
Abstract
Although numerous studies have revealed significant associations between variants in the nicotinic acetylcholine receptors (nAChR) subunits and nicotine dependence (ND), only few studies were performed in Chinese subjects. Here, we performed association and interaction analysis for 20 single nucleotide polymorphisms (SNPs) in the CHRNB3-CHRNA6 gene cluster with ND in a Chinese Han population (N = 5,055). We found nominally significant associations for all tested SNPs with ND measured by the Fagerström Test for Nicotine Dependence score; of these, 11 SNPs remained significant after Bonferroni correction for multiple tests (p = 9 × 10−4~2 × 10−3). Further conditional analysis indicated that no other SNP was significantly associated with ND independent of the most-highly significant SNP, rs6474414. Also, our haplotype-based association analysis indicated that each haplotype block was significantly associated with ND (p < 0.01). Further, we provide the first evidence of the genetic interaction of these two genes in affecting ND in this sample with an empirical p-value of 0.0015. Finally, our meta-analysis of samples with Asian and European origins for five SNPs in CHRNB3 showed significant associations with ND, with p-values ranging from 6.86 × 10−14 for rs13280604 to 6.50 × 10−8 for rs4950. This represents the first study showing that CHRNB3/A6 are highly associated with ND in a large Chinese Han sample.
Similar content being viewed by others


Introduction
Tobacco use continues to be a leading cause of preventable morbidity and death worldwide1. The World Health Organization predicts that if the current smoking trend continues, the annual number of deaths from smoking-related diseases will double to 10 million in 20202, 3. Because of the effect of legislation against tobacco smoking and public realization of its health consequences, the prevalence of cigarette smoking has declined in the United States and other developed countries4. On the other hand, despite heightened public awareness of the negative health consequences, smoking remains prevalent or has even increased in many developing countries5, especially in Asian nations such as China. The smoking rate in Chinese men aged 15 or older has been estimated to be 66%6, implying that more than 350 million Chinese men smoke tobacco, almost one third of the total smokers in the world7.
Cigarette smoking, initiation, nicotine dependence (ND) and cessation8, 9, are complex and multifactorial behaviors determined by genetic and environmental factors, as well as their interactions. Evidence from twin studies indicates that the mean heritability of ND is 0.56 in total and 0.59 for male smokers10.
Neuronal nicotinic acetylcholine receptors (nAChRs), which are ligand-gated ion channels consisting of five subunits, are the primary targets for nicotine to initiate the brain responses to smoking. Accordingly, it is biologically plausible that the genes encoding these nAChR subunits influence smoking-related behaviors such as initiation, intensity, and ND. During recent years, considerable efforts, especially genome-wide association studies (GWAS), have been conducted to identify genetic factors underlying ND, with several widely accepted successes, including the identification of the importance of AChR subunit gene clusters on chromosomes 15 (CHRNA5/A3/B4)11,12,13,14,15,16 and 8 (CHRNB3/A6)17,18,19,20 and the genes encoding nicotine-metabolizing enzymes on chromosome 19 (CYP2A6/A7)19, 21,22,23. As of October 2015, nine published GWAS and meta-GWASs have yielded 11 genetic loci carrying linked variants (p < 5 × 10−8) associated with relevant ND phenotypes16, 19, 21, 24,25,26,27,28,29. Among these, three genome-wide significant single nucleotide polymorphisms (SNPs) on chromosome 8p1119, 27 (rs13280604, rs6474412, and rs1451240, in perfect linkage disequilibrium; LD) in samples of European and African ancestries have been suggestively associated with the ND status of smokers30. Although the dichotomized Fagerström Test for Nicotine Dependence (FTND) appears to have an equivalent relation with rs1451240 across ethnicities, the relation between this SNP and the number of cigarettes smoked per day (CPD) is much weaker in African Americans (AAs) than in European Americans (EAs)27. The other two SNPs were both significantly associated with CPD in European samples19.
The CHRNA6 and CHRNB3 genes are located contiguously in a tail-to-tail configuration on chromosome 8p11. The protein products of the two subunit genes are co-localized in nAChRs in the substantia nigra, ventral tegmental area, striatum, and locus coeruleus31. Previous candidate gene-based association studies have found that SNPs in the CHRNB3-CHRNA6 cluster are significantly associated with the risk of ND and various tobacco-use behaviors18, 30, 32,33,34,35,36,37,38. For detailed information, please refer to our recent published reviews on this gene family39,40,41.
Although current findings have greatly improved our understanding of the etiology of ND, much is still unknown. In particular, there arises a critical question whether the identified associations are reproducible across different ancestral populations. Considering that nearly all subjects used in GWAS or candidate gene-based association studies were of European origin, it would be interesting to know whether SNPs in CHRNA6 and CHRNB3 are also involved in ND in smokers of other ethnicities. Given the demonstrated differences across ethnicities with respect to genetics, physiological processes, and behavior underlying ND42,43,44,45, it is necessary to conduct this kind of work with samples of other ethnicities, such as Chinese smokers, as we report in this study.
Materials and Methods
Subjects
We recruited Han Chinese participants aged 20 years and older from several communities in the cities of Jincheng and Taiyuan, Shanxi Province in 2013. Because few females (~4%) smoke tobacco in China, only males (n = 5055) were included in the study. A structured questionnaire on smoking behavior, demographics, medical history, lifestyle, environment, and dietary characteristics was administered to each subject by trained researchers in local hospitals of the two areas. Individuals with other psychiatric diagnoses, such as major depression or schizophrenia, were excluded. The subjects were biologically unrelated to each other. After a detailed explanation of the project and process of this study, informed written consent was obtained from each participant. The Institutional Review Board of the First Affiliated Hospital of Zhejiang University School had approved the project and all questionnaires used for it, and all experiments were performed in accordance with relevant guidelines and regulations of the institution. Table 1 and Supplementary Figure 1 provides the detailed characteristics of the subjects. We defined non-smokers as persons having smoked fewer than 100 cigarettes during their lifetimes, while smokers were assessed with the FTND46, 47, one of the commonly used measures of ND (0–10 scale).
SNP selection and genotyping
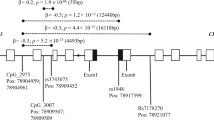
Twenty SNPs in the CHRNB3-CHRNA6 cluster (CHRNB3: rs10958725, rs10958726, rs13273442, rs4736835, rs1955186, rs6474413, rs7004381, rs4950, rs1530848, rs13280604, rs6474414, rs6474415, rs4954; CHRNA6: rs9298629, rs2304297, rs892413, rs10087172, rs2196128, rs2217732, rs1072003) were selected using the SNP Browser software from Applied Biosystems (http://www.appliedbiosystems.com) by searching dbSNP (http://www.ncbi.nlm.nih.gov/SNP/) and other published papers17, 18, 34, 35, 37, 48,49,50. The SNPs were chosen to span the two genes and to have moderate minor allele frequencies (MAF ≥ 5% in CHB (Chinese Han in Beijing) based on the NCBI SNP database) and to assay reliably (Supplementary Table 1; Fig. 1).

Schematic diagram of the human CHRNB3-CHRNA6 cluster for selective SNPs (not drawn to scale). Horizontal black arrows indicate the direction of transcription of each gene. Gray and black rectangles indicate exons separated by intronic regions and untranslated regions, respectively. The 20 SNPs that were genotyped are shown by vertical arrows, with their rsID and gene locations indicated.
Genomic DNA was extracted from the peripheral blood of each participant using the Qiagen DNA purification kit according to the manufacturer’s instructions. The concentration of DNA in each sample was determined by the optical density (OD) at 260 nm, and the quality was evaluated by the OD 260/280 ratio. SNP genotyping was performed by using the TaqMan OpenArray Genotyping Platform (Applied Biosystems, Inc.). It requires a pair of primers common to both wild-type and mutant sequences, as well as two minor groove binder probes, one being a probe for the normal sequence, labeled with the VIC fluorophore, and the other being a probe for the mutant sequence, labeled with the FAM fluorophore. For the samples with an A260/280 ratio between 1.7 and 2.0 and a DNA concentration of 50 ng/μl, about 2 μl (ca. 100 ng) of each genomic sample was placed in a well of a 384-well plate, together with 2 μl of OpenArray Master Mix, using a distribution pattern according to the manufacturer’s instructions. The sealed plate was then centrifuged briefly at 1000 rpm to remove any bubbles. The samples, tips, and the OpenArray plate (composed of 96 sub-arrays, each with 64 nano-wells) were placed in the AccuFill system, and the plates were customized to provide an even distribution. A total volume of 33 μl was dispensed into each nano-well. After the plates had been stabilized with the immersion fluid in a glass case, they were sealed with a photosensitive gum for 2 min under ultraviolet light in the sealing station. The amplification reaction was performed in the Dual Flat Block GeneAmp PCR System 9700 thermal cycler, a process that lasted approximately 4 h. The plates were then placed in an OpenArray NT Cycler for detection of the emitted fluorescence. After capture of the signal, the results were imported into TaqMan Genotyper software for SNP calling.
Quality control (QC) tests were performed on each DNA sample, with the goal of excluding samples with sex or chromosomal anomalies, a low call rate, or first- or second- degree relatedness. With this QC step, we removed 334 samples with a call rate of <95%. The genotyping rate for all tested SNPs was >95%, and the p value for the Hardy-Weinberg equilibrium was set to >0.05.
Statistical analysis
Individual SNP-based association analysis
For individual SNP-based association analysis, we used the PLINK program (v. 1.07)51 to perform linear regression for FTND score under an additive genetic model with age, body mass index (BMI), miner (whether the subject had worked as a miner or not), work years, and number of family members who smoke as covariates. We also performed association analysis by excluding “number of family members who smoke” as a covariate from the analysis, which yielded almost the same association results (data not shown). Statistically significant results (p < 0.05) for individual SNPs were adjusted for multiple testing using Bonferroni correction.
Haplotype-based association analysis
Pair-wise LD and haplotype blocks were assessed by Haploview (v. 4.2)52, 53, and their associations with the FTND score were analyzed using Haplo Stats (v. 1.6.8) through computing score statistics with the same covariates and genetic models used in the individual SNP-based association analysis54. The significance of each haplotype was adjusted by the number of major haplotypes (frequency > 5%) included in the analysis, a strategy used to find out if detected associations represent the same signal or not, as we did previously15, 37, 55.
Gene-gene interaction analysis
To further determine the genetic contribution of the CHRNB3-CHRNA6 cluster to ND, we performed a gene–gene interaction analysis for the total sample as we did previously for nAChR genes CHRNA5/A3/B4 14, 15, CHRNA4, and CHRNB256. We employed the GMDR-GPU program, which was developed by our group57, for the SNP-by-SNP interaction analysis of CHRNB3 and CHRNA6 with exhaustive searches by two- to five-way SNP combinations. The GMDR-GPU is an improved version of the original GMDR program58 that can run much faster because of the integration of more efficient computational algorithms and capacities. Similar to the association analysis described above, GMDR-GPU calculated a score statistic for each subject based on a generalized linear model under an assumption of normal distribution of the FTND scores. We used the following three criteria to determine the best statistical gene–gene interaction model: the cross-validation consistency (CVC), the prediction accuracy, and the significant p value, which was determined by 107 permutation tests for the selected SNP combinations.
Meta-analysis of individual SNP-based association with ND
We searched PubMed and Web of Science for all studies eligible for this meta-analysis according to the PRISMA guidelines59, a strategy similar to what we used previously60, 61. The key words used in the search were “CHRNB3” or “CHRNA6” or “cholinergic nicotinic receptor gene” combined with “nicotine dependence” or “ND.” The reports containing the chosen words were checked individually for references that could have been missed by the above-mentioned search protocol. The inclusion criteria for meta-analysis were as follows. First, the study presented sufficient information for sample size, effect size of the tested allele and p values. Also, each study had to be independent, with no duplication or overlap. To combine the published data with the association results from this study, we meta-analyzed each SNP individually under the random-effects model by METAL62. Heterogeneity among different ethnic samples was assessed withI 2 in METAL, which measures the degree of inconsistency among the results from different samples63.
Results
Individual SNP-based association analysis
As shown in Table 2, the MAF and the β of the SNPs tested for association with ND were almost the same for the Jincheng and Taiyuan samples. Because of the lack of heterogeneity between the samples from the two sites on the basis of principal component analysis (PCA), we calculated the linear regression of each SNP using the total sample with an additional covariate “site.” Of the 20 SNPs analyzed, 10 in CHRNB3 and 1 in CHRNA6 showed significant association with FTND after Bonferroni correction for multiple testing by SNP number, with the p value for rs6474414 being the smallest one. Further, we performed conditional analyses on rs6474414, which revealed no other SNPs that are significantly associated with ND.
Haplotype-based association analysis
Haplotype-based association analysis was performed for all SNPs in the CHRNB3-CHRNA6 cluster, regardless of the individual SNP-based association results. Similar to the results of the individual SNP-based association studies, we revealed no heterogeneity for the haplotype patterns detected at the two sites. According to the haplotype block criteria defined by Gabriel et al., only one block (D′ > 0.97) of CHRNB3 as well as CHRNA6 was identified with the pooled samples, and each site-specific sample (Fig. 2) appeared only once for the same block. Because the major haplotypes detected in each block were the same for the two sites, we present the association results for pooled samples.

LD structure for CHRNB3 and CHRNA6 SNPs in the pooled samples (Jincheng and Taiyuan). Haplotype blocks were defined according to Gabriel et al.52. The number in each box represents the r 2 value for each SNP pair as calculated by Haplotype (v. 4.2). The arrow on top of the figure represents the gene transcription direction from 5′ to 3′.
In CHRNB3, one haplotype of T-G-A-T-G-C-A-G-G-G-A-G, formed by SNPs rs10958725, rs10958726, rs13273442, rs4736835, rs1955186, rs6474413, rs7004381, rs4950, rs1530848, rs13280604, rs6474414, and rs6474415, with a frequency of 21.8%, was significantly associated with FTND (Hap Score = −2.8; p = 0.005) (Table 3). In CHRNA6, we found two significant haplotypes, T-C-A-C-C-G-G and G-G-C-T-T-A-C, with a frequency of 22.9% and 76.2%, respectively, formed by the SNPs rs9298629, rs2304297, rs892413, rs10087172, rs2196128, rs2217732, and rs1072003. These two associations remained significant after Bonferroni correction for multiple testing (Hap Scores = −2.8 and 2.6; p = 0.006 and 0.009, respectively) (Table 3).
Gene–gene interaction analysis
Based on the fact that CHRNB3, which is adjacent to CHRNA6 (Fig. 1), usually is co-expressed with α6, immunoprecipitation and high-affinity [125I] α-conotoxinMII (αCtxMII)-binding studies showed that the α6β3* pentamer was the predominant α6*-nAChRs in the striatum64, 65. The accessory subunit, β3, has an essential role in α6*-nAChRs biogenesis and function66, 67. Thus, there might be epistatic effects in ND between CHRNB3 and CHRNA6. Consistent with this hypothesis, we found the best interaction model of 5 SNPs in two genes (rs10958725 and rs1955186 in CHRNB3; rs10087172, rs2196128 and rs1072003 in CHRNA6), showing a CVC of 10/10, had a prediction accuracy of 52.71% and an empirical p value from 107 permutations of 0.0015 (Table 4).
Meta-analysis of individual SNP-based association with ND
Table 5 presents the association analysis results of each SNP among all samples, which include minor allele, β value, SE, and p. Because of the high heterogeneity between the ancestries of African and European or Asian (i.e., minor alleles of all five SNPs in African population were opposite to those in the European and Asian populations), we excluded AAs from our meta-analysis. The four samples included in this study were from: (1) the Chinese sample used in the current study; (2) the Korea Association Resource (KARE) study68; (3) European Americans (EAs) from MSTCC20, SAGE69, and CGEMS70; and (4) other EAs from the ENGAGE, TAG, and Ox-GSK consortia19. Because no FTND score was available for a few samples, the ND phenotype was measured by CPD15. Meta-analysis was performed for the five SNPs by combining the results from the included studies. Of the meta-analyzed SNPs, rs13280604 in CHRNB3 (p = 6.86 × 10−14) showed the strongest association with ND.
Discussion
In the current study, we examined genetic associations and epistatic effects in CHRNB3-CHRNA6 linked to ND in the Chinese Han population. Individual SNP-based association analyses revealed that 11 SNPs in this region showed a significant association with the FTND score after Bonferroni correction for multiple testing. Of these polymorphisms, rs6474414 in CHRNB3 showed the strongest association (p = 0.0009) in the pooled sample. Then we found one haplotype in CHRNB3 (p = 0.005) and two haplotypes in CHRNA6 (p = 0.006 and 0.009) that were associated significantly with FTND in the total sample. Furthermore, considering the fact that the protein products of these nAChR subunit genes must join in order to form functional nAChRs, we also performed gene–gene interaction analysis on all SNPs in this genomic region and found that the best interactive model involved 5 SNPs of the CHRNB3-CHRNA6 cluster. Finally, by conducting meta-analysis of published samples with Asian and European ancestry, we obtained confirmation of the previous findings and further promoted the genetic signals linking CHRNB3 with ND.
There are several highlights of this study. First, although both GWAS19, 30 and candidate gene-based association studies18, 32, 37 have implicated variants in the CHRNB3-CHRNA6 cluster in the development of ND, almost all subjects used in these studies were of European ancestry. Meanwhile, the reported largest GWAS of smoking with the African American samples did not identify this locus to be significant26. Given the known ethnic differences in physiological processes and behavior underlying ND42,43,44,45, 50, it is of great interest to determine whether this gene cluster is associated with ND in smokers of other populations, especially Asians. To attack this issue, we conducted the present study designed to investigate the association of candidate genes with ND in a Chinese Han population. To the best of our knowledge, this is the first genetic study of ND considering FTND score as a quantitative trait in a large Chinese sample.
Second, we provided additional support for a role of the CHRNB3-CHRNA6 cluster in mediating ND by meta-analyzing samples of smokers with different ethnicities, based on the hypothesis that the biological mechanisms underlying a disease of interest are shared by various human populations. There were multiple SNPs significantly associated with a decreased risk of tobacco addiction after Bonferroni correction in CHRNB3 (p value from 9 × 10−4 for rs6474414 to 2 × 10−3 for rs7004381). Of these, rs10958726 and rs13280604 achieved genome-wide significance (p < 5 × 10−8) for CPD in the combined analysis of more than 80,000 individuals19. Furthermore, these two variants participated in the subjective responses to initial tobacco use32, 36, the transition from occasional cigarette smoking to ND18, 27, and the number of quit attempts3. However, either none or nominal significance was identified regarding the association of these two variants with ND in African-American and Korean samples17, 71. By extracting data from other reports, we performed a meta-analysis with samples of Asian and European ancestry. All alleles with negative effects for five SNPs in CHRNB3 were the same in both ethnic populations. Significant association of ND with CHRNB3 was present in these ethnic samples, with p values ranging from 6.86 × 10−14 for rs13280604 to 6.50 × 10−8 for rs4950. Four SNPs reached genome-wide significance, providing convincing evidence of a role of CHRNB3 in ND by analyzing samples of two representative ancestries from throughout the world. Our meta-analysis results indicates that this region is homogeneous across populations, implying any causative variants identified in this gene could be important for almost all smokers, regardless of ancestry, except for AAs.
Third, we not only conducted association analyses of this region with ND at the SNP and haplotype levels, as in most reported studies of the association of the region with smoking, but also performed gene–gene interaction analysis exclusively in this region. As CHRNB3 and CHRNA6 lie tail to tail on chromosome 8 and are both components of nAChRs, it is biologically plausible that they function together to influence the etiology of ND. In addition, there are not many loci in CHRNA6 found to be significantly associated with smoking behavior so far, in contrast to the abundant results of the gene from in vivo studies (see below for details). One possible mechanism is the interaction between common variants within the CHRNB3-CHRNA6 cluster, similar to our previously detected interactions between CHRNA4 and CHRNB2 and CHRNA5/A3/B414, 56. In accordance with this hypothesis, we detected two loci in CHRNB3 and three loci in CHRNA6 that contributed interactively to FTND in our sample. This is the first evidence of an epistatic effect of these two genes discovered through genetic analysis in any ethnic sample. Statistically, interaction analysis within genes or within multigene regions is necessary to explore the mechanism of the observed association of common variants with complex traits.
Several functional studies by genetic manipulation in rodents have revealed the vital role of the α6 subunit in the establishment of ND despite the absence of evidence at the SNP-based association analysis level72,73,74. For example, α6 wild type (WT) mice self-administered nicotine in a unit dose of 26.3 μg/kg per infusion, whereas their α6 knockout (KO) drug-naive littermates did not. The α6-KO animals did not self-administer nicotine even in an extensive range of lower (8.7–17.5 μg/kg) and higher (35–52.6 μg/kg) doses. Importantly, when the α6 subunit was selectively re-expressed in the VTA of α6−/− mice using a lentiviral vector, the reinforcement property of nicotine was restored75. However, most genetic studies tested CHRNB3-CHRNA6 together in which significant association was more likely attributable to variants in CHRNB3. Moreover, the association of CHRNA6 SNPs typically did not survive correction for multiple testing32, 35. In the present study, we demonstrated that rs1072003 in CHRNA6 was still significantly associated with FTND score after Bonferroni correction (p = 0.002). To the contrary, Saccone et al.17, 35 did not find evidence of an association between this SNP and ND defined as an FTND score ≥ 4 in EA or AA samples, and neither did Zeiger et al.32 nor Ehringer et al.36 using subjective responses to tobacco as phenotypes.
By using an in vitro culture approach, Ehringer et al.36 conducted a luciferase reporter assay study for different haplotypes in the CHRNB3 promoter. Data from these experiments indicated that different alleles in the CHRNB3 upstream promoter region led to different degrees of RNA expression. Rs4950 dramatically diminished the binding capacity of a specific transcription factor, Oct-1, and greatly reduced the activity of the promoter of CHRNB3 76. This finding is consistent with the speculation that this SNP might have an effect on the expression of CHRNB3, based on prediction by the Transcriptional Element Search System (TESS), a publicly available program3, 32. Kamens et al.77 illustrated that the protective genetic variant at rs6474413, but not rs4950, reduced gene expression and decreased β3 gene expression in mice and reduced nicotine intake. This kind of work increases our understanding of the molecular mechanisms that contribute to the human genetic associations of tobacco use behaviors, making such work highly important.
One or more differences in study design, ascertainment criteria, assessment of ND, demographic variables, or sample size could contribute to differences in the association of SNPs in the nAChR genes with smoking behaviors. For example, in a Han Chinese sample, Wei et al.49 did not find any significant locus in the CHRNB3 gene except for rs4954. Although it is difficult for us to explain such a difference, we suspect that small sample size (i.e., 223 cases and 257 controls) used in their study might be one of the main reasons for them to detect any potential significant association. Furthermore, Wei and his associates classified their smokers into low ND (LND; FTND ≤ 3) and high ND (HND; FTND ≥ 6) groups, which differed from the definition of smoking used in our study in which we treated FTND as a quantitative trait. Furthermore, we genotyped more SNPs in the CHRNB3-CHRNA6 region in comparison of number of SNPs included in their study on this region. More importantly, we performed not only individual SNP- and haplotype-based association analysis but also interaction analysis among the variants within this CHRNB3-CHRNA6 region. Nevertheless, because of the high LD of the SNPs in each gene examined in our study, it remains to be determined which SNP indeed contributes to the significant association signal. As a matter of fact, SNPs involved in the present study are either intronic or outside the translational region of the gene cluster; thus, it is less clear whether one of these SNPs is the functional variant(s) responsible for the observed association. More effort is needed to find causal variant(s) in this region.
To improve detection power and reduce the possibility of population substructure which might influence our association findings, we recruited all our subjects from the same geographic areas and required all enrolled subjects to be Han Chinese and free of other diagnosed psychiatric disorders or using other substances. Using the subjects recruited from the same geographic areas imply that they are more homogeneous and have similar genetic background according to a recent reported study showing that the genetic clustering of Han Chinese is closely related to their geographic location78. Because majority of smokers are males in China and many other Asian countries79, we only performed our genetic analysis of variants in this cluster with ND in male smokers. We wish that we could not do similar genetic analysis for female smokers as well, which can be considered a potential limitation of this study. Another potential limitation of this study is that we did not genotype as many SNPs as we wish for this region including failed to genotype one of three SNPs with genome-wide significance (i.e., rs1451240). To have better coverage for these samples, more SNPs with a high density are needed for further analysis of this gene cluster in relation to ND. Furthermore, we identified only one block each in the CHRNB3 as well as in CHRNA6, and found that major haplotypes in each of the above-mentioned LD blocks were significantly associated with the FTND score. However, as reported by Saccone et al.17, there might be at least two distinct loci for ND in CHRNB3-CHRNA6. The first one appears to be driven by rs13277254 and other highly correlated SNPs such as rs10958726 and rs1955186. The other one was tagged by rs4952, the only known coding SNP in exon 5 of CHRNB3. Because of the extremely low minor allele frequency of rs4952 in Asian populations, we did not detect this signal in our sample. Finally, we failed to find another independent Chinese smoker sample to replicate our detected SNP-SNP interaction results in this paper. Thus, it is important and necessary to replicate our reported gene-by-gene interaction results in future study with independent samples.
In sum, our results indicate that both CHRNB3 and CHRNA6 are significantly associated with ND in the Chinese Han population. We derived this conclusion by genetic analysis at both the individual SNP and haplotype level, which made these genes excellent subjects for research on the molecular mechanisms of ND. This study represents a significant extension of earlier reports in which most subjects were of European origin and which promoted the association signals of CHRNB3 by meta-analysis. Meanwhile, we also detected gene-by-gene interaction between CHRNB3 and CHRNA6 statistically, consistent with the hypothesis that these two genes function together in mediating tobacco addiction. A better understanding of the role of these genetic variants will be key for pharmacologic targeting to reduce or possibly eliminate some of the addictive properties of nicotine in susceptible individuals.
References
CDC. Current cigarette smoking among adults–United States,. MMWR Morb Mortal Wkly Rep 2012, 889–894 2011.
Warren, C. W., J., N., Eriksen, M. P. & Asma, S. Global Tobacco Surveillance System (GTSS) collaborative group. Patterns of global tobacco use in young people and implications for future chronic disease burden in adults. Lancet, 749–753 (2006).
Hoft, N. R. et al. Genetic association of the CHRNA6 and CHRNB3 genes with tobacco dependence in a nationally representative sample. Neuropsychopharmacology 34, 698–706, doi:10.1038/npp.2008.122 (2009).
Nguyen, K. H., Marshall, L., Brown, S. & Neff, L. State-Specific Prevalence of Current Cigarette Smoking and Smokeless Tobacco Use Among Adults - United States, 2014. MMWR Morb Mortal Wkly Rep 65, 1045–1051, doi:10.15585/mmwr.mm6539a1 (2016).
Benowitz, N. L. Clinical pharmacology of nicotine: implications for understanding, preventing, and treating tobacco addiction. Clin Pharmacol Ther 83, 531–541, doi:10.1038/clpt.2008.3 (2008).
MOHC. Smoking and health report of China 2006. (2006).
CCDC. Smoking control report of China 2009. (2009).
Breslau, N., Johnson, E. O., Hiripi, E. & Kessler, R. Nicotine dependence in the United States: prevalence, trends, and smoking persistence. Archives of general psychiatry 58, 810–816 (2001).
Johnson, E. O., Chase, G. A. & Breslau, N. Persistence of cigarette smoking: familial liability and the role of nicotine dependence. Addiction 97, 1063–1070 (2002).
Li, M. D., Cheng, R., Ma, J. Z. & Swan, G. E. A meta-analysis of estimated genetic and environmental effects on smoking behavior in male and female adult twins. Addiction 98, 23–31 (2003).
Berrettini, W. et al. Alpha-5/alpha-3 nicotinic receptor subunit alleles increase risk for heavy smoking. Mol Psychiatry 13, 368–373 (2008).
Bierut, L. J. et al. Variants in Nicotinic Receptors and Risk for Nicotine Dependence. Am J Psychiatry 165, 1163–1171 (2008).
Chen, L. S. et al. Smoking and genetic risk variation across populations of European, Asian, and African American ancestry–a meta-analysis of chromosome 15q25. Genet Epidemiol 36, 340–351, doi:10.1002/gepi.21627 (2012).
Li, M. D. et al. Association and interaction analysis of variants in CHRNA5/CHRNA3/CHRNB4 gene cluster with nicotine dependence in African and European Americans. Am J Med Genet B Neuropsychiatr Genet 153B, 745–756, doi:10.1002/ajmg.b.31043 (2010).
Wang, J. & Li, M. D. Common and unique biological pathways associated with smoking initiation/progression, nicotine dependence, and smoking cessation. Neuropsychopharmacology 35, 702–719, doi:10.1038/npp.2009.178 (2010).
Tobacco & Genetics, C. Genome-wide meta-analyses identify multiple loci associated with smoking behavior. Nat Genet 42, 441–447, doi:10.1038/ng.571 (2010).
Saccone, N. L. et al. Multiple cholinergic nicotinic receptor genes affect nicotine dependence risk in African and European Americans. Genes Brain Behav 9, 741–750, doi:10.1111/j.1601-183X.2010.00608.x (2010).
Saccone, S. F. et al. Cholinergic nicotinic receptor genes implicated in a nicotine dependence association study targeting 348 candidate genes with 3713 SNPs. Hum Mol Genet 16, 36–49 (2007).
Thorgeirsson, T. E. et al. Sequence variants at CHRNB3-CHRNA6 and CYP2A6 affect smoking behavior. Nat Genet 42, 448–453, doi:10.1038/ng.573 (2010).
Cui, W. Y. et al. Significant association of CHRNB3 variants with nicotine dependence in multiple ethnic populations. Mol Psychiatry 18, 1149–1151, doi:10.1038/mp.2012.190 (2013).
Kumasaka, N. et al. Haplotypes with copy number and single nucleotide polymorphisms in CYP2A6 locus are associated with smoking quantity in a Japanese population. PLoS One 7, e44507, doi:10.1371/journal.pone.0044507 (2012).
Chen, L. S. et al. Pharmacotherapy effects on smoking cessation vary with nicotine metabolism gene (CYP2A6). Addiction 109, 128–137, doi:10.1111/add.12353 (2014).
Bloom, A. J. et al. Variants in two adjacent genes, EGLN2 and CYP2A6, influence smoking behavior related to disease risk via different mechanisms. Hum Mol Genet 23, 555–561, doi:10.1093/hmg/ddt432 (2014).
Liu, J. Z. et al. Meta-analysis and imputation refines the association of 15q25 with smoking quantity. Nat Genet 42, 436–440, doi:10.1038/ng.572 (2010).
Thorgeirsson, T. E. et al. A variant associated with nicotine dependence, lung cancer and peripheral arterial disease. Nature 452, 638–642 (2008).
David, S. P. et al. Genome-wide meta-analyses of smoking behaviors in African Americans. Translational psychiatry 2, e119, doi:10.1038/tp.2012.41 (2012).
Rice, J. P. et al. CHRNB3 is more strongly associated with Fagerstrom test for cigarette dependence-based nicotine dependence than cigarettes per day: phenotype definition changes genome-wide association studies results. Addiction 107, 2019–2028, doi:10.1111/j.1360-0443.2012.03922.x (2012).
Loukola, A. et al. A Genome-Wide Association Study of a Biomarker of Nicotine Metabolism. PLoS Genet 11, e1005498, doi:10.1371/journal.pgen.1005498 (2015).
Gelernter, J. et al. Genome-wide association study of nicotine dependence in American populations: identification of novel risk loci in both African-Americans and European-Americans. Biol Psychiatry 77, 493–503, doi:10.1016/j.biopsych.2014.08.025 (2015).
Bierut, L. J. et al. Novel genes identified in a high-density genome wide association study for nicotine dependence. Hum Mol Genet 16, 24–35 (2007).
Gotti, C., Riganti, L., Vailati, S. & Clementi, F. Brain neuronal nicotinic receptors as new targets for drug discovery. Current pharmaceutical design 12, 407–428 (2006).
Zeiger, J. S. et al. The neuronal nicotinic receptor subunit genes (CHRNA6 and CHRNB3) are associated with subjective responses to tobacco. Hum Mol Genet 17, 724–734, doi:10.1093/hmg/ddm344 (2008).
Greenbaum, L. et al. Why do young women smoke? I. Direct and interactive effects of environment, psychological characteristics and nicotinic cholinergic receptor genes. Mol Psychiatry 11, 312–322, 223 (2006).
Stevens, V. L. et al. Nicotinic receptor gene variants influence susceptibility to heavy smoking. Cancer Epidemiol Biomarkers Prev 17, 3517–3525, doi:10.1158/1055-9965.EPI-08-0585 (2008).
Saccone, N. L. et al. Multiple distinct risk loci for nicotine dependence identified by dense coverage of the complete family of nicotinic receptor subunit (CHRN) genes. Am J Med Genet B Neuropsychiatr Genet 150B, 453–466, doi:10.1002/ajmg.b.30828 (2009).
Ehringer, M. A. et al. Association of CHRN genes with “dizziness” to tobacco. Am J Med Genet B Neuropsychiatr Genet 153B, 600–609, doi:10.1002/ajmg.b.31027 (2010).
Wang, S. et al. Significant associations of CHRNA2 and CHRNA6 with nicotine dependence in European American and African American populations. Hum Genet 133, 575–586, doi:10.1007/s00439-013-1398-9 (2014).
Swan, G. E. et al. Varenicline for smoking cessation: nausea severity and variation in nicotinic receptor genes. Pharmacogenomics J 12, 349–358, doi:10.1038/tpj.2011.19 (2012).
Wen, L., Jiang, K., Yuan, W., Cui, W. & Li, M. D. Contribution of Variants in CHRNA5/A3/B4 Gene Cluster on Chromosome 15 to Tobacco Smoking: From Genetic Association to Mechanism. Mol Neurobiol 53, 472–484, doi:10.1007/s12035-014-8997-x (2016).
Wen, L., Yang, Z., Cui, W. & Li, M. D. Crucial roles of the CHRNB3-CHRNA6 gene cluster on chromosome 8 in nicotine dependence: update and subjects for future research. Translational psychiatry 6, e843, doi:10.1038/tp.2016.103 (2016).
Yang, J. & Li, M. D. Converging findings from linkage and association analyses on susceptibility genes for smoking and other addictions. Mol Psychiatry 21, 992–1008, doi:10.1038/mp.2016.67 (2016).
Payne, T. J. & Diefenbach, L. Characteristics of African American smokers: a brief review. Am J Med Sci 326, 212–215 (2003).
Perez-Stable, E. J., Herrera, B., Jacob, P. 3rd & Benowitz, N. L. Nicotine metabolism and intake in black and white smokers. JAMA 280, 152–156 (1998).
Keskitalo-Vuokko, K. et al. Associations of nicotine intake measures with CHRN genes in Finnish smokers. Nicotine Tob Res 13, 686–690, doi:10.1093/ntr/ntr059 (2011).
Hubacek, J. A., Lanska, V. & Adamkova, V. Lack of an association between SNPs within the cholinergic receptor genes and smoking behavior in a Czech post-MONICA study. Genet Mol Biol 37, 625–630, doi:10.1590/S1415-47572014005000023 (2014).
Fagerstrom, K. O. Measuring degree of physical dependence to tobacco smoking with reference to individualization of treatment. Addict Behav 3, 235–241 (1978).
Heatherton, T. F., Kozlowski, L. T., Frecker, R. C. & Fagerstrom, K. O. The Fagerstrom Test for Nicotine Dependence: a revision of the Fagerstrom Tolerance Questionnaire. British journal of addiction 86, 1119–1127 (1991).
Culverhouse, R. C. et al. Multiple distinct CHRNB3-CHRNA6 variants are genetic risk factors for nicotine dependence in African Americans and European Americans. Addiction 109, 814–822, doi:10.1111/add.12478 (2014).
Wei, J. et al. Association study of 45 candidate genes in nicotine dependence in Han Chinese. Addict Behav 37, 622–626, doi:10.1016/j.addbeh.2012.01.009 (2012).
Etter, J. F. et al. Association of genes coding for the alpha-4, alpha-5, beta-2 and beta-3 subunits of nicotinic receptors with cigarette smoking and nicotine dependence. Addict Behav 34, 772–775, doi:10.1016/j.addbeh.2009.05.010 (2009).
Purcell, S. et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet 81, 559–575, doi:10.1086/519795 (2007).
Gabriel, S. B. et al. The structure of haplotype blocks in the human genome. Science 296, 2225–2229 (2002).
Barrett, J. C., Fry, B., Maller, J. & Daly, M. J. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21, 263–265 (2005).
Schaid, D. J., Rowland, C. M., Tines, D. E., Jacobson, R. M. & Poland, G. A. Score tests for association between traits and haplotypes when linkage phase is ambiguous. Am J Hum Genet 70, 425–434, doi:10.1086/338688 (2002).
Tao, J. et al. Fine mapping analysis of HLA-DP/DQ gene clusters on chromosome 6 reveals multiple susceptibility loci for HBV infection. Amino Acids 47, 2623–2634, doi:10.1007/s00726-015-2054-6 (2015).
Li, M. D., Lou, X. Y., Chen, G., Ma, J. Z. & Elston, R. C. Gene-gene interactions among CHRNA4, CHRNB2, BDNF, and NTRK2 in nicotine dependence. Biol Psychiatry 64, 951–957 (2008).
Zhu, Z. et al. Development of GMDR-GPU for gene-gene interaction analysis and its application to WTCCC GWAS data for type 2 diabetes. PLoS One 8, e61943, doi:10.1371/journal.pone.0061943 (2013).
Lou, X. Y. et al. A Generalized Combinatorial Approach for Detecting Gene-by-Gene and Gene-by-Environment Interactions with Application to Nicotine Dependence. Am J Hum Genet 80, 1125–1137 (2007).
Moher, D., Liberati, A., Tetzlaff, J., Altman, D. G. & Group, P. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. Ann Intern Med 151(264-269), W264 (2009).
Ma, Y., Fan, R. & Li, M. D. Meta-Analysis Reveals Significant Association of the 3’-UTR VNTR in SLC6A3 with Alcohol Dependence. Alcohol Clin Exp Res 40, 1443–1453, doi:10.1111/acer.13104 (2016).
Ma, Y., Yuan, W., Cui, W. & Li, M. D. Meta-analysis reveals significant association of 3’-UTR VNTR in SLC6A3 with smoking cessation in Caucasian populations. Pharmacogenomics J 16, 10–17, doi:10.1038/tpj.2015.44 (2016).
Willer, C. J., Li, Y. & Abecasis, G. R. METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics 26, 2190–2191, doi:10.1093/bioinformatics/btq340 (2010).
Higgins, J. P., Thompson, S. G., Deeks, J. J. & Altman, D. G. Measuring inconsistency in meta-analyses. BMJ 327, 557–560, doi:10.1136/bmj.327.7414.557 (2003).
Champtiaux, N. et al. Subunit composition of functional nicotinic receptors in dopaminergic neurons investigated with knock-out mice. J Neurosci 23, 7820–7829 (2003).
Zoli, M. et al. Identification of the nicotinic receptor subtypes expressed on dopaminergic terminals in the rat striatum. J Neurosci 22, 8785–8789 (2002).
Cui, C. et al. The beta3 nicotinic receptor subunit: a component of alpha-conotoxin MII-binding nicotinic acetylcholine receptors that modulate dopamine release and related behaviors. J Neurosci 23, 11045–11053 (2003).
Gotti, C. et al. Expression of nigrostriatal alpha 6-containing nicotinic acetylcholine receptors is selectively reduced, but not eliminated, by beta 3 subunit gene deletion. Mol Pharmacol 67, 2007–2015, doi:10.1124/mol.105.011940 (2005).
Cho, Y. S. et al. A large-scale genome-wide association study of Asian populations uncovers genetic factors influencing eight quantitative traits. Nat Genet 41, 527–534, doi:10.1038/ng.357 (2009).
Bierut, L. J. et al. A genome-wide association study of alcohol dependence. Proc Natl Acad Sci USA 107, 5082–5087, doi:10.1073/pnas.0911109107 (2010).
Landi, M. T. et al. A genome-wide association study of lung cancer identifies a region of chromosome 5p15 associated with risk for adenocarcinoma. Am J Hum Genet 85, 679–691, doi:10.1016/j.ajhg.2009.09.012 (2009).
Won, W. Y. et al. Genetic Association of CHRNB3 and CHRNA6 Gene Polymorphisms with Nicotine Dependence Syndrome Scale in Korean Population. Psychiatry Investig 11, 307–312, doi:10.4306/pi.2014.11.3.307 (2014).
Exley, R. et al. Distinct contributions of nicotinic acetylcholine receptor subunit alpha4 and subunit alpha6 to the reinforcing effects of nicotine. Proc Natl Acad Sci USA 108, 7577–7582, doi:10.1073/pnas.1103000108 (2011).
Jackson, K. J., McIntosh, J. M., Brunzell, D. H., Sanjakdar, S. S. & Damaj, M. I. The role of alpha6-containing nicotinic acetylcholine receptors in nicotine reward and withdrawal. J Pharmacol Exp Ther 331, 547–554, doi:10.1124/jpet.109.155457 (2009).
Sanjakdar, S. S. et al. Differential roles of alpha6beta2* and alpha4beta2* neuronal nicotinic receptors in nicotine- and cocaine-conditioned reward in mice. Neuropsychopharmacology 40, 350–360, doi:10.1038/npp.2014.177 (2015).
Pons, S. et al. Crucial role of alpha4 and alpha6 nicotinic acetylcholine receptor subunits from ventral tegmental area in systemic nicotine self-administration. J Neurosci 28, 12318–12327, doi:10.1523/JNEUROSCI.3918-08.2008 (2008).
Bar-Shira, A. et al. CHRNB3 c.−57A > G functional promoter change affects Parkinson’s disease and smoking. Neurobiol Aging 35(2179), e2171–2176, doi:10.1016/j.neurobiolaging.2014.03.014 (2014).
Kamens, H. M. et al. The beta3 subunit of the nicotinic acetylcholine receptor: Modulation of gene expression and nicotine consumption. Neuropharmacology 99, 639–649, doi:10.1016/j.neuropharm.2015.08.035 (2015).
Chen, J. et al. Genetic structure of the Han Chinese population revealed by genome-wide SNP variation. Am J Hum Genet 85, 775–785, doi:10.1016/j.ajhg.2009.10.016 (2009).
Zhang, H. & Cai, B. The impact of tobacco on lung health in China. Respirology 8, 17–21 (2003).
Acknowledgements
This study was supported in part by the China Precision Medicine Initiative (2016YFC0906300), Research Center for Air Pollution and Health of Zhejiang University, the State Key Laboratory for Diagnosis and Treatment of Infectious Diseases of the First Affiliated Hospital of Zhejiang University, and Shanxi Key Laboratory of Ecological Animal Science and Environmental Veterinary Medicine of Shanxi Agricultural University. We thank Dr. David L. Bronson for excellent editing of this manuscript.
Author information
Authors and Affiliations
Contributions
L.W., H.H., K.S., Z.Y., W.C., W.Y., Y.M., K.J., X.J. and J.W. participated clinical data collection; H.H., Q.L., Z.Y., R.F., J.C. and X.J. performed laboratory experiments; L.W., H.H. and Q.L. participated data analysis; W.L., Q.L., T.J.P. and J.W. participated paper writing; M.D.L. conceived the study and was involved every step of this study; and all authors approved the paper.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wen, L., Han, H., Liu, Q. et al. Significant association of the CHRNB3-CHRNA6 gene cluster with nicotine dependence in the Chinese Han population. Sci Rep 7, 9745 (2017). https://doi.org/10.1038/s41598-017-09492-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-09492-8
- Springer Nature Limited
This article is cited by
-
Association and cis-mQTL analysis of variants in serotonergic genes associated with nicotine dependence in Chinese Han smokers
Translational Psychiatry (2018)