
Abstract
The potential hepatoprotective activities of two Lannea species were explored in vivo. Furthermore, the binding activities of their main polyphenols to the antiapoptotic protein Bcl-2 were investigated. Based on HPLC-MS/MS results, 22 secondary metabolites were characterized in L. stuhlmannii (mainly tannins), while 20 secondary metabolites (mainly sulphated tannins) were identified in L. humilis. Both extracts exhibited substantial antioxidant activities in vitro and counteracted D-galactosamine induced intoxication in rats in vivo and increased the total antioxidant capacity (TAC) of liver tissues. In addition to reducing the elevated levels of AST and total bilirubin, both extracts significantly attenuated the deleterious histopathologic changes in liver after D-galactosamine-intoxication. Also, both extracts protected hepatocytes from apoptotic cell death and increased the expression of the anti-apoptotic protein Bcl-2. The identified compounds from both extracts can bind to the Bcl-2: Bim (BH3) interface with an appreciable binding free energy. Hydrogen and ionic bonds and hydrophobic interactions with amino acid residues in the hydrophobic face of Bim (BH3) domain were discovered. To sum up, L. humilis and L. stuhlmanni exhibited promising hepatoprotective activities in vivo against D-GalN-induced liver injury and their hepatoprotection is due to the antioxidant and anti-apoptotic effects of tannins and proanthocyanidins.
Similar content being viewed by others


Introduction
Liver cirrhosis is considered a major health problem and a leading cause of death all over the world. This pathological condition is directly or indirectly connected to an overproduction of free radicals. Drug candidates able to combat free radicals and their negative impact on cells could serve as leads for treatment. Several studies have demonstrated that many plants produce secondary metabolites with antioxidant and proven powerful free radical scavenging activities. Such plants might have a potential for the treatment of liver diseases linked to oxidative stress and related conditions1,2.
The genus Lannea with 40 species is one of 69 genera (about 850 species) belonging to the Anacardiaceae. This genus, along with others such as Pistacia, Blepharocarya, Campylopetalum, and Dobinea, is widespread in the tropics3. Phytochemical investigations of several members of this genus revealed several new and rare natural products distributed over different phytochemical classes such as tannins, cyanidins, flavonoids, alkylphenols, and dihydroalkyhexenones4,5. For instance, a prenylated chromenflavanone was isolated from L. acida leaf extract6. Also, three dihydroalkylhexenones were discovered in L. edulis in a bio-guided fractionation5.
Several biological activities have been reported from the genus Lannea. In particular, a L. coromandelica leaf extract has shown anti-conceptive activities while its bark extract was found to have hypoglycemic activities, in addition to anti-inflammatory activities in rats7,8. Okoth et al. have reported antioxidant and antibacterial activities for L. alata root and the isolated compounds myricitrin, myricetin-3-O-a-arabinofuranoside, lupeol, and sitosterol4.
Lannea stuhlmannii and L. humilis are deciduous trees widely distributed in the tropics3. In Tanzania, Lannea species are being used traditionally for the treatment of cancer. Also, decoctions of L. stuhlmannii root are still in use against fever, asthma, stomachache, and dysentery while decoctions of the stembark are considered beneficial to counteract headache and stomach pain. The leaf extract is used to relieve abdominal pain and to hasten childbirth. Moreover, stembark extract has shown antibacterial activity9. In Namibia, L. stuhlmannii roots are used as ethnomedicine for the treatment of HIV/AIDS infections. In East Africa, the plant is employed traditionally in treatment of anaemia10.
The bark of L. humilis is used traditionally to treat fever, and diarrhea11. In Tanzania, a root decoction is drunk to treat anaemia, stomach pains, nausea and general body weakness12. L. humilis trees excrete a water-soluble gum rich in amino acids, galactose, and other polysaccharides13,14. The root of L. stuhlmannii and the bark from L. humilis demonstrated promising anti-trypanosomal and cytotoxic activities in a panel of 40 plants extracts15.
Phytochemical studies are missing for both species. However, a preliminary study by Chhabra et al. reported the presence of some alkaloids and flavonoids in L. stuhlmannii bark16. Herein we characterized the polyphenolic constituents of L. stuhlmannii and L. humilis bark extracts using HPLC-PDA-ESI-MS/MS. Moreover, in vitro antioxidant activities were studied and hepatoprotective activities were investigated in rats treated with the toxic D-galactosamine that causes liver damage. Molecular modelling was employed to find out if the main polyphenols of the plants bind to Bcl-2: Bim (BH3) interface that might be modulated by the drug.
Materials and Methods
Plant material
Lannea humilis (Oliv.) Engl. and Lannea stuhlmannii (Engl.) Engl. bark samples were collected from trees growing in Lupaga Site in Shinyanga, Tanzania and kept under accession number P7311 and P7317, respectively, at IPMB, Heidelberg University15. The bark samples were ground and extracted with 100% methanol at room temperature for three days (6 × 500 mL). The combined extracts were filtered and evaporated using a rotary evaporator at 40 °C. The residues were frozen at −70 °C, and then lyophilized to fine dried powder. The extraction yield was calculated based on initial dry weight and found to be 22% and 15% for L. humilis and L. stuhlmannii bark, respectively.
HPLC-PDA-MS/MS
The extracts were analyzed utilizing a ThermoFinnigan LCQ-Duo ion trap mass spectrometer (ThermoElectron Corporation, Waltham, Ma, USA) with an ESI source (ThermoQuest Corporation, Austin, Tx, USA). A ThermoFinnigan HPLC system using a C18 reversed-phase column (Zorbax Eclipse XDB-C18, Rapid resolution, 4.6 × 150 mm, 3.5 µm, Agilent, Santa Clara, CA, USA) was used17. Water and acetonitrile (ACN) (Sigma-Aldrich GmbH, Germany) (0.1% formic acid each) were used as a mobile phase. At 0 min, ACN was 5% and then increased to 30% in 60 min at 1 mL/min with a 1:1 split before the ESI source. Autosampler surveyor ThermoQuest was utilized to inject the extracts and the system was controlled by Xcalibur software (XcaliburTM 2.0.7, Thermo Fischer Scientific, Waltham, Ma, USA). The MS operated in the negative mode as reported before18. The ions were detected in a full scan mode and mass range of 50–2000 m/z.
Biological experiments
Antioxidant activities in vitro
The Folin-Ciocalteu method quantified the total phenolic content at the extracts as described before19. The 2,2′-diphenyl-1-picryl-hydrazyl-hydrate (DPPH) radical scavenging and ferric reducing antioxidant power (FRAP) assays were utilized to evaluate the antioxidant properties of the extracts and were carried out as described earlier19.
Animals and experimental design
Male Wistar rats (8 weeks of age, Faculty of Veterinary Medicine, Zagazig, Egypt) were used. Rats were housed in a light-controlled room with a 12-h light/dark cycle and had access to food and water. Experimental protocols and animal care methods were approved by Ethical Committee of the Faculty of Pharmacy, Zagazig University for Animal Use (Zagazig, Egypt) and conducted following the guidelines of the US National Institutes of Health on animal care and use. After a 1-week acclimatization period, rats were divided into seven groups: (1) Control, (2) D-galactosamine (D-GalN) treated with vehicle (0.9% saline), (3) Lannea humilis (100 mg/kg, oral), (4) L. humilis (200 mg/kg, oral), (5) Lannea stuhlmannii (100 mg/kg, oral), (6) L. stuhlmannii (200 mg/kg, oral) and (7) silymarin (100 mg/kg, oral). Six animals were allocated to each group (total animals: n = 42). To induce acute liver failure, the rats were injected with D-GalN (800 mg/kg, i.p). The animals were treated with vehicle, extracts or silymarin for three consecutive days before the injection of D-GalN. Blood samples were drawn from the retro-orbital plexus of the eye 24 h after D-GalN injection under pentobarbital anesthesia (50 mg/kg, i.p.). The serum was separated by centrifugation at 3500 × g for 15 min at 4 °C. These serum samples were used for measurement of AST activity and total bilirubin level. The livers were excised and used for light microscopic observations, oxidative stress markers measurement and immunohistochemical staining of Bcl-2.
Liver proteins
Serum aspartate aminotransferase (AST) activity was measured using a commercial assay kit, (Diamond diagnostic, Egypt). Total bilirubin levels were measured using commercially available analytical kit (BioMed Diagnostics, Egypt).
Determination of oxidative stress markers
The generation of reactive oxygen species following D-GalN injection was detected in liver tissues using commercially available kits provided by Biodiagnostic (Giza, Egypt) to measure the lipid peroxidation product content, malondialdehyde (MDA).Measurement of Total antioxidant capacity (TAC) was performed by reacting with the endogenous antioxidants in the serum sample with certain amount of hydrogen peroxide (H2O2). The remaining H2O2 was measured by an enzymatic reaction with 3,5-dichloro-2-hydroxy benzene sulphonate producing a colored compound. The intensity of the resulting color was measured by spectrophotometer (JENWAY 6105 UV/V Spectrophotometer) at wavelength of 505 nm.
Histology
Excised livers were processed for light microscopic examination, according to standard procedures. The livers were preserved in phosphate-buffered 10% formalin, after which the livers were cut into small pieces, embedded in paraffin wax, sectioned at 4 µm thickness, and stained with haematoxylin and eosin.
Immunohistochemical determination of Bcl-2 (anti-apoptotic marker)
Liver sections were prepared by soaking them in xylene to remove paraffin and graded alcohol was used (70%, 90%, 100%). The activity of endogenous peroxidase was blocked by addition of 5% H2O2 in absolute methanol for 10 min at room temperature. Sections were then rinsed by phosphate buffered saline (PBS). Thereafter, sections were incubated with primary antibodies against Bcl-2. Protein expression was measured using Streptavidin biotin peroxidase kit. Tissues were stained with diaminobenzidine (DAB) as chromogen for Bcl-2 detection and then counterstained with haematoxylin20.
Morphometric analysis
The number of Bcl-2 positive hepatocytes in five randomly selected high power microscopic fields were analysed using computerized image system (a Leica Qwin 500 image analyser connected to a Leica microscope). This number was expressed as cell number per µm².
Statistical analysis
Values are expressed as the mean ± standard error of the mean. Data were processed by Graphpad prism version 5 (GraphPad Software, San Diego, CA, U.S.A.). For statistical analysis, we used unpaired Student’s t-tests for two-group comparisons and one-way analysis of variance (ANOVA) followed by Tukey post hoc tests for multiple comparisons. For all comparisons, differences were considered significant at p < 0.05.
Results and Discussion
The methanol extracts obtained from the bark of L. stuhlmannii and L. humilis were analyzed by HPLC-PDA-ESI-MS/MS. A total of 22 and 20 secondary metabolites were tentatively identified in L. stuhlmannii and L. humilis bark, respectively. L. stuhlmannii extract is rich in condensed tannins, whereas sulphated condensed tannins and phenolic acids dominated in L. humllis (Table 1 and Fig. 1).

Total ion chromatogram of methanol bark extracts of (a) L. stuhlmannii (22 compounds, mainly tannins). (b) L. humilis (20 secondary metabolites, mainly sulphated tannins).
In the L. humilis extract, compound 27, (λmax = 278), demonstrated a [M – H]− at m/z 399 and main fragments at m/z 319 [M – H – 80, sulphate moiety], m/z 301[M – H – 80 – 18, sulphate and water moieties], and m/z 193 [M – H - C2H4 - C6H5O3]; it was tentatively characterized as (epi)gallocatechin 5-O-methyl 7-O-sulphate. Representative spectra and proposed fragmentation pattern are shown in Fig. 2(a,b). Also, compound 40 showed a [M – H]− at m/z 551 and main daughter ions at m/z 471 [M – H – 80, sulphate moiety], 319 [M – H – 80–152, loss of sulphate and gallate moieties], 301, and 193, it was tentatively assigned to (epi)gallocatechin gallate 5-O-methyl 7-O- sulphate, (Fig. 3a,b).

(a) MS2 of compound (27), (epi)gallocatechin 5-O-methyl 7-O-sulphate at [M – H]− at m/z 399. (b) A proposed fragmentation pattern.

(a) MS2 of compound (40), (epi)gallocatechin gallate 5-O-methyl 7 sulphate at [M – H]− at m/z 551. (b) A proposed fragmentation pattern.
Compound 30 exhibited a [M – H]−at m/z 383 and a main daughter ion at m/z 303 [M – H – 80, sulphate moiety]; it was tentatively identified as flavan 3-,4-,5-trihydroxy 5-O methyl 7 sulphate, (Fig. 4a,b). Compound 31 exhibited a [M – H]− at m/z 277 and a molecular ion peak at m/z 197 [M – H – 80]; it was assigned to syringic acid sulphate.

(a) MS2 of compound (30), flavan 3-,4-,5-trihydroxy 5-O methyl 7-O-sulphate at [M – H]− at m/z 383. (b) A proposed fragmentation pattern.
The peak at retention time 7.51 min (compound 34) showed a [M – H]− at m/z 397 and a main fragment ion at m/z 317. The latter exhibited a main fragment ion at m/z 289 in MS3. From the fragmentation pattern, we were not able to confirm the position of the ethyl group substituent, position 5 or position 8 on the flavan nucleus. After comparing its UV spectra with those of compound 27; it was identified as (epi)catechin-5-O-ethyl 7-O-sulphate. According to Woodward’s rules, the compound would have had a higher λmax than the current reported value if the ethyl group had been in position 8. A representative mass spectra and proposed fragmentation pattern are shown in Figs 5 and 6.

Negative ion ESI-MS/MS spectra of (epi)catechin-5-O-ethyl, 7-O-sulphate (a) MS2 of [M − H]−m/z 397. (b) MS3 of main daughter ion at m/z 317.

A proposed fragmentation pattern of (epi)catechin-5-O-ethyl 7-O-sulphate at [M − H]−m/z 397.
Peak 33 (retention time 6.87 min) has demonstrated a [M – H]− at m/z 559 and fragment ions at m/z 479 [M – H – 80, sulphate moiety], 397 [M – H – 162, hexose moiety], and 317 [M – H – 80–162] was tentatively assigned to (epi)catechin-5-O-ethyl 7-O-sulphate 3-O-hexoside, Fig. 7.

Negative ion ESI-MS/MS spectra of (epi)catechin-5-O-ethyl 7-O-sulphate 3-O-hexoside at [M − H]−m/z 559.
Both (epi)gallocatechin gallate-(epi)catechin (35) and (epi)catechin gallate-(epi)catechin gallate (41) were detected only in L. humilis at [M – H]− m/z 745, and 881, respectively, while caffeoylquinic acid (3) and feruloylquinic acid (4) were only detected in L. stuhlmannii at [M – H]− m/z 353, and 367 with a molecular ion peak at m/z 191, respectively22. Both extracts were found to contain (epi)catechin monomers, dimers and trimers17.
Biological activities
Antioxidant and hepatoprotective activities
Catechins are renowned for their powerful potential to scavenge various free radicals such as hydroxyl, peroxyl, superoxide, and other radicals. Antioxidant activity of catechins is mediated through different mechanisms. They are able to transfer an electron to bind a reactive radical, while they change to the more stable and less reactive phenoxyl radical. They are also able to chelate Cu2+ and Fe3+ ions, thus limiting free radicals generation. Indirectly, catechins exert an antioxidant effect via increasing the level of endogenous antioxidants such as glutathione reductase, catalase, and superoxide dismutase. Moreover, catechins are reported to have an inhibitory effect on xanthine oxidase that catalyzes the metabolism of purines into uric acid and reactive oxygen species. A direct relationship between the number of the hydroxyl groups a catechin possess and the exerted antioxidant potential has been demonstrated17,18.
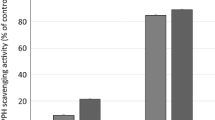
Here, the antioxidant potential of the methanol extract of bark from L. humilis and L. stuhlmannii was investigated using two assays, namely DPPH and FRAP. The total phenolic content (TPC) was determined using Folin Ciocalteu assay and expressed as mg gallic acid (GAE)/g extract. Both extracts showed substantial activities in all assays, data are shown in Table 2. Such promising scavenging activities were reported from other Lannea species such as L. alata4, other bark-tannins rich extracts from Albizia harveyi, Senna singueana, Turraea fischeri, and proanthocyanidin rich root extracts from Cassia abbreviata and Ximenia americana var. caffra17,18,21,23,24.
In another set of experiments, the in vivo antioxidant potential was examined in rats treated with toxic D-galactosamine. We determined the total antioxidant capacity (TAC) and lipid peroxidation product, MDA in liver tissues as markers for oxidative stress. Except for the low dose level of L. stuhlmannii, both extracts were able to counteract the D-galactosamine-intoxication and increased the TAC of liver tissues (Fig. 8A). However, the low dose levels of L. humilis and L. stuhlmannii (100 mg/kg b.w.) were able to attenuate the MDA increase (Fig. 8B).

Antioxidant activity in vivo of oral administration of L. humilis (LH) and L. stuhlmannii (LS) in a dose of 100 mg/kg and 200 mg/kg and silymarin in a dose of 100 mg/kg for 3 days against the oxidative stress caused by D-galactosamine (D-GalN, 800 mg/kg, ip). (A) Total antioxidant capacity (TAC) in control and treated rats. (B) Liver malondialdehyde (MDA) contents in control and treated rats. Each bar represents the mean ± standard error (n = 5). *p < 0.05 versus control. @p < 0.05 versus vehicle-treated D-GalN group.
Extract effects on markers of liver dysfunction and histopathological changes
To test whether the two studied Lannea species have hepatoprotective activities in vivo, the biochemical and histopathological changes were evaluated after a liver cell injury had been induced by single dose injection of D-galactosamine (D-GalN, 800 mg/kg, i.p.). All treatments reduced AST and total bilirubin levels (p < 0.05), except the high dose of L. humilis which failed to decrease serum AST activity (p > 0.05), Fig. 9.

Hepatoprotective action of oral administration of L. humilis (LH) and L. stuhlmannii (LS) in a dose of 100 mg/kg and 200 mg/kg and silymarin in a dose of 100 mg/kg for 3 days in rats after D-galactosamine (D-GalN, 800 mg/kg, ip) induced acute liver injury. (A) Serum enzyme profile of AST in control and treated rats. (B) Serum total bilirubin level in control and treated rats. Each bar represents the mean ± standard error (n = 5). *p < 0.05 versus control. @p < 0.05 versus vehicle-treated D-GalN group.
To confirm the in vivo antioxidant activities and the observed biochemical changes, a histopathological evaluation was performed. It is obvious from Fig. 10B that D-GalN has induced a mononuclear cellular infiltration indicating the development of inflammation in liver tissues (arrow head) in the portal area and congested blood vessel (BV). Together with the observed hyperplasia of bile ducts (arrow), these changes provide evidence for the successful establishment of animal liver injury model. In general, these deleterious changes were significantly attenuated by L. humilis and L. stuhlmannii treatment. However, a dose of 100 mg/kg L. humilis has shown little improvement and some mononuclear cellular infiltration (arrow) was spotted in the portal tract area and additionally a congested blood vessel (BV) was detected (Fig. 10C).

Light photomicrographs of the liver of rats treated with vehicle (0.9% saline), 200×. (A) Normal control. (B) The heptotoxic D-GaIN (800 mg/kg b.w.) (C,D) L. humilis (100 and 200 mg/kg, b.w.), (E,F) L. stuhlmannii (100 and 200 mg/kg, b.w.) (G) The hepatoprotective control silymarin (100 mg/kg, b.w.).
A higher dose of L. humilis (200 mg/kg) showed marked improvement indicated by little mononuclear cellular infiltration (arrow) and central vein (CV) with minimum congestion, (Fig. 10D). The low dose level (100 mg/kg) of L. stuhlmannii had little improvement as seen in Fig. 10E, where mononuclear cellular infiltration (arrow head) was clearly noticed in the portal area accompanied with bile duct hyperplasia (arrow). Figure 10F revealed that the high dose level (200 mg/kg) of L. stuhlmannii was able to exert some improvements where mononuclear cellular infiltration was observed only in the portal tract area (arrow head) and less congested blood vessel (BV) was also observed. Figure 10G showed that silymarin pretreatment (a lignin with known hepatoprotective properties) caused partial improvement.
Extract effects on anti-apoptotic marker Bcl-2
When cellular antioxidants are depleted and ROS accumulation reaches a critical threshold, mitochondrial damage occurs. This process leads to the release of cytochrome c (Cyt-c) from the mitochondria, which in turn activates the apoptotic pathway25. Several proteins can inhibit or decrease apoptosis, among them the anti-apoptotic members of Bcl-2 family proteins26. To clarify the mechanism that the extracts attenuated hepatotoxicity induced by D-GalN, we stained liver sections with avidin-biotin peroxidase stain with haematoxylin counter stain for Bcl-2, anti-apoptotic marker, and determined the percentage of positively stained hepatic cells. A significant increase in the number of Bcl-2 positive hepatocytes was observed in rats treated with all dose levels of the extracts (Fig. 11C–F), suggesting that both Lannea species protected hepatocyte from apoptotic cell death. The extracts produced similar effect to silymarin (Fig. 11G).

Immunohistochemical determination of Bcl-2 in paraffin-embedded liver tissues stained with Avidin-biotin peroxidase stain with haematoxylin counter stain, (A) Liver of rats treated with vehicle (0.9% saline). (B) Rats injected a single dose of D-GaIN (800 mg/kg, b.w.); (C and D) rats treated with two doses of L. humilis for 3 days (100 and 200 mg/kg, b.w.). (E and F) Rats treated with two doses of L. stuhlmannii for 3 days (100 and 200 mg/kg, b.w.). (G) The positive control silymarin (100 mg/kg, b.w.), magnification, 400×. The Bar graph represents mean ± standard error of the mean of the number of Bcl-2 positive hepatocytes (n = 6). *p < 0.05 versus control, @p < 0.05 versus vehicle-treated D-GalN group.
Molecular modelling study
The family of B-cell lymphoma-2 (Bcl-2) proteins is known for regulating programmed cell death through mediating mitochondrial or intrinsic initiated apoptosis. The Bcl-2 proteins are either pro-survival (anti-apoptotic) such as Bcl-XL, Bcl-2, Bcl-B, and Mcl-1 proteins or anti-survival (pro-apoptotic) proteins including Bim, Bad, Bak, Bax, and others27.
The molecular structures of the majority of these proteins have been resolved. The anti-apoptotic members have been found to share one or even more of the crucial BH domains namely BH1, BH2, BH3, and BH4. All the pro-apoptotic proteins, however, contain the BH3 domain that is very essential for cell killing activity28.
In order to get an insight how the plant extracts were able to protect against hepatic cells death, we decided to dock some of the identified compounds to the crystal structure of the interaction surface of Bcl-2:Bim (BH3) complex, pdb code: 4b4s, using molecular operating environment (MOE), 2013.08; Chemical Computing Group Inc., Montreal, QC, Canada, H3A 2R7, 2016 according to our previously applied protocol24. The binding site in the Bcl-2: Bim (BH3) crystal structure as well as the amino acid residues crucial for the efficient binding between the two proteins are well established, thus docking the plant major chemical components to this site would be beneficial to understand how they are able to hinder the Bcl-2: Bim complex formation and thus blocking apoptosis. The docked compounds are shown in Fig. 12 and docking results are summarized in Table 3.

Compounds docked to the Bcl-2: Bim (BH3) interface. Compounds are numbered according to Table 1.
Despite the fact that the actual mechanism by which these proteins regulate apoptosis is still not clearly understood, it is well accepted now that this happens through heterodimerization of two members of pro- and anti-apoptotic agents into a protein complex that activates apoptotic pathways29. Resolving the loop residues in the Bcl-2:Bim (BH3) complex for instance, has shown that Bcl-2 surface has a hydrophobic cleft in which the hydrophobic face of the BH3 of Bim binds. In this regard, Bim (BH3) domain was shown to extend from Arg 53 to Ala 74 amino acid residues. Within this domain, the side chains of five hydrophobic residues are interacting with the hydrophobic cleft of Bcl-2 and conserved in all pro-apoptotic BH3 domains, namely Ile 58, Leu 62, Ile 65, Phe 69, and Tyr 7330. Additionally, some more amino acid residues were later reported to contribute favorably to the binding free energy of the complex such as Arg 64 and Glu 68. The residue Tyr 72 contributes to a lesser extent to the complex formation as its side chain is more solvent exposed31. Studies have shown that a single mutation in any of these residues such as Leu 62, for instance, could lead to a severe impairment in the binding of the two opposing proteins and hence blocking the apoptotic cell death32,33.
As shown in Table 3, the docked compounds were able to bind to the Bcl-2: Bim (BH3) interface with a scoring function of a range between −11.98 to −15.94 reflecting an appreciable binding free energy. Additionally, the compounds have afforded different polar and hydrophobic interactions to the amino acid residues in the hydrophobic face of Bim BH3 domain, (Table 3). Except for 6c, all other compounds with S-configuration of the stereo-center(s) were able to interact with the essential conserved Tyr 73 and/or Tyr 72 residues. However, compounds 27b and 40b having stereo-centers with R- configuration have also shown to interact with Tyr 73 residue. This interaction stabilized by different other polar and hydrophobic interactions could result in conformational changes in the protein structure leading to interference with dimerization of the two opposing proteins Bcl-2: Bim, thus hindering apoptosis. Figures 13 and 14 show the 3D poses and the 2D-interactions, respectively, of two selected compounds34a, 40a, and their enantiomers 34b, and 40b docked to the Bcl-2: Bim (BH3) interface.

3D-docking poses of compounds 34a (top left), 34b (top right), 40a (bottom left), and 40b (bottom right) docked to Bcl-2: Bim (BH3) interface. Surface residues are coloured (grey hydrophobic, red acidic, blue basic, and cyan other).

2D-interactions of compounds 34a (top left), 34b (top right), 40a (bottom left), and 40b (bottom right) with amino acid residues on Bcl-2: Bim (BH3) interface.
In view of the immunohistochemical and molecular modeling studies, the anti-apoptotic effect of the extracts could be presumably attributed to their tannin content. This comes to agreement with other studies that have reported anti-apoptotic activity for proanthocyanidin rich extracts such as the bark extract of Senna sengueana21. Polyphenols are powerful biologically active natural products which can bind to several important proteins in cells by forming multiple hydrogen and ionic bonds that can modulate the 3D structure of proteins34. In summary, evidence is provided that L. humilis and L. stuhlmanni protect against D-GalN-induced liver injury. The hepatoprotection of the extracts is due to antioxidant and anti-apoptotic effects.
Conclusions
The chemical profiling of the methanol extracts of Lannea stuhlmannii and L. humilis bark was analyzed by HPLC-PDA-ESI-MS/MS. A total of 42 secondary metabolites were identified in the investigated extracts. Noteworthy, this work provides the first comprehensive phytochemical study of L. stuhlmannii and L. humilis bark and confirms their traditional use. The extracts could protect rat liver from intoxication with D galactosamine, apparently through antioxidant and increasing the antiapoptotic protein Bcl-2. Thus, Lannea is another candidate for the potential treatment of liver injury that needs further and deeper analyses.
References
van Wyk, B.-E. & Wink, M. Phytomedicines, Herbal Drugs, and Poisons. University of Chicago Press. 2015.
Van Wyk, B.-E. & Wink, M. Medicinal plants of the world. (2nd Ed.) 2017.
Mabberley, D. J. Mabberley’s plant-book: a portable dictionary of the vascular plants. Cambridge University Press, Cambridge, 2008.
Okoth, D. A. et al. Antibacterial and antioxidant activities of flavonoids from Lannea alata (Engl.) Engl. (Anacardiaceae). Phytochem Lett 6, 476–481, https://doi.org/10.1016/j.phytol.2013.06.003 (2013).
Queiroz, E. F. et al. New dihydroalkylhexenones from Lannea edulis. J Nat Prod 66, 578–580, https://doi.org/10.1021/np0202428 (2003).
Sultana, S. & Ilyas, M. A flavanone from Lannea acida. Phytochemistry 25, 963–964, https://doi.org/10.1016/0031-9422(86)80040-1 (1986).
Imam, M. Z. & Moniruzzaman, M. Antinociceptive effect of ethanol extract of leaves of Lannea coromandelica. J Ethnopharmacol 154, 109–115, https://doi.org/10.1016/j.jep.2014.03.032 (2014).
Singh, S. & Singh, G. B. Anti-inflammatory activity of Lannea coromandelica bark extract in rats. Phytother Res 8, 311–313, https://doi.org/10.1002/ptr.2650080513 (1994).
Chhabra, S. C. et al. Plants used in traditional medicine in eastern Tanzania. I. Pteridophytes and angiosperms (Acanthaceae to Canellaceae). J Ethnopharmac 21, 253–277, https://doi.org/10.1016/0378-8741(87)90103-6 (1987).
Chinsembu, K. C. & Hedimbi, M. An ethnobotanical survey of plants used to manage HIV/AIDS opportunistic infections in Katima Mulilo, Caprivi region, Namibia. JEthnobiol Ethnomed 6, 25, https://doi.org/10.1186/1746-4269-6-25 (2010).
Kudi, A. C. & Myint, S. H. Antiviral activity of some Nigerian medicinal plant extracts. J Ethnopharmacol 68, 289–294, https://doi.org/10.1016/S0378-8741(99)00049-5 (1999).
Maregesi, S. M. et al. Ethnopharmacological survey of the Bunda district, Tanzania: Plants used to treat infectious diseases. J Ethnopharmacol 113, 457–470, https://doi.org/10.1016/j.jep.2007.07.006 (2007).
Anderson, D. M. W. & Hendrie, A. The structure of Lannea humilis gum. Carbohyd Res 22, 265–279, https://doi.org/10.1016/S0008-6215(00)81277-X (1972).
Anderson, D. M. W. et al. The amino acid and amino sugar composition of some plant gums. Phytochemistry 11, 733–736, https://doi.org/10.1016/0031-9422(72)80039-6 (1972).
Nibret, E. et al. Screening of some Tanzanian medicinal plants for their trypanocidal and cytotoxic activities. Phytother Res 24(7), 945–947, https://doi.org/10.1002/ptr.3066 (2010).
Chhabra, S. C. et al. Phytochemical screening of tanzanian medicinal plants. I. J Ethnopharmacol 11, 157–179, https://doi.org/10.1016/0378-8741(84)90037-0 (1984).
Sobeh, M. et al. Polyphenolics from Albizia harveyi exhibit antioxidant activities and counteract oxidative damage and ultra-structural changes of cryopreserved bull semen. Molecules 22, 1993, https://doi.org/10.3390/molecules22111993 (2017).
Sobeh, M. et al. Hepatoprotective and hypoglycemic effects of a tannin rich extract from Ximenia americana var. caffra root. Phytomedicine 33, 36–42, https://doi.org/10.1016/j.phymed.2017.07.003 (2017).
Ghareeb, M.A. et al. HPLC-DAD-ESI-MS/MS analysis of fruits from Firmiana simplex (L.) and evaluation of their antioxidant and antigenotoxic properties. J Pharm Pharmacol, https://doi.org/10.1111/jphp.12843 (2017).
Zhou, X. et al. Effects of SSTF on the expression of apoptosis-associated gene Bcl-2 and Bax by cardiomyocytes induced by H2O2. Guangdong Med J28, 1590–1591 (2005).
Sobeh, M. et al. Senna singueana: Antioxidant, hepatoprotective, antiapoptotic properties and phytochemical profiling of a methanol bark extract. Molecules 22, 1502, https://doi.org/10.3390/molecules22091502 (2017).
Sobeh, M. et al. HPLC-PDA-MS/MS characterization of bioactive secondary metabolites from Turraea fischeri bark extract and its antioxidant and hepatoprotective activities in vivo. Molecules 22, 2089, https://doi.org/10.3390/molecules22122089 (2017).
Sobeh, M. et al. A proanthocyanidin-rich extract from Cassia abbreviata exhibits antioxidant and hepatoprotective activities in vivo. J Ethnopharmacol 213, 38–47, https://doi.org/10.1016/j.jep.2017.11.007 (2018).
Sobeh, M. et al. Albizia harveyi: phytochemical profiling, antioxidant, antidiabetic and hepatoprotective activities of the bark extract. Med Chem Res 26, 3091–3105, https://doi.org/10.1007/s00044-017-2005-8 (2017).
RomanRamos, R. et al. Monosodium glutamate neonatal intoxication associated with obesity in adult stage is characterized by chronic inflammation and increased mrna expression of peroxisome proliferatoractivated receptors in mice. Basic Clin Pharmacol Toxicol 108, 406–413, https://doi.org/10.1111/j.1742-7843.2011.00671.x (2011).
Billard, C. Design of novel BH3 mimetics for the treatment of chronic lymphocytic leukemia. Leukemia 26, 2032–2038, https://doi.org/10.1038/leu.2012.88 (2012).
Youle, R. J. & Strasser, A. The BCL-2 protein family: opposing activities that mediate cell death. Nature reviews. Molecul Cell Biol 9, 47, https://doi.org/10.1038/nrm2308 (2008).
Liu, X. et al. The structure of a Bcl-xL/Bim fragment complex: implications for Bim function. Immunity 19, 341–352, https://doi.org/10.1016/S1074-7613(03)00234-6 (2003).
Reed, J. C. Mechanisms of apoptosis. Am J Pathol 157, 1415–1430, https://doi.org/10.1016/S0002-9440(10)64779-7 (2000).
Rautureau, G. J. P. et al. The restricted binding repertoire of Bcl-B leaves Bim as the universal BH3-only prosurvival Bcl-2 protein antagonist. Cell Death Dis 3, e443, https://doi.org/10.1038/cddis.2012.178 (2012).
Delgado-Soler, L. et al. Molecular determinants of Bim (BH3) peptide binding to pro-survival proteins. J Chem Inf Model 52, 2107–2118, https://doi.org/10.1021/ci3001666 (2012).
Sattler, M. et al. Structure of Bcl-xL-Bak peptide complex: recognition between regulators of apoptosis. Science 275, 983–986, https://doi.org/10.1126/science.275.5302.983 (1997).
Boersma, M. D. et al. Hydrophile scanning as a complement to alanine scanning for exploring and manipulating protein–protein recognition: application to the Bim BH3 domain. Protein Science 17, 1232–1240, https://doi.org/10.1110/ps.032896.107 (2008).
Wink, M. Modes of action of herbal medicines and plant secondary metabolites. Medicines 2, 251–286, https://doi.org/10.3390/medicines2030251 (2015).
Acknowledgements
The authors received financial support from the Deutsche Forschungsgemeinschaft and Ruprecht-Karls-Universität Heidelberg within the funding programme Open Access Publishing.
Author information
Authors and Affiliations
Contributions
M.S. performed the extraction, chemical characterization of the extract, the antioxidant activities in vitro, analyzed the data, assisted in the molecular modelling experiments, wrote the paper, conceived and designed the project. M.F.M designed and performed the hepatoprotective experiments and wrote the paper. R.A.H. designed and performed the antiapoptotic experiments. M.A.O.A. performed the molecular modeling experiments and wrote the paper. O.M.S. and M.A.G. participated in the chemical characterization of the extract and revised the paper. A.M.E. analyzed the data and revised the paper and M.W. revised the paper, conceived and designed the project.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sobeh, M., Mahmoud, M.F., Hasan, R.A. et al. Tannin-rich extracts from Lannea stuhlmannii and Lannea humilis (Anacardiaceae) exhibit hepatoprotective activities in vivo via enhancement of the anti-apoptotic protein Bcl-2. Sci Rep 8, 9343 (2018). https://doi.org/10.1038/s41598-018-27452-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-27452-8
- Springer Nature Limited
This article is cited by
-
Ipomoea carnea mitigates ethanol-induced ulcers in irradiated rats via Nrf2/HO−1 pathway: an in vivo and in silico study
Scientific Reports (2024)
-
Potent hepatoprotective activity of common rattan (Calamus rotang L.) leaf extract and its molecular mechanism
BMC Complementary Medicine and Therapies (2023)
-
In silico and in vitro anti-inflammatory study of phenolic compounds isolated from Eucalyptus maculata resin
Scientific Reports (2023)
-
Colibri® insecticide induces male reproductive toxicity: alleviating effects of Lannea acida (Anacardiaceae) in rats
Basic and Clinical Andrology (2019)
-
Phytochemical analysis of Eucalyptus camaldulensis leaves extracts and testing its antimicrobial and schistosomicidal activities
Bulletin of the National Research Centre (2018)