
Abstract
Behavioral data can be important for effective management of endangered marine predators, but can be challenging to obtain. We utilized suction cup-attached biologging tags equipped with stereo hydrophones, triaxial accelerometers, triaxial magnetometers, pressure and temperature sensors, to characterize the subsurface behavior of an endangered population of killer whales (Orcinus orca). Tags recorded depth, acoustic and movement behavior on fish-eating killer whales in the Salish Sea between 2010–2014. We tested the hypotheses that (a) distinct behavioral states can be characterized by integrating movement and acoustic variables, (b) subsurface foraging occurs in bouts, with distinct periods of searching and capture temporally separated from travel, and (c) the probabilities of transitioning between behavioral states differ by sex. Using Hidden Markov modeling of two acoustic and four movement variables, we identified five temporally distinct behavioral states. Persistence in the same state on a subsequent dive had the greatest likelihood, with the exception of deep prey pursuit, indicating that behavior was clustered in time. Additionally, females spent more time at the surface than males, and engaged in less foraging behavior. These results reveal significant complexity and sex differences in subsurface foraging behavior, and underscore the importance of incorporating behavior into the design of conservation strategies.
Similar content being viewed by others


Introduction
Studying the behavior of organisms can yield insights that transcend disciplines, and is an urgent priority as global change imposes novel pressures on ecological patterns and processes1,2,3,4,5. For threatened and endangered populations, characterizing behavior can be integral to conservation outcomes3, yet particularly challenging for these populations. The rarity of such populations can make them paradoxically difficult to study6. Furthermore, behavioral studies are vulnerable to observer bias and the tendency to quantify behaviors through subjective lenses7. Additionally, for species that spend the majority of their time in habitats inaccessible to researchers, quantifying behavior can be especially challenging and confounded by time (i.e., only the behaviors that are observed are quantified)5.
Many populations of cetaceans (whales, dolphins and porpoises) are endangered globally8. Until recently it was difficult to obtain continuous observations of subsurface behavior to inform conservation efforts because these species spend the majority of their lives underwater. For endangered cetaceans that forage in groups, individuals need to balance the energetic gains of group-facilitated prey consumption with the metabolic costs of prey pursuit at depth9,10,11,12. The ways in which these species temporally structure their foraging activities to account for this paradox, however, are poorly understood yet fundamental to conservation.
We studied the foraging behavior of an endangered population of killer whales (Orcinus orca), which lives in the Northeastern Pacific Ocean, and spends the majority of its time in the inland and coastal waters along the west coast of North America between central California and southeastern Alaska. Individuals in the Southern Resident killer whale (SRKW) population spend their lifetimes in stable, matrilineal groups, and consume salmon, primarily Chinook (Oncorhynchus tshawytscha)13,14, that are routinely shared with group members15. Chinook salmon, the largest salmon species, provide a significant energetic return, but given that they occur deeper than other salmon species, up to several hundred meters16,17, SRKW must invest significant energy in the forms of breath holding and locomotion costs associated with fluking and drag11,12, in order to pursue these larger caloric payoffs. The challenge of acquiring sufficient Chinook salmon to meet caloric requirements is compounded by the fact that many Chinook salmon populations originating from the west coast of the United States and British Columbia are threatened or endangered18.
To study foraging behavior, past studies primarily relied upon observations of kills at the surface, collection of samples of prey floating in the water after kills, or collection of fecal samples indicating recent consumption e.g.13,14. Surface observations revealed distinct episodes of foraging activities, including increased swim speeds indicative of prey pursuit14, and aggregation behavior following prey capture, presumably to share prey among group members, particularly between dependent offspring and mothers. A recent study utilized movement variables of subsurface behavior to identify prey capture events, in order to explore sex differences in foraging ecology of fish-eating killer whales, revealing males made more prey capture dives than females, presumably to support the energetic requirements of a larger body size19. Currently, however, little is known about the extent to which these prey pursuits represent the culmination of continuous search efforts, or whether foraging occurs in distinct periods in space and time, allowing for other activities including recovery following metabolically costly deep dives, socialization, traveling, or other non-foraging activities. Additionally, given the sexual dimorphism between males and females and differences in metabolic rates, energetic requirements12 and foraging rates19, it is possible that males and females may partition behavioral activities or timing to minimize energy use while maximizing gain and meeting other needs including care of dependent offspring. Indeed, in many species of sexually dimorphic marine mammals, foraging behavior is sexually segregated20,21,22. Addressing these data gaps can inform management of the SRKW population in critcal habitat including the inland waters of Washington State and British Columbia, where many commercial and recreational interests coincide with the whales’ foraging activities from May through September.
We utilized multisensor biologging tags to characterize the subsurface behavior of SRKW. Specifically, tags were attached by suction cup to killer whales and recorded depth, movement, and sound, providing the opportunity to collect a continuous stream of data as the subjects traveled below the surface. To minimize the effect of subjective observer bias on behavioral categorization, we used hidden Markov models to systematically characterize latent behavioral states based on four movement and two acoustic variables computed from data recorded on the tags. This approach enabled quantification of behavioral states based on similarities identified through a latent Markov process rather than relying on subjective assignment by researchers. We then used these behavioral state allocations to test the hypotheses that (a) SRKW engage in distinct behavioral activities characterized by differences in their subsurface movement and acoustic behavior, (b) SRKW foraging behavior occurs in bouts, with distinct periods of searching and capture followed by rest, and (c) the probabilities of transitioning between behavioral states differ by sex.
Knowledge of the behavior of a species, the temporal patterning of activity budgets, and whether behavioral patterns differ by sex is valuable for guiding effective conservation5,23. The SRKW population was listed as Endangered in the United States24 and as a Species at Risk in Canada25. Though this population has been small (<100 animals) since 1975, recent declines have reduced the population to a near record low of 73 animals at present, prompting increased concern over its future viability26,27. One of the main threats to population persistence is availability and accessibility of prey14,27,28. Therefore, a better understanding of the temporal patterns of behavior, and whether foraging-related activities including shallow searching and deep prey pursuit differ by sex, will enable the design of effective SRKW conservation measures that promote foraging opportunities.
Results
Summary of deployments
We analyzed a total of 41.8 h from all 13 deployments (2010 = 9.3 h, n = 3; 2012 = 15.3 h, n = 6; 2014 = 17.2 h, n = 4) (Table 1). Mean analyzed time per deployment was 3.2 h (range = 0.6–6.9 h). We analyzed 3728 dives from these deployments. Females (f) and males (m) made a total of 1324 and 2404 dives, respectively (2010: 253 (f), 517 (m); 2012: 429 (f), 912 (m); 2014: 642 (f), 975 (m)). The number of dives analyzed per deployment ranged from 37 to 583 (Table 1).
State classifications of diving behavior
The unit of analysis was a dive, defined as any departure from and return to the surface. The best model identified five underlying behavioral states in Southern Resident killer whales (Fig. 1, Supplementary Figs S1 and S2). These states were characterized by: (1) deep dives with common occurrence of buzzing, nearly ubiquitous occurrence of clicking, and large values of jerk peak, roll and heading variance, (2) shallow dives with no buzzing, uncommon clicking, and small values of jerk peak, roll and heading variance, (3) shallow to intermediate dives with no buzzing, some clicking, and small-to-moderate values of jerk peak, roll and heading variance, (4) shallow dives with virtually no buzzing, abundant clicking, small values of jerk peak and heading variance, and small-to-moderate values of roll, and (5) shallow dives, no buzzing, rare clicking, and small values of jerk peak, roll and heading variance (Table 2, Fig. 2). There was a significant effect of state on dive duration (linear mixed model: F4, 3718 = 761.2, p < 0.0001) (Fig. 3).

Hidden Markov modeling revealed five distinct behavioral states across all individuals. Dives (n = 3728) are plotted for all deployments (state 1 = purple, state 2 = red, state 3 = orange, state 4 = green, state 5 = blue).

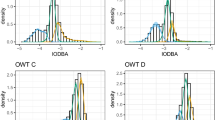
Fitted state-dependent distributions of the best model by sex.

Differences in dive duration by state and sex. Boxes indicate least square mean values for females (black) and males (gray), whiskers indicate confidence. Significance, determined by a Tukey HSD test across all states and sexes, is indicated by unshared letters above bars.
Transition probabilities
For both females and males, the largest transition probabilities observed were for state persistence on the subsequent dive, indicating that behavior was temporally clustered, as expected (Fig. 4, 5, Supplementary Fig. S1). Females and males were most likely to persist in states 2, 4 and 5 (different types of shallow behavior). Persistence in state 1 (deep dives) never occurred for females, and was rare for males. For state 3 (shallow-intermediate dives), persistence was the most likely outcome for males but not females (Fig. 5).

Dive profile of a deployment on a male killer whale, indicating behavioral state allocation over time. State 1 = purple, state 2 = red, state 3 = orange, state 4 = green, state 5 = blue. Gray dives indicate the omitted 5-min interval at the start of each deployment.

Transition probabilities between states for females and males. Line color indicates the state of origin, and line thickness scales positively with probability, which is indicated adjacent to the corresponding line.
For females and males, switching to state 1 was rare, whereas switching to state 2 was common from states 1 and 3. For females, switching to state 3 was moderately likely from states 1 and 2, less common from state 4, and rare from state 5. In contrast, after persistence in state 3, males were equally likely to switch to state 3 from any other state. Males were more likely than females to switch to state 4 from any state except state 1, suggesting males often returned to state 4 throughout a deployment. For both males and females, transitions into state 5 from states 1, 2 or 4 were rare, and infrequent from state 3 (Fig. 5).
State allocations
The multinomial logistic model supported the existence of differences between females and males in the frequency of different behavioral states (Table 3, Supplementary Table S1, Fig. 6A). The positive log odds ratio (with state 1 as the reference baseline) indicated both sexes were more likely to occur in states 2–4 than in state 1. Furthermore, the log odds ratio differed between sexes for all states except state 5, supporting the hypothesis of sex differences in state allocation, and was most different between sexes for state 2, indicating females were substantially more likely to occur in state 2 (respiration) vs. state 1 (deep prey pursuit) than were males.

Summaries of behavioral state allocation for all deployments. (A) Boxplots of proportional occurrence of dives by state, per deployment, for females (light blue) and males (dark blue). Horizontal bars indicate medians, box edges are interquartile ranges, whiskers indicate minimum and maximum values, and outliers are plotted. (B) Proportion of time spent per state, per deployment, arranged by sex. State 1 = purple, state 2 = red, state 3 = orange, state 4 = green, state 5 = blue.
We found a significant difference between males and females in the proportion of time per state (χ2 = 7883.1, df = 4, p < 0.0001; Table 3; Fig. 6B). Females spent substantially less time than males in state 1 or 4, and notably more time than males in states 2, 3, and 5.
Discussion
We used a latent Markov process to identify five temporally distinct behavioral states, demonstrating complexity in subsurface behavior of Southern Resident killer whales. From the most likely state sequence, we computed the probability of state persistence and state switching, revealing that behavior occurs in bouts. We show that females and males differ in their transition probabilities between states, and in the frequency and cumulative time in these states.
State 1 was characterized by deep dives, common buzz and nearly ubiquitous click occurrence, and large values of jerk peak, roll and heading variance. Additionally, state 1 dives had the greatest duration. Consistent with previous results19, state 1 is likely associated with close pursuit and capture attempts of salmonid prey, whereby prey are chased at depths typically greater than 30 m, producing substantial jerk movement due to abrupt acceleration and deceleration, as well as changes in musculature in the head region at prey interception29,30,31,32,33. Additionally, state 1 is associated with large values of roll, and large heading variance due to repeated direction changes34 while closing in on prey. Similarly, prey pursuit dives by Northern Resident killer whales, a partially-sympatric population with a similar diet13, tend to be deeper, longer, include a large roll angle, are more tortuous, and include faster swim speeds than non-pursuit dives35.
Dives allocated to state 2 were characterized by shallow depth, no buzzing and uncommon clicking, and small values of jerk peak, roll and heading variance. These dives were shortest in duration, reflecting their surface association, and typically followed intermediate to deep dives. These dives are likely associated with surface respiration, with small values of heading variance. The high probability of switching from state 1 to state 2, particularly in males, and moderate probability of switching from state 3 to state 2, as well as the high probability of persisting in state 2, supports the idea that state 2 dives likely function as respiration dives as in35 following depletion of oxygen stores during deep dives. The presence of occasional clicking by females and slightly more common clicking by males (Table 2, Fig. 2) suggests that a small amount of acoustic searching may be occurring during state 2 dives as well. State 5 dives were similarly characterized by shallow depth, no buzzes, rare occurrence of clicks, and small values of movement variables including heading variance, which is indicative of directional travel. These dives were short in duration but significantly longer than state 2 dives (Fig. 3), with the highest likelihood of persistence for any of the states. The notable occurrence of these dives in long bouts suggests that state 5 likely represents traveling.
State 4 was characterized by dives similar in depth and kinematics to states 2 and 5, but notably different was the nearly ubiquitous occurrence of clicks. Dives in this state were short as well, similar to those in states 2 and 5. State 4 is likely a prey searching state primarily, consistent with the acoustic behavior that precedes prey capture in many other toothed whales whereby an individual produces echolocation clicks that scan an area for possible targets prior to initiating a capture attempt e.g.29,30,36,37,38,39,40. Additionally, the tendency for these dives to occur in bouts further supports the interpretation that this is primarily a prey searching state. The absence of buzzes from nearly all dives in this state indicates these dives are unlikely to include capture attempts. It is important to note, however, that prey searching does not preclude the occurrence of other surface-associated behaviors that may occur simultaneously, such as traveling or socializing.
Finally, state 3 was characterized by intermediate depth, with no buzzes, relatively common click occurrence, and low to moderate values of jerk peak, roll and heading variance. These dives were intermediate in duration, suggesting there could be a variety of behaviors associated with dives in this state. The absence of buzzes indicates these dives are unlikely prey capture attempts, but it is possible that prey pursuit was initiated and then aborted at intermediate depth. This interpretation is unlikely to explain all dives classified to state 3, however, because approximately two thirds of female dives and half of male dives in this state did not contain clicks. It is possible that state 3 behaviors include socializing, such as engaging in tactile behavior and/or prey sharing, which could explain moderate values of heading variance, roll and jerk peak as individuals move vertically and horizontally and make physical contact with other group members41. Additionally, these behaviors may be best accomplished several meters below the surface, where positional alignment within a pod may be able to take on a three-dimensional formation whereas surface swimming arrangements are confined to essentially the same horizontal plane. Furthermore, acoustic interference with the surface can create zones that impede detection of signals42,43. Therefore, when communicating with individuals out of sight, using stereotyped pulsed calls and whistles with the majority of energy in mid-frequencies (1–20 kHz)44,45, it is possible that swimming approximately 15–25 m below the surface, as observed in many dives classified to state 3, may facilitate social behavior by optimizing passive listening for and active sending of important information.
For both females and males, behavioral bouts (sequences of consecutive dives of the same state) were common for states 2, 4 and 5, moderately common for state 3, and rare for state 1. This indicates that certain behaviors, particularly acoustic searching (state 4) and respiration and travel (states 2 and 5, respectively), are clustered in time. Surface travel behaviors are common in other social odontocetes e.g.35,46,47, supporting our finding of temporal patterning (bouts) of behavior. Indeed, subsurface behavior does not appear to be randomly distributed in time, but rather is temporally clumped, likely driven by other factors, including prey distribution48,49, bathymetry features that may aggregate prey50,51, time of day51,52, oceanographic parameters53, aerobic dive limits54 and anthropogenic pressures55,56,57,58. It is not possible to tease apart the relative importance of each of these factors in this study. However, the extreme sociality of resident-type killer whales likely plays a considerable role in mediating behavioral patterns. Resident-type killer whale movements are coordinated at a coarse scale, with pods often traveling together, or breaking apart briefly to reunite a few hours or a few days later. Furthermore, because of the existence of kin-directed prey sharing among resident-type killer whales15, coordinating behavior to maximize foraging success would benefit the population59. Indeed, studies have revealed synchronicity in diving behavior in other cetaceans41,60,61,62,63.
Notably, persistence in state 1 (final pursuit and capture attempt) between consecutive dives was rare. There are at least two, non-mutually exclusive explanations for this finding. First, deep diving for prey pursuit incurs significant energy expenditure through (1) the depletion of oxygen stores during breath-holding, (2) the kinematics of chasing prey including rapid acceleration and deceleration and sudden direction changes, and (3) withstanding the physiological effects of extreme pressure at depth. Rather than completing costly consecutive dives, individuals may enhance their overall foraging effectiveness by recovering near the surface (demonstrating behavior such as in dives classified to state 2) before initiating another prey pursuit dive. Indeed, many deep-foraging cetaceans make several recovery dives at the surface before returning to depth e.g.35,46,64,65. The second explanation for the paucity of state 1 persistence may be that a prey item is often captured during a deep foraging dive19. Upon capturing a prey item, it is then typically shared among pod-members near the surface15, precluding the occurrence of consecutive deep dives. If prey are not captured during pursuit, it is possible that their heightened vigilance may make consecutive deep prey pursuit dives less likely to be executed due to their potentially reduced likelihood of success.
We found sex differences in state switching behavior. Males were more likely than females to switch to acoustic searching (state 4) from any state except deep prey pursuit (state 1), suggesting males often returned to acoustic searching behavior. This may be driven by key deployments in which females did not engage in any deep prey pursuit behavior. Additionally, females were more likely than males to engage in non-foraging behavior, and spent notably more time in surface respiration, intermediate depth or traveling states than males, who spent substantially more time searching for and pursuing prey. It is possible that female foraging behavior is temporally compartmentalized, due to the time demands of caring for calves or other offspring or kin e.g.66,67, and due to the potentially increased costs of transport for females with calves4. Alternatively, since adult males are larger than females or juvenile males, and have the largest metabolic requirements in the population12, they may need to forage more often than females or juvenile males19 to meet their metabolic requirements, and consequently may engage in searching behavior more often. It is important to note our conclusions are limited by the sample size of deployments greater than three hours on females. We cannot rule out the possibility that collecting a greater number of longer deployments on females could reveal more equal time budgets of state allocation. Nonetheless, our results are consistent with known sex differences in prey capture and prey pursuit in Southern Resident killer whales19.
This investigation utilized Hidden Markov models to characterize the subsurface behavior of a population of killer whales, to shed light on the ways in which group foraging cetaceans temporally structure their foraging activities and to determine whether females and males differ in their behavioral patterns. We revealed that killer whale subsurface activity budgets are complex, involving at least five distinct behavioral states, and that females and males differ in subsurface activity, with females generally engaging in less foraging behavior than males, though we cannot rule out the possible influence of small sample size on this sex difference. We demonstrated that foraging is more than intermittent deep dives to acquire prey, following the general sequence: (1) an event causes a transition from the intermediate depth or traveling states into active searching at the surface using echolocation, (2) the individual identifies a potential target, and initiates a deep dive to investigate further or engage in initial prey pursuit, (3) the individual either abandons the investigation or pursues the target and initiates a chase and potentially a successful capture, and (4) the individual returns to the surface and continues searching or transitions to respiration, traveling or socializing behavior. Consequently, these findings underscore two important points. First, when determining how environmental or human factors may impact foraging, it is important to consider disturbance to multiple foraging-related behaviors, not just to the ultimate prey capture. Second, it is not possible to completely assess whether animals are engaged in foraging activities from surface observations alone. Studies that categorize behavior through focal observations may not be able to resolve differences between dive types, so they may not be able to quantify disruptions to foraging, for example impacts on searching effort, an integral component of foraging. Indeed, we demonstrate that searching appears kinematically similar to respiration or traveling, and is distinguished primarily by the presence of echolocation. Thus, studies quantifying human impact on cetacean foraging should combine surface observations with quantification of subsurface behavior, including both kinematic and acoustic data when possible. This approach, combined with modeling hidden states using latent Markov chains, can provide a valuable method for identifying patterns in behavioral data that can inform management of threatened and endangered species.
Methods
Data collection
We deployed multi-sensor biologging tags (‘Dtags’)68 on SRKW in the Salish Sea during September 2010 and 2014 (Version 2) and September 2012 (Version 3). Dtags were equipped with stereo hydrophones, triaxial accelerometers, triaxial magnetometers, and pressure and temperature sensors, and recorded subsurface acoustic and movement behavior. Dtags sampled audio at 192 kHz and depth, movement and temperature at 50 Hz (Version 2), or sampled audio at 240 kHz and depth, movement and temperature at 200 Hz (downsampled to 50 Hz during post-processing; Version 3). Dtags stored data to onboard flash memory, contained a VHF beacon to enable tag recovery, and were programmed to release before local sunset, if they had not already fallen off naturally. Tagging was conducted under federal research permits in the U.S.A. (NMFS No. 781–1824/16163) and in Canada (DFO SARA/Marine Mammal License No. MML 2010-01/ SARA-106B), and approved by the Northwest Fisheries Science Center’s Institutional Animal Care and Use Committee.
Twenty-three Dtags were attached by suction cup to the dorsal side of 21 killer whales using a 7 m carbon fiber pole held by a tagger standing on the bow pulpit of a 6.7 m rigid-hulled inflatable research vessel. Animal reactions to tagging consisted primarily of mild to moderate behavioral changes, and included flinching or diving for up to a few minutes, but resumed previous behavior within five minutes of tagging. Animals were tagged opportunistically, while ensuring a balanced representation between sexes. All animals were >1 year old. No animals were tagged multiple times within the same year, but two animals were tagged twice in different years. Focal follows of the tagged animal were conducted for the duration that the tag remained on the animal, described elsewhere19,69. These focal follows allowed for identification of changes in tag orientation for use in data calibration, and to facilitate tag retrieval. All tags were attached during only daylight hours.
Data processing
Following tag retrieval, data were downloaded and custom software (the 2014 Dtag Toolbox, www.soundtags.org/dtags/dtag-toolbox) along with custom scripts in Matlab version 2016a (The Mathworks, Natick, MA, USA) were used to calibrate acoustic and movement sensor data and convert pressure data to temperature-corrected depth, following established methods68, see details in19,69. All acoustic audits of sound files were conducted by the same experienced researcher. We excluded nine deployments due to acoustic quality. For these deployments, excessive noise from water flowing past the hydrophones, often due to suboptimal tag placement, prevented acoustic variables from being computed. Additionally, we excluded the first deployment due to inconsistencies in data collection methods and hydrophone gain settings with the rest of the deployments. We down-sampled pressure data to 5 Hz, and applied a custom dive detector that identified a dive as the interval containing a departure from the surface (0.5 m), a maximum depth ≥1 m, and a return to the surface. Due to the relatively high sample rate in the pressure data, occasionally noise in the pressure data caused false detections. Therefore, we checked all dive results manually, and corrected dives as needed. We excluded dives that began within the first five minutes of tag attachment in order to account for any short-term behavioral responses to tagging. We confirmed that this was a conservative estimate of the duration of behavioral responses to tagging, by visually inspecting all dive profiles.
Dive was the unit of analysis in this study, defined as the time between successive respirations at the surface. For each dive, we calculated start and end times of the whole dive and of the bottom phase (≥70% of maximum depth), following methods in Arranz et al.33. Next, we computed six variables for each dive: Maximum depth, the deepest point of a dive (m); Jerk peak, the maximum peak in the jerk (rate of change of acceleration); Roll, the median of the absolute value of the animal’s roll (dorsal-ventral); Heading variance, the circular variance in the animal’s heading (left-right); Buzz presence, the binary presence of echolocation pulses with an inter-click interval ≤10 ms within a click bout; Slow click presence, the binary presence of echolocation clicks with an inter-click interval >100 ms within a click bout. Both acoustic variables were localized to the tagged animal using angle of arrival (see details in19). Only clicks and buzzes assigned to the tagged whale were included in the analyses, and we followed methods in Arranz et al.33 to ensure that only foraging-related buzzes and not burst-pulses were selected. Each variable was calculated over the duration of an entire dive, with the exception of jerk peak, which was calculated over the bottom phase. Details about the derivation of these variables are published elsewhere19,70. These variables were selected because of their established roles in SRKW foraging activities. Slow echolocation click trains are produced during bouts of prey searching by odontocetes e.g.29,37,39. Buzzes occur during foraging, often just prior to prey interception30,33,36,37. Jerk peak, roll and heading variance have been used together to detect prey capture events with good accuracy19. Depth is a reliable indicator of foraging behavior, with resident-type fish-eating killer whales making deep foraging dives (i.e., often >30 m) to capture prey, particularly salmonids35.
Statistical analysis
We used multivariate Hidden Markov Models (HMMs) to characterize subsurface behavior71. This framework has been previously established for studying behavior in cetaceans46,47,72,73,74,75, and is well suited for analyzing behavioral time series, as it identifies the most likely underlying, non-observable (hidden) states that produce the observed behavior. We utilized a first-order Markov process with N hidden states, in which the probability of being in the current state was determined by the previous state. The six dive variables measured during a dive defined the observed behavior for that dive. The four continuous variables (maximum depth, jerk peak, roll, heading variance) were natural log-transformed and assumed to have an approximate Gaussian distribution, and the two binary variables were modeled with a binomial distribution and logit link. All dive variables were assumed to be independent of each other, which exploratory analysis revealed was a valid assumption, and each tag deployment (tag attached to an individual) was modeled as an independent time series.
We constructed models with three to five underlying hidden states, and did not consider higher order models because the quadratic increase in model parameters would result in biologically uninterpretable models. We did not allow models with six or more states because this was not supported by field observations of SRKW behavioral activities11,56,70,76,77,78. A similar approach of constraining modeling to a pre-determined number of states has been utilized previously, to balance the tradeoffs of statistical vs. biological interpretability47,74,75. Models assumed that observed behavior was conditionally independent given the states (contemporaneous conditional independence). We used the depmixS4 package79 in R v.3.3.3 (R Core Team, Foundation for Statistical Computing, Vienna, Austria) to construct multivariate Hidden Markov Models. This platform allows for flexibility in constructing a variety of dependent mixture models. Each HMM included the six dive variables as response variables, with sex as the single covariate on each response variable. We computed the transition matrix for each model, assuming that all state transitions were possible (no elements fixed at a priori values), and for each model we included one of four covariates on the transition matrix: age class (juvenile or adult), sex, year or a null model without a covariate. We constructed twelve separate HMMs using unique combinations of number of states and a single transition matrix covariate; for interpretability we did not use more than one transition matrix covariate at a time.
We fit the models by maximum likelihood estimation using the expectation-maximization (EM) algorithm in the depmixS4 package, which iteratively maximizes the expected log-likelihood of model parameters given the observations and underlying states. We specified a maximum of 2000 starting values to increase the chance that a solution was found during model fitting. We fit each model 200 times from random initializations to check for numerical stability and robustness due to different starting values, and retained the model with the lowest AIC score. We compared the twelve best models via log-likelihood values and AIC scores. All 5-state models had lower AIC scores than all 4-state models, which had lower scores than all 3-state models (Table 4). The 5-state model with sex as a covariate on the transition matrix had the overall lowest AIC score, and the second lowest log-likelihood value, so we accepted this model (Table 4). Finally, we calculated transition probabilities from the state transition matrix for this model.
We analyzed results of the best HMM in R v.3.5.3. We tested whether dive duration varied by state and sex, given the increased costs of transport for females with calves11 and the known differences in metabolic rate and energetic requirements12, and the potential temporal constraints this could have on behavior. To determine whether dive duration varied by state and sex, we constructed a linear mixed model using the lme480 package with dive duration (natural log-transformed to meet model assumptions) as the response variable, fixed effects of state and sex, and a random effect of deployment, followed by a Type III ANOVA test using the lmerTest package81 to identify significant effects, and a Tukey HSD test with adjusted p-values using the lsmeans82 and multcompView83 packages to compare levels of significant effects. To examine differences estimated between state occurrence by sex, we fit a hierarchical multinomial logistic regression model using the brms package84, with estimated state from the best HMM as the response variable, sex as a fixed effect, and deployment as a random effect. To determine if it was necessary to account for potential effects of deployment duration on the occurrence of the five states, we additionally fit multinomial logistic regression models as above, incorporating ln(deployment duration) as linear effects and as an offset, and compared these models to the one without deployment duration, by examining the posterior estimates of coefficients and comparing estimates using leave-one-out cross-validation (LOOIC)85. We found no strong support for including deployment duration as a predictor or offset in the model. Estimates of LOOIC were extremely similar across models and posterior estimates of the linear effects were uncertain, overlapping 0 in all cases, with large coefficients of variation ranging from 0.65 to 14.7. Therefore, we did not include deployment duration in the final model. The final hierarchical multinomial logistic regression model was run with four MCMC chains, a burn-in period of 2000 samples, and we retained another 1000 samples. The Rhat values of all parameters were 1.0, supporting model convergence86. To examine differences in cumulative time spent in each state by sex, we summed dive duration (in seconds) across state and deployment, for each sex, and implemented a Chi-square test on the cumulative time per state, between females and males.
Ethics statement
The research was conducted in accordance with all Research Permits (USA: NMFS No. 781-1824/16163; Canada: DFO SARA/Marine Mammal License No. MML 2010-01/SARA-106B), and was approved by Northwest Fisheries Science Center’s Institutional Animal Care and Use Committee.
Data availability
The datasets analyzed during the current study are available from the corresponding author upon reasonable request.
References
Sutherland, W. J. The importance of behavioural studies in conservation biology. Anim. Behav. 56, 801–809 (1998).
Caro, T. The behaviour–conservation interface. Trends Ecol. Evol. 14, 366–369 (1999).
Cooke, S. J. Biotelemetry and biologging in endangered species research and animal conservation: relevance to regional, national, and IUCN Red List threat assessments. Endanger. Species Res. 4, 165–185 (2008).
Cooke, S. J. et al. Physiology, behaviour and conservation. Physiol. Biochem. Zool. 87, 1–14 (2014).
Wilson, A. D. M., Wikelski, M., Wilson, R. P. & Cooke, S. J. Utility of biological sensor tags in animal conservation. Conserv. Biol. 29, 1065–1075 (2015).
Groom, M. J., Meffe, G. K. Carroll, C. R. & Andelman, S. J. Principles of Conservation Biology (Sinauer Associates, 2006).
Tuyttens, F. A. M. et al. Observer bias in animal behaviour research: can we believe what we score, if we score what we believe? Anim. Behav. 90, 273–280 (2014).
IUCN. The IUCN Red List of Threatened Species. Version 2019.2, http://www.iucnredlist.org. Accessed 26 August 2019.
Watwood, S. L., Miller, P. J., Johnson, M., Madsen, P. T. & Tyack, P. L. Deep‐diving foraging behaviour of sperm whales (Physeter macrocephalus). J. Anim. Ecol. 75, 814–825 (2006).
Aguilar Soto, N. et al. Cheetahs of the deep sea: deep foraging sprints in short‐finned pilot whales off Tenerife (Canary Islands). J. Anim. Ecol. 77, 936–947 (2008).
Williams, R. & Noren, D. P. Swimming speed, respiration rate, and estimated cost of transport in adult killer whales. Mar. Mam. Sci. 25, 327–350 (2009).
Noren, D. P. Estimated field metabolic rates and prey requirements of resident killer whales. Mar. Mam. Sci. 27, 60–77 (2011).
Ford, J. K. B. & Ellis, G. M. Selective foraging by fish-eating killer whales Orcinus orca in British Columbia. Mar. Ecol. Prog. Ser. 316, 185–199 (2006).
Hanson, M. B. et al. Species and stock identification of prey consumed by endangered southern resident killer whales in their summer range. Endanger. Species Res. 11, 69–82 (2010).
Wright, B. M., Stredulinsky, E. H., Ellis, G. M. & Ford, J. K. Kin-directed food sharing promotes lifetime natal philopatry of both sexes in a population of fish-eating killer whales, Orcinus orca. Anim. Behav. 115, 81–95 (2016).
Candy, J. R. & Quinn, T. P. Behavior of adult Chinook salmon (Oncorhynchus tshawytscha) in British Columbia coastal waters determined from ultrasonic telemetry. Can. J. Zool. 77, 1161–1169 (1999).
Riddell, B. et al. Ocean ecology of Chinook salmon in Ocean Ecology of Pacific Salmon and Trout (ed. Beamish, R. J.) 555–696 (American Fisheries Society, 2018).
Ford, M. J. et al. Status review update for Pacific salmon and steelhead listed under the Endangered Species Act: Pacific Northwest. National Marine Fisheries Service, Northwest Fisheries Science Center, http://www.westcoast.fisheries.noaa.gov/publications/status_reviews/salmon_steelhead/2016_status_review.html (2015).
Tennessen, J. B. et al. Kinematic signatures of prey capture from archival tags reveal sex differences in killer whale foraging activity. J. Exp. Biol. 222, jeb191874 (2019).
Ruckstuhl, K. & Neuhaus, P. Sexual Segregation in Vertebrates. (Cambridge University Press, 2005).
Wearmouth, V. J. & Sims, D. W. Sexual segregation in marine fish, reptiles, birds and mammals: behaviour patterns, mechanisms and conservation implications. Adv. Mar. Biol. 54, 107–170 (2008).
Salton, M., Kirkwood, R., Slip, D. & Harcourt, R. Mechanisms for sex-based segregation in foraging behaviour by a polygynous marine carnivore. Mar. Ecol. Prog. Ser. 624, 213–226 (2019).
Berger-Tal, O. et al. Integrating animal behavior and conservation biology: a conceptual framework. Behav. Ecol. 22, 236–239 (2011).
NMFS. Endangered and threatened wildlife and plants. Endangered status for southern resident killer whales. Fed. Reg. 70, 69903–69912 (2005).
COSEWIC. COSEWIC assessment and update status report on the killer whale Orcinus orca in Canada. Committee on the Status of Endangered Wildlife in Canada. 47 pp (Ottawa, 2001).
NMFS. Southern Resident Killer Whales (Orcinus orca) 5 Year Review: Summary and Evaluation, https://www.fisheries.noaa.gov/resource/document/southern-resident-killer-whales-orcinus-orca-5-year-review-summary-and.72 pp (2016).
Lacy, R. C. et al. Evaluating anthropogenic threats to endangered killer whales to inform effective recovery plans. Sci. Rep. 7, 14119 (2017).
Ward, E. J., Holmes, E. E. & Balcomb, K. C. Quantifying the effects of prey abundance on killer whale reproduction. J. Appl. Ecol. 46, 632–640 (2009).
Johnson, M., Madsen, P. T., Zimmer, W. M., Aguilar de Soto, N. & Tyack, P. L. Beaked whales echolocate on prey. Proc. Royal Soc. B. 271, S383–S386 (2004).
Wisniewska, D. M., Johnson, M., Nachtigall, P. E. & Madsen, P. T. Buzzing during biosonar-based interception of prey in the delphinids Tursiops truncatus and Pseudorca crassidens. J. Exp. Biol. 217, 4279–4282 (2014).
Ydesen, K. S. et al. What a jerk: prey engulfment revealed by high-rate, super-cranial accelerometry on a harbour seal (Phoca vitulina). J. Exp. Biol. 217, 2239–2243 (2014).
Allen, A. N., Goldbogen, J. A., Friedlaender, A. S. & Calambokidis, J. Development of an automated method of detecting stereotyped feeding events in multisensor data from tagged rorqual whales. Ecol. Evol. 6, 7522–7535 (2016).
Arranz, P. et al. Discrimination of fast click-series produced by tagged Risso’s dolphins (Grampus griseus) for echolocation or communication. J. Exp. Biol. 219, 2898–2907 (2016).
DeRuiter, S. L. et al. First direct measurements of behavioural responses by Cuvier’s beaked whales to mid-frequency active sonar. Biol. Lett. 9, 20130223 (2013).
Wright, B. M. et al. Fine-scale foraging movements by fish-eating killer whales (Orcinus orca) relate to the vertical distributions and escape responses of salmonid prey (Oncorhynchus spp.). Mov. Ecol. 5, 3 (2017).
Miller, P. J., Johnson, M. P. & Tyack, P. L. Sperm whale behaviour indicates the use of echolocation click buzzes ‘creaks’ in prey capture. Proc. Royal Soc. B. 271, 2239–2247 (2004).
DeRuiter, S. L. et al. Acoustic behaviour of echolocating porpoises during prey capture. J. Exp. Biol. 212, 3100–3107 (2009).
Madsen, P. T., de Soto, N. A., Arranz, P. & Johnson, M. Echolocation in Blainville’s beaked whales (Mesoplodon densirostris). J. Comp. Physiol. A. 199, 451–469 (2013).
Fais, A. et al. Sperm whale echolocation behaviour reveals a directed, prior-based search strategy informed by prey distribution. Behav. Ecol. Sociobiol. 69, 663–674 (2015).
Arranz, P. et al. Diving behavior and fine-scale kinematics of free-ranging Risso’s dolphins foraging in shallow and deep-water habitats. Front. Ecol. Evol. 7, 53 (2019).
Aoki, K., Sakai, M., Miller, P. J., Visser, F. & Sato, K. Body contact and synchronous diving in long-finned pilot whales. Behav. Processes 99, 12–20 (2013).
Urick, R. J. Principles of Underwater Sound. 2nd ed. (McGraw-Hill, 1983).
Richardson, W. J., Greene, C. R. Jr., Malme, C. I. & Thomson, D. H. Marine Mammals and Noise (Academic Press, 1995).
Miller, P. J. Diversity in sound pressure levels and estimated active space of resident killer whale vocalizations. J. Comp. Physiol. A. 192, 449–459 (2006).
Holt, M. M., Noren, D. P. & Emmons, C. K. Effects of noise levels and call types on the source levels of killer whale calls. J. Acoust. Soc. Am. 130, 3100–3106 (2011).
Isojunno, S. et al. Individual, ecological, and anthropogenic influences on activity budgets of long‐finned pilot whales. Ecosphere 8, e02044 (2017).
Quick, N. J. et al. Hidden Markov models reveal complexity in the diving behaviour of short-finned pilot whales. Sci. Rep. 7, 45765 (2017).
Friedlaender, A. S. et al. Diel changes in humpback whale Megaptera novaeangliae feeding behavior in response to sand lance Ammodytes spp. behavior and distribution. Mar. Ecol. Prog. Ser. 395, 91–100 (2009).
Hazen, E. L. et al. Fine-scale prey aggregations and foraging ecology of humpback whales Megaptera novaeangliae. Mar. Ecol. Prog. Ser. 395, 75–89 (2009).
Friedlaender, A. S. et al. Whale distribution in relation to prey abundance and oceanographic processes in shelf waters of the Western Antarctic Peninsula. Mar. Ecol. Prog. Ser. 317, 297–310 (2006).
Johnston, D. W. et al. Temporal patterns in the acoustic signals of beaked whales at Cross Seamount. Biol. Lett. 4, 208–211 (2008).
Baird, R. W., Webster, D. L., Schorr, G. S., McSweeney, D. J. & Barlow, J. Diel variation in beaked whale diving behavior. Mar. Mam. Sci. 24, 630–642 (2008).
Gill, P. C. et al. Blue whale habitat selection and within-season distribution in a regional upwelling system off southern Australia. Mar. Ecol. Prog. Ser. 421, 243–263 (2011).
Noren, S. R. & Williams, T. M. Body size and skeletal muscle myoglobin of cetaceans: adaptations for maximizing dive duration. Comp. Biochem. Physiol. A 126, 181–191 (2000).
Constantine, R., Brunton, D. H. & Dennis, T. Dolphin-watching tour boats change bottlenose dolphin (Tursiops truncatus) behaviour. Biol. Cons. 117, 299–307 (2004).
Lusseau, D., Bain, D. E., Williams, R. & Smith, J. C. Vessel traffic disrupts the foraging behavior of southern resident killer whales Orcinus orca. Endanger. Species Res. 6, 211–221 (2009).
Christiansen, F., Rasmussen, M. & Lusseau, D. Whale watching disrupts feeding activities of minke whales on a feeding ground. Mar. Ecol. Prog. Ser. 478, 239–251 (2013).
Senigaglia, V. et al. Meta-analyses of whale-watching impact studies: comparisons of cetacean responses to disturbance. Mar. Ecol. Prog. Ser. 542, 251–263 (2016).
Hoelzel, A. R. Foraging behaviour and social group dynamics in Puget Sound killer whales. Anim. Behav. 45, 581–591 (1993).
Perelberg, A. & Schuster, R. Coordinated breathing in bottlenose dolphins (Tursiops truncatus) as cooperation: Integrating proximate and ultimate explanations. J. Comp. Psych. 122, 109–120 (2008).
Sakai, M., Morisaka, T., Kogi, K., Hishii, T. & Kohshima, S. Fine-scale analysis of synchronous breathing in wild Indo-Pacific bottlenose dolphins (Tursiops aduncus). Behav. Processes 83, 48–53 (2009).
Senigaglia, V., de Stephanis, R., Verborgh, P. & Lusseau, D. The role of synchronized swimming as affiliative and anti-predatory behavior in long-finned pilot whales. Behav. Processes 91, 8–14 (2012).
Senigaglia, V. & Whitehead, H. Synchronous breathing by pilot whales. Mar. Mam. Sci. 28, 213–219 (2012).
Acevedo-Gutiérrez, A., Croll, D. A. & Tershy, B. R. High feeding costs limit dive time in the largest whales. J. Exp. Biol. 205, 1747–1753 (2002).
Tyack, P. L., Johnson, M., Soto, N. A., Sturlese, A. & Madsen, P. T. Extreme diving of beaked whales. J. Exp. Biol. 209, 4238–4253 (2006).
Whitehead, H. Babysitting, dive synchrony, and indications of alloparental care in sperm whales. Behav. Ecol. Sociobiol. 38, 237–244 (1996).
Augusto, J. F., Frasier, T. R. & Whitehead, H. Characterizing alloparental care in the pilot whale (Globicephala melas) population that summers off Cape Breton, Nova Scotia, Canada. Mar. Mam. Sci. 33, 440–456 (2017).
Johnson, M. P. & Tyack, P. L. A digital acoustic recording tag for measuring the response of wild marine mammals to sound. IEEE J. Oceanic. Eng. 28, 3–12 (2003).
Holt, M. M., Hanson, M. B., Giles, D. A., Emmons, C. K. & Hogan, J. T. Noise levels received by endangered killer whales Orcinus orca before and after implementation of vessel regulations. Endanger. Species Res. 34, 15–26 (2017).
Holt, M. M., Noren, D. P. & Emmons, C. K. An investigation of sound use and behavior in a killer whale (Orcinus orca) population to inform passive acoustic monitoring studies. Mar. Mam. Sci. 29, E193–E202 (2012).
Zucchini, W., MacDonald, I. L. & Langrock, R. Hidden Markov Models for Time Series: an Introduction using R (Chapman and Hall/CRC, 2016).
Isojunno, S. & Miller, P. J. Sperm whale response to tag boat presence: biologically informed hidden state models quantify lost feeding opportunities. Ecosphere 6, 1–46 (2015).
Isojunno, S. et al. Sperm whales reduce foraging effort during exposure to 1–2 kHz sonar and killer whale sounds. Ecol. Appl. 26, 77–93 (2016).
Quick, N., Scott-Hayward, L., Sadykova, D., Nowacek, D. & Read, A. Effects of a scientific echo sounder on the behavior of short-finned pilot whales (Globicephala macrorhynchus). Can. J. Fish. Aquat. Sci. 74, 716–726 (2016).
DeRuiter, S. L. et al. A multivariate mixed hidden Markov model for blue whale behaviour and responses to sound exposure. Ann. Appl. Stat. 11, 362–392 (2017).
Noren, D. P., Johnson, A. H., Rehder, D. & Larson, A. Close approaches by vessels elicit surface active behaviors by southern resident killer whales. Endang. Species Res. 8, 179–192 (2009).
Williams, R., Lusseau, D. & Hammond, P. S. Estimating relative energetic costs of human disturbance to killer whales (Orcinus orca). Biol. Conserv. 133, 301–311 (2006).
Williams, R., Erbe, C., Ashe, E., Beerman, A. & Smith, J. Severity of killer whale behavioral responses to ship noise: A dose–response study. Mar. Poll. Bull. 79, 254–260 (2014).
Visser, I. & Speekenbrink, M. depmixS4: An R package for hidden Markov models. J. Stat. Softw. 36, 1–21, http://www.jstatsoft.org/v36/i07/ (2010).
Bates, D., Maechler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48, https://doi.org/10.18637/jss.v067.i01 (2015).
Kuznetsova, A., Brockhoff, P. B. & Christensen, R. H. B. lmerTest Package: Tests in linear mixed effects models. J. Stat. Softw. 82, 1–26, https://doi.org/10.18637/jss.v082.i13 (2017).
Lenth, R. V. Least-squares means: The R Package lsmeans. J. Stat. Softw. 69, 1–33, https://doi.org/10.18637/jss.v069.i01 (2016).
Graves, S., Piepho, H.-P., Selzer, L. & Dorai-Raj, S. multcompView: Visualizations of paired comparisons. R package, version 0.1–7, https://CRAN.R-project.org/package=multcompView (2015).
Bürkner, P.-C. brms: An R package for Bayesian multilevel models using Stan. J. Stat. Softw. 80, 1–28, https://doi.org/10.18637/jss.v080.i01 (2017).
Vehtari A., Gabry, J., Yao, Y. & Gelman, A. loo: Efficient leave-one-out cross-validation and WAIC for Bayesian models. R package version 2.1.0, https://CRAN.R-project.org/package=loo (2019).
Gelman, A. & Rubin, D. B. Inference from iterative simulation using multiple sequences. Stat. Sci. 7, 457–472 (1992).
Acknowledgements
We thank M. Johnson, S. DeRuiter, A. Bocconcelli, A. Allen, F. Jensen, A. Stimpert, P. Wensveen, P. Miller and D. Haas for assistance with tagging logistics and data processing, and the University of Washington’s Friday Harbor Laboratory for field support. We thank D. Noren, M. Ford, R. Baird and T. Tennessen for valuable feedback. We are grateful for the assistance provided by many dedicated field volunteers and technicians. Any use of trade, firm or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government. This work was supported by funding from the National Oceanic and Atmospheric Administration’s Ocean Acoustics Program (to M.M.H. and M.B.H.) and Northwest Fisheries Science Center, the National Fish and Wildlife Foundation’s Killer Whale Research and Conservation Program with support from SeaWorld Entertainment, and the Port of Vancouver.
Author information
Authors and Affiliations
Contributions
M.M.H. and M.B.H. secured funding and designed the study. M.M.H., M.B.H., C.K.E., D.A.G. and J.T.H. conducted the fieldwork. J.B.T., M.M.H. and E.J.W. analyzed the data. J.B.T. wrote the manuscript. All authors edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tennessen, J.B., Holt, M.M., Ward, E.J. et al. Hidden Markov models reveal temporal patterns and sex differences in killer whale behavior. Sci Rep 9, 14951 (2019). https://doi.org/10.1038/s41598-019-50942-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-50942-2
- Springer Nature Limited