerates nutrients, dietary fiber, minerals, phytochemicals and antioxidant activity in Amaranthus tricolor leaves. PLoS ONE https://doi.org/10.1371/journal.pone.0206388 (2018).
Sarker, U. & Oba, S. Augmentation of leaf color parameters, pigments, vitamins, phenolic acids, flavonoids and antioxidant activity in selected A. tricolor under salinity stress. Sci. Rep. 8, 12349. https://doi.org/10.1038/s41598-018-30897-6 (2018).
Jansen, P. C. M. Amaranthus hypochondriacus L. [Internet] Record from PROTA4U. In Grubben, G. J. H. & Denton, O. A. (Eds.), PROTA (Plant Resources of Tropical Africa/Ressources vegetales de l’Afrique tropicale), Wageningen, Netherlands (2004). https://www.prota4u.org/search.asp. Accessed March 9th 2020.
Rajan, S. & Markose, B. L. Horticultural science series-6. In Propagation of Horticultural Crops (ed. Peter, K. M. V.) (New India Publishing Agency, New Delhi, India, 2007).
Dusgupta, N. & De, B. Antioxidant activity of some leafy vegetables of India: a comparative study. Food Chem. 101, 471–474 (2007).
Steffensen, S. K. et al. Variations in the polyphenol content of seeds of field grown Amaranthus genotypes. Food Chem. 129, 131–138 (2011).
Rice-Evans, C. A., Miller, N. J. & Papanga, G. Antioxidant properties of phenolic compounds. Trends Plant. Sci. 2, 152–159 (1997).
Sun, H., Mu, T., Xi, L., Zhang, M. & Chen, J. Sweet potato (Ipomoea batatas L.) leaves as nutritional and functional foods. Food Chem. 156, 380–389 (2014).
Sarker, U. & Oba, S. Protein, dietary fiber, minerals, antioxidant pigments and phytochemicals, and antioxidant activity in selected red morph Amaranthus leafy vegetable. PLoS ONE https://doi.org/10.1371/journal.pone.0222517 (2019).
Sarker, U. & Oba, S. Nutraceuticals, antioxidant pigments, and phytochemicals in the leaves of A. spinosus and A. viridis weedy species. Sci. Rep. 9, 20413. https://doi.org/10.1038/s41598-019-50977-5 (2019).
Sarker, U., Hossain, M. M. & Oba, S. Nutritional and antioxidant components and antioxidant capacity in green morph Amaranthus leafy vegetable. Sci. Rep. 10, 1336. https://doi.org/10.1038/s41598-020-57687-3 (2020).
Sarker, U., Oba, S. & Daramy, M. A. Nutrients, minerals, antioxidant pigments and phytochemicals, and antioxidant capacity of the leaves of stem amaranth. Sci. Rep. 10, 3892. https://doi.org/10.1038/s41598-020-60252-7 (2020).
Sarker, U. & Oba, S. Nutrients, minerals, pigments, phytochemical, and radical scavenging activity in Amaranthus blitum leafy vegetable. Sci. Rep. 10, 3868. https://doi.org/10.1038/s41598-020-59848-w (2020).
Jimenez-Aguilar, D. M. & Grusak, M. A. Minerals, vitamin C, phenolics, flavonoids and antioxidant activity of Amaranthus leafy vegetables. J. Food Compos. Anal. 58, 33–39 (2017).
Madruga, M. S. & Camara, F. S. The chemical composition of “Multimistura” as a food supplement. Food Chem. 68, 41–44 (2000).
Shahidi, F., Chavan, U. D., Bal, A. K. & McKenzie, D. B. Chemical composition of beach pea (Lathyrus maritimus L.) plant parts. Food Chem. 64, 39–44 (1999).
Khanam, U. K. S. & Oba, S. Bioactive substances in leaves of two amaranth species, Amaranthus lividus and A. hypochondriacus. Can. J. Plant Sci. 93, 47–58 (2013).
Khanam, U. K. S., Oba, S., Yanase, E. & Murakami, Y. Phenolic acids, flavonoids and total antioxidant capacity of selected leafy vegetables. J. Funct. Foods. 4, 979–987 (2012).
Rashad, M. M. I. & Sarker, U. Genetic variations in yield and yield contributing traits of green amaranth. Genetika 52, 393–407 (2020).
Sarker, U. & Oba, S. Antioxidant constituents of three selected red and green color Amaranthus leafy vegetable. Sci. Rep. 9, 18233. https://doi.org/10.1038/s41598-019-52033-8 (2019).
Prior, R., Wu, X. & Schaich, K. Standardized methods for the determination of Antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 53, 4290–4302 (2005).
Author information
Authors and Affiliations
Contributions
U.S. initiated the research work and conceived the study; U.S. performed the experiments; biochemical analysis and statistical analysis; U.S. drafted, edited, interpreted data and prepared the manuscript; S.O. edited the manuscript, provided valuable suggestions during the experiment.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sarker, U., Oba, S. Nutritional and bioactive constituents and scavenging capacity of radicals in Amaranthus hypochondriacus. Sci Rep 10, 19962 (2020). https://doi.org/10.1038/s41598-020-71714-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-71714-3
- Springer Nature Limited
This article is cited by
-
Bioactive polyphenolic compounds and antioxidant potentials of two leafy vegetables in Bangladesh: the Momordica charantia and the Ipomoea aquatica
Food Production, Processing and Nutrition (2024)
-
Nutraceuticals as components of sustainable poultry production systems for food and nutrition security in Africa: a review
Agriculture & Food Security (2024)
-
Changes in polyphenolic composition, physiological characteristics, and yield-related traits of Moshgak (Ducrosia anethifolia Boiss.) populations in response to drought stress
Protoplasma (2023)
-
Color attributes, betacyanin, and carotenoid profiles, bioactive components, and radical quenching capacity in selected Amaranthus gangeticus leafy vegetables
Scientific Reports (2021)
-
Nutraceuticals, phytochemicals, and radical quenching ability of selected drought-tolerant advance lines of vegetable amaranth
BMC Plant Biology (2020)