
Abstract
Reinvestigation of the Kushk and Chahmir areas (Bafq and Behabad regions) of central Iran has yielded a diverse assemblage of Ediacaran fossils, including several new species, just prior to the Cambrian explosion of complex animals. The Kushk series consists mainly of shallow marine carbonate deposits followed by deep-water calcareous marine shales. Ediacaran fossils occur commonly in the shale deposits and include biostratigraphically-important taxa Cloudina and Corumbella, which confirms a latest Ediacaran age for these deposits, the youngest examples of Kimberellomorphs (stem-group molluscs) that helps bridge the gap between their first occurrence in the middle-Ediacaran and the crown diversification in the Cambrian, and likely sponges, which are rare prior to the Cambrian.
Similar content being viewed by others


Introduction
The Ediacara biota represent an enigmatic group of large, multicellular, soft-bodied organisms with a global distribution in the latest Ediacaran (~570-541 Ma)1. Although their phylogenetic affinities are poorly resolved2,3, consensus is emerging that they most likely represent a diverse assemblage of stem and crown group animals in addition to extinct higher-order clades4,5. Their disappearance prior to the Cambrian explosion4 has been explained in many ways6, from an environmentally-driven terminal Ediacaran mass extinction7,8, to a preservational bias resulting from the fossilization (or lack thereof) of soft-tissues9. It has also been suggested that competition with emerging animals may have disrupted classic Ediacara biota, ultimately resulting in their extinction10,11. Importantly, the youngest Ediacaran assemblages (Nama Assemblage)8,12,13,14 are typically faunally depauperate10, dominated by a handful of modular Erniettomorpha15 and fractal Rangeomorpha16, and variety of tubular, often calcified, animals17.
Late Ediacaran fossils from the Lower Shale Member (Chopoqlu Shale) of the Soltanieh Formation of northern Iran18 and shale deposits of the Kushk Series in central Iran19,20,21 include the type material of Permedusites changazensis Hahn and Pflug 1980, however previous reports of Dickinsonia?, Palaeoplatoda?, Pteridinium?, Spriggina?, Yazdia?, and Kushkia? require further investigation in light of recent taxonomic and taphonomic studies18,19,20,21. Here we report the discovery of several new field sites from central Iran (Fig. 1a) that host a strikingly diverse assemblage of coexisting Ediacara biota and likely animals.

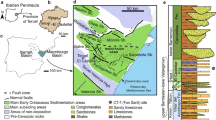
(a) Location of new field sites in central Iran. The Kushk Series accumulated in the Zarigan-Chahmir Basin (map by S.H.V., CorelDRAW Graphic Suite). (b) Stratigraphic column of the Kushk Series, which comprises 13 sub-units consisting of volcanic rhyolites, microdiorites, tuffs, dolomitic limestones, shales, and sandstones. Fossil occurrences are known from sub-units 6 (argillaceous shales).
Geological Setting
The Kushk Series outcrops widely along the western margin of the Lut Block and the eastern margin of the Central Iranian Yazd Block. Sediments accumulated in the Zarigan-Chahmir Basin, which is bounded by the Kuhbanan Fault to the east and the Posht-e-Badam Fault to the west22,23. The Kushk Series reaches a maximum thickness of ~520 m in our sections, although the lower boundary is not exposed and the upper boundary is faulted with the Hashem Formation (Lower Cambrian). The Kushk Series comprises 13 sub-units consisting of volcanic rhyolites, microdiorites, tuffs, dolomitic limestones, dolomite, shales, and sandstones (Fig. 1b) interpreted as a deep, open-marine facies shallowing upwards into a carbonate platform24 resulting from a transgression-regression sequence. Of interest to this study is the fossiliferous sub-unit 6 (30–35 m), which is exposed in multiple areas (Kushk mine site, Chahgaz, Wedge, Chahmir, and Darehdehu). It is comprised of grey, thin-bedded argillaceous shales (containing trace fossils) with intercalation of grey, medium-bedded sandstones and massive sulfide mineralization containing pyritic intercalations, and green tuffs. These grey argillaceous shales can often weather to green (as seen in much of the study area). In the Wedge locality, sub-unit 6 is composed of grey argillaceous shales with intercalation of sandstones, tuff and dolomitic tuffs, while in the Chahmir area it consists of grey, thin-bedded siltstone and argillaceous shales with intercalation of tuff and very thin-bedded mudstone. This sub-unit can be correlated with the Lower Shale Member (Chopoqlu Shale) of the Soltanieh Formation25 in northern Iran. The extensive accumulations of volcanic material and tuffs, combined with the rapid perceived deepening of the platform, suggests that the Kushk basin may have represented an extensional rift basin related to opening of the Proto-Paleotethys sea24 in northeastern Gondwana. Geochronological constraints in the region are limited, with four 207Pb/206Pb dates of 595, 690 and 715 Ma (all ± 120 Ma)26 and 581 ± 8.6 Ma27 from the Kushk area, and a single 207Pb/206Pb date of 540.7 ± 4.8 Ma27 from the Chahmir area. Preliminary sampling of ash beds was unfortunately inconclusive; however, the discovery of classic Precambrian (Chuaria Fig. 2a) and terminal-Ediacaran index fossils (Cloudina, and Corumbella; Fig. 2b,c) support Hamdi and Jiang Zhiwen28 in assigning a latest Ediacaran (to earliest Cambrian) age. Of the three index fossils, Chuaria (n = 1) spans the greatest temporal interval (Tonian to Ediacaran) however it remains restricted to the Precambrian. The lightly-calcifying metazoan Cloudina (n = 37; Fig. 2b) is represented in these sections by external molds morphologically identical to Cloudina molds from Namibia29. Given the taxonomic difficulty of assigning species to dissolution impressions, the range of “cloudinids” is latest Ediacaran30 to possibly the basal Cambrian31. Finally, Corumbella (n = 108, Fig. 2c) is, to the best of our knowledge, restricted to the terminal Ediacaran32,33. As such, the only likely timeframe for the overlap of these three index fossils is latest Ediacaran.

Ediacaran fossils from the Kushk Series, Kushk and Chahmir areas. (a) Organic-walled Chuaria (Ku/14/49), (b) Cloudina (deep negative impression, Ku/14/03a), (c) Corumbella (Ch/14/47), (d) Persimedusites chahgazensis (Ku/14/01), (e) Kuckaraukia (Ch/14/30), (f) unknown tubular organism (Ku/14/09a), (g) Erniettomorph (Ku/14/67), (h,i) Rangeomorph with at least 4 petaloid leaves (1-4) (Ku/14/15), (j) Kimberella persii n.sp. (Ch/14/62), (k) Possible trace fossil Radulichnus (Ku/14/114), (j) Gibbavasis kushkii n.sp. (Ch/14/56a). White (1 cm) and black (0.5 cm) scale bars.
Results
Our new field sites represent the most diverse assemblage of late Ediacaran (Nama-Assemblage ~545-541 Ma) fossils known6,10,12, and includes the previously endemic Ediacaran fossil Persimedusites chahgazensis (n = 41, Fig. 2d;), which has been subsequently described (albeit identified as “Ediacaran discs”) from Argentina34. In addition to Cloudina, and Corumbella, Ediacaran fossils include the recently described Kuckaraukia35 (n = 4, Fig. 2e) from the Russian Urals, rare Erniettomorpha (n = 2, Fig. 2g) and Rangeomorpha (n = 1, Fig. 2h,i), and a diverse assemblage of tubular fossils (n = 16, Fig. 2f).
Of particular importance are two new species that help bridge the gap between the high diversity Ediacaran assemblages of South Australia and Russia, with Cambrian assemblages of sponges and diverse crown bilaterians. Kimberella persii n.sp. (n = 9; Figs 2j, 3) consists of an oval to dumbbell-shaped fossil with anterior-posterior differentiation and clear segmentation along the midline. Although the impressions are compressed, at least three different sediment heights are found within the impression, implying differences in tissue lability and structural rigidity. Kimberella persii n.sp. shares the implied tissue differentiation and segmentation typical of Kimberella quadrata, however lacks the organic frill that outlines the periphery of the implied organic dorsal shell36 (Fig. 3). These characters suggest a bilaterian (stem mollusc) affinity36. Gibbavasis kushkii n.sp. (n = 9; Figs 2i, 4) consists of a small, goblet- to oval-shaped form preserved in negative epirelief with distinct rows of round protrusions, which originally represented external openings later infilled with sediment37,38. The overall columnar shape with distinct incurrent pores is suggestive of a poriferan-grade organism capable of effective filter feeding, however in the absence of siliceous or carbonaceous spicules, it is difficult to assign this species to an existing clade.

Specimens of Kimberella persii n.sp. from the Kushk Series, Chahmir area. (a,b) Specimen Ch/14/62, (c,d) specimen Ch/14/49a, (e,f) specimen Ch/14/49b. Scale bars: 1 cm.

Specimens of Gibbavasis kushkii n.sp. from the Kushk Series, Chahmir area. (a,b) Specimen Ch/14/56a, (c,d) specimen Ch/14/56b, (e) specimen Ch/14/57a, (f) specimen Ch/14/57b. Scale bars: 1 cm.
Discussion
Recent studies of late-Ediacaran sections from Namibia10,11 and southwestern USA8,14 have demonstrated a distinctly depauperate global diversity of classic Ediacara biota (consisting almost exclusively of Erniettomorpha and Rangeomorpha), combined with an increase in diversity and abundance of mineralizing and organic tubular organisms17. The discovery of a high-diversity terminal Ediacaran population from central Iran is pivotal in assessing the nature of the end-Ediacaran extinction, in addition to a possible post White-Sea extinction of Ediacara biota10,12.
What is most striking about this end-Ediacaran assemblage is that it consists almost exclusively of animals. The rare and diminutive (in size) Ernitettomorpha (Fig. 2g) and Rangeomorpha (Fig. 2h,i) in this assemblage are dwarfed by the abundance of metazoans such as Corumbella32, Cloudina39, Kimberella, and Gibbavasis. The discovery of Kimberella persii extends the range of this group up to the Ediacaran-Cambrian boundary, filling the stratigraphic gap between the diverse White-Sea assemblage6 and the diverse crown molluscs known from many Cambrian Lagerstätte40,41. As such, the terminal Ediacaran of Iran showcases a changing Ediacaran ecosystem with a thriving metazoan community.
Systematic Paleontology
Phylum Mollusca Linnaeus, 1758
Genus Kimberella Glaessner and Wade 1966
Kimberella persii n.sp.
Type species
Kimberella quadata Glaessner and Wade 1966.
Diagnosis
Elongate to oval-shaped, bilaterally-symmetrical form with segmented internal divisions represented as elliptical positive ridges along longest positive relief axis. Internal structure consists of four zones including central segments, internal, medial and marginal zones.
Description
Small (9–25 mm in length and 5–15 mm in width) oval (rarely cylindrical) fossil with anterior-posterior differentiation and prominent central segmented ridge (7–25 mm) consisting of 7–25 segments along the major axis. Adjacent to the internal segmented ridge are two parallel-sided smooth furrows 0.5–2 mm in width, surrounded by a slightly elevated and segmented 1–6 mm outer margin that strongly terminates at a distinct marginal rim.
Material
Nine specimens; figured specimens Ch/14/49a, b, 62 (Fig. 3), Chahmir area.
Locality
From gray argillaceous shales (sub-unit 6) of the Kushk Series in the Chahgaz (Dargazin) area near Kushk (Bafq) and Chahmir (Behabad), Central Iran.
Occurrence
Kimberella is known from the Ediacara Hills, Flinders Ranges (South Australia)42; Suz’ma, Karakhta and Solza rivers, Zimnii Bereg, White Sea (Russia)36,43; Kushk Series, Kushk area (Chahgaz (Dargazin) locale) in the Bafq region and Chahmir area in the Behabad region, Central Iran (this study).
Etymology
Kimberella persii, i.e. Kimberella from Persia.
Phylum Uncertain
Genus Gibbavasis n.gen.
Gibbavasis kushkii n.sp.
Diagnosis
Cylindrical to vase-shaped fossils with prominent positive nubs evenly spaced along parallel tracts around circumference.
Description
Small (4–14 mm long, 2–7 mm wide) vase-shaped, negative-relief impression consisting of regularly spaced positive relief and hemispherical bumps that run longitudinally as seven to nine rows across the width of the vase. Rows are also apparent latitudinally, resulting in a lattice-like appearance.
Remarks
Prominent positive-relief bosses likely represent a taphonomic inversion of an originally negative pit infilled with sediment. Pits may have served as an entryway for water currents. Specimens similar in construciton to the terminal Ediacaran fossil Ausia (from the Nama Assemblage), however Gibbavasis is noticeably smaller and vase-shaped rather than fan-shaped. The affinities of Ausia are unknown.
Material
Nine specimens; figured specimens Ch/14/56a, b, 57a, b (Fig. 4), Chahmir area.
Locality
From gray argillaceous shales (sub-unit 6) of the Kushk Series in the Kushk area (Chahgaz (Dargazin) locale in the Bafq region and Chahmir area in the Behabad region, Central Iran.
Etymology
Gibbavasis (Latin Gibba (bumpy) vasis (vase)) kushkii (from the Kushk Series of Central Iran).
Methods
More than 250 Ediacaran specimens were collected (in situ and also from float) from the bed tops of the argillaceous shales (sub-unit 6; Fig. 1b) of the Kushk Series in Kushk area (Chahgaz (Dargazin) and Wedge locales in the Bafq region, and Chahmir and Darehdehu areas in the Behabad region, Central Iran. 95 specimens from the Kushk area and 150 specimens from the Chahmir area were investigated and identified. All specimens were washed and photographed, after which detailed morphological data were collected for all specimens.
References
Narbonne, G. M., Xiao, S., Shields, G. A. & Gehling, J. G. The Ediacaran Period. In Felix M. Gradstein, James G. Ogg, Mark Schmitz and Gabi Ogg, The Geologic Time Scale 2012. Published by Elsevier BV. Chapter 18, 413–435 (2012).
Xiao, S. & Laflamme, M. On the eve of animal radiation: Phylogeny ecology and evolurion of the Ediacara biota. Trends in Ecology and Evolution 24, 31–40 (2009).
Budd, G. E. & Jensen, S. The origin of the animals and a “Savannah” hypothesis for early bilaterian evolution. Biological Reviews, https://doi.org/10.1111/brv.12239 (2015).
Erwin, D. H., Laflamme, M., Tweedt, S. M. & Sperling, E. A. The Cambrian conundrum: Early Divergence and Later Ecological Success in the Early History of Animal. Science 334, 1091–1097 (2011).
Dececchi, T., Narbonne, G. M., Greentree, C. & Laflamme, M. Relating Ediacaran Fronds. Paleobiology 43(2), 171–180 (2017).
Laflamme, M., Darroch, S. A. F., Tweedt, S. M., Peterson, K. J. & Erwin, D. H. The end of the Ediacara biota: Extinction, biotic replacement, or Cheshire Cat? Gondwana Research 23, 558–573 (2013).
Amthor, J. E. et al. Extinction of Cloudina and Namacalathus at the Precambrian-Cambrian boundary in Oman. Geology 31(5), 431–434 (2003).
Smith, E. F. et al. The end of the Ediacaran: Two new exceptionally preserved body fossil assemblages from Mount Dunfee, Nevada, USA. Geology 44(11), 911–914 (2016).
Gehling, J. G. & Droser, M. L. How well do fossil assemblages of the Ediacara Biota tell time? Geology 41(4), 447–450 (2013).
Darroch, S. A. et al. Biotic replacement and mass extinction of the Ediacara biota. Proceedings of the Royal Society B 282, 20151003 (2015).
Darroch, S. A. F. et al. A mixed Ediacaran-metazoan assemblage from the Zaris sub-basin, Namibia. Palaeogeography, Palaeoclimatology, Palaeoecology 459, 198–208 (2016).
Boag, T. H., Darroch, S. A. F. & Laflamme, M. Ediacaran distributions in space and time: testing assemblage concepts of earliest macroscopic body fossils. Paleobiology 42(4), 574–594 (2016).
Narbonne, G. M., Saylor, B. Z. & Grotzinger, J. P. The youngest Ediacaran fossils from Southern Africa. Journal of Paleontology 71, 953–967 (1997).
Smith, E. F., Nelson, L. L., Tweedt, S. M., Zeng, H. & Workman, J. B. A cosmopolitan late Ediacaran biotic assemblage: new fossils from Nevada and Namibia support a global biostratigraphic link. Proceedings of the Royal Society B 284, 20170934, https://doi.org/10.1098/rspb.2017.0934 (2017).
Ivantsov, A. Y., Narbonne, G. M., Trusler, P. W., Greentree, C. & Vickers-Rich, P. Elucidating Ernietta: new insights from exceptional specimens in the Ediacaran of Namibia. Lethaia 49, 540–554 (2016).
Vickers-Rich, P. et al. New Discoveries form the Ediacaran of southern Namibia. Journal of Paleontology 87, 1–15 (2013).
Schiffbauer, J. D. et al. The latest Ediacaran Wormworld Fauna: Setting the ecological stage for the Cambrian explosion. GSA Today 26, 4–11 (2016).
Jafari, S. M., Shemirani, A. & Hamdi, B. Microbiostratigraphy of the Late Ediacaran to Ordovician in NW Iran (Takab area). Geological Society of London, Special Publication, 286, 433–437 (2007).
Hahn, G. & Pflug, H. D. Ein neuer Medusen-Fund aus dem Jung-Präkambrium von Zentral-Iran. Senckenbergiana Lethaea 60(4/6), 449–461 (1980).
Glaessner, M. F. The Dawn of Animal Life. Cambridge University Press, 296p (1984).
Hamdi, B. Sedimentary rocks of Precambrian-Cambrian in Iran, Geological Survey of Iran press, no. 20, 353p [in Persian] (1995).
Gibbs, A. Geology and genessis of the Bafq lead-zinc deposit. Mines and Metals B., 205–220 (1976).
Rajabi, A., Rastad, E., Alfonso, P. & Canet, C. Geology, ore facies and sulphur isotopes of the Koushk vent-proximal sedimentary-exhalative deposi, Posht-e-Badam Block, Central Iran. International Geology Review 54, 1635–1648 (2012).
Lasemi, Y. Facies analysis, depositional environments and sequences stratigraphy of the Upper Pre-Cambrian and Paleozoic rocks of Iran. Geological Survey of Iran press, no. 78, 180p [in Persian] (2001).
Stöcklin, J., Ruttner, A. & Navavi, M. New data on the Lower Paleozoic and Precambrian of North Iran. Geological Survey of Iran, report no. 1, 13 (1964).
Huchriede, R., Kursten, M. & Venzlaff, H. ZurGeologie de Gebietes Zwischen Kerman und Sagand (Iran). Geol. Jb. Beih. 51, 1–197, Abb. 1–50, Taf. 1–10: Hannover (1962).
Vickers-Rich, P. et al A preliminary report on new Ediacaran fossils from Iran. Alcheringa: An Australasian Journal of Paleontology, https://doi.org/10.1080/3115518.2017.1384061 (2017).
Hamdi, B. & Jiang Zhiwen Paleozoic fossils from the Morad and Rizu series in Central Iran. Journal of Geosciences, Geological Survey of Iran, 1 ( 4 ), 26-35 [in Persian with English abstract] (1992).
Grotzinger, J. P., Bowring, S. A., Saylor, B. Z. & Kaufman, A. J. Biostratigraphy and Geochronologic Constraints on Early Animal Evolution. Science 270(5236), 598–604 (1995).
Xiao, S. et al. Toward an Ediacaran time scale: problems, protocols, and prospects. Episodes 39(4), 540–555 (2016).
Zhu, M. et al. A deep root for the Cambrian explosion: Implications of new bio- and chemostratigraphy from the Siberian Platform:. Geology 45(5), 459–462 (2017).
Babcock, L. E., Grunow, A. W., Sadowski, G. R. & Leslie, S. A. Corumbella, an Ediacaran-grade organism from the Late Neoproterozoic of Brazil. Palaeogeography, Palaeoclimatology, Palaeoecology 220, 7–18 (2005).
Warren, L. V. et al. The dawn of animal skeletogenesis: ultrastructural analysis of the Ediacaran metazoan Corumbella werneri. Geology 40(8), 691–694 (2012).
Arrouy, M. J. et al. Ediacaran discs from South America: probable soft-bodied macrofossils unlock the paleogeography of the Clymene Ocean. Scientific Reports 6, 30590, https://doi.org/10.1038/srep30590 (2016).
Razumovskiy, A. A., Ivantsov, A. Y., Novikov, I. A. & Korochantsev, A. V. Kuckaraukia multituberculata: A new Vendian fossil from the Basa Formation of the Asha Group in the South Urals. Paleontological Journal 49(5), 449–456 (2015).
Fedonkin, M. A., Simoneta, A. & Ivantsov, A. Y. New data on Kimberella, the Vendian mollusc-like organism (White Sea region, Russia): palaeoecological and evolutionary implications. Geological Society of London, Special Publication 286, 157–179 (2007).
Gehling, J. G. Microbial Mats in Terminal Proterozoic Siliciclastics: Ediacaran Death Masks. Palaios 14, 40–57 (1999).
Kenchington, C. G. & Wilby, P. R. Of time and taphonomy: Preservation in the Ediacaran. Paleontological Society Paper 20, 101–122 (2014).
Penny, A. M. et al. Ediacaran metazoan reefs from the Nama Group, Namibia. Science 344(6191), 1504–1506, https://doi.org/10.1126/science.1253393 (2014).
Paterson, J. R. et al. The Emu Bay Shale Konservat-Lagerstätte: a view of Cambrian life from East Gondwana. Journal of the Geological Society 173, 1–11 (2015).
Vinther, J., Sperling, E. A., Briggs, D. E. G. & Peterson, K. J. Amolecular palaeobiological hypothesis for the origin of aplacophoran molluscs and their derivation from chiton-like ancestors. Proceedings of the Royal Society B 279, 1259–1268 (2012).
Glaessner, M. F. & Daily, B. The geology and Late Precambrian fauna of the Ediacara fossil reserve. Records of the South Australian Museum 13, 369–401 (1959).
Fedonkin, M. A. & Waggoner, B. M. The Late Precambrian fossil Kimberlla is a molluse-like bilaterian organism. Nature 388, 868–871 (1997).
Acknowledgements
Research supported by an Early Researcher Award (ERA), Natural Science and Engineering Research Council of Canada (NSERC) Discovery Grant (RGPIN 435402), and a Research and Scholarly Activity Fund to Laflamme. Thanks are extended to the Islamic Azad University to award a sabbatical to Vaziri at the University of Toronto Mississauga and B. Hamdi, Y. Lasemi, D. Jahani, N. Kohansal Ghadimvand, M.M. Zand, H. Omidi, A. Shariati and H. Zamani for useful discussions and support.
Author information
Authors and Affiliations
Contributions
Field work: S.H.V. and M.R.M. Geology: S.H.V. Laboratory study: S.H.V. Systematic paleontology: S.H.V. and M.L. Data analysis: S.H.V. and M.L. Wrote the paper: S.H.V. and M.L. Drawings, photography, and figures: S.H.V. and M.L.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vaziri, S.H., Majidifard, M.R. & Laflamme, M. Diverse Assemblage of Ediacaran fossils from Central Iran. Sci Rep 8, 5060 (2018). https://doi.org/10.1038/s41598-018-23442-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-23442-y
- Springer Nature Limited
This article is cited by
-
A late-Ediacaran crown-group sponge animal
Nature (2024)
-
Cryogenian and Ediacaran integrative stratigraphy, biotas, and paleogeographical evolution of the Qinghai-Tibetan Plateau and its surrounding areas
Science China Earth Sciences (2024)