
Abstract
Declines in the abundance of long-lived and habitat-forming species on continental shelves have attracted particular attention given their importance to ecosystem structure and function of marine habitats. The study of undisturbed habitats defined as “pristine areas” is essential in creating a frame of reference for natural habitats free of human interference. Gorgonian species are one of the key structure-forming taxa in benthic communities on the Antarctic continental shelf. Current knowledge of the diversity, distribution and demography of this group is relatively limited in Antarctica. To overcome this lack of information we present original data on pristine and remote populations of gorgonians from the Weddell Sea, some of which display the largest colony sizes ever recorded in Antarctica. We assessed the distribution patterns of seven gorgonian species, a morphogroup and a family in front of the Filchner Ronne Ice Shelf (Weddell Sea) by means of quantitative analysis of video transects. Analysis of these videos showed a total of 3140 colonies of gorgonians with the highest abundance in the southern section and a significantly clumped distribution. This study contributes to the general knowledge of pristine areas of the continental shelf and identifies the eastern Weddell Sea as a hotspot for habitat-forming species.
Similar content being viewed by others
Introduction
The current state of the oceans is very different from what it was in the past1,2. Actually, most marine ecosystems are affected by climate change (e.g. ocean warming, acidification, sea level rise)3,4,5 and other multiple human-derived threats (e.g. overfishing, pollution, habitat destruction)6,7,8 which threaten marine global biodiversity and modify oceanic environments5 to the point of being considered “unnatural oceans”9 nearly devoid of “pristine” areas10. Such pristine areas are minimally affected by major human threats, thus providing a unique opportunity to better understand how marine ecosystems are structured and behave11,12. They are also essential to study the effects of climate change on benthic communities13, particularly on the Antarctic continental shelf where one can find still relatively undisturbed environments14,15. The potential impact of trawling activity has also become a major concern due to its extensive damage to continental shelves and deep cold-water coral reefs16,17. Although there is evidence of fishing activity as by-catch from longline fisheries in South Georgia18 and in the Ross Sea19, most of the Antarctic continental shelf has been little influenced by industrial fishing20. The lack of terrigenous sediments21, the relative constancy of its physical conditions22,23 and the relative absence of human-derived impacts5, make the Antarctic continental shelf a highly favourable environment for the development of high-density benthic megafauna communities.
In the last few decades, studies carried out on the continental shelf of the northeastern Weddell Sea have generated key insights on the diversity24,25, the degree of heterogeneity26,27, and the impact of iceberg scouring28,29,30. Iceberg scouring constitutes one of the major natural disturbances for high-Antarctic shelf fauna and it is increasingly apparent that iceberg scouring events may be altered by iceberg calving associated with regional atmospheric warming31,32. The estimated rate of disturbance of the Antarctic continental shelf by grounding icebergs is approximately 5%26, although still considerably less than the 53% attributable to trawling in other continental shelves33.
Gorgonians are among the main structural species of many benthic communities across all latitudes and depths, from shallow sublittoral habitats to continental shelves and deep seas34,35,36. Hence, the Commission for the Conservation of Antarctic Marine Living Resources (CCAMLR) has recognized gorgonians as a Vulnerable Marine Ecosystems (VME) indicator taxon37. These organisms contribute to the structure of benthic communities adding three-dimensional complexity to the habitat38,39. During the last decade, knowledge about diversity, distribution, ecology and state of conservation of gorgonian populations on the continental shelf has significantly increased in the Mediterranean Sea40,41, the Pacific Ocean42,43 and the Atlantic Ocean44. In Antarctica, most of the studies of this group of organisms have focused on taxonomy45,46, trophic ecology47,48, growth rates49,50 and reproductive ecology51,52. However, despite the high abundance of gorgonians in some locations of the Weddell Sea and their ecological role in Antarctic benthic communities30,53, there is still an important lack of knowledge on their ecological characteristics such as spatial distribution, abundance and demographic processes54.
Non-destructive sampling techniques like video-equipped towed gear, Remotely Operated Vehicles (ROVs) or manned submersibles are commonly used to study coastal areas55, deep reefs of cold-water corals56, seamounts57, and mesophotic areas58 to assess biodiversity patterns, characterize communities, evaluate spatial and temporal changes, and assess benthic ecosystem health status59,60. Although the majority of studies on Antarctic benthos have been carried out using semi-quantitative techniques like Agassiz and bottom trawls61,62, non-destructive image methodology has also been commonly used in the high Antarctic to provide quantitative information on the distributional patterns of benthic megafauna communities over large spatial and bathymetrical domains28,53,58,63,64,65.
Knowledge of demographic processes and spatial distribution patterns is a prerequisite to understand their role in benthic communities and provide basic information on their underlying dynamics and resilience, as well as to facilitate their management and conservation66,67. The major aim of the present study was to assess the health status of Antarctic gorgonian assemblages in a pristine and remote area in the southernmost part of the Weddell Sea continental shelf. Specifically, (1) we characterized the diversity and the abundance of gorgonians group; (2) explored their distribution patterns, and (3) assessed their population size structure. This study attempts to be a benchmark for the investigation of continental shelf habitats modified by anthropogenic pressure and to contribute to the general knowledge of pristine areas with habitat-forming species.
Results
Abundance of gorgonian populations
A total of 3140 colonies (1402 in the north and 1738 in the south), comprising seven gorgonian species (Ainigmaptilon sp., Dasystenella acanthina, Fannyella rossii, Fannyella spinosa, Thouarella sp.1, Thouarella sp.2, and Thouarella variabilis), an unbranched morphogroup (which included specimens from the genus Onogorgia, Armadillogorgia, Primnoella and Arntzia) and the family Isididae, were counted along six transects (Supplementary Table S1), occurring with a frequency of 64.4% in 1836 sampling units of 1 m2. Based on our count data, we estimated more than 46000 and more than 97000 colonies for the north and the south region, respectively. Overall, Thouarella sp.1 was the most abundant (n = 597 total colonies across all transects) and the second most frequent species representing 19% of observed colonies present in 20% of the sampling units. Thouarella sp.2 was the second most abundant (n = 572), but the most frequent (20%) species. The family Isididae (n = 535, 17.8%), Fannyella rossii (n = 474, 17.5%) and Thouarella variabilis (n = 438, 15.4%) were the third, fourth and fifth most abundant and frequent species, respectively. The unbranched group (n = 280) was more abundant than Dasystenella acanthina (n = 189), but less frequent (5.4% and 7.5%, respectively). The other species accounted for less than 2% of the observed colonies, occurring in less than 3% of the sampling units. Generally, abundance of the gorgonians differed between the two study sections, being lower in the northern part. Only Isididae (n = 277) and Dasystenella acanthina (n = 87) abundance showed high values in the northern stations (Fig. 1).

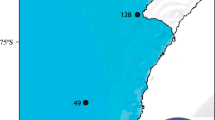
Composition, distribution and abundance of gorgonian species. The pie charts display the percentage of gorgonian taxa at each transect (n = number of colonies per transect). The size of the pie charts represents the abundance of the gorgonians. The histogram shows the abundance of gorgonian species per region (north: stations 128, 136, 170; south 49, 81, 86). Map of Antarctica was downloaded from http://www.ibcso.org/data.html 100. The figure was generated with QGIS Version 2.12 http://www.qgis.org/it/site/.
Spatial distribution
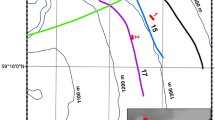
Ripley’s K analysis revealed a significantly clumped distribution of the family Isididae, Thouarella sp.1 and Thouarella sp.2 colonies at all scales (from 1 m2 to whole transect). Thouarella variabilis showed a clumped distribution in the north, but a random distribution in the south. An opposite result for spatial distribution was found for Dasystenella acanthina, Fannyella rossii and Unbranched (Fig. 2). Gorgonian abundances varied markedly among the various stations (F5,1179 = 53.3, p < 0.001) (Fig. 3). In stn. 49, the Unbranched morphogroup was the group of gorgonians with the highest abundance (47 col/m2) and mean density of 7.3 ± 11.5 col/m2, while the least abundant species were Fannyella spinosa and Ainigmaptillon sp., with highest abundance of 20 and 19 col/m2 and mean density of 1.3 ± 0.6 and 1.2 ± 0.4 col/m2, respectively (Fig. 3).

L-function (Ripley’s K) for the most abundant species. Values below 95% confidence interval (dotted lines) indicate a statistically significant clumped distribution of colonies; values within the confidence interval indicate a random distribution; values above the confidence interval indicate a statistically significant over-dispersed distribution (n = number of colonies).

Density plots. Densities of the most abundant species are plotted at each station (n = number of colonies). Substrate type is indicated as black (coarse sediment), dark grey (intermediate sediment) and light grey (fine sediment).
Population size structure
The size-frequency distributions of the 7 species populations were unimodal (Fig. 4). Most of the gorgonian populations were positively skewed, indicating an asymmetrical distribution of size frequency and a high proportion of small colonies (0–10 cm) (Fig. 4 and Supplementary Table S2). In contrast, the two populations of the Unbranched morphogroup were not skewed, being dominated by medium-sized colonies (10–20 cm in the south and 50–60 cm in the north) (Fig. 4 and Supplementary Table S2). Most of the northern populations showed no significant kurtosis while in the south all the gorgonian populations, except Unbranched, showed significant kurtosis (Supplementary Table S2). Finally, in both studied areas, all gorgonian populations displayed the same size class distributions (Fig. 4).

Size-frequency distribution of gorgonian populations (n = number of colonies).
Population structure
The structure of gorgonian populations gradually differed in both areas, with significant differences between north and south (F1,1183 = 65.06, p < 0.001) (Fig. 5).

Non-metric multidimensional scaling (nMDS) ordination plot of abundance of gorgonian species in the south and north region of the SE Weddell Sea. Analysis performed on Bray-Curtis dissimilarities for abundance (colonies/m2). Each point represents a subsample of 200 m length for each station.
The SIMPER analysis showed an average similarity in species composition which ranged from 17.89% to 31.68% (Supplementary Table S3). The number of species contributing up to 90% of the similarity was the same in the two sections (Supplementary Table S3). Thouarella sp.1 contributed most (25.14%) to the similarity in the northern transects, while the family Isididae was especially relevant in the southern transects (22.43% of contribution). Focusing on taxa dissimilarities, the family Isididae was the most important contributing with 17% of the average dissimilarity between north and south.
Discussion
This study focused on previously unknown extensive gorgonian assemblages in the southeastern Weddell Sea. Our results indicate that this region is a hotspot for gorgonian diversity in terms of both number of species and their abundance. The diversity of the studied gorgonian assemblages was similar to those reported on subtropical68, Mediterranean shelf41 and other Antarctic coastal areas69. Seven different species, a gorgonian morphogroup and a gorgonian family Isididae were observed to dwell between 250 and 350 m depth within the study area (Fig. 1), in agreement with previous findings in coastal areas of the Antarctic Peninsula69. On the other hand, gorgonian density observed in these multi-specific assemblages clearly exceeds (by ten-fold; 47 colonies/m2) those reported for other Antarctic and Arctic shelf areas70,71 (Supplementary Table S4). These high density values were similar to those found in temperate72,73 and tropical74,75 coastal assemblages (Supplementary Table S4). Despite extreme environmental conditions and the general theory that species richness decreases with increasing latitude, it is also generally accepted that this theory is not strictly true and varies with species in the Southern Ocean76,77. In addition, benthic biomass in some Antarctic areas is larger than in temperate and subtropical areas78. Such dense three-dimensional communities cover large sections of the Antarctic continental shelf as well as the Mediterranean Sea41 and deep undisturbed North Atlantic coral banks44. Fannyella rossii and the three species of genus Thouarella showed high abundances in each video transect (Supplementary Table S1). This highlights the unique abundance of these Antarctic gorgonian species45. Of additional note is their high density, with maximum values of 6 ind/m2 for Fannyella rossii and 10 ind/m2 for Thouarella sp.1 in the southern section (Supplementary Table S1).
All populations in this study were represented by many small colonies with a positively skewed colony size distribution (Fig. 4). The size structure of a population results from the action of biotic and abiotic factors and from the type, intensity, and frequency of disturbance to which individuals are exposed79,80. Positively skewed size frequency distribution implies that a population is in a healthy state and growing, since it includes an abundance of juveniles81,82. On the contrary, a negative skewness indicates a lack of recent recruitment and therefore it implies a risk of population decline81,83. Population size structure of all Thouarella and Fannyella species was mostly asymmetrical with many small colonies (Fig. 4), suggesting high recruitment rates82. The size structure also reflects the growth and the development of each individual within the population, as well as past recruitment and mass mortality events. Unfortunately, due to their inaccessibility, only a few studies of gorgonian population size structure have been done on continental shelves41. Unbranched individuals seem to grow slowly and older without being replaced (low recruitment) with few small colonies (sexual juveniles) and dominance of large-sized individuals41.
As a rule of thumb, because of their slow growth rate84,85 and reproduction type, gorgonians are especially vulnerable to iceberg scouring86,87, making their recovery very slow17. All gorgonian species described in this study reproduce by internal brooding. This means that the settlement of the larvae occurs at short distances from the parents88 explaining the patchy distribution along all video transects. Some gorgonian species such as Ainigmaptillon antarcticum and Primnoisis antarctica, which are internal brooders, are also pioneer taxa appearing during the first stage of recolonization after iceberg scouring events with patchy distribution86,87.
The high diversity and abundance of gorgonian assemblages on the Antarctic continental shelf, and the vast area covered by high densities of genus Thouarella are probably related to the low iceberg scouring pressure and oceanographic-ice conditions. Constant hydrodynamic conditions that increase particle suspension in the near-bottom water layers may also imply enhanced food availability for gorgonians on the continental shelf89. Strong currents are advantageous to the establishment of this group of organisms supplying them with food and continuously keeping them completely clear of sediment90,91. Moreover, based on our findings of high abundance and large sizes in the southern section of our study area, we hypothesize that it is little affected by iceberg scouring, thereby favouring the establishment of well-developed pristine gorgonian populations.
Reduced abundance of long-lived and habitat-forming species from the deep sea and continental shelves in shallow sublittoral habitats have attracted particular attention, given their disproportionate importance to ecosystem structure and function, and the social value of marine habitats14. Yet, factors responsible for such decline are mainly overexploitation and habitat destruction by bottom trawling and by-catch fishing92. Evidence of fishing activities with Thouarella spp. as by-catch has been reported in South Georgia18. Moreover, specimens from the genus Primnoa and the family Isididae from longline fisheries were found in the Ross Sea19. To our knowledge, our results are the first to show pristine populations of gorgonians with the highest abundance and largest size ever recorded on the Antarctic continental shelf. These populations are far more mature and better preserved than any other known population in Antarctica. Clearly, more research is needed to determine the locations of such refuges and to devise strategies to protect such gorgonian populations as well as the many other species interacting with them. The study of these pristine gorgonian populations may also provide basic knowledge on how other continental shelf and upper slope communities may have thrived in the decades before bottom trawling fishing ensued.
Methods
Study area
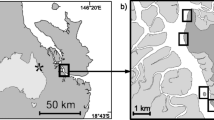
The study area was sampled as part of the multidisciplinary PS82 (ANT XXIX/9) expedition on board R/V Polarstern from December 19, 2013, to March 5, 201493. It is located in front of the Filchner Ronne Ice Shelf in the southernmost part of the Weddell Sea; a region poorly investigated due to the heavy sea ice conditions93. The small amount of data available from this area has made it an area of special relevance to better understand oceanographic conditions and to gain new insights into biodiversity patterns in this remote and pristine region94. The study area was divided into a south and a north section due to Brunt Ice Shelf, which may produce different oceanographic conditions93.
ROV sampling procedure
In order to study the composition and distribution of gorgonians, an inspection-class ROV (Remotely Operated Vehicle, Ocean Modules V8 Sii) was deployed at six stations in the area of the Filchner Trough (Supplementary Table S5). Three random stations (stn. 49, stn. 81, stn. 86) were recorded in the southern part of the continental shelf and three random stations (stn. 128, stn. 136, stn. 170) in its northern part closer to the shelf break. The ROV was equipped with a High Definition (HD) video camera (Kongsberg oe14–502) looking forward in an angle of 40–45° and two parallel lasers providing a reference scale of 4 cm on the video (see Knust and Schröder 201493 for more details on the ROV procedure). The ROV video material is available from the data publisher PANGEA at www.pangea.de (see Table S5 for DOIs).
Species identification
In order to confirm the taxonomic identification of the species observed in the videos, colonies of gorgonians were collected with an Agassiz Trawl (AGT) after the ROV deployments. Colonies were fixed and preserved in 10% formalin until analysed in the laboratory (see Supplementary Table S6 for taxonomic remarks on the identification of some groups of species). We identified 7 gorgonian species belonging to the family Primnoidae (Ainigmaptilon sp., Dasystenella acanthina, Fannyella rossii, Fannyella spinosa, Thouarella sp.1, Thouarella sp.2, and Thouarella variabilis), an unbranched morphogroup (flagelliform colonies with polyps distributed in whorls along the main stem of the colony), which included specimens of the genera Onogorgia, Armadillogorgia, Primnoella and Arntzia, and a bamboo coral group of the Family Isididae, (Supplementary Figures S1 and S2).
Video analysis
Quantitative video analysis was performed using the software SONY XDCAM Viewer. Every gorgonian observed within a width of 0.3 m (based on the laser beams) along each video transect was identified with a distance from the beginning of the transect according to the ROV’s ultra-short baseline (USBL) position data.
Spatial distribution and size structure
We examined the species composition and quantified the frequency as the relative proportion of each species present for each sampling unit of the transect and the abundance as the total number of colonies across all the transects (see below). The most abundant species of gorgonians were used to compare their abundance, spatial distribution and size class in both the north and south areas. These results were displayed in density plots, obtained by transforming each transect into a string of contiguous quadrats of 1 m2 (0.3 × 3.33 m) and counting the number of colonies of each species only inside each quadrat. A total of 1836 useful sampling units were obtained from the 6 transects.
The significance of the deviation from a random distribution was analysed with the one-dimensional version of Ripley’s K-function second-order spatial statistic95,96. When the sample statistic is found within the bounds of the confidence interval at any point, it indicates complete spatial randomness; a significant positive deviation of the sample statistic indicates over-dispersion of the colonies, whereas a significant negative deviation indicates a clumped distribution67.
To study population size structure, the maximum height of each observed gorgonian colony was measured using the Macnification 2.0.1 software on still images extracted from recorded footage97. The distance between the two laser beams was used to calibrate extracted images and measurements were performed on still images in which the laser beams were in the same plane as the colony base to reduce the error due to the perspective55. Based on previous studies, colony size class was defined for each 10 cm55,82. We considered as young colonies the smallest colonies that could be distinguished using the video analysis (2–5 cm in height)80. Size structure was also analysed in terms of descriptive statistics using distribution parameters such as skewness and kurtosis. Skewness is a measure of the symmetry of a distribution using its mean, reflecting the proportion of small versus large colonies in a gorgonian population; if skewness is significant the distribution is asymmetric. Kurtosis is a measure of the peakedness of a distribution near its central mode. A significant kurtosis value indicates longer tails than would be expected for a normal distribution, and therefore a particular colony size prevails in the population. Only transects with more than 40 colonies were studied for population size structure in order to generate meaningful skewness and kurtosis estimates.
Population size structure and density data from other areas of the continental shelf
To compare our data with that of other gorgonian populations dwelling on the continental shelf of other seas, we compiled data on maximum abundance, mean density and maximum height from previous studies using ROV observations. Overall, we compiled population structure data for 36 taxa and 12 different study areas (Supplementary Table S4).
Assemblage structure
A non-metric multi-dimensional scaling (nMDS) ordination analysis was performed based on the Bray–Curtis similarity measure using square-root-transformed abundance data. For visualization purposes, data are presented for each 200 m length. Furthermore, a similarity percentage procedure analysis, SIMPER98, was performed to identify the relative contribution of each species to average similarities between areas. A non-parametric analysis of variance, PERMANOVA99, was applied using Bray-Curtis distance for the multivariate analyses. Statistical analyses were computed using the program Primer v6 with the PERMANOVA + add-on package.
References
Jackson, J. B. C. Reefs since columbus. Coral reefs 16, 23–32 (1997).
Halpern, B. S. et al. An index to assess the health and benefits of the global ocean. Nature 488, 615–620 (2012).
Blunier, T. & Brook, E. J. Timing of millenial-scale climate change in Antarctica and Greenland during the last glacial period. Science 291, 109–112 (2001).
Hoegh-Guldberg, O. P. J. et al. Coral reefs under rapidclimate change and ocean acidification. Science 318, 1737–1742 (2007).
Halpern, B. S. et al. A global map of human impact on marine ecosystems. Science 319, 948–952 (2008).
Jackson, J. B. C. et al. Historical overfishing and the recent collapse of coastal ecosystems. Science 293, 629–638 (2001).
Worm, B. et al. Impacts of biodiversity loss on ocean ecosystem services. Science 314, 787–790 (2006).
Estes, J. A. et al. Trophic downgrading of planet Earth. Science 333, 301–306 (2011).
Jackson, J. B. C. & Sala, E. Unnatural oceans. Sci Mar 65, 273–281 (2001).
Cressey, D. Antarctic seas in the balance: plans to protect swathes of ocean face tough test. Nature 490, 324–325 (2012).
Sandin, S. A. et al. Baselines and degradation of coral reefs in the northern Line Islands. PloS One 3(2), e1548 (2004).
Knowlton, N. & Jackson, J. B. C. Shifting baselines, local impacts, and global change on coral reefs. PLoS Biol 6(2), e54 (2008).
Levin, L. A. & Dayton, P. K. Ecological theory and continental margins: where shallow meets deep. Trends ecol evol 24(11), 606–617 (2009).
Clarke, A. & Harris, C. M. Polar marine ecosystems: major threats and future change. Environ Conserv 30, 1–25 (2003).
Smetacek, V. & Nicol, S. Polar ocean ecosystems in a changing world. Nature 437, 362–68 (2005).
Hall–Spencer, J., Allain, V. & Fosså, J. H. Trawling damage to Northeast Atlantic ancient coral reefs. P Roy Soc Lond B Bio 269, 507–511 (2002).
Althaus, F. et al. Impacts of bottom trawling on deep-coral ecosystems of seamounts are long-lasting. Mar Ecol Prog Ser 397, 279–294 (2009).
Taylor, M. L. Distribution and diversity of octocorals from longline by-catch around South Georgia, UK. PhD Thesis. Imperial College London (2011).
Parker, S. J. & Bowden, D. A. Identifying taxonomic groups vulnerable to bottom longline fishing gear in the Ross Sea region. CCAMLR Science 17, 105–127 (2010).
Ainley, D. G. & Pauly, D. Fishing down the food web of the Antarctic continental shelf and slope. Polar Record 50, 92–107 (2014).
Anderson, J. B. Antarctic marine geology. Cambridge University Press (1999).
Hedgpeth, J. W. The Antarctic marine ecosystem. In Adaptations within Antarctic ecosystems, Proceedings of the third SCAR Symposium on Antarctic Biology. (ed. Llano, G. A.) 3–10 (Gulf Publishing, Houston 1977).
Picken, G. B. Marine habitats-benthos. In Key environments Antarctica. (ed. Bonner, W. N. & Walton, D. W. H.) 154–172 (Pergamon Press, Oxford, 1985).
Voß, J. Zoogeographie und Gemeinschaftsanalyse des Makrozoobenthos des Weddellmeeres (Antarktis). Berichte Polarforsch 45, 1–145 (1988).
Gutt, J. & Starmans, A. Structure and biodiversity of megabenthos in the Weddell and Lazarev Seas (Antarctica): ecological role of physical parameters and biological interactions. Polar Biol 20, 229–247 (1998).
Gutt, J. Some “driving forces” structuring communities of the sublittoral Antarctic macrobenthos. Ant Sci 12, 297–313 (2000).
Teixidó, N., Garrabou, J. & Arntz, W. E. Spatial pattern quantification of Antarctic benthic communities using landscape indices. Mar Ecol Prog Ser 242, 1–14 (2002).
Gutt, J. & Starmans, A. Quantification of iceberg impact and benthic recolonization patterns in the Weddell Sea (Antarctica). Polar Biol 24, 615–619 (2001).
Gutt, J. & Piepenburg, D. Scale-dependent impact on diversity of Antarctic benthos caused by grounding of icebergs. Mar Ecol Prog Ser 253, 77–83 (2003).
Teixidó, N., Garrabou, J., Gutt, J. & Arntz, W. E. Iceberg disturbance and successional spatial patterns: the case of the shelf Antarctic benthic communities. Ecosystems 10, 143–158 (2007).
Scambos, T. A., Hulbe, C., Fahnestock, M. & Bohlander, J. The link between climate warming and break-up of ice shelves in the Antarctic Peninsula. J Glaciol 46, 516–530 (2000).
Barnes, D. K. A., Fenton, M. & Cordingley, A. Climate-linked iceberg activity massively reduces spatial competition in Antarctic shallow waters. Curr Biol 24, R553–R554 (2014).
Mc Allister, D. E. Status of the world ocean and its biodiversity. Sea Wind 9, 1–72 (1995).
Gili, J. M., Murillo, J. & Ros, J. The distribution pattern of benthic cnidarians in the western Mediterranean. Sci Mar 53, 19–35 (1989).
Jones, C. G., Lawton, J. H. & Shachak, M. Organisms as ecosystem engineers. Oikos 69, 373–386 (1994).
Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. Oceanogr Mar Biol 48, 123–195 (2006).
Jones, C. G. & Lockhart, S. J. Detecting Vulnerable Marine Ecosystems in the Southern Ocean using research trawls and underwater imagery. Mar Policy 35, 732–736 (2011).
Jones, C. G., Lawton, J. H. & Shachak, M. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78, 1946–1957 (1997).
Orejas, C. et al. Benthic suspension feeders, key players in Antarctic marine ecosystems? Contrib Sci 1, 299–311 (2000).
Salomidi, M., Smith, C., Katsanevakis, S., Panayotidis, P. & Papathanassiou, V. Some observations on the structure and distribution of gorgonian assemblages in the eastern Mediterranean Sea in Proceedings of the 1st Mediterranean symposium on the conservation of the coralligenous and other calcareous bio-concretions 242–245 (2009).
Grinyó, J. et al. Diversity, distribution and population size structure of deep Mediterranean gorgonian assemblages (Menorca Channel, Western Mediterranean Sea). Prog Oceanogr 145, 42–56 (2016).
Matsumoto, A. K., Iwase, F., Imahara, Y. & Namikawa, H. Bathymetric distribution and biodiversity of cold-water octocorals (Coelenterata: Octocorallia) in Sagami Bay and adjacent waters of Japan. Bull Mar Sci 81, 231–251 (2007).
Stone, R. P., Masuda, M. M. & Karinen, J. F. Assessing the ecological importance of red tree coral thickets in the eastern Gulf of Alaska. ICES J Mar Sci 72, 900–915 (2014).
Mortensen, P. B. & Buhl-Mortensen, L. Distribution of deep-water gorgonian corals in relation to benthic habitat features in the Northeast Channel (Atlantic Canada). Mar Biol 144, 1223–1238 (2004).
Zapata-Guardiola, R. & López-González, P. J. Two new species of Antarctic gorgonians (Octocorallia: Primnoidae) with a redescription of Thouarella laxa Versluys, 1906. Helgoland Mar Res 64, 169–180 (2009).
Zapata-Guardiola, R. & López-González, P. J. Two new gorgonian genera (Octocorallia: Primnoidae) from Southern Ocean waters. Polar Biol 33, 313–320 (2010).
Orejas, C., Gili, J. M., López-González, P. J. & Arntz, W. E. Feeding strategies and diet composition of four Antarctic cnidarian species. Polar Biol 24, 620–627 (2001).
Orejas, C., Gili, J. M. & Arntz, W. Role of small-plankton communities in the diet of two Antarctic octocorals (Primnoisis antarctica and Primnoella sp.). Mar Ecol Prog Ser 250, 105–116 (2003).
Peck, L. S. & Brockington, S. Growth of the Antarctic octocoral Primnoella scotiae and predation by the anemone Dactylanthus Antarcticus. Deep Sea Res II 92, 73–78 (2013).
Martínez-Dios, A., Dominguez-Carrió, C., Zapata-Guardiola, R. & Gili, J. M. New insights on Antarctic gorgonians’ age, growth and their potential as paleorecords. Deep Sea Res I 112, 57–67 (2016).
Orejas, C. et al. Distribution and reproductive ecology of the Antarctic octocoral Ainigmaptilon antarcticum in the Weddell Sea. Mar Ecol Prog Ser 231, 101–114 (2002).
Orejas, C., Gili, J. M., López-González, P. J., Hasemann, C. & Arntz, W. E. Reproduction patterns of four Antarctic octocorals in the Weddell Sea: an inter-specific, shape, and latitudinal comparison. Mar Biol 150(4), 551–563 (2007).
Starmans, A., Gutt, J. & Arntz, W. E. Mega-epibenthic communities in Arctic and Antarctic shelf areas. Mar Biol 135, 269–280 (1999).
Dueñas, L. F. et al. The Antarctic Circumpolar Current as a diversification trigger for deep-sea octocorals. BMC evolutionary biol 16, 2 (2016).
Gori, A. et al. Spatial distribution patterns of the gorgonians Eunicella singularis, Paramuricea clavata and Leptogorgia sarmentosa (Cap of Creus, Northwestern Mediterranean Sea). Mar Biol 158, 143–158 (2011a).
Orejas, C. et al. Cold-water corals in the Cap de Creus canyon, northwestern Mediterranean: spatial distribution, density and anthropogenic impact. Mar Ecol Prog Ser 397, 37–51 (2009).
Bo, M. et al. Characteristics of a black coral meadow in the twilight zone of the central Mediterranean Sea. Mar Ecol Prog Ser 397, 53–61 (2009).
Rooney, J. et al. Mesophotic coral ecosystems in the Hawaiian Archipelago. Coral Reefs 29, 361–367 (2010).
Gutt, J. Antarctic macro-zoobenthic communities: a review and an ecological classification. Antarct Sci 19, 165–182 (2007).
Ludvigsen, M., Sortland, B., Johnsen, G. & Singh, H. Applications of geo-referenced underwater photo mosaics in marine biology and archaeology. Oceanography 20, 140–149 (2007).
Thurston, M. H., Bett, B. J., Rice, A. L. & Jackson, P. A. B. Variations in the invertebrate abyssal megafauna in the North Atlantic Ocean. Deep Sea Res I 41, 1321–1348 (1994).
Arnaud, P. M. et al. Semi-quantitative study of macrobenthic fauna in the region of the South Shetland Islands and the Antarctic Peninsula. Polar Biol 19, 160–166 (1998).
Gerdes, D., Klages, M., Arntz, W. E., Galeron, J. & Hain, S. Quantitative investigations on macrobenthos communities of the southeastern Weddell Sea shelf based on multibox corer samples. Polar Biol 12, 291–301 (1992).
Piepenburg, D., Voß, J. & Gutt, J. Assemblages of sea stars (Echinodermata: Asteroidea) and brittle stars (Echinodermata: Ophiuroidea) in the Weddell Sea (Antarctica) and off Northeast Greenland (Arctic): A comparison of diversity and abundance. Polar Biol 17, 305–322 (1997).
Teixidó, N., Rossi, S. & López-González, P. J. A unique assemblage of epibenthic sessile suspension feeders with archaic features in the high-Antarctic. Deep Sea Res II 53, 1029–1052 (2006).
Gili, J. M. & Coma, R. Benthic suspension feeders: their paramount role in littoral marine food webs. Trends Ecol Evol 13, 316–321 (1998).
Fortin, M. J. & Dale, M. R. T. Spatial analysis: a guide for ecologists. 365 (Cambridge University Press, Cambridge 2005).
Opresko, D. Abundance and distribution of shallow-water gorgonians in the area of Miami, Florida. Bull Mar Sci 23, 535–558 (1973).
Raguá-Gil, J. M., Gutt, J., Clarke, A. & Arntz, W. E. Antarctic shallow-water mega-epibenthos: shaped by circumpolar dispersion or local conditions? Mar Biol 144, 829–839 (2004).
Stone, R. P. Coral habitat in the Aleutian Islands of Alaska: depth distribution, fine scale species associations, and fisheries interactions. Coral Reefs 25, 229–238 (2006).
Clark, M. R. & Bowden, D. A. Seamount biodiversity: high variability both within and between seamounts in the Ross Sea region of Antarctica. Hydrobiol 761, 161–180 (2015).
Grigg, R. W. Population dynamics of tow gorgonian corals. Ecology 58, 278–290 (1977).
Cúrdia, J. et al. Spatial and depth-associated distribution patterns of shallow gorgonians in the Algarve coast (Portugal, NE Atlantic). Helgoland Mar Res 67, 521–534 (2013).
Lasker, H. R. & Coffroth, M. A. Octocoral distributions at Carrie Bow Cay, Belize. Mar Ecol Prog Ser 13, 21–28 (1983).
Yoshioka, P. M. & Yoshioka, B. B. Effects of wave energy, topographic relief and sediment transport on the distribution of shallow-water gorgonians of Puerto Rico. Coral Reefs 8, 145–152 (1989).
Arntz, W., Brey, T. & Gallardo, V. A. Antarctic zoobenthos. Oceanogr Mar Biol 32, 241–304 (1994).
Convey, P. et al. The spatial structure of Antarctic biodiversity. Ecol Monogr 84(2), 203–244 (2014).
Brey, T. & Clarke, A. Population dynamics of marine benthic invertebrates in Antarctic and subantarctic environments: are there unique adaptations? Antarct Sci 5, 253–266 (1993).
Gilmour, J. P. Size-structures of populations of the mushroom coral Fungia fungites: the role of disturbance. Coral Reefs 23, 493–504 (2004).
Gori, A. et al. Size and spatial structure in deep versus shallow populations of the Mediterranean gorgonian Eunicella singularis (Cap de Creus, northwestern Mediterranean Sea). Mar Biol 158, 1721–1732 (2011).
Meesters, E. H., Hilterman, M., Kardinaal, E., Keetman, M. & Bak, R. P. M. Colony size-frequency distributions of scleractinian coral populations: spatial and interspecific variation. Mar Ecol Prog Ser 209, 43–54 (2001).
Linares, C., Coma, R., Garrabou, J., Díaz, D. & Zabala, M. Size distribution, density and disturbance in two Mediterranean gorgonians: Paramuricea clavata and Eunicella singularis. J App Ecol 45, 688–699 (2008).
Bak, R. P. M. & Meesters, E. H. Coral population structure: the hidden information of colony size-frequency distributions. Mar Ecol Prog Ser 162, 301–306 (1998).
Coma, R., Ribes, M., Zabala, M. & Gili, J. M. Growth in a modular colonial marine invertebrate. Estuar Coast Shelf S 47, 459–470 (1998).
Sherwood, O. A. & Edinger, E. N. Ages and growth rates of some deep-sea gorgonian and antipatharian corals of Newfoundland and Labrador. Can J Fish Aquat Sci 66, 142–152 (2009).
Gutt, J., Starmans, A. & Dieckmann, G. Impact of iceberg scouring on polar benthic habitats. Mar Ecol Prog Ser 137, 311–316 (1996).
Teixidó, N., Garrabou, J., Gutt, J. & Arntz, W. E. Recovery in Antarctic benthos after iceberg disturbance: trends in benthic composition, abundance and growth forms. Mar Ecol Prog Ser 278, 1–16 (2004).
Kahng, S. E., Benayahu, Y. & Lasker, H. R. Sexual reproduction in octocorals. Mar Ecol Prog Ser 443, 265–83 (2011).
Thiem, Ø., Ravagnan, E., Fosså, J. H. & Berntsen, J. Food supply mechanisms for cold-water corals along a continental shelf edge. J Mar Syst 60, 207–219 (2006).
Genin, A., Dayton, P. K., Lonsdale, P. F. & Spiess, F. N. Coral on seamount peaks provide evidence of current acceleration over deep-sea topography. Nature 322, 59–61 (1986).
Rogers, A. D. The biology of seamounts. Adv Mar Biol 30, 305–350 (1994).
Mortensen, P. B., Buhl-Mortensen, L., Gebruk, A. V. & Krylova, E. M. Occurrence of deep-water corals on the Mid-Atlantic Ridge based on MAR-ECO data. Deep Sea Res II 55, 142–152 (2008).
Knust, R. & Schröder, M. The Expedition PS82 of the Research Vessel POLARSTERN to the southern Weddell Sea in 2013/2014. Berichte zur Polar und Meeresforschung. Reports on polar and marine research. pp. 680 (2014).
Vaughan, D. G. et al. Recent rapid regional climate warming on the Antarctic Peninsula. Climatic change 60(3), 243–274 (2003).
Ripley, B. D. The second order analysis of stationary point processes. J Appl Probab 13, 255–266 (1976).
Haase, P., Pugnaire, F. I., Clark, S. C. & Incoll, L. D. Spatial patterns in a two-tiered semi-arid shrubland in southeastern Spain. J Veg Sci 7, 527–534 (1996).
Schols, P. & Lorson, D. Macnification. Orbicule, Leuven, Belgium, http://www.orbicule.com/ (2008).
Clarke, K. R. & Warwick, R. M. Change in Marine Communities: and approach to statistical analysis and interpretation. Natural Environment Research Council, United Kingdom (1994).
Anderson, M. J. A new method for non-parametric multivariate analysis of variance. Austral Ecol 26, 32–46 (2001).
Arndt, J. E. et al. The International Bathymetric Chart of the Southern Ocean (IBCSO) Version 1.0 - A new bathymetric compilation covering circum-Antarctic waters. Geophys Res Lett 40, 3111–3117 (2013).
Acknowledgements
We are grateful to the captain and crew of R/V Polarstern cruise PS82 (ANT-XXIX/9) and AWI for their technical and logistical support. We thank Nils Owsianowski for engineering and for piloting the ROV, and Anna Kluibenschedl, Santiago Pineda Metz and Pilar Casado de Amezua for their help during ROV deployments. This research was partially funded by ECOWED Project (CTM2012-39350-C02-01), PACES I 1.6, PACES II 1.6 and Polarstern Expedition PS82 with grant no. AWI_PS82_03.
Author information
Authors and Affiliations
Contributions
J.M.G. and N.T. conceived the research. R.Z.G. identified the species. L.F. and C.R. led the R.O.V. surveys and provided the video material, S.A. assisted in the surveys. J.S. and S.A. analysed the videos and treated the data. S.A. and N.T. wrote the manuscript. All authors revised the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ambroso, S., Salazar, J., Zapata-Guardiola, R. et al. Pristine populations of habitat-forming gorgonian species on the Antarctic continental shelf. Sci Rep 7, 12251 (2017). https://doi.org/10.1038/s41598-017-12427-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-12427-y
- Springer Nature Limited
This article is cited by
-
Are there distribution patterns and population structure differences among demersal fish species in relation to Antarctic benthic communities? A case study in the Weddell Sea
Polar Biology (2023)
-
Macroscopic characteristics facilitate identification of common Antarctic glass sponges (Porifera, Hexactinellida, Rossellidae)
Polar Biology (2020)
-
The importance of coastal gorgonians in the blue carbon budget
Scientific Reports (2019)