

Abstract
Komodo dragons (Varanus komodoensis) are the largest extant predatory lizards and their ziphodont (serrated, curved and blade-shaped) teeth make them valuable analogues for studying tooth structure, function and comparing with extinct ziphodont taxa, such as theropod dinosaurs. Like other ziphodont reptiles, V. komodoensis teeth possess only a thin coating of enamel that is nevertheless able to cope with the demands of their puncture–pull feeding. Using advanced chemical and structural imaging, we reveal that V. komodoensis teeth possess a unique adaptation for maintaining their cutting edges: orange, iron-enriched coatings on their tooth serrations and tips. Comparisons with other extant varanids and crocodylians revealed that iron sequestration is probably widespread in reptile enamels but it is most striking in V. komodoensis and closely related ziphodont species, suggesting a crucial role in supporting serrated teeth. Unfortunately, fossilization confounds our ability to consistently detect similar iron coatings in fossil teeth, including those of ziphodont dinosaurs. However, unlike V. komodoensis, some theropods possessed specialized enamel along their tooth serrations, resembling the wavy enamel found in herbivorous hadrosaurid dinosaurs. These discoveries illustrate unexpected and disparate specializations for maintaining ziphodont teeth in predatory reptiles.
Similar content being viewed by others
Main
Ziphodont teeth evolved convergently in several extinct apex predators, including the ‘pelycosaur’ Dimetrodon1 and theropod dinosaurs1,2,3,4,5,6. These teeth have serrated cutting edges composed of dentine-cored serrations (denticles) capped with a thin veneer of hard enamel. The slenderness of the hard enamel cap is most striking in extinct ziphodont species, where even the largest predatory dinosaurs have only 10–20% of the absolute enamel thickness of a human tooth and lack the structural complexity of mammalian enamel2,7. Despite having thin, structurally simpler enamel, amniotes repeatedly evolved ziphodont teeth2, suggesting that this tooth morphology is linked to the success of many extinct carnivores. This also suggests that there may be common specializations hidden in ziphodont enamel which enable these teeth to effectively cut and tear into animal tissue.
Unfortunately, our poor understanding of the material properties of non-mammalian dental tissues in general7,8 and of the confounding effects of fossilization, limit our ability to infer structure–function relationships in extinct lineages. However, the Komodo dragon (Varanus komodoensis) is a rare example of an extant reptile bearing ziphodont teeth and is, therefore, a key taxon for understanding their function5,9,10,11,12. Here, we describe the unique specializations within the enamel of V. komodoensis and compare them with those of other squamates, crocodylians and theropod dinosaurs. Through state-of-the-art elemental, structural and nanomechanical characterization, we (1) reveal surprising structural and chemical complexity of V. komodoensis teeth; (2) compare adaptations for maintaining cutting-edge enamel in varanids and extant crocodylians; and (3) contrast these adaptations in extant reptiles with those of extinct theropod dinosaurs.
Results
Iron-coated Komodo dragon teeth
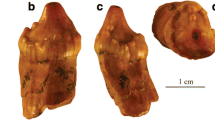
Our initial examination of the teeth of skeletal and fluid-preserved specimens of V. komodoensis revealed an overlooked feature of their dentition: all the unworn tooth crown tips and serrations are pigmented orange (Fig. 1 and Supplementary Table 1). Erupted and unerupted teeth are equally pigmented, indicating that it is not staining from feeding (Fig. 1 and Supplementary Fig. 1). The orange colouration is most intense along serrations closer to the crown tips, whereas the rest of the crown is coated in transparent enamel, making these regions white from the underlying dentine. The pigmentation is even more pronounced when using laser-stimulated fluorescence (LSF)13 to image both erupted and unerupted teeth, in which the surrounding enamel fluoresces brightly and the pigmented serrations and tips appear dark (Fig. 1d,e).

a, Lateral view of the skull of V. komodoensis (Natural History Museum, London, NHMUK 1934.9.2.1). b, Lingual view of a dentary tooth position showing several unerupted replacement teeth with orange pigmentation (American Museum of Natural History, AMNH 37912). c, Dorsal view of two erupted teeth from a fluid-preserved specimen (Zoological Society of London) showing pigmented cutting edges and apices. d, White light (WL) image of an erupted and unerupted tooth in the same specimen. e, Laser stimulated fluorescence (LSF) image of the same specimen, showing the pronounced serration pigmentation in both the erupted and unerupted teeth. f, Dorsal view of three left dentary teeth in NHMUK 1934.2.1 showing identical pigmentation on the tooth apices and mesial serrations. g, Lateral view of an isolated replacement tooth (Museum of Life Sciences, MoLS X263). h, Close-up of tooth apex in g showing the orange pigmentation along the tooth tip. i, Distal view of tooth serrations of MoLS X263 showing orange serrations and tooth apex. j, Polished thick section through mesial denticles of a tooth (Queensland Museum, Australia, J94036-2) showing orange pigmentation restricted to the enamel. k, SEM image of three mesial serrations of J94036-2. l, Close-up of serration enamel showing the bright coating. m, Close-up of the crown apex enamel showing the same nanocrystalline coating. Asterisks indicate pigmented regions. de, dentine; en, enamel; et, erupted tooth; gi, gingiva; ut, unerupted tooth.
Similar enamel pigmentation is found in specialized mammal teeth14; however, this feature has never been reported before in a carnivorous reptile. Scanning electron microscope (SEM) imaging of acid-etched sections of the serrations and tooth apices of V. komodoensis showed that the enamel is only 20 µm thick (Fig. 1k–m). Moreover, it is composed of parallel-crystallite enamel; a simple, prismless enamel type where bundles of crystallites are oriented perpendicular to the outer surface (Fig. 1m). However, the outermost 1–2 µm in V. komodoensis enamel consists of a smooth coating which appears bright under SEM. This coating is found along the serrations and crown tips.
To determine the composition of the pigmented regions, we subjected functional and replacement teeth to several complimentary imaging and characterization techniques (Methods). The orange colouration of the enamel is similar to the iron oxide-enriched outer layers in mammals with pigmented teeth14,15. Indeed, every technique detected iron in the outer 1–2 µm of the enamel and occasionally an associated concentration of zinc (Supplementary Figs. 2 and 3). The high zinc content in the outermost enamel may be a product of normal enamel secretion and maturation, similar to that in mammalian enamel16. However, the iron was consistently concentrated along the mesial and distal serrations in horizontal sections but not along enamel further away from the serrations (Fig. 2g–i and Supplementary Fig. 2). SEM–energy-dispersive X-ray spectroscopy (EDS) and laser ablation inductively coupled plasma mass spectrometry (LA-ICP-MS) analyses confirmed that the iron-enriched layer coincides with the thin outer coating we identified along the crown tips and serrations under SEM, which overlies the calcium-rich enamel (Fig. 2j,k and Supplementary Figs. 2 and 3). The colocation of high iron and higher oxygen signals (Supplementary Fig. 2k,l) suggests an iron oxide composition which coats and colours the outer enamel orange.

a, Polished thick section of a tooth apex and distal serrations (MoLS X263). b, S-µXRF elemental map for iron along the tooth apex and serrations in MoLS X263. c,d, High-resolution S-µXRF map of iron along two apical serrations (c), compared with the calcium map of the same region (d), showing that iron is restricted to the outer enamel surface. e,f, High-resolution S-µXRF map of iron along the tip of the tooth (e), compared with the calcium map of the same region (f). g, High-resolution S-µXRF map of iron along a horizontal section through a distal serration in the same tooth. h, Wholeview image of horizontal section through MoLS X263. i, Higher resolution S-µXRF map of iron along the horizontal section through a mesial serration in MoLS X263. j, SEM–EDS map of iron from a thick section of a functional tooth (J94036-2) showing that the iron is most abundant in the coating overlying the enamel. k, SEM–EDS map of calcium from the same view as in j, showing reduced calcium signal in the outer coating. l, STEM–EDS map of iron from an FIB-milled portion of the outer enamel coating of J94036-2 (outer surface of tooth is towards the top). m, STEM–EDS map of calcium in the same region, showing lack of calcium in the iron-rich coating. n, STEM image of the interface between the iron-rich coating (above) and crystalline enamel (below) and iron-rich material interspersed between enamel crystallites (middle). In all elemental maps, brighter colours indicate higher counts. In the illustrations, red represents the pigmented regions, white is dentine and grey is enamel. am, amorphous region; cr, crystalline region; ecr, enamel crystallites; edj, enamel–dentine junction; Fe, iron.
We further isolated a portion of this iron coating using focused ion beam (FIB) milling to conduct higher resolution scanning transmission electron microscope (STEM) and STEM–EDS analyses to determine how this iron-rich layer is incorporated into the enamel. These data revealed that the iron-enriched layer consists of a thin (100–200 nm) inner region which is interspersed between individual enamel crystallite bundles and a thicker outer layer, devoid of calcium, coating the enamel (Fig. 2l–n).
Iron in other extant reptile teeth
Given the surprising and consistent orange colouration of both the replacement and functional teeth in V. komodoensis, we further surveyed skeletal collections to determine the prevalence of this feature across a sample of Varanus species (Fig. 3a). Orange cutting edges were present in V. salvadorii, V. rosenbergi and V. giganteus to varying degrees and occasionally in V. varius, V. salvator and V. indicus teeth (Supplementary Fig. 4). These species all bear serrations, albeit smaller than those of V. komodoensis. Pigmented cutting edges were not visible in five other Varanus species, including some with small serrations and others that had non-serrated cutting edges. For comparisons, the cutting edges of the teeth of the anguimorph lizard Heloderma and a small sample of other squamate specimens we examined were not pigmented (Supplementary Fig. 5 and Supplementary Table 1). This suggests that iron-pigmented cutting edges are found in several ziphodont Varanus species but are most pronounced in V. komodoensis.

a, Simplified phylogeny of Varanus species examined in this study (modified from ref. 40, position of V. priscus taken from ref. 41). Red branches indicate taxa with obvious, consistently pigmented cutting edges. Black branches indicate those with no obvious pigmentation. Blue branches indicate taxa with occasional or inconsistently pigmented teeth. Orange branch indicates unknown level of pigmentation due to fossilization. Scale bars, 1 mm. b, Isolated tooth crown of the extinct V. priscus (SAMP 54739) showing no signs of pigmentation on the distal serrations. c, A second isolated crown assigned to V. priscus (SAMP 54739) showing no evidence of pigmentation.
The prevalence of pigmented cutting edges, particularly in the larger serrated varanid teeth, led us to further predict that the largest ziphodont squamate, the extinct Varanus priscus (‘Megalania’) may have also possessed iron-coated serrations. However, these fossilized teeth showed no visual evidence of pigmentation (Fig. 3b,c). Indeed, the varanid fossil record in general includes many fossilized teeth bearing serrations17,18,19,20,21; however, none of these occurrences shows any obvious pigmentation similar to their extant counterparts. Furthermore, the small absolute sizes of varanid teeth and the paucity of fossil teeth available for destructive analyses make it difficult to assess the material properties of iron coatings in extant taxa or how fossilization has hindered our ability to visually detect them in extinct species. We therefore sought to test for iron-pigmented enamel in another reptilian group with much larger teeth and a more abundant dental fossil record: modern and fossil crocodylians and the ziphodont teeth of theropod dinosaurs.
We first examined the teeth of several species of crocodylians to determine if extant toothed archosaurs are capable of sequestering iron in a similar fashion to Varanus. We subjected shed teeth of zoo-kept crocodylians (Alligator mississippiensis, Crocodylus porosus, Osteolaemus tetraspis and Tomistoma schlegelii) to similar elemental analyses. Although their teeth are not serrated, we observed orange-coloured carinae (non-serrated cutting edges) in some but not all crocodylian teeth we sampled. However, even in crocodylian teeth lacking obvious pigmentation under white light, we were able to identify similar fluorescence patterns along the carinae to those we identified in V. komodoensis teeth using LSF (Fig. 4 and Supplementary Figs. 6 and 7). LA-ICP-MS and synchrotron-based X-ray microfluorescence (S-µXRF) analyses confirmed the presence of an outer iron-enriched enamel layer in all four crocodylian species, similar in distribution to those seen in V. komodoensis (Fig. 4 and Supplementary Figs. 6–8). In some cases, these appeared to be more concentrated along the carinae, whereas in others the iron layer was more evenly distributed along the enamel surface. Thus, extant toothed archosaurs appear to be able to sequester iron in their enamel and to varying degrees along their cutting edges, albeit in more subtle arrangements compared with those of V. komodoensis.

a,b, Close-up of a posterior tooth of Tomistoma schlegelii under WL (a) and LSF (b), revealing a distinctive fluorescence pattern along the carina (asterisks). c, Close-up of the tooth tip of an anterior T. schlegelii tooth showing an orange carina under WL. d, Polished horizontal thick section through the carina of the tooth in c showing orange pigmentation (asterisks). e,f, LA-ICP-MS map of iron (e) and calcium (f), taken over a horizontal section through a carina from the tooth in a. g, Polished thick section of a caniniform tooth of A. mississippiensis cut parallel to the carinae, showing orange pigmentation along the outer enamel region (asterisks). h,i, LA-ICP-MS maps of iron (h) and calcium (i), from the enamel and dentine of the tooth in g. j, Welch test of hardness of pigmented and non-pigmented enamel in the same tooth measured using nano-indentation (n = 38 (non-pigmented) and 36 (pigmented) indents in one tooth). Central lines represent medians (3.33, 3.63), upper and lower bounds of boxes represent lower quartiles (3.14, 3.38) and upper quartiles (3.55, 4.06), minima (2.47, 2.68) and maxima (4.6, 5.63) for non-pigmented and pigmented enamel, respectively. P value (two-tailed) was 0.0058. k,l, S-µXRF maps of iron (k) and calcium (l) along the tip of a tooth section from a C. porosus tooth cut along the carina. m, Mesial view of the tooth sectioned and mapped in k and l. n, Section used in k and l, showing region of interest. o,p, S-µXRF maps of iron (o) and calcium (p) taken from a fossilized crocodylian tooth from Dinosaur Provinical Park (Canada) (UALVP 60550). q, Mesial view of UALVP 60550 before sectioning showing no evidence of pigmentation along the carina. r, Section used in o and p showing region of interest. In all elemental maps, brighter colours indicate higher counts. cn, carina.
Composition of the iron coating in V. komodoensis
We conducted iron-edge X-ray absorption near-edge spectroscopy (Fe-XANES) to characterize the chemical speciation of the iron coating in V. komodoensis and compare it to other iron-enriched enamels. Comparison with standards suggested that the white line (7,132 eV) and post-edge features, including a peak at 7,147 eV, were most consistent with ferrihydrite (Supplementary Fig. 9), which is also found in pigmented mammal teeth15. Combinations with other iron oxide species cannot be excluded; however, haematite exhibits several post-edge features between 7,132 and 7,160 eV and magnetite exhibits a small shift in the principal absorption peak energy, both of which were not observed in the V. komodoensis iron coating. The Fe K-edge XANES spectra were also compared with an extant crocodylian tooth (Crocodylus porosus) and iron-enriched beaver (Castor canadensis) enamel. These spectra were different across taxa, suggesting variation in iron speciation across species with pigmented enamel.
Searching for iron coatings in fossil reptile teeth
We similarly subjected Late Cretaceous-aged crocodylian teeth from Dinosaur Provincial Park (Alberta, Canada) to LA-ICP-MS and S-µXRF to examine and compare the distribution of iron and calcium between extant and fossilized crocodylian teeth. The resulting maps consistently showed increased calcium and iron signals throughout the dentine of each tooth with no indication of increased iron concentrations along the outer enamel (Fig. 4o,p and Supplementary Fig. 7). These results indicate that even if these crocodylian teeth possessed iron in their enamel, fossilization may obscure this signal, as a result of the abundance of iron and other exogenous elements in fossilized dental tissues.
We then undertook elemental imaging of several tyrannosaurid and dromaeosaurid theropod specimens (Supplementary Table 2) to examine elemental composition and heterogeneity along their serrations. Given the ability of extant archosaurs to sequester iron in their enamel and the intensity of the colouration in ziphodont Varanus teeth, we expected to see some level of increased iron signal within the serration enamel of ziphodont dinosaurs. However, preliminary white light and LSF imaging of a sample of theropod teeth showed no evidence of differential fluorescence patterns along the cutting edges (Fig. 5a–f and Supplementary Fig. 10). Furthermore, LA-ICP-MS and S-µXRF analyses of theropod teeth showed similar results to those of fossil crocodylian teeth: the dentine was enriched in iron, whereas the enamel was relatively depauperate, except in regions of diagenetic crack infilling (Fig. 5 and Supplementary Figs. 10–14). A similar phenomenon occurred for other elements, which had comparable or higher signals in the dentine compared with the enamel, whereas the opposite occurs in extant reptile teeth. These findings suggest that, similar to our crocodylian sample, either diagenetic alterations mask this feature in fossil theropod teeth or theropods did not normally sequester iron along their serrations and may have used an alternative adaptation to reinforce their serration enamel.

a, Labiolingual view of a tooth of the tyrannosaurid Albertosaurus (NHMUK R12599). b,c, Distal view of NHMUK R12599 under WL (b) and LSF imaging (c), showing a lack of differential fluorescence between the serrations or the rest of the tooth. d, Labiolingual view of a dromaeosaurid tooth (UALVP 61165). e,f, Distal view of a dromaeosaurid tooth (UALVP 61165) under WL (e) and LSF imaging (f), showing no fluorescence difference on- or off-serration. g, Polished horizontal thick section through a distal serration of a tyrannosaurid tooth (UALVP 60554). h, LA-ICP-MS map of iron along the distal serration in g showing no evidence of iron sequestration along the serration enamel. i, Polished horizontal thick section through a distal serration of UALVP 61165. j, LA-ICP-MS map of iron along the same region as in g. k, Close-up view of serration enamel in a tyrannosaurid tooth (UALVP 60553). l, Serration enamel in a dromaeosaurid tooth (UALVP 61165). m, Polished thick section through distal serrations of a tyrannosaurid tooth (UALVP 60555). n, LA-ICP-MS map of iron along the serrations in UALVP 60555. o, Polished thick section through the distal serrations of a dromaeosaurid tooth (UALVP 61165). p, LA-ICP-MS map of iron along the serrations in UALVP 61165. In all elemental maps, brighter colours indicate higher counts.
Structural complexity in the enamel of theropod teeth
To assess microstructural features within theropod serrations, we next undertook qualitative analyses (light and scanning electron microscopy) to characterize theropod enamel microstructure. Compared to V. komodoensis, theropod enamel is often thicker (50–200 µm) and can be composed of a more complex microstructure (Fig. 6 and Supplementary Figs. 15 and 16). Tyrannosaurid enamel in particular is organized into discrete columns of herringboned stacks of crystallites, which extend out from the enamel–dentine junction7,22 (Fig. 6c,d). However, this enamel structure differed along the serrations: crystallites within the outer half of the serration enamel form clusters that spiral towards the outer enamel surface (Fig. 6g,h). This enamel type is reminiscent of wavy enamel found in hadrosaurs, a herbivorous group of dinosaurs with grinding teeth7,22. Wavy enamel forms characteristic wave patterns under cross-polarized light23,24 which are also present along tyrannosaurid serrations in thin section (Fig.6f and Supplementary Fig. 17). By comparison, this wavy pattern was largely absent from the smaller teeth of dromaeosaurids, where the enamel was usually of a simpler, parallel crystallite type across the whole tooth (Supplementary Fig. 16).

a, Close-up image of an isolated tyrannosaurid tooth (UALVP 60556), with illustrations of the distributions of columnar (grey) and wavy (blue) enamel based on structural analyses. b, Polished thick section through the distal serrations of UALVP 60555 showing positions of sections in subsequent analyses. c, SEM image of columnar enamel in a tyrannosaurid tooth in horizontal section (UALVP 60556). d, Higher magnification SEM image showing herringbone stacks of enamel crystallites within each column. e, S-µXRD-generated diffraction pattern taken through a region of columnar enamel in a tyrannosaurid tooth (UALVP 60554), showing three principal enamel crystallite orientations in the 002 reflection. f, Cross-polarized light image of serration enamel in a thin section of a tyrannosaurid tooth (UALVP 60398) showing wavy pattern in enamel. g, SEM image of enamel along a serration of a tyrannosaurid tooth in longitudinal section (UALVP 60557). h, Higher magnification image of the outer enamel surface showing the spiralled arrangements of serration enamel. i, S-µXRD-generated diffraction pattern taken through a region of wavy enamel in a tyrannosaurid tooth, showing two principal enamel crystallite orientations in the 002 reflection and a higher divergence angle between them compared with the columnar enamel (e). j, Composite apatite crystal texture map of two serrations in longitudinal section (UALVP 53472) derived from diffraction patterns. k, Composite texture map of enamel crystallites along a single serration in horizontal section (UALVP 60554). Lines within each pixel in j and k indicate principal enamel crystallite orientations. Hotter colours indicate more highly textured regions (lower full-width half maxima). Wavy enamel manifests as larger divergences between principal orientations and more textured regions along the serration. ce, columnar enamel; ec, enamel column; FWHM, full-width at half-maximum; oes, outer enamel surface; we, wavy enamel.
To quantify the variation in enamel structure and texture along tyrannosaurid tooth crowns, we performed synchrotron-based X-ray microdiffraction (S-µXRD) mapping of thinned tooth samples. Previous studies of fossil reptile enamel using S-µXRD have studied powdered theropod dinosaur teeth or small transect maps of fossil and modern crocodylian teeth8,25,26 and thus were unable to provide spatial information along a tooth crown. In contrast, our mapping of apatite crystallite orientations as well as the texture (variance in orientation) in situ along and across entire tyrannosaurid serrations allowed us to quantify structural variation along the section, a method that has previously been used on human teeth27 (Supplementary Figs. 18–20). In tyrannosaurids, the dentine was comparatively poorly textured, with no obvious preferred crystallite orientations (Fig. 6j,k). Crystallites within the serration enamel, by comparison, showed one to three principal orientations, depending on the region of enamel. Along the serrations, the inner layers possessed a single principal apatite crystal orientation with crystal c axes largely parallel to the enamel–dentine junction but had a bidirectional crystallite orientation which increasingly diverges towards the serration surface (Fig. 4i,j). In horizontal section, the enamel textures on- and off-serration were significantly different (Supplementary Fig. 20 and Supplementary Table 3). The columnar enamel observed by SEM along most of the crown retains three principal orientations at acute angles to one another, reflecting the herringbone arrangements of the enamel crystallites within each column (Fig. 6e,k). In contrast, serration enamel possessed more textured, divergent, bidirectional orientations associated with the wavy enamel we observed under SEM (Fig. 6h–k).
Mechanical testing of reptilian enamels
Nanomechanical testing of cut and polished V. komodoensis and A. mississippiensis teeth revealed that enamel hardness and elastic moduli of extant reptile teeth were comparable to various mammalian enamel types14 (Supplementary Figs. 21 and 22). Reliably testing the iron coatings of the serration enamel in V. komodoensis proved difficult because it was <1 µm thick in polished sections; however, we were able to directly test thicker iron-enriched regions in an A. mississippiensis tooth (Supplementary Fig. 22). The indentation hardness of these iron-enriched enamel regions (\(\bar{x}=\) 3.70 GPa) was slightly (9%) but significantly higher than that of underlying enamel (\(\bar{x}=\) 3.36 GPa; t = 2.89, P = 0.0054). However, the indentation moduli (a measure of stiffness) did not differ greatly across the enamel (Fig. 4j, Supplementary Fig. 22 and Supplementary Table 4), which is consistent with measurements in some iron-pigmented mammal teeth14. Tyrannosaurid enamel and dentine were typically twice as hard as analogous regions in the extant V. komodoensis and A. mississippiensis teeth, probably reflecting the effects of fossilization (Supplementary Figs. 23 and 24).
Discussion
Despite their use as analogues for theropod dinosaur feeding behaviour9,11,28, our study demonstrates a striking and previously overlooked predatory adaptation in the Komodo dragon, V. komodoensis: iron-enriched, protective layers along their tooth serrations and tips. Iron sequestration is found in the dental enamel of specialized mammals14,15,29,30; however, the ability to sequester iron into a discrete coating along the cutting edges of a tooth has never been observed, let alone in a reptile. Furthermore, unlike in pigmented rodent teeth where mixed-phase iron oxides are incorporated into the intergranular spaces within enamel, iron appears to be concentrated into a distinct coating of ferrihydrite which is bonded to the underlying crystalline enamel in V. komodoensis (Fig. 2 and Supplementary Fig. 9). This was unexpected, given that V. komodoensis possesses only 15–20 µm of enamel and replaces its teeth rapidly12.
Iron-pigmented enamel has been studied in beavers15, shrews14, some fish31 and salamanders32,33 but its function remains unclear14. In some cases, iron sequestration hardens the enamel15; in others the mechanical properties of pigmented and unpigmented regions are statistically indistinguishable14. The iron-enriched regions in A. mississippiensis teeth provided us with thick enough material for nanomechanical testing and we determined that the pigmented regions exhibited slightly higher indentation hardness compared with unpigmented enamel. This local increase in hardness indicates that iron sequestration may serve to make the outer enamel layers along the cutting edges more wear-resistant compared with other regions of the tooth. Inferring a similar function to the serrated teeth of V. komodoensis is more difficult, given that the iron coating (and the enamel in general) is so thin. However, the coating covers each serration and is most pronounced in extant varanid species with highly serrated cutting edges, further indicating a role in supporting ziphodont tooth function. Consistent with this, the serrations also show different mechanical wear compared with other regions of the tooth crown: most of a worn crown of V. komodoensis can show spalling of enamel and exposure of the underlying dentine, whereas the serration enamel appears to wear gradually (Supplementary Fig. 25). This thin iron coating must therefore be enough to retain a sharp cutting edge on each tooth before it is replaced by a new generation, which would have occurred very frequently12. Other proposed functions for iron pigmentation include increased abrasion-, acid- or microfracture-resistance14,30. The iron layer is acid-resistant in V. komodoensis, based on SEM observations of acid-etched teeth (Supplementary Fig. 26), and it may therefore also protect tooth serrations from digestive acids.
Our ability to visually detect the same pigmented cutting edges in closely related species of Varanus suggests that iron sequestration may also be widespread in reptile teeth. We therefore predicted to see this feature in the large ziphodont teeth of closely related extinct taxa as well (for example, V. (‘Megalania’) priscus). However, our broader comparisons show that iron sequestration is neither a presence-or-absence phenomenon in extant reptiles nor is it easily detectable in fossil reptile teeth. Even where pigmentation was absent from the cutting edges in extant reptile teeth under white light, fluorescence imaging techniques revealed that even extant crocodylians can possess iron-enriched outer enamel layers, particularly along the cutting edges. This suggests that many reptiles may have iron-enriched enamel but that only some species have evolved prominent iron coatings along specific parts of their tooth crowns, presumably as feeding adaptations. Under high enough concentrations, the iron layers are visible under white light, as is the case in V. komodoensis and some of its ziphodont relatives.
Given that extant archosaurs can sequester iron within their enamel and that V. komodoensis, the largest extant ziphodont reptile, possessed iron-coated tooth serrations, we predicted that we may be able to detect similar iron coatings in extinct ziphodont varanids and theropod dinosaurs. However, even in cases where we might expect to observe this adaptation, diagenesis may obscure any site-specific iron signal within the enamel. We did not detect any colour changes along the serrations of V. priscus and we did not see any consistent iron-enrichment within the serration enamel of theropod dinosaurs using any technique.
Despite the lack of evidence for iron coatings along theropod tooth serrations, our sample of tyrannosaurid teeth demonstrated a second surprising adaptation within ziphodont reptile teeth. SEM imaging and S-µXRD mapping consistently showed microstructural differences between the serration and non-serration enamel in tyrannosaurid teeth. Tyrannosaurids possessed a form of wavy enamel along the tooth serrations that is elsewhere only found on the grinding teeth of herbivorous ornithopod dinosaurs7,22. Unlike in ornithopods, wavy enamel is restricted to the serrations in tyrannosaurid teeth, rather than along the entire crown (Fig. 6). This specialization of the serration enamel in tyrannosaurids suggests a role in supporting the cutting edges of the teeth. In ornithopods, this staggered, sinusoidal arrangement of crystallite bundles may have hindered crack propagation and avoided larger scale fracturing of the enamel, owing to the convoluted pathways in which microfractures could form7. As in V. komodoensis, tyrannosaurid teeth frequently show evidence of enamel spalling34, however this is not typically seen on the serrations themselves (Supplementary Fig. 25).
Alternative feeding strategies between V. komodoensis9 and some tyrannosaurids10,35 may explain these divergent strategies for maintaining serrated teeth but these differences could also reflect scaling effects between these two ziphodont taxa. Dromaeosaurid dinosaur teeth provide partial evidence for this. The dromaeosaurid teeth sampled in this study were of similar sizes to those of V. komodoensis and most showed comparably thin and simple enamel microstructure between the serrations and the rest of the crown (Supplementary Fig. 16). Only in a dromaeosaurid tooth with thicker enamel (40 µm or more) did we see any qualitative differences in enamel structure on- and off-serration but these were still not as pronounced as in the larger tyrannosaurid teeth (Supplementary Fig. 16k–o). Despite having relatively thin enamel coatings, it is possible that only the larger teeth of theropod dinosaurs possess enough enamel over which the microstructure can influence the mechanical wear of the serrations. Conversely, the enamel is probably too thin in extant ziphodont lizards to influence tooth wear on a similar scale to tyrannosaurids. The thin, structurally simple enamel in V. komodoensis probably plays a limited role in the cutting function of each tooth and natural selection may have alternatively favoured individuals with more elaborate iron coatings to mitigate wear along their rapidly replaced teeth. If theropod dinosaurs did similarly incorporate iron into their ziphodont teeth, then we may expect to find clearer evidence in smaller species or individuals with comparably thin enamel. Identifying these iron coatings in fossil reptile teeth, however, remains a challenge; we did not see any evidence for iron sequestration, even in small dromaeosaurid teeth.
Our results highlight two unexpected and disparate evolutionary adaptations in ziphodont reptiles. On the one hand, V. komodoensis evolved prominent iron coatings along the tooth tips and serrations to reinforce their cutting teeth. By comparison, some theropod dinosaurs developed structural adaptations in enamel along their tooth serrations. In both cases, the cutting edges of the teeth in these ziphodont reptiles are either structurally or chemically modified. Despite having remarkably thin enamel, Komodo dragon and tyrannosaurid teeth both highlight the reptilian capacity to modify specific regions of the dental hard tissues to maintain sharp cutting edges. Whereas structurally and chemically complex enamel are traditionally associated with derived mammalian groups14,15,30, it is evident that similar adaptations lie hidden within the thin veneer of enamel of carnivorous reptiles and probably contributed to their success as apex predators.
Methods
Light microscopy
Imaging of Natural History Museum London (NHMUK), Museum of Life Sciences (MoLS) and South Australian Museum (SAM) specimens was done using a Nikon D810 DSLR camera or a Keyence VHX 7000 digital microscope (King’s College, London). American Museum of Natural History (AMNH) specimens were imaged using a Leica digital dissecting microscope.
Sample preparation for analyses
Samples were all first embedded in epoxy resin (EpoThin2, Beuhler) and placed under vacuum. Samples were then sectioned along planes of interest using an Isomet 1000 low-speed saw (Buehler) and polished using a LaboForce (Struers) grinding and polishing wheel, using P1200, P2000 and P4000 grinding papers and aluminium oxide polishing powders on cloth plates. Samples used for SEM were etched for 15–30 s using 1 M hydrochloric acid, placed in distilled water for several minutes, then mounted to SEM stubs. Samples prepared for S-µXRF and S-µXRD analyses were embedded in the same resin and cut using the Isomet saw to thicknesses of ~500 µm, then glued to blank epoxy resin pucks for mounting on to the LaboForce polishing machine. Samples were then ground to thicknesses of <300 µm using the same grinding papers and polishing powders before being removed from the resin pucks using a dental drill.
Scanning electron microscopy–energy dispersive spectroscopy
SEM imaging was conducted in two facilities: Centre for Ultrastructural Imaging (CUI) at King’s College London and Harvey Flowers Electron Microscopy Suite, Imperial College London. Samples were coated with 6–10 nm of gold and imaged using a JEOL JCM-7000 or a JEOL JSM-7800 F Prime SEM (CUI). These samples were imaged under high-vacuum setting using a 15 kV beam acceleration voltage on a JCM-7000 SEM (JEOL). SEM–EDS analysis was performed on a Sigma 300 SEM (Zeiss) equipped with an XFlash 6–60 EDS detector (Bruker), using 10 kV beam accelerating voltage (Harvey Flowers Electron Microscopy Suite).
STEM cross-sections were prepared from the SEM sample using a Helios 5 CX (ThermoFisher Scientific) with a Ga ion beam following standard milling procedures. Samples were milled to a thickness of 70–100 nm.
Electron microscopy–energy dispersal spectroscopy
TEM cross-sections were prepared from the SEM sample using a ThermoFisher Scientific (TFS) Helios 5 CX FIB-SEM with a Ga ion beam following standard milling procedures. Samples were milled to electron transparency with a thickness of 70–100 nm.
STEM imaging and EDS analysis was carried out on a TFS Spectra 300 TEM equipped with a monochromator and probe corrector. High-angle annular dark-field (HAADF) images were collected on a Fischione HAADF detector where the contrast in the image is roughly proportional to the atomic number squared (Z2) and the mass-thickness of the sample. EDS was acquired at 300 kV from a Dual-X windowless EDX detector with a solid angle of 1.8 srad. TFS Velox software (ThermoFisher Scientific) was used to process the data.
Laser-stimulated fluorescence
LSF of fossil theropod teeth was performed with a 405 nm diode laser projected through a lens as a line following a previously published methodology13. The line was scanned over the specimen in a dark room and a Nikon D850 camera with a 450 nm blocking filter recorded the image. LSF imaging of crocodylian teeth was conducted using the same 405 nm diode laser projected as a cone and imaged using the Keyence VHX 7000 microscope with a 505 nm blocking filter. Images were colour equalized in Photoshop (Adobe, v.25.5), applied equally to the entire image.
Synchrotron radiation X-ray microfluorescence spectroscopy
S-µXRF analyses were undertaken at the Diamond Light Source (Oxford Harwell Campus) beamline B16 (experiments M22284-2019 and MM26050-2020) and European Synchrotron Radiation Facility (Grenoble, France) beamlines BM28 (experiments 28-01-1286 and LS-3093) and ID21 (experiment LS-3074). All experiments were conducted using a monochromatic beam. Data acquisition parameters for each experiment and sample used in this study are detailed in Supplementary Table 2.
For data processing, raw XRF data were imported into PyMCA (v.5.6.3), calibrated to the beam energy, batch fitted and normalized to incident beam intensity for each scan. Maps were then generated in Matplotlib36 in Python (v.3.9.7).
Synchrotron radiation X-ray microdiffraction mapping
Synchrotron X-ray microdiffraction (S-µXRD) mapping measurements were conducted on the B16 beamline37 at Diamond Light Source and the BM28 beamline at the European Synchrotron Radiation Facility. An incident X-ray energy of 15.5 keV was used, equivalent to a wavelength of 0.799 Å, with a beam size of 24 × 19 µm2 (horizontal × vertical). Specimens were mounted in a transmission geometry normal to the impinging X-rays onto a travelling x–y sample stage to enable measurements in two orthogonal directions perpendicular to the X-ray beam. A single S-µXRD measurement had an exposure time of 20 s and was collected using a two-dimensional (2D) area detector (Image Star 9000, Photonic Science) with a 3,056 × 3,056 pixel resolution (31 × 31 µm2 optical pixel size), placed 144 mm behind the sample to give a q range of 0.15–3.46 Å−1 (where q = 4π/λ sinθ). For each specimen, raster mapping measurements were collected at spatial increments of 20 µm in the x–y directions, with corresponding values for beam transmission recorded throughout using a beam stop mounted photo-diode. Additional measurements were taken for the direct beam and empty sample holders to allow the diffraction patterns to be corrected for background and normalization effects. Instrument parameters including the X-ray wavelength and experimental geometry were accurately determined by calibrating to a lanthanum hexaboride (LaB6) (Sigma Aldrich) standard sample. Other experimental parameters are summarized in Supplementary Table 2.
Two-dimensional diffraction images presented in Fig. 4j,k were processed using the DAWN (v.2.27.0, 2022) software package38,39. Diffraction images were normalized with respect to count time, monitor intensity and transmission and background corrected. Data were then remapped to polar coordinates, that is cake remapping, to generate images of azimuthal angle (°) versus q (Å−1) (x versus y) to enable 2D fitting of the (002) diffraction peak located at ~1.83 Å−1. The (002) plane was used to measure the c lattice parameter as a function of location within the sample map and preferred orientation as it is normal to the c axes of the fluorapatite unit cell. Here, preferred orientation is determined from intensity variations in the (002) diffraction ring with respect to the azimuth. To facilitate batch fitting of relatively large diffraction data of highly textured specimens, a machine learning based approach was used to classify data according to preferred orientation (Supplementary Figs. 18 and 19), which demonstrated greater variance in the diffraction images compared with the peak positions in q space. Remapped data were sectioned about the (002) diffraction peak and averaged along the azimuthal axis to produce one-dimensional plots of intensity versus the azimuthal angle (Supplementary Fig. 18a) representing the preferred orientation of constituent crystallite populations. Principal component analysis (PCA) was applied to the orientation data (Supplementary Fig. 18b), following normalization and mean centring, to aid in texture classification as a function of position within the specimen. Orientation data were grouped according to their similarity using k-means clustering applied to the PCA data (Supplementary Fig. 18c), with ~40–50 unique clusters required to classify each S-µXRD map.
To improve data clarity and aid in the identification of orientation peaks, data were reconstructed from their principal components to remove components which explained little variance (only applied for peak identification purposes) and data were subsequently averaged in each cluster. For each averaged data cluster, peak positions and peak widths were determined from the second derivative of the intensity of the orientation data (Supplementary Fig. 18d). Troughs in the second derivative corresponded to underlying peak positions while peak widths were estimated from trough widths, where the width of a second derivative trough is approximately one-third of a peak width in the intensity domain. Values for peak positions and widths were used as parameters to initialize 2D pseudo-Voigt peak models on a plane background to fit the 002 peaks in the original (sectioned) remapped diffraction images for each cluster (Supplementary Fig. 18e). The quality of data fitting was determined by least-squares methods where the goodness-of-fit increased as the chi-square value approached unity. The computational analysis for the processing and fitting pipeline was performed using Python programming software (v.3.9.12, 2022). The 2D peak positions in the x and y axis of the diffraction images yielded the direction of preferred orientation and the (002)-peak position in q space, respectively, for each constituent crystallite population. The positions of the (002) peaks were used to calculate the corresponding unit cell c axis lattice parameters. A hexagonal unit cell was assumed for fluorapatite, where the relationship between the d spacing (d = 2π/q), Miller indices for a given Bragg peak and lattice parameters are given as:
where h, k and l refer to the Miller indices for a particular Bragg peak while a and c represent the relevant lattice parameters. Substituting the (002) indices into equation (1) gives the individual c lattice parameter as a function of d.
Nano-indentation
A V. komodoensis (J94036-2) and tyrannosaurid (UALVP 60555) tooth were cut in longitudinal section to expose the mesial and distal serrations. These were polished using a LaboForce (Struers) grinding and polishing wheel, using P1200, P2000 and P4000 grinding papers and aluminium oxide polishing powders on cloth plates. A Bruker Hysitron Ti950 with a Berkovitch indenter tip was used, equipped with a load-controlled, trapezoidal function (4,000 µN of force, 5 s loading onto the sample, 5 s sustain, 5 s release). Analyses were conducted on regions of interest along mesial and distal serrations of each tooth. Owing to the order of magnitude difference in size and thickness of enamel between the V. komodoensis and tyrannosaurid teeth, different spacings between indents for both samples were used. For V. komodoensis, successive indents were 10 µm apart to ensure that several measurements were taken in the thin enamel. For the tyrannosaurid tooth, indents were spaced 15 µm apart.
Two shed teeth from an A. mississippiensis individual from the Crocodiles of the World Zoo (Brize Norton, UK) were also sectioned longitudinally along the carinae. ‘Tooth 1’ was a posterior, bulbous tooth, whereas ‘tooth 2’ was a conical anterior tooth. These teeth were polished following the same protocol and a series of transects of indentations were taken through the enamel of each tooth (Supplementary Fig. 22). Indents were spaced 15 µm apart using the same loading function as with the other tooth samples (4,000 µN of force, 5 s loading onto the sample, 5 s sustain, 5 s release). Following the LA-ICP-MS experiments on ‘tooth 2’ (Fig. 4g–j), we conducted a second indentation experiment on the outer enamel layers of the tooth to ensure that we indented the iron-rich, slightly pigmented regions in the outer 30–40 µm. We conducted a series of indentations along the pigmented and unpigmented enamel regions just below the two regions we sampled for LA-ICP-MS (Supplementary Fig. 22e,o–q). We recorded a total of 36 indents within the pigmented enamel regions and 38 indents in adjacent, underlying enamel regions. We then compared the indentation hardness and elastic moduli between the unpigmented and pigmented enamel regions using a two-sample Welch test and a pooled variance t-test on the hardness and elastic modulus data, respectively. Effect size was calculated using Cohen’s d (Supplementary Table 4).
Iron-edge X-ray absorption near-edge structure
Fe K-edge XANES measurements were performed at the I-18 beamline (Diamond Light Source, experiment SP35162-1) with a 2 × 2 μm2 beam footprint. XRF mapping was first undertaken to identify Fe-rich regions of interest within the enamel of V. komodoensis, C. porosus and C. canadensis before point XANES measurements (three to five per sample). For XANES, the energy of the incoming beam was scanned from 6.96 to 7.25 keV with increments of 0.5 eV over the pre-edge, edge and post-edge features. Energy calibration was performed using an Fe foil. Spectra were analysed in Athena XAS Data Processing (Demeter v.0.9.26). Spectral standards for ferrihydrite (Fe5HO8•4H2O), magnetite (Fe3O4) and haematite (Fe2O3) were sourced from https://xaslib.xrayabsorption.org (International X-ray Absorption Society) and were energy-normalized using Fe foil measurements before analysis.
Laser ablation inductively coupled mass spectrometry
An Iridia 193 nm ArF*excimer-based LA system (Teledyne Photon Machines) was used, equipped with the cobalt long-pulse ablation cell. The LA system was coupled to a ThermoFisher Scientific iCAPTQ ICP-mass spectrometer (ThermoFisher Scientific) by means of the Aerosol Rapid Introduction System. Full operational parameters for both the Iridia and iCAPTQ ICP-MS are provided in Supplementary Table 5.
Tuning of the instrument settings was performed using a NIST SRM 612 glass certified reference material (National Institute for Standards and Technology), optimizing for low laser-induced elemental fractionation by monitoring 238U+/232Th+, oxide formation rates (<1%) through the 232Th16O+/232Th+ ratio and the sensitivity of 59Co+, 115In+ and 238U+. LA-ICP-MS image was acquired in a fixed dosage mode, with a vertical and horizontal spatial resolution of 5 µm. Imaging was performed on extant crocodylian and fossil tyrannosaurid tooth samples. The selected isotopes of interest 24Mg, 44Ca16O, 56Fe, 66Zn, 88Sr16O, 89Y16O and 138Ba16O were chosen to maximize sensitivity while minimizing isobaric/polyatomic interferences and increasing signal-to-noise ratio. dynamic reaction collision mode with oxygen as the reaction gas was used in the collision reaction cell to reduce the contribution of polyatomic interferences on imaging for all isotopes. The reactive oxygen species measured are proxies for the elements of interest (Ca, Sr, Y and Ba) and are therefore labelled in the LA-ICP-MS maps accordingly. To correct for instrumental drift, a series of NIST 612 standard ablation scans were performed before and after experimental samples.
ICP-MS and positional data were reconstructed to generate elemental images using the hierarchical data format (HDF)-based image processing software (HDIP, Teledyne Photon Machines). A bespoke in-house pipeline, written in Python (v.4), was used to analyse the reconstructed data and produce comparable elemental images. The pipeline consisted of removing negative values, attributed to instrumental noise and replacing with zeros. A mask was applied to each image removing noise and reconstruction artefacts surrounding the samples. All data are presented on the same intensity scale for comparability (95% percentile).
We then used the Keyence digital microscope to generate 2D profiles of the ablated regions and determined that the laser removed 5.4 times more dentine than enamel in the regions of interest, due to material differences between enamel and dentine in the extant crocodylian teeth. A correction factor was applied to the LA-ICP-MS data to account for this difference and generated corrected relative counts for the elements of interest (Supplementary Fig. 8).
V. komodoensis serrations were imaged using LA-ICP-TOF (time-of-flight)-MS using a 193 nm Iridia (Teledyne) coupled with a Vitesse (Nu Instruments). The Iridia was equipped with the cobalt short-pulse ablation cell achieving a single-shot washout of <2 ms. Laser parameters of 500 Hz (repetition rate), dosage of ten shots and a 4 µm spot size were used for imaging. Data were processed using Nu Quant software, HDIP and Python.
Statistics and reproducibility
Electron microscope, S-µXRF and S-µXRD, nano-indentation and LA-MS experiments were all conducted once on different regions of interest on individual teeth. Several of the teeth were used for several experiments to verify elemental and structural results from different methodologies (Supplementary Information).
Animal ethics
All museum specimens are housed in publicly accessible institutions. The fluid-preserved V. komodoensis from the Zoological Society of London in Fig. 1 was made available for study to A.R.H.L. following medical euthanasia at the London Zoo, the reasons for which were unrelated to this study.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data, measurements and images are either in the manuscript or Supplementary Information. Raw synchrotron XRF and XRD datasets are made publicly available through the relevant synchrotron facility 3 years after each experiment under a CC-BY-4 license (see Supplementary Table 2 for experiment numbers).
Code availability
All scripts used to analyse the synchrotron X-ray diffraction data are provided at GitHub https://github.com/Freeman163/LeBlanc.-et-al.-2024.-Nat.-Ecol.-Evol.
References
Brink, K. S. & Reisz, R. R. Hidden dental diversity in the oldest terrestrial apex predator Dimetrodon. Nat. Commun. 5, 3269 (2014).
Whitney, M. R., LeBlanc, A. R. H., Reynolds, A. R. & Brink, K. S. Convergent dental adaptations in the serrations of hypercarnivorous synapsids and dinosaurs. Biol. Lett. 16, 20200750 (2020).
de Andrade, M. B., Young, M. T., Desojo, J. B. & Brusatte, S. L. The evolution of extreme hypercarnivory in Metriorhynchidae (Mesoeucrocodylia: Thalattosuchia) based on evidence from microscopic denticle morphology. J. Vertebr. Paleontol. 30, 1451–1465 (2010).
Abler, W. L. The serrated teeth of tyrannosaurid dinosaurs and biting structures in other animals. Paleobiology 18, 161–183 (1992).
Brink, K. S. et al. Developmental and evolutionary novelty in the serrated teeth of theropod dinosaurs. Sci. Rep. 5, 12338 (2015).
Torices, A., Wilkinson, R., Arbour, V. M., Ruiz-Omenaca, J. I. & Currie, P. J. Puncture–pull biomechanics in the teeth of predatory coelurosaurian dinosaurs. Curr. Biol. 28, 1467–1474 (2018).
Sander, P. M. The Microstructure of Reptilian Tooth enamel: Terminology, Function and Phylogeny München Geowissenschaftliche Abhandlungen Vol. 38 (Verlag Dr. Friedrich Pfeil, 1999).
Enax, J. et al. Characterization of crocodile teeth: correlation of composition, microstructure and hardness. J. Struct. Biol. 184, 155–163 (2013).
D’Amore, D. C. & Blumenschine, R. J. Komodo monitor (Varanus komodoensis) feeding behavior and dental function reflected through tooth marks on bone surfaces and the application to ziphodont paleobiology. Paleobiology 35, 525–552 (2009).
D’Amore, D. C. A functional explanation for denticulation in theropod dinosaur teeth. Anat. Rec. 292, 1297–1314 (2009).
Janeczek, M. et al. Macroanatomical, histological and microtomographical study of the teeth of the Komodo dragon (Varanus komodoensis)—adaptation to hunting. Biology 12, 247 (2023).
Maho, T. & Reisz, R. R. Exceptionally rapid tooth development and ontogenetic changes in the feeding apparatus of the Komodo dragon. PLoS ONE 19, e0295002 (2024).
Kaye, T. G. et al. Laser-stimulated fluorescence in paleontology. PLoS ONE 10, e0125923 (2015).
Dumont, M., Tütken, T., Kostka, A., Duarte, M. J. & Borodin, S. Structural and functional characterization of enamel pigmentation in shrews. J. Struct. Biol. 186, 38–48 (2014).
Gordon, L. M. et al. Amorphous intergranular phases control the properties of rodent tooth enamel. Science 347, 746–750 (2015).
Müller, W. et al. Enamel mineralization and compositional time-resolution in human teeth evaluated via histologically-defined LA-ICPMS profiles. Geochim. Cosmochim. Acta 255, 105–126 (2019).
Georgalis, G. L., Mennecart, B. & Smith, K. T. First fossil record of Varanus (Reptilia, Squamata) from Switzerland and the earliest occurrences of the genus in Europe. Swiss J. Geosci. 116, 9 (2023).
Georgalis, G. L., Cernansky, A. & Klembara, J. Osteological atlas of new lizards from the Phosphorites du Quercy (France), based on historical, forgotten, fossil material. Geodiversitas 43, 219–293 (2021).
Hocknull, S. A. et al. Dragon’s paradise lost: palaeobiogeography, evolution and extinction of the largest-ever terrestrial lizards (Varanidae). PLoS ONE 4, e7241 (2009).
Smith, K. T. A diverse new assemblage of Late Eocene squamates (Reptilia) from the Chadron Formation of North Dakota, USA. Palaeontol. Electron. 9, 1–44 (2006).
Georgalis, G. L., Villa, A., Ivanov, M., Vasilyan, D. & Delfino, M. Fossil amphibians and reptiles from the Neogene locality of Maramena (Greece), the most diverse European herpetofauna at the Miocene/Pliocene transition boundary. Palaeontol. Electron. 22, 1–99 (2019).
Hwang, S. H. The evolution of dinosaur tooth enamel microstructure. Biol. Rev. 86, 183–216 (2011).
Chen, J., LeBlanc, A. R. H., Jin, L., Huang, T. & Reisz, R. R. Tooth development, histology and enamel microstructure in Changchunsaurus parvus: implications for dental evolution in ornithopod dinosaurs. PLoS ONE 13, e0205206 (2018).
Godefroit, P. et al. Extreme tooth enlargement in a new Late Cretaceous rhabdodontid dinosaur from Southern France. Sci. Rep. 7, 13098 (2017).
Feng, R. et al. Chemical and structural information from the enamel of a Troodon tooth leading to an understanding of diet and environment. Appl. Spectrosc. 70, 1883–1890 (2016).
Vallcorba, O. et al. Synchrotron X-ray microdiffraction to study dental structures in Cretaceous crocodylomorphs. Cretac. Res. 128, 104960 (2021).
Al-Jawad, M. et al. 2D mapping of texture and lattice parameters of dental enamel. Biomaterials 28, 2908–2914 (2007).
D’Amore, D. C., Moreno, K., McHenry, C. R. & Wroe, S. The effects of biting and pulling on the forces generated during feeding in the Komodo dragon (Varanus komodoensis). PLoS ONE 6, e26226 (2011).
Smith, T. & Codrea, V. Red iron-pigmented tooth enamel in a multituberculate mammal from the Late Cretaceous Transylvanian ‘Hateg Island’. PLoS ONE 10, e0132550 (2015).
Selvig, K. A. & Halse, A. The ultrastructural localization of iron in rat incisor enamel. Scand. J. Dent. Res. 83, 88–95 (1975).
Sparks, N. H. C., Motta, P. J., Shellis, R. P., Wade, V. J. & Mann, S. An analytical electron microscopy study of iron-rich teeth from the Butterflyfish (Chaetodon ornatissimus). J. Exp. Biol. 151, 371–385 (1990).
Randall, M. Electron microscopical demonstration of ferritin in the dental epithelial cells of urodeles. Nature 210, 1325–1326 (1966).
Anderson, M. A. & Miller, B. T. Early iron deposition in teeth of the streamside salamader, Ambystoma barbouri. J. Herpetol. 45, 336–338 (2011).
Schubert, B. W. & Ungar, P. S. Wear facets and enamel spalling in tyrannosaurid dinosaurs. Acta Palaeontol. Pol. 50, 93–99 (2005).
Jacobsen, A. R. Feeding behaviour of carnivorous dinosaurs as determined by tooth marks on dinosaur bones. Hist. Biol. 13, 17–26 (1998).
Hunter, J. D. Matplotlib: a 2D graphics environment. Comput. Sci. Eng. 9, 90–95 (2007).
Sawhney, K. J. S. et al. A test beamline on Diamond Light Source. AIP Conf. Proc. 1234, 387–390 (2010).
Basham, M. et al. Data analysis WorkbeNch (Dawn). J. Synchrotron Radiat. 22, 853–858 (2015).
Filik, J. et al. Processing two-dimensional X-ray diffraction and small-angle scattering data in ıt DAWN 2. J. Appl. Crystallogr. 50, 959–966 (2017).
Vidal, N. et al. Molecular evidence for an Asian origin of monitor lizards followed by Tertiary dispersals to Africa and Australasia. Biol. Lett. 8, 853–855 (2012).
Head, J. J., Barrett, P. M. & Rayfield, E. J. Neurocranial osteology and systematic relationships of Varanus (Megalania) prisca Owen, 1859 (Squamata: Varanidae). Zool. J. Linn. Soc. 155, 445–457 (2009).
Acknowledgements
For assistance with specimen acquisition and loans, we thank P. J. Currie, H. Gibbins, C. Coy (University of Alberta) and L. Rowden (Zoological Society of London, London Zoo). We also thank J. Streicher and S. Maidment (NHMUK), G. Sales and S. Ryder (MoLS), D. Kizirian and L. Vonnahme (AMNH), M. S. Y. Lee, M. Hutchinson and M. -A. Binnie (SAM) for collections access. E. Radvar (King’s College London), P. Machado and L. Allison (CUI) assisted with SEM imaging and we thank the London Metallomics Facility for LA-ICP-MS and Nu Instruments and Teledyne for access to ICP-TOF-MS (Vitesse and Iridia). P. Shaw (Nu Instruments) and G. Foster, J. A. Milton (University of Southampton) and T. Stewart (King’s College London) assisted with the Vitesse imaging. We are grateful to the synchrotron beamline staff E. Villalobos, H. Castillo Michel (ESRF-ID21), L. Bouchenoire (ESRF-BM28) and O. Fox (DLS-B16) for their assistance with the synchrotron experiments. O.A. was supported by the Natural Sciences and Engineering Research Council of Canada (grant no. 2017-05862). A.R.H.L. was supported by a Crispian Scully Research Award from the Oral and Dental Research Trust and the British Society of Dental Research. F. Giuliani was supported by the Engineering and Physical Sciences Research Council (grant no. EP/V007661/1). M. Pittman was supported by The School of Life Sciences of The Chinese University of Hong Kong. This study is part of a project that received funding from the European Research Council under the European Union’s Horizon 2020 research and innovation programme Horizon (grant agreement no. 894331 to A.R.H.L. and O.A.). Part of this study also received funding from the European Research Council’s Horizon 2020 programme (grant agreement no. 851705 to D.L.).
Author information
Authors and Affiliations
Contributions
A.R.H.L. and O.A. conceived the project. A.R.H.L., A.P.M., S. Sirovica, M.A-.J. and O.A. collected synchrotron diffraction and fluorescence data. A.P.M. and S. Sirovica. processed fluorescence and diffraction data. A.R.H.L., S.W., F.G. and C.M.M. collected electron microscopy data. M.P. and T.G.K. collected LSF data. A.R.H.L. and D.C.D. collected data on pigmented enamel in lizards. A.P.M. collected laser ablation mass spectrometry data. D.C.D., C.C., C.S., S. Spiro, B.T., J.C. and D.L. provided specimens and data for extant reptile teeth. A.R.H.L. collected the histology data. A.R.H.L. and D.L. conducted nano-indentation experiments. All authors assisted in interpreting the data and edited previous drafts of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Ecology & Evolution thanks Georgios Georgalis and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
LeBlanc, A.R.H., Morrell, A.P., Sirovica, S. et al. Iron-coated Komodo dragon teeth and the complex dental enamel of carnivorous reptiles. Nat Ecol Evol (2024). https://doi.org/10.1038/s41559-024-02477-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41559-024-02477-7
- Springer Nature Limited