
Abstract
Plant cell wall degrading enzymes (PCWDEs) are the primary virulence determinants of soft rotting bacteria such as the potato pathogen, Pectobacterium atrosepticum. The regulation of secondary metabolite (Rsm) system controls production of PCWDEs in response to changing nutrient conditions. This work identified a new suppressor of an rsmB mutation – ECA1172 or rsmS (rsmB suppressor). Mutants defective in rsmB (encoding a small regulatory RNA), show reduced elaboration of the quorum sensing molecule (N-3-oxohexanoyl-homoserine lactone; OHHL) and PCWDEs. However, OHHL and PCWDE production were partially restored in an rsmB, rsmS double mutant. Single rsmS mutants, overproduced PCWDEs and OHHL relative to wild type P. atrosepticum and exhibited hypervirulence in potato. RsmS overproduction also resulted in increased PCWDEs and OHHL. Homology searches revealed rsmS conservation across pathogens such as Escherichia coli (ybaM), Dickeya solani, Klebsiella pneumoniae and Shigella flexneri. An rsmS mutant of Pectobacterium carotovorum ATCC39048 showed bypass of rsmB-dependent repression of PCWDEs and OHHL production. P. carotovorum ATCC39048 produces the β-lactam antibiotic, 1-carbapen-2-em-3-carboxylic acid (a carbapenem). Production of the antibiotic was repressed in an rsmB mutant but partially restored in an rsmB, rsmS double mutant. This work highlights the importance of RsmS, as a conserved pleiotropic regulator of virulence and antibiotic biosynthesis.
Similar content being viewed by others

Introduction
Members of the Pectobacteriaceae are the causative agents of soft rot in potato tubers and blackleg in potato plants. These Gram-negative bacteria, such as Pectobacterium atrosepticum SCRI1043 (Pba) and Pectobacterium carotovorum (Pcc), are responsible for diminished yields due to diseased plants, seed potato infection, and soft rot decay of tubers post-harvest during storage. Both soft rot and blackleg diseases are caused by the secretion of plant cell wall degrading enzymes (PCWDEs), such as pectate lyases, polygalacturonase, cellulases and proteases1. Flagellum-dependent motility is also important for plant virulence1,2. The production and secretion of PCWDEs and other virulence determinants is tightly controlled by many interconnected regulatory networks including an acyl-homoserine lactone quorum sensing (QS) system and the regulation of secondary metabolite (Rsm) system3,4,5.
Cell density-dependent expression of PCWDEs in Pba and Pcc is regulated by the ExpI-VirR (ExpR2 in some species) QS system3,6,7,8. ExpI, an acyl-homoserine lactone synthase, produces the freely diffusible signalling molecule N-3-(oxohexanoyl)-homoserine lactone (OHHL) which acts as a proxy for population density, and VirR represses transcription of target genes3. Above a threshold concentration, OHHL interacts with VirR, relieving repression, resulting in transcription of genes encoding PCWDEs, siderophore biosynthesis, and flagellar motility3,6,7,9. Furthermore, in the absence of OHHL, VirR binds the promoter of rsmA, another important virulence regulator, activating its transcription and linking the Rsm and QS systems9.
The Rsm regulatory system acts post-transcriptionally to regulate translation of target mRNAs by blocking the Shine-Dalgarno site and influencing transcript stability4,6,7. The primary component of the system is the RNA-binding protein RsmA (CsrA in Escherichia coli). In Pectobacterium spp., RsmA interacts with transcripts for virulence determinants, thereby influencing in planta infection10. RsmA/CsrA binds to GGA motifs of specific transcripts, blocking ribosomal access, resulting in decreased translation11,12. RsmA-binding can be influenced by the untranslated RNA, rsmB13. When processed and folded, rsmB forms a multi-stem-loop structure with GGA motifs exposed at the end of each loop. Therefore, a single copy of rsmB can titrate many copies of RsmA simultaneously and this stoichiometry has a crucial impact on regulatory functionality14,15. In Pba and Pcc, an rsmB mutant shows constitutively active RsmA and produces almost no PCWDEs; even less than an expI, QS-defective, mutant13,14,16.
A previous study attempted to identify a mutation in rsmA by isolating suppressors of rsmB16. Unexpectedly, this strategy identified a mutant with a transposon insertion in metJ that was found to suppress the rsmB mutation. MetJ is a metabolic regulator known to bind DNA at met-boxes, often found upstream of methionine biosynthetic genes17,18,19,20. While there had been several studies investigating metJ and its role in metabolism20,21,22, Cubitt et al. demonstrated a link between MetJ, virulence, and the Rsm system16. Furthermore, a mutation in metJ resulted in pleiotropic transcriptional changes, suggesting that it affected multiple cellular pathways16. We postulated that there might be other suppressors of rsmB, still to be identified.
To investigate suppression of the Rsm system further, a random transposon mutagenesis of an rsmB mutant of Pba was conducted and insertion mutants with increased protease (caseinase) production were selected for further analysis. From this screen, transposon insertions in priC and ECA1172, a small open reading frame (ORF) of unknown function immediately downstream of priC, were identified. We demonstrated that suppression of rsmB was via mutation in ECA1172, a gene we named rsmS for rsmB suppressor.
Results
Identification of ECA1172, a new regulator of rsmB-dependent virulence repression
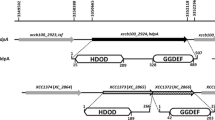
A Pba rsmB mutant produces no detectable protease activity16. To identify suppressors of this phenotype, over 20 000 random transposon insertion mutants in an rsmB mutant background were screened for restored protease production. A single transposon insertion 12 bp within the priC gene, (encoding the primosomal replication protein N), and several insertions in the ORF immediately downstream, ECA1172 (a gene of unknown function) were identified (Fig. 1a). Each of these transposon insertions partially restored protease production (Fig. 1 and Supplemental Fig. 1). As the ECA1172 insertion produced an rsmB suppressor phenotype, we named it rsmS. Both priC and rsmS were novel suppressors of rsmB-dependent repression of PCWDEs and, therefore, warranted further characterization.

Mutation of rsmS affects biosynthesis of PCWDEs. (a) Genomic context of rsmS within Pba. Genes are indicated with arrows and names are written within. The numbers below indicate the number of bp between each gene. (b) The priC-rsmS promoter region. The −10 and −35 sites are indicated in bold. Putative binding sites for Crp, OxyR, and Lrp are indicated with a line under the site. A putative ArcA binding site is indicated in italics as it overlaps both the OxyR and −10 site. Putative Shine-Dalgarno sites are indicated as RBS and the translational start and stop sites are indicated in bold. (c) PCWDE plate assays showing production of protease (left), pectate lyase (middle) and cellulase (right) for the wild type, rsmB (NW155), rsmS (KA32) or rsmB rsmS (KA35) after incubation at 25 °C.
Mutation of priC causes polar effects on rsmS
Genomic analysis suggested that the priC and rsmS genes were within an operon (Fig. 1a). The two ORFs were separated by 37 bp, transcribed in the same direction, and we identified a candidate promoter upstream of priC but no promoter within 250 bp of the rsmS translational start site (Fig. 1b). The transposon used in the screen contained a transcriptional terminator23, thus, if priC and rsmS were in an operon, a transposon insertion within priC, would also affect production of the rsmS product. To determine if suppression of rsmB observed in the priC transposon mutant was due to a polar effect on rsmS, protease production was observed when a plasmid-borne copy of either priC or rsmS was expressed in a priC mutant. No change in production was detected when PriC was expressed from a plasmid, but protease production decreased when RsmS was expressed in a priC mutant background, suggesting that the mutation had been complemented (Supplemental Fig. 1). Furthermore, we had identified transposon insertions within rsmS¸ the ORF immediately downstream, which also partially restored protease production in an rsmB mutant. We concluded that the two genes were transcribed as an operon and that it was the downstream effect on rsmS in the priC mutant that partially suppressed the rsmB mutation.
Mutation of rsmS restores production of PCWDEs in an rsmB mutant
To exclude the possibility of a secondary mutation elsewhere in the chromosome affecting protease production, a new mutation in rsmS was created by allelic exchange and was transduced into an rsmB mutant (creating strain KA35) and into wild type Pba (creating KA32). Production of protease was still partially restored in KA35 (rsmB, rsmS), suggesting that it was the deletion of rsmS and not another mutation that was responsible for this phenotype (Fig. 1c). We continued to work with KA35 and KA32 for the remainder of this study. As we had been able to phenotypically complement a mutation in priC with a copy of rsmS in trans, we knew that this plasmid-borne copy of rsmS was functional. We introduced the same plasmid into KA35 (rsmB, rsmS) and found that protease production was no longer detectable (Supplemental Fig. 2a). We concluded that mutation of rsmS alone was capable of restoring protease production in an rsmB mutant and that expression of rsmS in trans complemented this phenotype.
Cellulase and pectate lyase are both important for potato infection and production of both enzymes is significantly reduced in an rsmB mutant1,16. As observed for protease production above, in an rsmB, rsmS double mutant, significantly more cellulase or pectate lyase activity was detected than in an rsmB mutant (Fig. 1c). Though enzymatic activity was not as high as in wild type Pba, an rsmS, rsmB double mutant produced more PCWDEs than a strain mutated for rsmB alone (Fig. 1c). At the same time, we examined strain KA32 (rsmS) and found that secreted enzyme levels were increased, relative to wild type (Fig. 1c). We concluded that RsmS repressed PCWDEs in Pba.
Swimming motility is an important virulence determinant in Pba2,10,24, thus we examined whether mutation of rsmS affected flagellum-dependent swimming. Using low percentage agar, we found no significant difference in swimming motility between wild type Pba, an rsmB mutant, an rsmS mutant or an rsmB rsmS double mutant (data not shown). This observation was consistent with other work examining rsmB mutant phenotypes in Pba16.
An rsmS mutation bypasses repression of PCWDEs and OHHL in an rsmB mutant
To ascertain if growth rate or cell density-dependent expression of virulence factors was altered by mutation of rsmS, the activities of PCWDEs throughout growth in wild type, KA32 (rsmS), KA35 (rsmB, rsmS) or NW155 (rsmB) strains were determined. No significant differences in optical density or growth rate were observed between any of the strains, suggesting that mutation of rsmS caused no discernible growth defect in these media (Fig. 2). Sterile supernatant samples, taken at each time point, were analysed for cellulase, pectate lyase or protease activity. Mutants defective in rsmB showed reduced levels of all enzymes compared to those in wild type Pba. Supernatants taken from strain KA35 (rsmB, rsmS) showed significantly greater enzyme activity than those observed in an rsmB mutant (Fig. 2). These data were consistent with observations on plates and in the original screen: a mutation in rsmS phenotypically suppressed the effects of an rsmB mutation.

An rsmS mutation alters PCWDE and OHHL production. Levels of protease (a), cellulase (b), pectate lyase (c) and OHHL (d) in the culture supernatant determined at two-hour intervals throughout growth in PMB at 25 °C. Wild type (black), rsmS (KA32, blue), rsmB (NW155, green) and rsmB rsmS (KA35, red) cultures were assessed for growth (OD600 – dashed lines) and sterile supernatant samples taken every two hours. Solid lines indicate the enzyme activity or relative OHHL amounts determined from sterile supernatants. Values indicate the average +/− SD (n = 3).
We found that mutation of rsmS in an otherwise wild type background resulted in significantly increased secreted enzymes but no growth defect, reinforcing the findings of earlier plate-based assays (Figs 1 and 2). For example, after 16 hours of growth, secreted pectate lyase activity of an rsmS mutant was 2.5-fold higher than that of wild type Pba (Fig. 2c). Similar patterns were observed for cellulase and protease production (Fig. 2a,b). These data, together with plate-based assays, suggested that, although rsmS was originally identified as a suppressor of rsmB in double mutants, an rsmS mutation may be sufficient to cause hypervirulence (Fig. 2).
Mutation of rsmS alters levels of the QS signalling molecule OHHL
An rsmB mutant of Pba produces reduced amounts of OHHL16. We wanted to determine whether mutation of rsmS, in an rsmB mutant background, perturbed OHHL production, as this would influence production of PCWDEs. The same sterile supernatant samples (see previous section) were assessed for the presence of OHHL using a bioluminescent reporter assay in Escherichia coli25. In an rsmB, rsmS double mutant, production of OHHL was indistinguishable from that of wild type (Fig. 2d). Therefore, in addition to suppressing the effects of rsmB on production of virulence determinants, an rsmS mutation completely relieved rsmB-dependent repression of OHHL production. In the rsmS mutant, OHHL biosynthesis was significantly greater than that observed in wild type cultures (Fig. 2d), suggesting that RsmS was also a repressor of OHHL production (and thus QS).
Overexpression of rsmS resulted in increased enzyme secretion
The plasmid-based expression system (pBAD30) used for complementing protease production in an rsmS mutant, provided rsmS on a plasmid under the control of the Para promoter (Supplemental Fig. 2). Expression from this promoter is induced by increasing arabinose concentrations26. Using pBAD30-rsmS, we assessed protease production when rsmS was overexpressed in wild type Pba, an rsmB mutant, an rsmB, rsmS double mutant or an rsmS mutant. In all strains, overexpression of rsmS led to an increase in secreted protease activity, though again this was difficult to quantify on a plate (Supplemental Fig. 2). Surprisingly, even overexpression of rsmS in an rsmB mutant resulted in increased protease production (Supplemental Fig. 2). Therefore, these results showed that both loss of RsmS or an excess of RsmS resulted in increased secreted protease.
To investigate whether increased RsmS altered other virulence-associated phenotypes, we examined pectate lyase, cellulase and OHHL production in sterile culture supernatants from wild type Pba or an rsmS mutant carrying either pBAD30 (empty vector) or pBAD30-rsmS under increasing levels of induction. In wild type cultures, even in uninduced conditions, all three phenotypes were elevated when RsmS was expressed from a plasmid, compared to the empty vector control (Fig. 3a–c). As production of RsmS was induced, a progressive increase in cellulase and pectate lyase activity was observed (Fig. 3a,b). In contrast, OHHL levels increased when an extra copy of rsmS was present on a plasmid but did not change upon RsmS induction. These data suggested that, in wild type Pba, even a low level of rsmS transcription, through leaky expression and without induction, resulted in maximal OHHL production (Fig. 3c). In an rsmS mutant containing pBAD30-rsmS under uninduced conditions, reduced levels of OHHL, pectate lyase and cellulase were observed. These phenotypes were indistinguishable from those of wild type Pba, indicating that the mutation had been complemented. However, when RsmS was induced, levels of cellulase, pectate lyase and OHHL increased back to those observed in an rsmS mutant without induction (Fig. 3). Together these results showed that the relative abundance of RsmS in the cell, and not just its absence, impacted the production of PCWDEs and OHHL.

Perturbation of RsmS alters production of PCWDEs and OHHL. Pectate lyase (a), cellulase (b) or OHHL (c) production in wild type Pba (WT) or an rsmS mutant (KA32) carrying either pBAD30 or pBAD-rsmS grown in PMB with 0% arabinose (circle), 0.01% arabinose (square) or 0.1% arabinose (triangle) for 16 hours at 25 °C. Plots all show individual data points, the mean and error bars indicate +/− SD (n = 3).
Mutation of rsmS results in increased potato rot
Pba causes tuber rot1. Our earlier results demonstrated that mutation of rsmS resulted in increased production of PCWDEs (Fig. 2). Consequently, we assessed in planta virulence of this strain. Surface sterilized potatoes were inoculated with wild type Pba and an rsmS mutant at two inoculation sites on opposite sides of the potato. After four days of growth, the level of rot was measured at the inoculation sites. The weight of rot at the rsmS inoculation site was significantly larger than at the wild type site (Fig. 4), confirming our earlier results that the rsmS mutant was hypervirulent.

Mutation of rsmS alters in planta virulence. Values plotted represent the average amounts of potato rot (+/− SD) harvested from wild type (blue) and rsmS (KA32, green) inoculations after four days of incubation at 25 °C from four potato tubers.
RsmS is conserved across many model organisms, though its genomic context can differ
Given the importance of RsmS in regulating plant virulence in Pba and the widespread presence of the Rsm/Csr regulation system across different bacterial species4,27,28, we determined whether rsmS was also widely distributed. We identified rsmS homologues (or ybaM as it is designated in E. coli) across many different bacteria and found that the gene was always immediately downstream of priC (Figs 5 and 6). The predicted size of RsmSPba was 6.1 kDa and, using Pfam29, we were able to identify a single conserved domain pfam10689 spanning amino acids 2–44. However, this domain has no known function. We aligned the RsmS amino acid sequences from many different bacteria and found that, at the amino acid level, the sequence had been conserved, especially in the region covered by the pfam10689 domain (Fig. 6). Additionally, a Shine-Dalgarno sequence was identified immediately upstream of rsmS, suggesting that the ORF can be translated (Fig. 1). We examined the promoter region of priC in Pba to determine whether there were any known regulator binding sites in this area. We identified putative −10 and −35 sites and potential binding sites for the regulators Crp, OxyR, ArcA and Lrp, suggesting that some, or all, of these regulators may influence transcription of priC, and thus, rsmS (Fig. 1b).

RsmS is conserved in different bacteria. Amino acid (a) and nucleotide (b) alignments of RsmS or rsmS from Pba, Pcc39048, D. solani MK10, K. pneumoniae, E. coli K 12 and S. flexneri 2a 2457 T. In (a) positions conserved across all sequences are indicated with an * below, amino acids that share strongly similar properties across all sequences are indicated with a :, and a . indicates amino acids that share weakly similar properties across the sequences. (b) Nucleotides that are conserved across all sequences are indicated by * and nucleotides that did not align and represent a gap are indicated with a -.

Genomic contexts of rsmS in different bacterial species. Arrows are used to indicate each ORF in the surrounding area of rsmS with the ORF name inside. Red arrows indicate rsmS or its homologue in each organism. ORFs that are similar are indicated in the same colour of text. Repeat regions are indicated with stick arrows.
Mutation of rsmS is also capable of suppressing an rsmB mutant in Pcc39048
Our initial focus had been the Pba Rsm system, however, in the related phytopathogen Pcc, extensive research has been undertaken to understand the Rsm system4,13,14,30. We were particularly interested in the strain Pcc39048, as it produces a simple carbapenem antibiotic31. Pcc39048 contained homologues of rsmS (Fig. 5) and rsmB, though neither had been investigated. To isolate a mutant of rsmB, random transposon mutagenesis of Pcc39048 was first undertaken and colonies were screened for decreased protease (caseinase) production. A transconjugant containing a transposon insertion within rsmB was identified using this method. The transposon insertion was transduced back into wild type Pcc39048, generating strain SBEB. The resulting transductants exhibited no protease production and reduced cellulase and pectate lyase production (Fig. 7a).

Mutation of rsmS affects virulence in Pcc39048. (a) Protease (left), pectate lyase (middle) and cellulase (right) of wild type, rsmB (SBEB), rsmS (SBES) and rsmB rsmS (SBEBS) mutants. Production of each enzyme is indicated by a halo surrounding the strain spot. (b) Carbapenem production of wild type, rsmB, rsmS and rsmB, rsmS double mutants. OHHL (c) and Carbapenem production (d) in Pcc39048 of wild type (black), rsmB (green), rsmS (blue) and rsmB, rsmS (red) throughout growth in PMB at 30 °C. Strains were monitored for optical density (Dashed line, OD600) and supernatant samples were examined for OHHL and carbapenem activity (solid lines). Plotted points indicate the average value of each data point +/− SD (n = 3).
A second transposon mutagenesis was performed on the rsmB mutant (SBEB) in Pcc39048, to identify random transposon insertions capable of restoring caseinase production. Over 14 000 colonies were screened and two showing increased protease activity relative to the progenitor strain (SBEB) were chosen for further analysis. One of these transposon insertions was within hexY (rsmC). Mutations in hexY have been identified previously, and are known to bypass the Rsm system16,32,33. The second transposon insertion was within a small ORF encoding a protein showing 82% amino acid identity to RsmS from Pba (Fig. 6a). The surrounding genomic location was the same in Pba and Pcc39048 (Fig. 5). The transposon in rsmS was transduced back into the rsmB mutant and into the wild type background. We examined these strains for pectate lyase, cellulase and protease activity and found that mutation of rsmS phenotypically suppressed the effects of the rsmB mutation (Fig. 7a). These data confirmed that RsmS was conserved between Pcc39048 and Pba and had the same function in both bacterial species.
PCWDE expression in Pcc39048 is also under QS control3,34 and our experiments in Pba suggested that mutation of rsmS affected OHHL levels. In Pcc39048, we found that OHHL production was reduced in an rsmB mutant but partially restored in an rsmB, rsmS double mutant (Fig. 7c). In an rsmS mutant, OHHL levels were slightly higher than in wild type, though not significantly different.
Carbapenem antibiotic production is affected by both rsmB and rsmS
Carbapenem production is under QS control in Pcc3904835. As the Rsm system affected production of OHHL, it was expected that rsmS and rsmB might also affect carbapenem production. Carbapenem activity was assessed as growth inhibition surrounding a test culture on an E. coli super sensitive (ESS) top lawn. Mutants of rsmB showed reduced haloes of inhibition and an rsmB, rsmS double mutant showed wild type carbapenem biosynthesis. No significant difference in antibiotic production was observed between the rsmS mutant and wild type (Fig. 7b). To gain information on carbapenem production throughout growth, sterile supernatant samples taken at regular time intervals were assayed for antibiotic activity and compared to those of wild type. In an rsmB mutant, carbapenem production was less than that of wild type throughout growth. This was partially restored in an rsmB, rsmS double mutant. Carbapenem production was slightly increased and activity was observed earlier in the growth curve in an rsmS mutant, following a similar pattern to that observed for PCWDEs in Pba (Fig. 7d).
Discussion
This work describes the identification and characterization of RsmS, a novel input to the Rsm system in both Pba and Pcc39048. Using a previously successful rsmB suppression screen16, several transposon insertions within the priC-rsmS operon were identified. The transposon used contained a transcriptional terminator, therefore insertions within priC also knocked out rsmS by polarity. Furthermore, we were only able to complement a priC insertion with rsmS, consistent with the operonic organization (Supplemental Fig. 1). This genetic organization is conserved across many different bacterial species, including E. coli. The one exception is another potato pathogen, D. solani, where an additional ORF, miaE, is found upstream of priC, perhaps forming a single miaE-priC-rsmS operon (Fig. 6). Furthermore, a study in E. coli examining knockouts of small proteins, found that mutation of ybaM (a homologue of rsmS) required the promoter upstream of priC for complementation36.
Given our identification of an insertion mutation in priC, it appears that it is not essential in Pba. More importantly, as priC and rsmS appear to be operonic, they are likely coregulated in many different bacterial species. As our bioinformatic interrogation identified, several conserved candidate regulator-binding sites were identified in the priC promoter region, including for Crp, OxyR and ArcA (Fig. 1). The impact of each of these regulators remains to be investigated, but the presence of putative binding sites suggests that this priC-rsmS operon is highly regulated in response to different environmental and nutritional inputs.
Small proteins such as RsmS/YbaM have been understudied37,38. However, proteomic techniques such as ribosome profiling have facilitated the identification of small proteins in a systematic way and research has begun to examine their roles in microbial physiology39,40,41. Some of the most well studied small proteins regulated sporulation in Bacillus subtilis, such as Sda, SpoVM and CmpA. Sda inhibits the kinase activity of KinA, part of the phosphorelay system that regulates sporulation, in response to DNA damage42,43. Another well studied small protein is AcrZ, a small E. coli protein associated with the AcrAB-TolC efflux pump43. Mutants defective in acrZ are more sensitive to tetracycline, puromycin and chloramphenicol but not all antibiotics transported by the AcrAB-TolC efflux pump, suggesting that this small protein alters the specificity of the pump44. Here we identified a new small protein RsmS: an important regulator of virulence and antibiotic production in Pectobacterium. With their varied roles in the cell and importance in drug resistance, virulence and other physiological impacts, the biology of small proteins clearly deserves further study and interrogation.
In Pba and Pcc, the QS system is also an important regulator of virulence factor production. Previous work demonstrated that OHHL production was significantly reduced in an rsmB mutant13,16. Addition of exogenous OHHL does not restore PCWDE production in an rsmB mutant, suggesting that the mutation does not act solely through the QS system16. During this work, we found that OHHL production was partially restored in an rsmB, rsmS mutant and that precocious OHHL production was observed in an rsmS mutant in both Pba and Pcc39048. Other studies have observed that addition of excess OHHL alone does not generally induce PCWDEs precociously3,45,46. Thus, although mutation of rsmS restored OHHL production in an rsmB mutant background, this is unlikely to be responsible for all the phenotypic changes observed. It is interesting in this context that a study investigating ppGpp-dependent regulation of PCWDEs in Pba also identified a mutation in rsmS (then referred to as ECA1172) as one that was capable of bypassing virulence suppression in a relA/spoT mutant background46.
While rsmS, or its homologues in other organisms, has not been examined extensively, altered ybaM (rsmS) transcription has been observed as part of transcriptomic studies investigating unrelated phenotypes. For example, under simulated microgravity in E. coli, ybaM transcript abundance was significantly increased47. Work examining changes in the E. coli commensal transcriptome during intestinal inflammation or colitis in mice found that ybaM mRNAs decreased 2.2-fold in IL10(−/−) mice, those suffering from colonic inflammation, compared to wild type mice48. Another study (mentioned above) created a ybaM mutant as part of an effort to systematically examine E. coli small proteins of unknown function. That work reported that a mutation of ybaM resulted in a competitive disadvantage compared to wild type E. coli when exposed to cell envelope stress, though this was not investigated further36. Notably, a csrA mutant is also more susceptible to cell envelope stress, though the priC-rsmS locus has not been identified as a binding target of CsrA12. In the present work, widespread changes in virulence phenotypes upon rsmS deletion or overexpression were observed. Taken together with the studies from E. coli, it is likely that rsmS is an important part of enterobacterial/pectobacterial stress response regulons and worthy of further interrogation in different model organisms.
Several studies have identified, indirectly, links between YbaM and the E. coli Csr regulatory system. For example, the half-life of ybaM is significantly reduced in a csrA51 mutant and the abundance of priC and ybaM mRNAs are halved in a csrD mutant - another component of the Rsm/Csr regulatory system27. Perhaps most interesting, given our work in Pba, is the experimental evidence suggesting YbaM and CsrA interact. This was found as part of a report establishing the complete protein-protein interaction network in E. coli using yeast two-hybrid system analysis49. Though the finding has not been validated using complementary techniques, it suggests a connection exists between YbaM/RsmS and the Csr/Rsm regulatory systems.
Taking our experimental results together with published literature, we propose the following tentative model in Pba (Fig. 8). Under normal conditions, in the absence of rsmB, RsmA binds to target transcripts blocking or activating their translation. When rsmB is transcribed, it can titrate many copies of RsmA, ultimately leading to expression of PCWDEs. In the absence of rsmB, no competition exists for RsmA and it can continually repress expression of PCWDEs. However, when a mutation in rsmB is combined with one in rsmS, partial restoration of PCWDEs is observed, suggesting that RsmA binding in this strain background may be less efficient. Finally, in a strain mutated in rsmS alone, precocious induction of OHHL and increased PCWDEs are observed. One possibility is that RsmS is required for efficient RsmA binding to target transcripts but not essential for binding of rsmB. Thus, if RsmS is absent or is in excess, RsmA is not able to efficiently access its binding site on a particular transcript, favouring binding to rsmB. Another possible explanation is that RsmS affects the stability of RsmA, targeting it for degradation. Alternatively, RsmS may act via an undiscovered pathway, and this remains to be investigated further. For example, comparing the stability of the rsmA transcript in wild type versus an rsmS mutant might determine whether the rsmA mRNA is targeted for degradation. Conversely, examining RsmA protein levels in an rsmS mutant would determine whether this mutation perturbs the protein’s stability. Finally, as dose-dependency of RsmS is important, it would also be interesting to examine any interaction between RsmS and RsmA in vitro. In summary, this study has conclusively demonstrated a link between RsmS, virulence, and antibiotic production in both Pba and Pcc39048. Given the evidence from other work in E. coli, we think it likely that RsmS/YbaM may be involved in the stress response in many different bacterial species, reflecting a widespread physiological importance.

Speculative model for the function of RsmS in the Rsm regulatory system. (a) Under normal conditions, when rsmB is not present, RsmA, facilitated by RsmS, binds target transcripts, halting or slowing translation. When the untranslated regulatory RNA rsmB is present, it titrates RsmA away, allowing translation and production of virulence factors. (b) In an rsmB mutant, RsmA, facilitated by RsmS, is efficiently bound to any target transcripts, shutting down translation. (c) In an rsmB, rsmS double mutant, RsmA is still able to bind target transcripts, though not as efficiently as previously, leading to a partially restored translation of virulence factors. (d) In an rsmS mutant, RsmA preferentially binds rsmB over target transcripts, increasing translation of target transcripts.
Methods
Bacterial strains and culture conditions
All bacterial strains, plasmids and bacteriophages used in this study are listed in Table 1. Unless otherwise indicated, bacterial strains were grown in Lysogeny broth-Lennox (LB, 10 g l−1 tryptone, 5 g l−1 NaCl, 5 g l−1 yeast extract) or Pel Minimal Broth (PMB, 0.1% (w/v) yeast extract, 0.1% (w/v) (NH4)2SO4, 1 mmol l−1 MgSO4, 0.5% (v/v) glycerol, 0.5% polygalacturonic acid, 40 mmol l−1 K2HPO4, 15 mmol l−1 KH2PO4, pH 7), supplemented with 1.5% agar (LBA) when in plates. E. coli strains were grown at 37 °C, Pcc39048 at 30 °C and Pba at 25 °C. Overnight cultures were grown in 30 ml sealed plastic universals on a roller wheel and aerated growth cultures contained 25 ml of culture in a 250 ml flask, shaken at 215 revolutions per minute (rpm). Optical density (OD600) was monitored using a Unicam Helios spectrophotometer. Where appropriate, antibiotics and supplements were added to media at the following concentrations: ampicillin (Ap), 50 µg ml−1; streptomycin (St), 50 µg ml−1; chloramphenicol (Cm), 50 µg ml−1; kanamycin (Kn), 15 µg ml−1 (E. coli strain β2163) or 50 µg ml−1 (otherwise); and diaminopimelic acid (DAPA), 0.3 mM.
Bacterial strain and plasmid construction
A mutation of rsmS was constructed by allelic exchange50. The upstream and downstream regions of rsmS were amplified by PCR using oligonucleotide pairs oKA33/oKA34 and oKA35/oKA36 respectively (oligonucleotides used in this study are listed in Table 2). A BglII site was artificially introduced within oKA34 and oKA35. The two PCR products were then used to perform overlap PCR generating a construct containing the rsmS flanking regions with the introduced BglII site between them. This construct was digested with BamHI/XhoI and ligated with compatibly digested pBluescript to create pBluescript-ΔrsmS. Oligonucleotides oKA42/oKA43 were then used to amplify the cat gene from pACYC184. This fragment and pBluescript-ΔrsmS were digested with BglII and ligated to create pBluescript-ΔrsmS-Cm. This plasmid was sequenced, digested with BamHI/XhoI, and ligated with compatibly digested pKNG101 to create pKNG101-ΔrsmS-Cm. This construct was introduced into E. coli strain β2163 by transformation and a biparental mating set up with Pba and the E. coli donor cells on LBA + DAPA. The mating patches were incubated at 25 °C overnight and resuspended in one ml of LB. Serial dilutions of the mating patch were plated onto LBA + Cm + St. Double crossover mutants were identified by selection on LBA + Cm containing sucrose50. Colonies were subsequently screened by PCR and sequenced to confirm that the rsmS gene has been removed and replaced with the chloramphenicol acetyltransferase gene.
Plasmid pBAD30-rsmS was created as follows: oKA17 and oKA28 were used to amplify the rsmS ORF from Pba, this product was digested with EcoRI/HindIII and ligated with compatibly digested pBAD30 to create pBAD30-rsmS. The same procedure was used to create pBAD30-priC but with amplification of priC using oligonucleotides oKA89 and oKA91.
Random transposon mutagenesis
Transposon mutagenesis was undertaken using the pDS1028 plasposon system23 or using the TnphoA’2 transposon system34,51. Mutants bypassing rsmB repression of protease production were screened on caseinase agar (Nutrient broth, supplemented with 10 g l−1 Marvel skimmed milk powder and 1.5% agar). The rsmB mutant of Pcc39048 was identified by screening random transposon mutants for reduced protease production on caseinase agar as described previously16. Transposon insertions were transduced into different genetic backgrounds using the generalized transducing phage φM1 for Pba52 or φKP for Pcc3904853. Chromosomal transposon insertion sites were mapped by Random Primed PCR54 or by replicon cloning23.
Phenotypic assays
Plate based assays assessing activities of PCWDEs were carried out by first normalizing the number of cells from an overnight culture for each strain to OD600 1.0. Ten µl of normalised cells were spotted onto indicator plates and incubated at 25 °C for 48 hours. Cellulase and pectate lyase plates were prepared and developed as described previously55 and protease (gelatinase) plates were prepared and developed as described by Hankin and Anagnostaki56. Following development, haloes representing the respective enzyme activity were imaged. Swimming motility was assessed by spotting five µl of a normalized culture (OD600 = 0.1) onto a soft agar plate as described previously9. Carbapenem antibiotic production was monitored using an ESS biosensor seeded top lawn (0.7% agar). A normalized culture (OD600 = 1.0, five µl) of the strain tested was spotted onto the top lawn for live cell based assays, or a cut well assay was used57. For the cut well assay, culture supernatants were sterilized by passage through a 0.22 µm filter and were applied into standard sized wells of an ESS seeded top lawn. After incubation overnight at 30 °C, areas of haloes where ESS growth was inhibited surrounding each well were measured and normalized to OD600.
Liquid PCWDE assays were carried out on culture supernatants grown in PMB. At the indicated timepoint, a one ml sample of the culture was taken, separated by centrifugation at 16 000 g, the supernatant removed, snap frozen in liquid nitrogen and stored at −80 °C until assayed. Pectate lyase activity was determined by measuring the breakdown of polygalacturonic acid as described by Starr and colleagues58. Cellulase and protease activities of samples were determined using the methods described previously by Coulthurst et al.59 and Cubitt et al.16 respectively.
Tuber virulence assays
All tuber virulence assays were performed using Maris Piper potatoes. Tubers were surface sterilized in 1% Virkon™ for five minutes and washed three times in sterile water. Each side of the sterilized tuber was then stabbed with a 200 µl sterile tip to create two inoculation sites. Overnight cultures of either the wild type or rsmS mutant were normalized so that 103 cells were contained within 20 µl and this volume was inoculated into the potato into one of the sites. Then this process was repeated for the other strain at the second inoculation site. Each site of inoculation was sealed using sterile vacuum grease and the potato wrapped three times in damp paper towel. The potatoes were then incubated at 25 °C for four days. The weight of rotting tissue was then measured after removing the rotten area. Four biological replicates were performed and the average weight of rot compared for wild type Pba or an rsmS mutant.
Acyl-homoserine lactone assays
Levels of OHHL were determined using an E. coli biosensor strain JM109 + pSB401 as described previously25,59. Culture supernatants were taken throughout growth and filtered by passing the sample through a 0.22 µm filter. One hundred µl of diluted (1:100) culture supernatants were mixed with 100 µl of JM109 + pSB401 cultures and incubated in black microtiter plates at 37 °C for three hours. Light emission (RLU) was measured using a Lucy Anthos luminometer (Pba) or a FLUOstar Omega plate reader (Pcc39048) and normalized to the original Pba culture OD600.
Bioinformatics and Statistics
Alignments of nucleotide or amino acid sequences were performed using Clustal Omega60. Promoter regions were mapped using BPROM61. All amino acid searches were performed using PSI-BLAST62, conserved domains identified using Pfam9, and genomic locations were examined using existing tools on NCBI. The genome sequence of Pcc39048 used was Genbank accession number QHMC0000000063. Graphpad PRISM was used to perform statistical tests, as indicated in the text, and p-values were reported. Reported values represent the average of a minimum of three independent biological replicates, reported as n = 3.
Data Availability
All data generated or analysed in this study are included in the manuscript.
References
Toth, I. K., Bell, K. S., Holeva, M. C. & Birch, P. R. J. Soft rot erwiniae: from genes to genomes. Mol. Plant Pathol. 4, 17–30 (2003).
Toth, I. K. et al. Mutation in a gene required for lipopolysaccharide and enterobacterial common antigen biosynthesis affects virulence in the plant pathogen Erwinia carotovora subsp. atroseptica. Mol Plant Microbe Interact 12, 499–507 (1999).
Burr, T. et al. Identification of the central quorum sensing regulator of virulence in the enteric phytopathogen, Erwinia carotovora: the VirR repressor. Mol. Microbiol. 59, 113–125 (2006).
Mukherjee, A., Cui, Y., Liu, Y., Dumenyo, C. K. & Chatterjee, A. K. Global regulation in Erwinia species by Erwinia carotovora rsmA, a homologue of Escherichia coli csrA: repression of secondary metabolites, pathogenicity and hypersensitive reaction. Microbiology 142, 427–434 (1996).
Barnard, A. M. et al. Quorum sensing, virulence and secondary metabolite production in plant soft-rotting bacteria. Philos. Trans. R. Soc. London B Biol. Sci. 362 (2007).
Sjoblom, S., Brader, G., Koch, G. & Palva, E. T. Cooperation of two distinct ExpR regulators controls quorum sensing specificity and virulence in the plant pathogen Erwinia carotovora. Mol. Microbiol. 60, 1474–1489 (2006).
Cui, Y., Chatterjee, A., Hasegawa, H. & Chatterjee, A. K. Erwinia carotovora subspecies produce duplicate variants of ExpR, LuxR homologs that activate rsmA transcription but differ in their interactions with N-acylhomoserine lactone signals. J. Bacteriol. 188, 4715–26 (2006).
Valente, R. S., Nadal-Jimenez, P., Carvalho, A. F. P., Vieira, F. J. D. & Xavier, K. B. Signal integration in quorum sensing enables cross-species induction of virulence in Pectobacterium wasabiae. MBio 8, e00398–17 (2017).
Monson, R. et al. Identification of genes in the VirR regulon of Pectobacterium atrosepticum and characterization of their roles in quorum sensing-dependent virulence. Env. Microbiol 15, 687–701 (2013).
Chatterjee, A., Cui, Y., Chakrabarty, P. & Chatterjee, A. K. Regulation of Motility in Erwinia carotovora subsp. carotovora: Quorum-Sensing Signal Controls FlhDC, the Global Regulator of Flagellar and Exoprotein Genes, by Modulating the Production of RsmA, an RNA-Binding Protein. Mol. Plant-Microbe Interact. 23, 1316–1323 (2010).
Romeo, T., Gong, M., Liu, M. Y. & Brun-Zinkernagel, A. M. Identification and molecular characterization of csrA, a pleiotropic gene from Escherichia coli that affects glycogen biosynthesis, gluconeogenesis, cell size, and surface properties. J. Bacteriol. 175, 4744–55 (1993).
Potts, A. H. et al. Global role of the bacterial post-transcriptional regulator CsrA revealed by integrated transcriptomics. Nat. Commun. 8 (2017).
Chatterjee, A., Cui, Y. & Chatterjee, A. K. RsmA and the quorum-sensing signal, N-[3-oxohexanoyl]-L-homoserine lactone, control the levels of rsmB RNA in Erwinia carotovora subsp. carotovora by affecting its stability. J. Bacteriol. 184, 4089–95 (2002).
Ma, W. et al. Molecular characterization of global regulatory RNA species that control pathogenicity factors in Erwinia amylovora and Erwinia herbicola pv. gypsophilae. J. Bacteriol. 183, 1870–80 (2001).
Liu, Y., Cui, Y., Mukherjee, A. & Chatterjee, A. K. Characterization of a novel RNA regulator of Erwinia carotovora ssp. carotovora that controls production of extracellular enzymes and secondary metabolites. Mol. Microbiol. 29, 219–234 (1998).
Cubitt, M. F. et al. A metabolic regulator modulates virulence and quorum sensing signal production in Pectobacterium atrosepticum. Mol Plant Microbe Interact 26, 356–366 (2013).
Cai, X. Y. et al. Methionine synthesis in Escherichia coli: effect of the MetR protein on metE and metH expression. Proc. Natl. Acad. Sci. USA 86, 4407–11 (1989).
Weissbach, H. & Brot, N. Regulation of methionine synthesis in Escherichia coli. Mol. Microbiol. 5, 1593–7 (1991).
Belfaiza, J. et al. Evolution in biosynthetic pathways: two enzymes catalyzing consecutive steps in methionine biosynthesis originate from a common ancestor and possess a similar regulatory region. Proc. Natl. Acad. Sci. USA 83, 867–71 (1986).
Marincs, F., Manfield, I. W., Stead, J. A., McDowall, K. J. & Stockley, P. G. Transcript analysis reveals an extended regulon and the importance of protein-protein co-operativity for the Escherichia coli methionine repressor. Biochem. J. 396, 227–34 (2006).
Leyn, S. A. et al. Comparative Genomics of Transcriptional Regulation of Methionine Metabolism in Proteobacteria. PLoS One 9, e113714 (2014).
Shoeman, R. et al. Regulation of methionine synthesis in Escherichia coli: Effect of metJ gene product and S-adenosylmethionine on the expression of the metF gene. Proc. Natl. Acad. Sci. USA 82, 3601–5 (1985).
Monson, R. et al. A Plasmid-Transposon Hybrid Mutagenesis System Effective in a Broad Range of Enterobacteria. Front. Microbiol. 6, 1442 (2015).
Mulholland, V. et al. A pleiotropic reduced virulence (Rvi-) mutant of Erwinia carotovora subspecies atroseptica is defective in flagella assembly proteins that are conserved in plant and animal bacterial pathogens. Mol Microbiol 9, 343–356 (1993).
Winson, M. K. et al. Construction and analysis of luxCDABE-based plasmid sensors for investigating N-acyl homoserine lactone-mediated quorum sensing. FEMS Microbiol. Lett. 163, 185–92 (1998).
Guzman, L. M., Belin, D., Carson, M. J. & Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 177, 4121–30 (1995).
Esquerré, T. et al. The Csr system regulates genome-wide mRNA stability and transcription and thus gene expression in Escherichia coli. Sci. Rep. 6, 25057 (2016).
Gore, A. L. & Payne, S. M. CsrA and Cra Influence Shigella flexneri Pathogenesis. Infect. Immun. 78, 4674–4682 (2010).
Finn, R. D. et al. The Pfam protein families database: towards a more sustainable future. Nucleic Acids Res. 44, D279–D285 (2015).
Cui, Y. et al. ExpR, a LuxR Homolog of Erwinia carotovora subsp. carotovora, activates transcription of rsmA, which specifies a global regulatory RNA-binding protein. J. Bacteriol. 187, 4792–4803 (2005).
Parker, W. L. et al. SQ 27,860, a simple carbapenem produced by species of Serratia and Erwinia. J. Antibiot. (Tokyo). 35, 653–60 (1982).
Bowden, S. D. et al. Surface swarming motility by Pectobacterium atrosepticum is a latent phenotype that requires O antigen and is regulated by quorum sensing. Microbiology 159, 2375–2385 (2013).
Shih, Y.-L., Harris, S. J., Borner, G., Rivet, M. M. & Salmond, G. P. C. The hexY genes of Erwinia carotovora ssp. carotovora and ssp. atroseptica encode novel proteins that regulate virulence and motility co-ordinately. Environ. Microbiol. 1, 535–547 (1999).
McGowan, S. J. et al. Carbapenem antibiotic biosynthesis in Erwinia carotovora is regulated by physiological and genetic factors modulating the quorum sensing-dependent control pathway. Mol. Microbiol. 55, 526–545 (2004).
McGowan, S. et al. Carbapenem antibiotic production in Erwinia carotovora is regulated by CarR, a homologue of the LuxR transcriptional activator. Microbiology 141, 541–550 (1995).
Hobbs, E. C., Astarita, J. L. & Storz, G. Small RNAs and small proteins involved in resistance to cell envelope stress and acid shock in Escherichia coli: analysis of a bar-coded mutant collection. J. Bacteriol. 192, 59–67 (2010).
Storz, G., Wolf, Y. I. & Ramamurthi, K. S. Small proteins can no longer be ignored. Annu. Rev. Biochem. 83, 753–77 (2014).
Duval, M. & Cossart, P. Small bacterial and phagic proteins: an updated view on a rapidly moving field. Curr. Opin. Microbiol. 39, 81–88 (2017).
Aspden, J. L. et al. Extensive translation of small Open Reading Frames revealed by Poly-Ribo-Seq. Elife 3, e03528 (2014).
Hemm, M. R., Paul, B. J., Schneider, T. D., Storz, G. & Rudd, K. E. Small membrane proteins found by comparative genomics and ribosome binding site models. Mol. Microbiol. 70, 1487–1501 (2008).
Shell, S. S. et al. Leaderless Transcripts and Small Proteins Are Common Features of the Mycobacterial Translational Landscape. PLOS Genet. 11, e1005641 (2015).
Burkholder, W. F., Kurtser, I. & Grossman, A. D. Replication initiation proteins regulate a developmental checkpoint in Bacillus subtilis. Cell 104, 269–79 (2001).
Rowland, S. L. et al. Structure and mechanism of action of Sda, an inhibitor of the histidine kinases that regulate initiation of sporulation in Bacillus subtilis. Mol. Cell 13, 689–701 (2004).
Hobbs, E. C., Yin, X., Paul, B. J., Astarita, J. L. & Storz, G. Conserved small protein associates with the multidrug efflux pump AcrB and differentially affects antibiotic resistance. Proc. Natl. Acad. Sci. 109, 16696–16701 (2012).
Byers, J. T., Lucas, C., Salmond, G. P. C. & Welch, M. Nonenzymatic turnover of an Erwinia carotovora quorum-sensing signaling molecule. J. Bacteriol. 184, 1163–71 (2002).
Bowden, S. D. et al. Virulence in Pectobacterium atrosepticum is regulated by a coincidence circuit involving quorum sensing and the stress alarmone, (p)ppGpp. Mol. Microbiol. 90, 457–471 (2013).
Arunasri, K. et al. Effect of Simulated Microgravity on E. coli K12 MG1655 Growth and Gene Expression. PLoS One 8, e57860 (2013).
Patwa, L. G. et al. Chronic Intestinal Inflammation Induces Stress-Response Genes in Commensal Escherichia coli. Gastroenterology 14, 1842–1851, https://doi.org/10.1053/j.gastro.2011.06.064 (2011).
Rajagopala, S. V. et al. The binary protein-protein interaction landscape of Escherichia coli. Nat. Biotechnol. 32, 285–290 (2014).
Kaniga, K., Delor, I. & Cornelis, G. R. A wide-host-range suicide vector for improving reverse genetics in Gram-negative bacteria: inactivation of the blaA gene of Yersinia enterocolitica. Gene 109, 137–141 (1991).
Wilmes-Riesenberg, M. R. & Wanner, B. L. TnphoA and TnphoA’ elements for making and switching fusions for study of transcription, translation, and cell surface localization. J. Bacteriol. 174, 4558–75 (1992).
Toth, I. K. et al. Generalized transduction in the potato blackleg pathogen Erwinia carotovora subsp. atroseptica by bacteriophage M1. Microbiology 143, 2433–2438 (1997).
Toth, I., Perombelon, M. & Salmond, G. Bacteriophage phiKP mediated generalized transduction in Erwinia carotovora subspecies carotovora. J. Gen. Microbiol. 139, 2705–2709 (1993).
Fineran, P. C., Everson, L., Slater, H. & Salmond, G. P. C. A GntR family transcriptional regulator (PigT) controls gluconate-mediated repression and defines a new, independent pathway for regulation of the tripyrrole antibiotic, prodigiosin, in Serratia. Microbiology 151, 3833–3845 (2005).
Andro, T. et al. Mutants of Erwinia chrysanthemi defective in secretion of pectinase and cellulase. J. Bacteriol. 160, 1199–203 (1984).
Hankin, L. & Anagnostakis, S. L. The Use of Solid Media for Detection of Enzyme Production by Fungi. Mycologia 67, 597 (1975).
Bainton, N. J. et al. N -(3-oxohexanoyl)-l-homoserine lactone regulates carbapenem antibiotic production in Erwinia carotovora. Biochem. J., https://doi.org/10.1042/bj2880997 (1992).
Starr, M. P., Chatterjee, A. K., Starr, P. B. & Buchanan, G. E. Enzymatic degradation of polygalacturonic acid by Yersinia and Klebsiella species in relation to clinical laboratory procedures. J. Clin. Microbiol. 6, 379–86 (1977).
Coulthurst, S. J. et al. DsbA plays a critical and multifaceted role in the production of secreted virulence factors by the phytopathogen Erwinia carotovora subsp. atroseptica. J Biol Chem 283, 23739–23753 (2008).
Sievers, F. et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7, 539 (2011).
Solovyev, V. & Salamov, A Automatic annotation of microbial genomes and metagenomic sequences. In In: Li. R.W. (ed.), Metagenomics and its applications in agriculture, biomedicine and environmental studies Nova Science Publishers. 61–78 (2011).
Altschul, S. F. et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25, 3389–3402 (1997).
Monson, R. E., Jones, K. & Salmond, G. P. C. Draft Genome Sequence of Pectobacterium carotovorum subsp. carotovorum ATCC 39048, a Carbapenem-Producing Phytopathogen. Microbiol. Resour. Announc. 7, e00825–18 (2018).
Bell, K. S. et al. Genome sequence of the enterobacterial phytopathogen Erwinia carotovora subsp. atroseptica and characterization of virulence factors. Proc. Natl. Acad. Sci. USA 101, 11105–10 (2004).
Demarre, G. et al. A new family of mobilizable suicide plasmids based on broad host range R388 plasmid (IncW) and RP4 plasmid (IncPα) conjugative machineries and their cognate Escherichia coli host strains. Res. Microbiol. 156, 245–255 (2005).
McClean, K. H. et al. Quorum sensing and Chromobacterium violaceum: exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 143(Pt 1), 3703–11 (1997).
Acknowledgements
The authors acknowledge the technical help of Sarah Barker, Alison Rawlinson and Diana Breitmaier. Maria Stroyakovski and Amy Hill provided comments on the manuscript and Jessica Bergman provided helpful discussions throughout the writing process. NS was supported by a postgraduate research studentship from the MRC, UK. KA, SB, and MC were supported by postgraduate research studentships from the BBSRC, UK and the GPCS lab was in receipt of generous grant support from the BBSRC, UK (awards BB/N008081/1, BB/H013261/1 and BB/F009666/1) and from the James Hutton Institute, Scotland.
Author information
Authors and Affiliations
Contributions
All authors helped to design the experiments and performed critical data analysis. R.E.M., K.A., N.R.W. and M.F.C. performed the experiments in Pba. R.E.M., K.A., S.B., N.S. and S.H. performed the experiments in Pcc39048. R.E.M., S.B., N.S. and S.H. performed the bioinformatic analysis. R.E.M. wrote the manuscript. R.E.M., G.P.C.S. and M.F.C. edited the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Monson, R.E., Apagyi, K., Bowden, S.D. et al. The rsmS (ybaM) mutation causes bypass suppression of the RsmAB post-transcriptional virulence regulation system in enterobacterial phytopathogens. Sci Rep 9, 4525 (2019). https://doi.org/10.1038/s41598-019-40970-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-40970-3
- Springer Nature Limited