
Abstract
Identifying the mechanisms of action of anticancer drugs is an important step in the development of new drugs. In this study, we established a comprehensive screening platform consisting of 68 oncogenes (MANO panel), encompassing 243 genetic variants, to identify predictive markers for drug efficacy. Validation was performed using drugs that targeted EGFR, BRAF, and MAP2K1, which confirmed the utility of this functional screening panel. Screening of a BRCA2-knockout DLD1 cell line (DLD1-KO) revealed that cells expressing SMO and GLI1 were resistant to olaparib. Gene set enrichment analysis identified genes associated with DNA damage repair that were enriched in cells overexpressing SMO and GLI1. The expression of genes associated with homologous recombination repair (HR), such as the FANC family and BRCA1/2, was significantly upregulated by GLI1 expression, which is indicative of PARP inhibitor resistance. Although not all representative genes of the nucleotide excision repair (NER) pathway were upregulated, NER activity was enhanced by GLI1. The GLI1 inhibitor was effective against DLD1-KO cells overexpressing GLI1 both in vitro and in vivo. Furthermore, the combination therapy of olaparib and GLI1 inhibitor exhibited a synergistic effect on DLD1-KO, suggesting the possible clinical application of GLI1 inhibitor targeting cancer with defective DNA damage repair. This platform enables the identification of biomarkers associated with drug sensitivity, and is a useful tool for drug development.
Similar content being viewed by others

Introduction
Treatments targeting specific genes and proteins that help cancer cells survive and proliferate have proven effective. However, not all patients with targeted lesions respond to therapy; various factors such as DNA alterations, gene expression, and epigenetic marks contribute to drug resistance. Since drug resistance invariably occurs in advanced cancers, it is essential to identify the resistance mechanism in order to administer the appropriate drug to the patient.
Currently, molecular targeted therapies vary by tumor type, making the options diverse and complex. In the case of lung cancer, EGFR tyrosine kinase inhibitors (TKIs) have varying effects on tumors with different EGFR mutations; therefore, effective agents must be selected for individual mutations [1,2,3,4]. In the case of melanoma, BRAF inhibitors are commonly administered to treat tumors with BRAF mutations. As the RAS/CRAF/MEK pathway is a known resistance mechanism to BRAF inhibitors, combination therapy using BRAF and MEK inhibitors has been introduced [5,6,7]. In addition to targeting oncogenes that drive tumor growth, targeting loss-of-function mutations in tumor suppressor genes is another strategy for anticancer drug development. Tumor cells deficient in BRCA, a double-strand break (DSB) repair factor, are sensitive to PARP inhibitors (PARPi) through a mechanism of synthetic lethality [8,9,10,11].
Several causes for resistance to PARPi have been reported [12, 13]. The most common mechanism is the restoration of homologous recombination (HR), such as the appearance of a BRCA reversion mutation [14, 15] and increased levels of RAD51C [16]. Other mechanisms associated with PARPi resistance are switching to alternative repair mechanisms, such as non-homologous end joining (NHEJ), replication fork stabilization, drug efflux, and reduced PARP expression. Inactivation of 53BP1, replication timing regulatory factor 1 (RIF1), mitotic arrest-deficient 2-like 2 (MAD2L2), the shieldin complex that inhibits DNA end resection, and dynein light chain LC8-Type 1 (DYNLL1) are known to be involved in partial restoration of HR [17,18,19,20,21].
As drug resistance mechanisms are complicated and remain largely unknown, we established a comprehensive screening platform to identify predictive markers for targeted therapies using the mixed-all-nominated-mutants-in-one (MANO) method, which was previously developed in our laboratory2. The screening platform was validated using several targeted therapies with known predictive biomarkers and was utilized to explore novel mechanisms for olaparib resistance.
Results
Assessment of the transforming activity of 243 variants in the MANO panel
Cancer-related genes (n = 68) were selected based on the COSMIC v90 database of somatic mutations in cancer. Wild-type and 1–4 hotspot variants were selected for each gene to generate a total of 243 variants (MANO panel). A 3T3 focus formation assay and a Ba/F3 transformation assay were performed to assess the transforming potential of the variants in the MANO panel. The results of the transformation assay are summarized in Fig. 1A, Supplementary Fig. 2, and Supplementary Table S6 with the information of the variants, including the number of COSMIC counts, variant type, and OncoKB annotation. Based on the focus formation assay, 43 variants (17.7%) had an FFA score of 2 (mild transformation), 20 (8.2%) had an FFA score of 3 (moderate transformation), 32 (13.2%) had an FFA score of 4 (severe transformation), and the remaining 148 variants (61.0%) had FFA scores of 1 (no transformation). Among the variants with FFA scores of 2–4, 17 (7.0%) were identified for the first time as transforming competent. Based on the Ba/F3 transformation assay, 47 (19.3%) variants resulted in IL-3-independent growth.

Information for each variant. i) The number of COSMIC database counts, ii) mutation type, iii) 3T3 FFA scores, iv) transformation in Ba/F3 cells, and v) OncoKB annotation.
Identification of predictive biomarkers using the MANO panel
To validate the capability of the MANO panel to assess the drug sensitivity of the variants, 3T3 cells and their variants were treated with osimertinib, a third-generation EGFR TKI (Fig. 2A). According to the assessment, known TKI-sensitizing mutations, such as EGFR exon 19 deletions (E746_A750del) and L858R, were sensitive to osimertinib (IC50 < 5 nM), whereas known TKI-resistant mutations, including exon 20 insertions (V769_D770insASV), were resistant to osimertinib (IC50 > 100 nM). Similarly, some ERBB2 variants, such as S310F and V777L were moderately sensitive (IC50 < 50 nM), whereas L755S was relatively resistant to osimertinib (IC50 > 50 nM), which was consistent with a previous report [22].

A The sensitivity to osimertinib in the screening panel using 3T3 Cells. Cells were treated with osimertinib for four days. KRAS_G12D was used as the control for resistant variant. Representative cell viability is color-coded. B Screening panel using PC9 cells. Cells were treated with osimertinib for four days. Each row indicates individual variants. The data for EGFR variants is enlarged. C Cell viability assay of osimertinib using 3T3 cells. 3T3 cells introduced the indicated variants were treated for four days. Cell viability is shown as the mean with SD (error bars). D Cell viability assay of osimertinib using PC9 cells. PC9 cells introduced the indicated variants were treated for four days. Cell viability is shown as the mean with SD (error bars). E Screening panel for the combination treatments using A375 cells. Cells were treated with trametinib and dabrafenib for 4 days. KRAS as the control and CRAF variants are described in the red frame. F Cell viability assay of dabrafenib plus trametinib in A375 cells. Viability is presented in a two-dimensional heat map. Details of the drug sensitivity data (A, B, E) are provided in Supplementary Table S6.
Next, an assay using PC9, a human lung cancer cell line, was conducted to validate the capability of the MANO panel in identifying drug resistance markers. PC9 harbored EGFR delE746-750, a driver mutation, and was sensitive to EGFR TKIs. PC9 cells expressing 243 variants were treated with osimertinib for four days. Similar to the assay in 3T3 cells, EGFR exon 19 deletions and L858R were sensitive (IC50 < 5 nM), whereas EGFR exon 20 insertion was resistant (IC50 > 100 nM). In addition, HRAS and RET variants were resistant (IC50 > 1 μM) to osimertinib, whereas CSF1R variants were partially resistant (IC50 = 11 nM for CSF1R WT) (Fig. 2B). Individual cell viability assays for several variants confirmed the results of screening (Fig. 2C, D, Supplementary Fig. S3A, B).
The A375 melanoma cell line harboring the BRAF V600E mutation was used to test the utility of the screening panel for evaluating drug combinations, because combination therapies of BRAF inhibitor and MEK inhibitor are in practice for this indication. The cells were treated with the BRAF inhibitor dabrafenib and MEK inhibitor trametinib. CRAF variants were sensitive to trametinib (IC50 < 10 nM) but resistant to dabrafenib (IC50 > 500 nM) (Fig. 2E). In a previous study, trametinib was used to treat cancer cells harboring CRAF variants, which are known to be dabrafenib resistant [5]. The results of the individual cell viability assays were concordant with the screening results (Fig. 2F).
Identification of novel resistance markers for PARPi
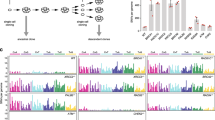
Screening with DLD1-KO was performed to identify novel mechanisms of resistance to olaparib, a PARPi (Fig. 3A). Interestingly, MYCN and SMO variants were resistant to olaparib. Individual assays confirmed that SMO variants conferred resistance, whereas MYCN and MYC variants did not significantly change the sensitivity to olaparib (Supplementary Fig. S4). GLI1, a transcription factor downstream of SMO, also conferred resistance to PARPi (Fig. 3B). In addition, olaparib resistance induced by SMO and GLI1 was assessed in MIAPaCa2-KO cells, a BRCA1 knockout pancreatic cell line, and UWB1.289, an ovarian carcinoma cell line with BRCA1 deficiency. Overexpression of SMO and GLI1 rendered MIAPaCa2-KO and UWB1.289 cells resistant to PARPi (Fig. 3C).

A The olaparib sensitivity of the variants in the screening panel using DLD1-KO Cells. Cells were treated with olaparib for six days. Representative cell viability is color-coded. B Cell viability assay of three PARPi using DLD1-KO cells. DLD1-KO cells introduced the indicated variants were treated for six days. Cell viability is shown as the mean with SD (error bars). C Cell viability assay of olaparib using MIAPaCa2-KO and UWB1.289, a BRCA1 deficient cell line. The cell lines introduced the indicated variants were treated for six days. Cell viability is shown as the mean with SD (error bars). D Phosphorylation of H2AX was determined by western blot analysis. Relative ratios were calculated using GAPDH as a control and comparing to the value of DLD1-KO treated with olaparib (1 μM) for 48 or 72 h. E Immunostaining of γH2AX and 53BP1. Cells were treated with 1 μM olaparib for 48 h. A representative fluorescence image is shown on the left. The white scale bars indicate 10 µm. Other images (DAPI and 53BP1) are shown in Supplementary Fig. 5B. The quantitative values of γH2AX are indicated on the right. The number of foci per nucleus is plotted with the mean and SD (error bars). The p value was calculated (*p < 0.05; Welch’s t-test).
SMO and GLI1 induce PARPi resistance by reducing DNA double-strand breaks
To determine the mechanism of resistance to olaparib, H2AX phosphorylation, an indicator of DSB, was measured by western blot analysis (Fig. 3D and Supplementary Fig. S5A). Overexpression of SMO and GLI1 reduced olaparib-induced H2AX phosphorylation in DLD1-KO cells. Furthermore, the number of foci for γH2AX and 53BP1, key mediators of DSB repair, was quantified by immunofluorescence (IF). After 48-h of treatment with olaparib, fewer γH2AX foci per nucleus were observed in DLD1-KO cells overexpressing SMO and GLI1 overexpression (DLD1-KO-SMO and DLD1-KO-GLI1) than in DLD1-KO cells (*p < 0.05; Welch’s t-test) (Fig. 3E and Supplementary Table S7). The same result was observed for IF of 53BP1 (Supplementary Fig. S5B, C).
SMO and GLI1 regulate the expression of genes associated with DNA damage repair
RNA-seq was conducted to identify the genes regulated by SMO and GLI1 during olaparib treatment. GSEA was performed to compare DLD1-KO, DLD1-KO-SMO, and DLD1-KO-GLI1. Compared with DLD1-KO cells, 456 and 419 gene sets were enriched in SMO or GLI1 expressing cells, respectively, and 343 gene sets were common (Fig. 4A). Of the commonly enriched gene sets, 20 (5.8%) were associated with DNA damage repair, 29 (8.5%) with DNA replication, and 22 (6.4%) with the cell cycle (Fig. 4B and Supplementary Table S8). Representative gene sets for DNA damage repair are shown in Fig. 4B. Without PARPi treatment, fewer gene sets associated with DNA damage repair were enriched in the DLD1-KO-SMO and DLD1-KO-GLI1 cells (Supplementary Fig. S6), suggesting that the functions of SMO and GLI1 are more apparent when DNA damage repair is required. Only three gene sets related to DNA damage repair or DNA replication were enriched in both cell types without olaparib treatment (Supplementary Table S9).

A GSEA was performed to identify the gene sets associated with SMO and GLI1. Compared with DLD1-KO cells, 456 and 419 gene sets were enriched in SMO or GLI1-expressing cells, respectively, and 343 gene sets were in common. The analysis was conducted in the presence of olaparib (1 μM) for 48 h. B The representative gene sets for DNA damage repair. DNA damage repair gene sets enriched in both DLD1-KO-SMO and DLD1-KO-GLI1 are shown. The detailed gene sets are listed in supplementary Table S8. The mRNA expression after olaparib treatment for 48 h. The expression of genes associated with DNA double-strand break repair (C) and the genes fundamentally involved in DNA repair, DNA damage checkpoints, and DNA replication (D) were evaluated (*p < 0.05; Student’s t-test). The expression of other genes is shown in Supplementary Fig. 7. E The expression change by the knockdown of GLI1 (left panel) and GANT61 treatment (right panel). Expression measured by real-time PCR is shown as the mean with SD (error bar). Fold change was calculated relative to the siControl or no treatment group. F NER activity assay. CPD, an UV-irradiated product, was measured by immunostaining. Fold change was calculated relative to the value immediately after irradiation as the control. Significant differences were in KO vs. KO + GLI1 and WT vs. KO + GLI1 (*p < 0.05; Welch’s t-test).
Next, we analyzed the expression of individual genes associated with DNA double-strand break repair, such as BRCA1, FANCD2, and PALB2. These genes as well as FANCA, FANCB, FANCC, FANCG, and RAD51C were upregulated in DLD1-KO-SMO and DLD1-KO-GLI1 cells (Fig. 4C and Supplementary Fig. S7A). Furthermore, the expression of the genes fundamentally involved in DNA repair, DNA damage checkpoints, and DNA replication, such as RPA1, RPA2, and PCNA, was also upregulated by SMO and GLI1 overexpression (Fig. 4D), whereas XPC was the only gene among the NER genes that were significantly upregulated in both DLD1-KO-SMO and DLD1-KO-GLI1 cells (Supplementary Fig. S7B).
To confirm the involvement of GLI1 in the regulation of FANCD2, GLI1 knockdown by siRNA was performed in SJSA1, a cell line with high GLI1 expression based on the DepMap (https://depmap.org/portal/ccle/). Not only FANCD2, but also BRCA1 and BRCA2 expression, were downregulated by siGLI1 as well as treatment with GANT61, which is a SMO inhibitor (Fig. 4E). In addition, the correlation between GLI1 expression and the expression of genes associated with DNA damage repair was determined using the DepMap RNA expression data. Cell lines with high GLI1 expression tended to have a high expression of genes associated with DNA damage repair (Supplementary Fig. S8). Above, GLI1 knockdown also repressed the expression of FANCD2, BRCA1 and BRCA2 (p = 1.3 × 10−3, 5.7 × 10−5, 5.7 × 10−3, respectively; Student’s t-test) in DLD1 cells, whose GLI1 expression is not high as SJSA1 according to DepMap data (Supplementary Fig. S9).
Similar to previous reports on the relationship between GLI1 and the NER pathway [23, 24], genes associated with NER were partially upregulated by SMO or GLI1 overexpression. To evaluate the change in NER activity by GLI1 expression, an NER activity assay was conducted by immunostaining CPD, a UV-irradiated product. GLI1 overexpression significantly reduced the CPD amount induced by UV irradiation (*p < 0.05; Welch’s t-test) (Fig. 4F).
GLI1 expression in ovarian cancer specimens
To assess the clinical relevance of the GLI1 pathway, we examined three cases of ovarian cancer with acquired resistance to olaparib (Supplementary Fig. S10A–C). RNA-seq of these specimens before and after olaparib treatment revealed that GLI1 expression increased following olaparib treatment in two cases. IPA analysis using RNA expression data was performed to identify the pathways that were activated by the treatment. A total of 3,550 genes were upregulated (fold change > 1.5) after treatment in at least two cases (Supplementary Fig. S10D). The basal cell carcinoma pathway, which includes the hedgehog (Hh) signaling pathway to which GLI1 belongs, was activated following olaparib treatment (Supplementary Fig. S10E and Supplementary Table S10).
In the GENIE Cohort v13.0 database, containing 107,543 samples, GLI1 amplification was identified in 704 cases (0.65%) and GLI1 mutations in 2,752 cases (2.5%); however, most of the GLI1 mutations were variants of unknown significance, and only 15 cases had R380Q, a known oncogenic mutation [25]. The frequency of GLI1 alterations is high in skin cancer, bone and soft tissue tumors, and tumors of the nervous system (Supplementary Fig. S11 and Supplementary Table S11).
Assessment of combination therapy of PARPi with GLI1 inhibitor
To determine whether GLI1 suppression potentiates olaparib treatment, the sensitivity of cells to olaparib in combination with GANT61 was evaluated (Fig. 5A). Treatment with 10 µM GANT61 affected both DLD1-KO and DLD1-KO-GLI1 cells. Olaparib at 50 nM and GANT61 at 5 µM exhibited a synergistic effect in DLD1-KO cells (synergy score >10 in ZIP, HAS and Bliss methods) (Supplementary Fig. S12 and Supplementary Table S12).

A The sensitivity of olaparib in combination with GANT61. The cell viability of indicated cells treated with several concentration of olaparib and 10 µM of GANT61 is shown as the mean with SD (error bars). In vivo assessment of PARPi and GLI1 inhibitor. Assay for DLD1-WT cells (B), DLD1-KO cells (C) and DLD1-KO + GLI1 cells (D) are shown. Tumor growth of each group is shown in the left panel. The weight of the resected tumor from the indicated groups is shown in the right panel. The tumor volume and weight are shown as the mean with SD (error bars). The p value was calculated (#p < 0.05; *p < 0.01; **p < 0.001; Student’s t-test). E The relationship between pathways and drugs examined in this study is shown as a schema. Overexpression of SMO and GLI activates the transcriptional function of GLI1 resulting in the promotion of DNA repair through the Fanconi anemia and NER pathway, which renders cells resistant to PARPi.
DLD1, DLD1-KO, and DLD1-KO-GLI1 were injected into the flanks of mice to assess the effects of a PARPi and a GLI1 inhibitor in combination. Mice were treated with olaparib, GANT61, or a combination of both. No loss in body weight was observed throughout the experiment (Supplementary Fig. S13A). Mice with the DLD1 parental cell tumor were sacrificed on day 23 because of rapid tumor growth, whereas observations of the other mice continued until day 42. For the DLD1 parental cells, no significant differences in tumor volume (tv) and tumor weight (tw) were observed regardless of the treatment (Fig. 5B and Supplementary Fig. S13B). DLD1-KO tumors significantly responded to olaparib (tv; p = 1.3 × 10−3, tw; p = 8.5 × 10−3) as well as the combination (tv; p = 1.0 × 10−3, tw; p = 7.8 × 10−4) on day 42 (Fig. 5C). For DLD1-KO-GLI1 tumors, olaparib or GANT61 treatment did not significantly reduce tv and tw compared with vehicle treatment, whereas the combination therapy significantly inhibited tumor growth (tv; p = 1.2 × 10−3, tw; p = 6.0 × 10−3) (Fig. 5D).
The relationship between the pathways and drugs examined in this study is shown as a schema (Fig. 5E). In summary, overexpression of SMO and GLI activates the transcriptional function of GLI1. As a result, DNA repair is promoted through the Fanconi anemia and the NER pathway, rendering cells resistant to PARPi. Therefore, the inhibition of GLI1 may suppress DNA repair pathway activation and have a synergistic effect with PARPi.
Discussion
We constructed the MANO panel as a platform to search for predictive markers in variants representing cancer-related genes. This is the first study to examine the feasibility of the MANO method for evaluating the functions of variants consisting of multiple genes, whereas the method has been used to individually evaluate variants of EGFR, ERBB2, MAP2K1, FGFR1-3, BRCA2 and sarcoma-related fusion genes [1, 22, 26,27,28,29]. The sensitive and resistant markers to osimertinib and the combination of dabrafenib and trametinib were consistent with previous reports. CSF1R variants were first identified to be partially resistant to osimertinib. Interestingly, even high concentrations of osimertinib could not eradicate tumor cells with CSF1R variants, suggesting that it may be involved in preventing apoptosis in the subclone of the tumor (Supplementary Fig. S3B).
The MANO panel identified GLI1 as a novel resistance marker for PARPi. GLI1 is a downstream transcription factor of Hh signaling that regulates multiple cellular functions such as cell differentiation, stem cell maintenance, and regeneration. The Hh signaling pathway consists of the ligands sonic, Indian, and desert hedgehog (Shh, Ihh, and Dhh, respectively) and the cell surface molecules Patched (PTCH) and Smoothened (SMO). In the absence of Hh ligands, PTCH suppresses SMO; however, upon ligand binding to PTCH, the SMO protein activates the transcription factor GLI1, which in turn translocates to the nucleus, resulting in the expression of Hh induced genes [30, 31].
Gene expression analysis using GSEA revealed that GLI1 and SMO were associated with HR and NER, which we hypothesized resulted in cancer cell resistance to olaparib. FANCD2 has a binding site for GLI1 (5′-GACCACCCA-3′) in its promoter region and may be transcriptionally regulated by GLI1 [32, 33]. In fact, increased expression of FANCD2 by GLI1 was observed (Fig. 4E). BRCA1 and BRCA2 expression was also altered by GLI1, although there was no putative binding site for GLI1 in their promoters, suggesting indirect regulation.
Although previous studies have reported that NER is activated by GLI1 [23, 34], our data indicated that GLI1 activates NER without increasing the transcription of representative factors for NER, such as XPA, ERCC1, ERCC2, ERCC3, and ERCC4 (Fig. 4D and Supplementary Fig. S7). NER activation may be caused by an increase in RPA1, RPA2, and PCNA, which are involved in various DNA damage repair pathways including NER. Interestingly, changes in gene expression regulated by GLI1 were more evident when the cells were treated with olaparib, suggesting that a specific mechanism may exist to promote the function of GLI1 as a transcription factor in response to DNA damage.
While our data revealed that GLI1 was mainly involved in HR and NER, DNA damage was repaired by at least five major pathways: base excision repair (BER), NER, mismatch repair (MMR), HR, and NHEJ. HR is a precise repair mechanism associated with a few errors, whereas NHEJ is an error-prone pathway that leads to genetic aberrations, chromosomal instability, cell cycle arrest, and apoptosis [35, 36]. When either pathway is defective, homology repair by microhomology-mediated end-joining (MMEJ), known as alteration-NHEJ, serves as a backup pathway but can also cause mutations resulting from DNA defects [37,38,39]. BER, NER, and MMR are activated in response to DNA single strand breaks (SSBs) [40]. Other pathways, such as the Fanconi pathway, are activated by DNA interstrand crosslinks, which are often induced by anticancer drugs [41]. The translesion synthesis (TLS) pathway is activated in conjunction with NER to respond to the replication stress caused by UV products and anticancer drugs [42, 43]. GLI1 involvement in other DNA repair pathways remains to be investigated.
In clinical samples, despite only a slight increase in SMO and GLI1 expression in olaparib-resistant cases, basal cell carcinoma signaling, which is closely associated with Hh signaling, was significantly activated. Although the frequency of SMO and GLI1 gene alterations in solid tumors is quite low according to the GENIE Cohort v13.0 database, an association with Hh signaling has been reported in ovarian [44, 45], breast [46], colorectal cancer [47], and other tumor types [48].
With respect to treatment with a GLI1 inhibitor, two intriguing insights were evident. First, GLI1 inhibitor monotherapy was effective against DLD1-KO cells, suggesting that GLI1 inhibition may induce apoptosis in BRCA-deficient cells by downregulating HR pathway activity. Second, the combination of a GLI1 inhibitor and olaparib was effective against DLD1-KO-GLI1, which indicates that GLI1 inhibitors may be used in olaparib-resistant cases with high GLI1 expression. Combination therapies have been addressed in previous studies [33, 34, 49].
This study had some limitations. First, because the genes were overexpressed by retrovirus infection, the expression level was far higher than that of endogenous expression and may not recapitulate the actual features of the variants. Therefore, individual experiments should be performed to validate the screening results. Second, although a mild synergistic effect was observed in the combinational treatment of GANT61 and olaparib, we would like to investigate the synergy using Patient-derived xenograft model in the next study. Third, the incidence of olaparib resistance acquired by GLI1 activation remains unknown.
In conclusion, we constructed a system to screen predictive markers of targeted therapies. Using this screening method, GLI1 was found to be involved in resistance to PARPi by enhancing the DNA damage repair capacity, which could be reversed with a GLI1 inhibitor. Therefore, GLI1 inhibitors may be effective in the treatment of BRCA1-deficient tumors. The MANO panel is expected to be a basic platform for future drug development.
Materials and methods
Cell lines
Human embryonic kidney (HEK) 293T cells, mouse 3T3 fibroblasts, Ba/F3 (IL-3-dependent mouse Pro-B cells), A375 (human melanoma), UWB1.289 (BRCA1-deficient ovarian cancer), UWB1.289 + BRCA1 (BRCA1-overexpressed UWB1.289), SJSA1 (human osteosarcoma), and MIAPaCa2 (human pancreatic carcinoma) cells were obtained from the American Type Culture Collection (Manassas, VA, USA). PC9 (human lung adenocarcinoma) was provided by the RIKEN BRC through the National BioResource Project of the MEXT/AMED, Japan. MIAPaCa2 cells with artificially knocked out BRCA1 (MIAPaCa2-KO) were kindly provided by Dr. Yuka Sasaki. DLD1 (human colorectal adenocarcinoma) and DLD1 cells with artificially knocked-out BRCA2 (DLD1-KO) were purchased from Horizon Discovery. 293T, 3T3, A375, and MIAPaca2 cells were cultured in Dulbecco’s Modified Eagle’s medium-F12 (DMEM-F12) supplemented with fetal bovine serum (FBS), and 2 mmol/L glutamine (Thermo Fisher Scientific, Waltham, MA, USA). PC9, SJSA1 and DLD1 cells were cultured in RPMI 1640 medium (Thermo Fisher Scientific) supplemented with 10% FBS and 2 mM glutamine. Ba/F3 cells were cultured in RPMI 1640 medium supplemented with 10% FBS, 2 mmol/L glutamine, and mouse IL-3 (20 U/mL; Sigma-Aldrich, St. Louis, MO, USA). UWB1.289 cells were cultured in 50% RPMI 1640 medium and a 50% Mammary Epithelial cell Growth Medium Bullet Kit (Lonza, Basel, Switzerland) supplemented with 3% FBS and 2 mmol/L glutamine.
Establishment of a retroviral vector with random barcodes
The pCX6 vector was created by inserting random 10 base pair (bp) DNA barcode sequences upstream of the start codon of selected genes in the pCX4 vector [50] (Supplementary Table S1). All plasmids were constructed using the GeneArt Gene Synthesis system (Thermo Fisher Scientific) and subjected to next-generation sequencing to confirm their sequences. Three clones with individual barcodes were constructed to obtain triplicate data for each assay.
Creation of retrovirus and infection into cell lines
The plasmids were transduced with packaging plasmids (Takara Bio, Shiga, Japan) into HEK293T cells to produce recombinant retroviral particles. The 3T3 cells and cancer cells were infected with ecotropic recombinant retroviruses using 4 µg/mL Polybrene (Sigma-Aldrich) for 24 h. Ba/F3 cells were seeded on RetroNectin-coated (Takara Bio) plates and infected with retroviruses in a medium supplemented with 20 U/mL of IL-3.
3T3 focus formation assay and Ba/F3 transformation assay
To evaluate transforming potential, 3T3 cells expressing the variants were cultured in DMEM-F12 supplemented with 5% bovine calf serum for two weeks, and the cells were stained with Giemsa solution [51]. The transforming activity of each variant was measured using focus formation assay (FFA) score. This score classified the focus-forming potential into the following four groups: score 1 (negative), no focus formation; score 2 (mild), partial focus formation of transformed cells; score 3 (moderate), diffusely transformed cells piled up in bundles; and score 4 (severe), round-shaped and anchorage-independent foci diffusely observed. Ba/F3 cells have also been used to screen for oncogenic variants [52]. Cells expressing a transforming variant should proliferate in the culture medium without IL-3 supplementation.
The MANO method
The schema for the MANO method is shown in Supplementary Fig. S1. For this method, a retroviral vector stably integrates individual genes into the genome of cells with a 10-bp barcode sequence. Individually transduced cells were cultured competitively to assess their transforming potential or drug sensitivity. Genomic DNA was extracted from the cells using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany), followed by amplification by polymerase chain reaction (PCR) using specific primers (Supplementary Table S2), including the indices and adapter sequences for the Illumina platform (Illumina, San Diego, CA, USA). Sequencing libraries were prepared using the NEBNext Q5 Hot Start HiFi PCR Master Mix (New England Biolabs, Ipswich, MA, USA) according to the manufacturer’s instructions, and the resulting products were purified using AMPure Beads (Beckman Coulter, Brea, CA, USA). The quality of the libraries was assessed using a Qubit 2.0 Fluorometer (Thermo Fisher Scientific) and an Agilent 2200 TapeStation System (Agilent Technologies, Santa Clara, CA, USA). The products were sequenced on the Illumina MiSeq using Reagent Kit V2 (300 cycles) and 150-bp paired-end reads were generated (Supplementary Table S3). The barcode sequence 5′-CTAGACTGCCXXXXXXXXXXGGATCACTCT-3′ (where X denotes any nucleotide) was included in the sequencing results and the number of barcodes in each variant was quantified.
Drug sensitivity assay using the MANO method in vitro
3T3 cells or human cancer cells expressing each variant were mixed in equal amounts and treated with the indicated concentrations of osimertinib (1 nM to 1 µM), trametinib (100 pM to 10 nM), dabrafenib (100 pM to 500 nM), olaparib (10 nM to 1 µM), or 10% dimethyl sulfoxide (DMSO) as the vehicle control. The cells were treated for four days (osimertinib, trametinib, and dabrafenib) or 6 days (olaparib). Drugs used for the assays were purchased from Selleck Chemicals (Houston, TX, USA). The experiments were conducted in both biological and technical triplicate. The number of barcodes was calculated using MANO (Supplementary Table S4). The most resistant variant of each drug was used as a reference control for scaling the barcode count. The relative growth inhibition of each variant was calculated as the ratio of the average read number of triplicates to that of the control. Variants with fewer than 100 barcode sequence reads were excluded from the analysis.
PrestoBlue cell viability assay
Cells expressing each variant were seeded into 96-well plates with 100 µL of culture medium per well, and drugs were added at different concentrations (described in each figure). After treatment for a few days, 10 µL of PrestoBlue (Thermo Fisher Scientific) was added to the plates, and fluorescence was measured (excitation 530 nm, emission 590 nm) after 2 h of incubation at 0.1 s. The data were analyzed and presented as a sigmoid curve using GraphPad Prism software v9. The R package SynergyFinder and ZIP synergy model were used to calculate the drug synergy score [53]. The score for each concentration was calculated, and the average of the scores for all concentrations was calculated. Synergy scores greater than 10 suggest that the interaction between the two drugs is likely synergistic.
Quantitative real-time PCR
Total RNA was extracted from SJSA1 cells using the RNeasy kit (Qiagen) according to the manufacturer’s protocol. DNase I digestion was performed to remove genomic DNA. The amount of total RNA was measured using NanoDrop One (Thermo Fisher Scientific) and cDNA was prepared by reverse transcription using SuperScript IV VILO reverse transcriptase (Thermo Fisher Scientific). Real-time PCR was performed using Power SYBR Green PCR Master Mix (Thermo Fisher Scientific) on a QuantStudio 3 real-time PCR system (Thermo Fisher Scientific) under the following conditions: 50 °C for 2 min, 95 °C for 10 min, and 40 cycles of 95 °C for 15 s and 60 °C for 60 s. Primers specific for BRCA1, BRCA2, GLI1, FANCD2, and ACTB are listed in Supplementary Table S5. The relative expression of the target mRNA was normalized to that of ACTB.
Western blot analysis
DLD1 parent, DLD1-KO, and DLD1-KO with individual variants were treated. Cells were lysed in 1% NP-40 lysis buffer containing protease and phosphatase inhibitors for 15 min on ice. Cell lysates were subjected to 7.5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and immunoblotting using primary antibodies against γH2AX (1:1000, #9718) and GAPDH (1:1000, #2118) (Cell Signaling Technology, Danvers, MA, USA). The secondary antibody used was horseradish peroxidase-linked anti-rabbit IgG (1:10,000, NA934V; GE Healthcare, Chicago, IL, USA). Band intensity was measured using ImageJ software. Ratios were corrected based on GAPDH expression levels. For the validation western blot, protein separation and detection were performed using an automated capillary electrophoresis system (Simple Western system and Compass software; ProteinSimple). Antibodies against the γH2AX (1:50) and GAPDH (1:50) were used. Signals were detected with an HRP-conjugated secondary anti-rabbit antibody and visualized using ProteinSimple software.
Immunostaining
DLD1 parent, DLD1-KO, and DLD1-KO overexpressing SMO and GLI1 were cultured on coverslips in 24-well plates. At the indicated time points, cells were fixed with 4% paraformaldehyde for 10 min and washed with PBS. The fixed cells were permeabilized with 0.1% Triton X-100/PBS for 15 min on ice, incubated with a blocking buffer (2% goat serum in 0.3% Triton X-100/PBS) for 10 min, and washed with PBS. The slides were incubated with primary antibodies (mouse anti-γH2AX (CA#613402; BioLegend, San Diego, CA, USA) and rabbit anti-53BP1 (Merck, PC712)) in blocking buffer for 60 min. After washing with PBS, the cells were incubated with the corresponding secondary antibodies (goat anti-mouse IgG (Alexa Fluor 488) (ab150113, Abcam, Cambridge, UK) or goat anti-rabbit IgG (Alexa Fluor 594) (Abcam, ab150080)) in 0.3% Triton X-100/PBS for 60 min in the dark. The slides were then washed with PBS, and coverslips were mounted using ProLong Diamond Antifade Mountant with DAPI (Thermo Fisher Scientific). Immunofluorescence imaging was performed using a confocal laser microscope (Olympus FV10i; OLYMPUS CORPORATION, Tokyo, Japan) and Leica TCS SP8 (Leica, Wetzlar, Germany).
Nucleotide excision repair (NER) activity assay
Cells (2 × 105) were seeded onto coverslips in a 35 mm culture dish one day before irradiation. After washing with PBS, the cells were irradiated with UVC at 15 J/m2 and cultured in fresh medium for various times to allow repair of photolesions. Cells were fixed for 10 min at room temperature with 4% paraformaldehyde in PBS. Immunostaining with anti-cyclobutane pyrimidine dimer (CPD) antibody (TDM-2; Cosmo Bio Co., Ltd, Tokyo, Japan) was performed according to the manufacturer’s protocol. Briefly, the cells were permeabilized with PBS containing 0.5% TritonX-100 on ice for 5 min and incubated with 2 M HCl (to denature DNA) for 30 min at room temperature. After blocking with 20% FBS in PBS for 30 min at 37 °C, the cells were incubated with an anti-CPD antibody (1:1500 dilution) for 30 min at 37 °C, followed by incubation with Alexa Fluor 594-labeled anti-mouse IgG antibody (1:100 dilution; Abcam) for 30 min at 37 °C. Coverslips were mounted onto glass slides with drops of ProLong Diamond Antifade Mountant containing DAPI (Invitrogen). Fluorescent images were acquired using a confocal laser scanning microscope (Leica TCS SP8), and the fluorescence intensities for CPD and genomic DNA in individual cell nuclei were measured using the Fiji software. Alexa Fluor 594 fluorescence (CPD) was divided by DAPI fluorescence (DNA) to obtain the relative CPD levels for each nucleus.
Patient specimens
Formalin-fixed paraffin-embedded (FFPE) surgically resected tumors or biopsy specimens were obtained from three patients. This study was conducted in accordance with the Declaration of Helsinki and the protocol was approved by the ethics committee of National Cancer Center (No. 2015-202). All subjects were provided written informed consent, except for those who could not be reached because of loss of follow-up or death at registration. In these cases, the institutional review board granted permission for the existing tissue samples to be used for research purposes. No samples from the patients who had opted out of participation were used in this study.
RNA sequencing (RNA-seq) and expression analysis
Total RNA was extracted using the RNeasy kit (Qiagen) according to the manufacturer’s protocol. DNase I digestion was performed to remove genomic DNA. Next, 750 ng of total RNA was used for RNA-seq analysis. Sequencing libraries were constructed using the NEBNext Ultra Directional RNA Library Prep Kit for Illumina (#E7420), and quality was evaluated using the Qubit 2.0 Fluorometer and Agilent 2200 TapeStation System. Sequencing was conducted using NovaSeq 6000 (Illumina). RNA-seq data were aligned to the hg19 genome using TopHat v2.1.1 with default settings. Gene expression was quantified using Cufflinks v2.0.9, using the default settings. GSEA v4.2.2 software was used to calculate the normalized enrichment score and statistical significance against the Molecular Signatures Database (MSigDB) hallmark, C2, C5, and C6. Analyses were performed on normalized counts generated using DESeq2. The expression datasets are shown in Supplementary Tables S6–9. Settings of perform; number of permutations: 1000; dataset to gene symbols: collapse; permutation type: gene set; enrichment statistic: weighted; metric for ranking genes: Signal2Noise; gene list sorting mode: real; gene list ordering mode: descending; maximum size of gene sets: 500; and minimum size of gene sets: 15. Pathway analysis was performed using Ingenuity Pathway Analysis (IPA) (Qiagen).
Tumor xenograft assay
Animal studies were conducted according to protocols approved by the Animal Ethics Committee of the National Cancer Research Center (Tokyo, Japan). The DLD1 parental, DLD1-KO, and DLD1-KO cells expressing GLI1 wild-type (2.0 × 106) were injected into 6-week-old female nude mice. Olaparib was dissolved in 5% dimethyl sulfoxide (DMSO), 30% polyethylene glycol (PEG) 300, and 65% sterile ultrapure water. GANT61 was dissolved in 5% ethanol, 40% polyethylene glycol (PEG) 300, 5% Tween 80, and 50% sterile ultrapure water. Each group of six mice was administered an intraperitoneal injection of olaparib (50 mg/kg body weight), GANT61 (50 mg/kg body weight), the combination of olaparib (50 mg/kg body weight), GANT61 (50 mg/kg body weight), or vehicle every other day. Body weight and tumor volume were measured twice per week. Tumor volume was calculated using the following formula: 0.5 × (large diameter) × (small diameter)2. The mice were sacrificed on day 42 or when the tumor volume reached 2000 mm3.
Data availability
The data generated in this study are available within the article and its supplementary table files.
References
Kohsaka S, Nagano M, Ueno T, Suehara Y, Hayashi T, Shimada N. et al. A method of high-throughput functional evaluation of EGFR gene variants of unknown significance in cancer. Sci Transl Med. 2017;9:eaan6566.
Ikeuchi H, Hirose T, Ikegami M, Takamochi K, Suzuki K, Mano H, et al. Preclinical assessment of combination therapy of EGFR tyrosine kinase inhibitors in a highly heterogeneous tumor model. Oncogene. 2022;41:2470–9.
Hayes TK, Aquilanti E, Persky NS, Yang X, Kim EE, Brenan L, et al. Comprehensive mutational scanning of EGFR reveals TKI sensitivities of extracellular domain mutants. Nat Commun. 2024;15:2742.
Robichaux JP, Le X, Vijayan RSK, Hicks JK, Heeke S, Elamin YY, et al. Structure-based classification predicts drug response in EGFR-mutant NSCLC. Nature. 2021;597:732–7.
Robert C, Karaszewska B, Schachter J, Rutkowski P, Mackiewicz A, Stroiakovski D, et al. Improved overall survival in melanoma with combined dabrafenib and trametinib. N Engl J Med. 2015;372:30–9.
Planchard D, Besse B, Groen HJM, Souquet PJ, Quoix E, Baik CS, et al. Dabrafenib plus trametinib in patients with previously treated BRAF(V600E)-mutant metastatic non-small cell lung cancer: an open-label, multicentre phase 2 trial. Lancet Oncol. 2016;17:984–93.
Subbiah V, Kreitman RJ, Wainberg ZA, Gazzah A, Lassen U, Stein A, et al. Dabrafenib plus trametinib in BRAFV600E-mutated rare cancers: the phase 2 ROAR trial. Nat Med. 2023;29:1103–12.
Bryant HE, Schultz N, Thomas HD, Parker KM, Flower D, Lopez E, et al. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nature. 2005;434:913–7.
Farmer H, McCabe N, Lord CJ, Tutt AN, Johnson DA, Richardson TB, et al. Targeting the DNA repair defect in BRCA mutant cells as a therapeutic strategy. Nature. 2005;434:917–21.
Lord CJ, Ashworth A. PARP inhibitors: synthetic lethality in the clinic. Science. 2017;355:1152–8.
Petropoulos M, Karamichali A, Rossetti GG, Freudenmann A, Iacovino LG, Dionellis VS, et al. Transcription-replication conflicts underlie sensitivity to PARP inhibitors. Nature. 2024;628:433–41.
Li H, Liu ZY, Wu N, Chen YC, Cheng Q, Wang J. PARP inhibitor resistance: the underlying mechanisms and clinical implications. Mol Cancer. 2020;19:107.
Lee E, Matulonis UA. PARP inhibitor resistance mechanisms and implications for post-progression combination therapies. Cancers (Basel). 2020;12:2054
Pettitt SJ, Frankum JR, Punta M, Lise S, Alexander J, Chen Y, et al. Clinical BRCA1/2 reversion analysis identifies hotspot mutations and predicted neoantigens associated with therapy resistance. Cancer Discov. 2020;10:1475–88.
Lin KK, Harrell MI, Oza AM, Oaknin A, Ray-Coquard I, Tinker AV, et al. BRCA reversion mutations in circulating tumor DNA predict primary and acquired resistance to the PARP inhibitor rucaparib in high-grade ovarian carcinoma. Cancer Discov. 2019;9:210–9.
Liu Y, Burness ML, Martin-Trevino R, Guy J, Bai S, Harouaka R, et al. RAD51 mediates resistance of cancer stem cells to PARP inhibition in triple-negative breast cancer. Clin Cancer Res. 2017;23:514–22.
Xu G, Chapman JR, Brandsma I, Yuan J, Mistrik M, Bouwman P, et al. REV7 counteracts DNA double-strand break resection and affects PARP inhibition. Nature. 2015;521:541–4.
Noordermeer SM, Adam S, Setiaputra D, Barazas M, Pettitt SJ, Ling AK, et al. The shieldin complex mediates 53BP1-dependent DNA repair. Nature. 2018;560:117–21.
Gupta R, Somyajit K, Narita T, Maskey E, Stanlie A, Kremer M, et al. DNA repair network analysis reveals shieldin as a key regulator of NHEJ and PARP inhibitor sensitivity. Cell. 2018;173:972–988 e923.
Jaspers JE, Kersbergen A, Boon U, Sol W, van Deemter L, Zander SA, et al. Loss of 53BP1 causes PARP inhibitor resistance in Brca1-mutated mouse mammary tumors. Cancer Discov. 2013;3:68–81.
He YJ, Meghani K, Caron MC, Yang C, Ronato DA, Bian J, et al. DYNLL1 binds to MRE11 to limit DNA end resection in BRCA1-deficient cells. Nature. 2018;563:522–6.
Nagano M, Kohsaka S, Ueno T, Kojima S, Saka K, Iwase H, et al. High-throughput functional evaluation of variants of unknown significance in ERBB2. Clin Cancer Res. 2018;24:5112–22.
Kudo K, Gavin E, Das S, Amable L, Shevde LA, Reed E. Inhibition of Gli1 results in altered c-Jun activation, inhibition of cisplatin-induced upregulation of ERCC1, XPD and XRCC1, and inhibition of platinum-DNA adduct repair. Oncogene. 2012;31:4718–24.
Avery JT, Zhang R, Boohaker RJ. GLI1: a therapeutic target for cancer. Front Oncol. 2021;11:673154.
Infante P, Mori M, Alfonsi R, Ghirga F, Aiello F, Toscano S, et al. Gli1/DNA interaction is a druggable target for Hedgehog-dependent tumors. EMBO J. 2015;34:200–17.
Hirose T, Ikegami M, Endo M, Matsumoto Y, Nakashima Y, Mano H, et al. Extensive functional evaluation of exon 20 insertion mutations of EGFR. Lung Cancer. 2021;152:135–42.
Mizuno S, Ikegami M, Koyama T, Sunami K, Ogata D, Kage H, et al. High-throughput functional evaluation of MAP2K1 variants in cancer. Mol Cancer Ther. 2023;22:227–39.
Nakamura IT, Kohsaka S, Ikegami M, Ikeuchi H, Ueno T, Li K, et al. Comprehensive functional evaluation of variants of fibroblast growth factor receptor genes in cancer. NPJ Precis Oncol. 2021;5:66.
Ikegami M, Kohsaka S, Ueno T, Momozawa Y, Inoue S, Tamura K, et al. High-throughput functional evaluation of BRCA2 variants of unknown significance. Nat Commun. 2020;11:2573.
Xia R, Xu M, Yang J, Ma X. The role of Hedgehog and Notch signaling pathway in cancer. Mol Biomed. 2022;3:44.
Suchors C, Kim J. Canonical Hedgehog pathway and noncanonical GLI transcription factor activation in cancer. Cells. 2022;11.
Mani C, Tripathi K, Omy TR, Reedy M, Manne U, Palle K. GLI1-targeting drugs induce replication stress and homologous recombination deficiency and synergize with PARP-targeted therapies in triple negative breast cancer cells. Biochim Biophys Acta Mol Basis Dis. 2022;1868:166300.
Mani C, Tripathi K, Chaudhary S, Somasagara RR, Rocconi RP, Crasto C, et al. Hedgehog/GLI1 transcriptionally regulates FANCD2 in ovarian tumor cells: its inhibition induces HR-deficiency and synergistic lethality with PARP inhibition. Neoplasia. 2021;23:1002–15.
Palle K, Mani C, Tripathi K, Athar M. Aberrant GLI1 activation in DNA damage response, carcinogenesis and chemoresistance. Cancers. 2015;7:2330–51.
Yoshioka KI, Kusumoto-Matsuo R, Matsuno Y, Ishiai M. Genomic instability and cancer risk associated with erroneous DNA Repair. Int J Mol Sci. 2021;22:12254.
Qin S, Kitty I, Hao Y, Zhao F, Kim W. Maintaining genome integrity: protein kinases and phosphatases orchestrate the balancing act of DNA double-strand breaks repair in cancer. Int J Mol Sci. 2023;24:10212.
Brambati A, Sacco O, Porcella S, Heyza J, Kareh M, Schmidt JC, et al. RHINO directs MMEJ to repair DNA breaks in mitosis. Science. 2023;381:653–60.
Fleury H, MacEachern MK, Stiefel CM, Anand R, Sempeck C, Nebenfuehr B, et al. The APE2 nuclease is essential for DNA double-strand break repair by microhomology-mediated end joining. Mol Cell. 2023;83:1429–1445 e1428.
Chandramouly G, Jamsen J, Borisonnik N, Tyagi M, Calbert ML, Tredinnick T, et al. Pollambda promotes microhomology-mediated end-joining. Nat Struct Mol Biol. 2023;30:107–14.
Apelt K, Lans H, Scharer OD, Luijsterburg MS. Nucleotide excision repair leaves a mark on chromatin: DNA damage detection in nucleosomes. Cell Mol Life Sci. 2021;78:7925–42.
Renaudin X, Rosselli F. The FANC/BRCA pathway releases replication blockades by eliminating DNA interstrand cross-links. Genes (Basel). 2020;11:585
Nayak S, Calvo JA, Cantor SB. Targeting translesion synthesis (TLS) to expose replication gaps, a unique cancer vulnerability. Expert Opin Ther Targets. 2021;25:27–36.
Peake JD, Noguchi E. Fanconi anemia: current insights regarding epidemiology, cancer, and DNA repair. Hum Genet. 2022;141:1811–36.
Nieddu V, Melocchi V, Battistini C, Franciosa G, Lupia M, Stellato C, et al. Matrix Gla Protein drives stemness and tumor initiation in ovarian cancer. Cell Death Dis. 2023;14:220.
Karin-Kujundzic V, Covarrubias-Pinto A, Skrtic A, Vranic S, Serman L. New insight into the role of PTCH1 protein in serous ovarian carcinomas. Int J Oncol. 2022;61:145.
Kuehn J, Espinoza-Sanchez NA, Teixeira F, Pavao MSG, Kiesel L, Gyorffy B, et al. Prognostic significance of hedgehog signaling network-related gene expression in breast cancer patients. J Cell Biochem. 2021;122:577–97.
Guo P, Chen Q, Peng K, Xie J, Liu J, Ren W, et al. Nuclear receptor coactivator SRC-1 promotes colorectal cancer progression through enhancing GLI2-mediated Hedgehog signaling. Oncogene. 2022;41:2846–59.
Kotulak-Chrzaszcz A, Kmiec Z, Wierzbicki PM. Sonic Hedgehog signaling pathway in gynecological and genitourinary cancer (Review). Int J Mol Med. 2021;47.
Lama-Sherpa TD, Lin VTG, Metge BJ, Weeks SE, Chen D, Samant RS, et al. Hedgehog signaling enables repair of ribosomal DNA double-strand breaks. Nucleic Acids Res. 2020;48:10342–52.
Akagi T, Sasai K, Hanafusa H. Refractory nature of normal human diploid fibroblasts with respect to oncogene-mediated transformation. Proc Natl Acad Sci USA. 2003;100:13567–72.
Hirose T, Ikegami M, Kojima S, Yoshida A, Endo M, Shimada E, et al. Extensive analysis of 59 sarcoma-related fusion genes identified pazopanib as a potential inhibitor to COL1A1-PDGFB fusion gene. Cancer Sci. 2023;114:4089–100.
Koga T, Suda K, Mitsudomi T. Utility of the Ba/F3 cell system for exploring on-target mechanisms of resistance to targeted therapies for lung cancer. Cancer Sci. 2022;113:815–27.
Zheng S, Wang W, Aldahdooh J, Malyutina A, Shadbahr T, Tanoli Z, et al. SynergyFinder plus: toward better interpretation and annotation of drug combination screening datasets. Genomics Proteom Bioinform. 2022;20:587–96.
Acknowledgements
The authors thank the members of our division, particularly A. Maruyama-Shiino, Y. Ono, and M. Takeyama for technical assistance.
Funding
This study was supported by the grants of Practical Research for Innovative Cancer Control (grant no. JP22ck0106536), Program for Promoting Platform of Genomics based Drug Discovery (grant no. JP23kk0305018), and Moonshot Research and Development Program (grant no. JP22zf0127009) from Japan Agency for Medical Research and Development (AMED), and also supported by the JSPS Grants-in-Aid for Scientific Research (B) (grant no. JP21H02795).
Author information
Authors and Affiliations
Contributions
SK conceived of the project and designed the study. SK and HM developed the study methodology. HI, YM., and RKM performed experiments. HO, YK, KY, and YY provided clinical samples. HI, TU, MI, and SK analyzed and interpreted the data. KY, KT, KS, and HM provided the administrative and technical support. HI, YM, RKM, HM, and SK wrote and edited the manuscript with feedback from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ikeuchi, H., Matsuno, Y., Kusumoto-Matsuo, R. et al. GLI1 confers resistance to PARP inhibitors by activating the DNA damage repair pathway. Oncogene (2024). https://doi.org/10.1038/s41388-024-03105-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41388-024-03105-1
- Springer Nature Limited