

Abstract
Functional centromeres are essential for proper cell division. Centromeres are established largely by epigenetic processes resulting in incorporation of the histone H3 variant CENP-A. Here, we demonstrate the direct involvement of H2B monoubiquitination, mediated by RNF20 in humans or Brl1 in Schizosaccharomyces pombe, in centromeric chromatin maintenance. Monoubiquinated H2B (H2Bub1) is needed for this maintenance, promoting noncoding transcription, centromere integrity and accurate chromosomal segregation. A transient pulse of centromeric H2Bub1 leads to RNA polymerase II–mediated transcription of the centromere's central domain, coupled to decreased H3 stability. H2Bub1-deficient cells have centromere cores that, despite their intact centromeric heterochromatin barriers, exhibit characteristics of heterochromatin, such as silencing histone modifications, reduced nucleosome turnover and reduced levels of transcription. In the H2Bub1-deficient cells, centromere functionality is hampered, thus resulting in unequal chromosome segregation. Therefore, centromeric H2Bub1 is essential for maintaining active centromeric chromatin.








Similar content being viewed by others


References
Sansó, M. et al. A positive feedback loop links opposing functions of P-TEFb/Cdk9 and Histone H2B ubiquitylation to regulate transcript elongation in fission yeast. PLoS Genet. 8, e1002822 (2012).
Sun, Z.W. & Allis, C.D. Ubiquitination of histone H2B regulates H3 methylation and gene silencing in yeast. Nature 418, 104–108 (2002).
Trujillo, K.M. & Osley, M.A. A role for H2B ubiquitylation in DNA replication. Mol. Cell 48, 734–746 (2012).
Chen, S., Li, J., Wang, D.L. & Sun, F.L. Histone H2B lysine 120 monoubiquitination is required for embryonic stem cell differentiation. Cell Res. 22, 1402–1405 (2012).
Karpiuk, O. et al. The histone H2B monoubiquitination regulatory pathway is required for differentiation of multipotent stem cells. Mol. Cell 46, 705–713 (2012).
Fuchs, G. et al. RNF20 and USP44 regulate stem cell differentiation by modulating H2B monoubiquitylation. Mol. Cell 46, 662–673 (2012).
Chernikova, S.B. et al. Deficiency in mammalian histone H2B ubiquitin ligase Bre1 (Rnf20/Rnf40) leads to replication stress and chromosomal instability. Cancer Res. 72, 2111–2119 (2012).
Moyal, L. et al. Requirement of ATM-dependent monoubiquitylation of histone H2B for timely repair of DNA double-strand breaks. Mol. Cell 41, 529–542 (2011).
Nakamura, K. et al. Regulation of homologous recombination by RNF20-dependent H2B ubiquitination. Mol. Cell 41, 515–528 (2011).
Svensson, J.P., Fry, R.C., Wang, E.M., Somoza, L.A. & Samson, L.D. Identification of novel human damage response proteins targeted through yeast orthology. PLoS ONE 7, e37368 (2012).
Ma, M.K.W., Heath, C., Hair, A. & West, A.G. Histone crosstalk directed by H2B ubiquitination is required for chromatin boundary integrity. PLoS Genet. 7, e1002175 (2011).
Zofall, M. & Grewal, S.I.S. HULC, a histone H2B ubiquitinating complex, modulates heterochromatin independent of histone methylation in fission yeast. J. Biol. Chem. 282, 14065–14072 (2007).
Norris, D., Dunn, B. & Osley, M.A. The effect of histone gene deletions on chromatin structure in Saccharomyces cerevisiae. Science 242, 759–761 (1988).
Maruyama, T., Nakamura, T., Hayashi, T. & Yanagida, M. Histone H2B mutations in inner region affect ubiquitination, centromere function, silencing and chromosome segregation. EMBO J. 25, 2420–2431 (2006).
Latham, J.A., Chosed, R.J., Wang, S.Z. & Dent, S.Y.R. Chromatin signaling to kinetochores: transregulation of Dam1 methylation by histone H2B ubiquitination. Cell 146, 709–719 (2011).
Black, B.E. & Bassett, E.A. The histone variant CENP-A and centromere specification. Curr. Opin. Cell Biol. 20, 91–100 (2008).
Mellone, B.G. & Allshire, R.C. Stretching it: putting the CEN(P-A) in centromere. Curr. Opin. Genet. Dev. 13, 191–198 (2003).
Fachinetti, D. et al. A two-step mechanism for epigenetic specification of centromere identity and function. Nat. Cell Biol. 15, 1056–1066 (2013).
Chan, F.L. et al. Active transcription and essential role of RNA polymerase II at the centromere during mitosis. Proc. Natl. Acad. Sci. USA 109, 1979–1984 (2012).
Choi, E.S. et al. Identification of noncoding transcripts from within CENP-A chromatin at fission yeast centromeres. J. Biol. Chem. 286, 23600–23607 (2011).
Ohkuni, K. & Kitagawa, K. Endogenous transcription at the centromere facilitates centromere activity in budding yeast. Curr. Biol. 21, 1695–1703 (2011).
Gent, J.I. & Dawe, R.K. RNA as a structural and regulatory component of the centromere. Annu. Rev. Genet. 46, 443–453 (2012).
Ferri, F., Bouzinba-Segard, H., Velasco, G., Hube, F. & Francastel, C. Non-coding murine centromeric transcripts associate with and potentiate Aurora B kinase. Nucleic Acids Res. 37, 5071–5080 (2009).
Topp, C.N., Zhong, C.X. & Dawe, R.K. Centromere-encoded RNAs are integral components of the maize kinetochore. Proc. Natl. Acad. Sci. USA 101, 15986–15991 (2004).
Scott, K.C. Transcription and ncRNAs: at the cent(rome)re of kinetochore assembly and maintenance. Chromosome Res. 21, 643–651 (2013).
Ekwall, K. Epigenetic control of centromere behavior. Annu. Rev. Genet. 41, 63–81 (2007).
Ribeiro, S.A. et al. A super-resolution map of the vertebrate kinetochore. Proc. Natl. Acad. Sci. USA 107, 10484–10489 (2010).
Sullivan, B.A. & Karpen, G.H. Centromeric chromatin exhibits a histone modification pattern that is distinct from both euchromatin and heterochromatin. Nat. Struct. Mol. Biol. 11, 1076–1083 (2004).
Volpe, T.A. et al. Regulation of heterochromatic silencing and histone H3 lysine-9 methylation by RNAi. Science 297, 1833–1837 (2002).
Shema, E. et al. The histone H2B-specific ubiquitin ligase RNF20/hBRE1 acts as a putative tumor suppressor through selective regulation of gene expression. Genes Dev. 22, 2664–2676 (2008).
Wang, E. et al. Histone H2B ubiquitin ligase RNF20 is required for MLL-rearranged leukemia. Proc. Natl. Acad. Sci. USA 110, 3901–3906 (2013).
Shema, E., Kim, J., Roeder, R.G. & Oren, M. RNF20 inhibits TFIIS-facilitated transcriptional elongation to suppress pro-oncogenic gene expression. Mol. Cell 42, 477–488 (2011).
Eymery, A. et al. A transcriptomic analysis of human centromeric and pericentric sequences in normal and tumor cells. Nucleic Acids Res. 37, 6340–6354 (2009).
Ting, D.T. et al. Aberrant overexpression of satellite repeats in pancreatic and other epithelial cancers. Science 331, 593–596 (2011).
Sadeghi, L., Bonilla, C., Stralfors, A., Ekwall, K. & Svensson, J.P. Podbat: a novel genomic tool reveals Swr1-Independent H2A.Z incorporation at gene coding sequences through epigenetic meta-analysis. PLOS Comput. Biol. 7, e1002163 (2011).
Scott, K.C., Merrett, S.L. & Willard, H.F. A heterochromatin barrier partitions the fission yeast centromere into discrete chromatin domains. Curr. Biol. 16, 119–129 (2006).
Noma, K., Cam, H.P., Maraia, R.J. & Grewal, S.I.S. A role for TFIIIC transcription factor complex in genome organization. Cell 125, 859–872 (2006).
Pidoux, A.L. et al. Fission yeast Scm3: a CENP-A receptor required for integrity of subkinetochore chromatin. Mol. Cell 33, 299–311 (2009).
Williams, J.S., Hayashi, T., Yanagida, M. & Russell, P. Fission yeast Scm3 mediates stable assembly of Cnp1/CENP-A into centromeric chromatin. Mol. Cell 33, 287–298 (2009).
Tanny, J.C., Erdjument-Bromage, H., Tempst, P. & Allis, C.D. Ubiquitylation of histone H2B controls RNA polymerase II transcription elongation independently of histone H3 methylation. Genes Dev. 21, 835–847 (2007).
Chandrasekharan, M.B., Huang, F. & Sun, Z.W. Ubiquitination of histone H2B regulates chromatin dynamics by enhancing nucleosome stability. Proc. Natl. Acad. Sci. USA 106, 16686–16691 (2009).
Fleming, A.B., Kao, C.F., Hillyer, C., Pikaart, M. & Osley, M.A. H2B ubiquitylation plays a role in nucleosome dynamics during transcription elongation. Mol. Cell 31, 57–66 (2008).
Wang, S.W., Asakawa, K., Win, T.Z., Toda, T. & Norbury, C.J. Inactivation of the pre-mRNA cleavage and polyadenylation factor Pfs2 in fission yeast causes lethal cell cycle defects. Mol. Cell. Biol. 25, 2288–2296 (2005).
Choi, E.S., Shin, J.A., Kim, H.S. & Jang, Y.K. Dynamic regulation of replication independent deposition of histone H3 in fission yeast. Nucleic Acids Res. 33, 7102–7110 (2005).
Aygün, O., Mehta, S. & Grewal, S.I. HDAC-mediated suppression of histone turnover promotes epigenetic stability of heterochromatin. Nat. Struct. Mol. Biol. 20, 547–554 (2013).
Emre, N.C.T. et al. Maintenance of low histone ubiquitylation by Ubp10 correlates with telomere-proximal Sir2 association and gene silencing. Mol. Cell 17, 585–594 (2005).
Henry, K.W. et al. Transcriptional activation via sequential histone H2B ubiquitylation and deubiquitylation, mediated by SAGA-associated Ubp8. Genes Dev. 17, 2648–2663 (2003).
Jung, I. et al. H2B monoubiquitylation is a 5′-enriched active transcription mark and correlates with exon-intron structure in human cells. Genome Res. 22, 1026–1035 (2012).
Fierz, B. et al. Histone H2B ubiquitylation disrupts local and higher-order chromatin compaction. Nat. Chem. Biol. 7, 113–119 (2011).
Shema-Yaacoby, E. et al. Systematic identification of proteins binding to chromatin-embedded ubiquitinylated H2B reveals recruitment of SWI/SNF to regulate transcription. Cell Rep. 4, 601–608 (2013).
Pavri, R. et al. Histone H2B monoubiquitination functions cooperatively with FACT to regulate elongation by RNA polymerase II. Cell 125, 703–717 (2006).
Lejeune, E. et al. The chromatin-remodeling factor FACT contributes to centromeric heterochromatin independently of RNAi. Curr. Biol. 17, 1219–1224 (2007).
Choi, E.S. et al. Factors that promote H3 chromatin integrity during transcription prevent promiscuous deposition of CENP-A(Cnp1) in fission yeast. PLoS Genet. 8, e1002985 (2012).
Du, Y., Topp, C.N. & Dawe, R.K. DNA binding of centromere protein C (CENPC) is stabilized by single-stranded RNA. PLoS Genet. 6, e1000835 (2010).
Wong, L.H. et al. Centromere RNA is a key component for the assembly of nucleoproteins at the nucleolus and centromere. Genome Res. 17, 1146–1160 (2007).
Bergmann, J.H. et al. Epigenetic engineering shows H3K4me2 is required for HJURP targeting and CENP-A assembly on a synthetic human kinetochore. EMBO J. 30, 328–340 (2011).
Keller, C., Kulasegaran-Shylini, R., Shimada, Y., Hotz, H.R. & Bühler, M. Noncoding RNAs prevent spreading of a repressive histone mark. Nat. Struct. Mol. Biol. 20, 994–1000 (2013).
Fisher, A.G. & Merkenschlager, M. Gene silencing, cell fate and nuclear organisation. Curr. Opin. Genet. Dev. 12, 193–197 (2002).
Sato, H., Masuda, F., Takayama, Y., Takahashi, K. & Saitoh, S. Epigenetic inactivation and subsequent heterochromatinization of a centromere stabilize dicentric chromosomes. Curr. Biol. 22, 658–667 (2012).
Durand-Dubief, M., Persson, J., Norman, U., Hartsuiker, E. & Ekwall, K. Topoisomerase I regulates open chromatin and controls gene expression in vivo. EMBO J. 29, 2126–2134 (2010).
Jurka, J. Repbase update: a database and an electronic journal of repetitive elements. Trends Genet. 16, 418–420 (2000).
Lantermann, A.B. et al. Schizosaccharomyces pombe genome-wide nucleosome mapping reveals positioning mechanisms distinct from those of Saccharomyces cerevisiae. Nat. Struct. Mol. Biol. 17, 251–257 (2010).
Haider, S. et al. The landscape of DNA repeat elements in human heart failure. Genome Biol. 13, R90 (2012).
Acknowledgements
Reagents were kindly provided by S. Grewal (US National Cancer Institute), K.C. Scott (Duke University), C. Norbury (University of Oxford), K. Gull (University of Oxford) and R.C. Allshire (University of Edinburgh). This study was supported by the Swedish Research Council (VR-M2579 and VR-NT4448) (K.E.) and the Swedish Cancer Society (CAN2012/238) (K.E.). The authors would like to thank S. Le Guyader at the Live Cell Imaging unit, Department of Biosciences and Nutrition, Karolinska Institutet, which is supported by grants from the Knut and Alice Wallenberg Foundation (KAW2006.0265) and the Swedish Research Council (822-2007-902 and K2008-661-21010-01-1). We are also grateful to P. Prasat for technical assistance.
Author information
Authors and Affiliations
Contributions
L. Sadeghi performed all experiments in S. pombe, and L. Siggens performed all experiments in human cells. L. Sadeghi, L. Siggens, J.P.S. and K.E. designed the study, analyzed the data and wrote the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
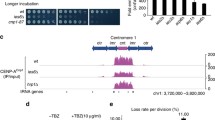
Supplementary Figure 1 Genome-browser views.
Genome browser view of (a) centromere 1 and (b) centromere 3 using anti-H3K9me2, anti-H3, and anti-CENP-ACnp1 relative to input; WT (top), htb1-K119R mutant (middle), and htb1-K119R/WT ratio (bottom). (c) ChIP levels of H3K9me2, H3, and CENP-ACnp1 at two centromeric loci (central core domain, cnt1; innermost repeat, imr2) in WT and htb1-K119R determined ChIP-qPCR relative to empty beads. Error bars indicate s.e.m. (n=3 independent experiments). (d) ChIP-qPCR of H3 and H3K9me2 at control locus LTR(n=3 independent experiments).
Supplementary Figure 2 Global effects of the htb1-K119R mutation on transcription and chromatin compaction.
H2Bub1-levels correlate with transcription (a) Scatterplot of H2Bub1 and RNA Pol II versus RNA Pol II and RNA expression. heterochromatin mark H3K9me2 in WT and H2Bub1-deficient cells (htb1-K119R). Pearson's correlation coefficient is indicated at the bottom right of each panel. (b) Transcripts have been divided into 4 categories depending on their expression level (high: 12–14, medium: 10–12, low: 8–10, background: 6–8). Transcripts were aligned after their transcription start site (TSS) and the levels of H2Bub1 and H3K4me2 (from ref. 35) were plotted as a function of position relative to TSS. (c) Scatterplot of RNA levels in WT and htb1-K119R. Grey line: unchanged between the samples, red dashed line: up-regulated in mutant >2-fold, green dashed line: down-regulated >2-fold. Genes encoding kinetochore proteins are marked (x) (d) MNase digestion of chromatin from WT and htb1-K119R cells. * marks oligonucleosomes present in WT but absent in htb1-K119R, arrow points at regions of undigested product in the htb1-K119Rextracts.
Supplementary Figure 3 No htb1-K119R effect on centromere transcript stability or nucleosome stability.
(a) Transcripts from the centromere core are rapidly degraded. Three primers pairs (1–3) were used in qRT-PCR to quantify the transcription of the pericentric (1) and central domain (2–3) of the centromere in WT and htb1-K119R cells. Error bars indicate s.e.m. (n=3 independent experiments) (b) (left) Nucleosomal turnover in WT and htb1-K119R cells as measured by ChIP-qPCR using antibody to hemagglutinin (HA) 2h and 0h after induction of H3-HA expression at two genes, sua1 and cdm1. Bars indicate range (n=2 independent experiments) (right) Expression levels of sua1 and cdm1 as determined by qRT-PCR. Error bars indicate s.e.m.(n=3 independent experiments).
Supplementary Figure 5 Colocalization of proteins with centromere-specific histone CENP-A in human cells.
Representative images of HeLa cells and in situ proximity ligation assay using one antibody against CENP-A and a second antibody against H2Bub1, RNF20 or RNA Pol II (n=2 independent experiments). Proximity between the two antibodies (<40 nm) result in a signal (red). Nuclei are counterstained with DAPI (blue).
Supplementary Figure 6 RNA-seq in human NCCIT cells afterRNF20siRNA knockdown (GSE25882).
(a) Repeat transcript levels (α-satellite: ALR, ALR1; Pol II-transcribed L1 repetitive sequences; and Pol III-transcribed ALU repeats). Reads from high-throughput sequencing were aligned to repetitive sequences from Repbase. (b) Genome wide correlation of mRNA expression in si-RNF20 cells compared to control. (c) Transcript expression correlation of centromeric genes only, identified as having one or more centromere or kinetochore associated functions in AMIGO (http://amigo.geneontology.org/cgi-bin/amigo/go.cgi).
Supplementary Figure 7 Full-length gel images and immunoblots.
(a) Agarose gel electrophoresis of RT-PCR products corresponding to Fig. 3f. (b) Immunoblot analysis of RNF20 expression in HEK293T cells stably expressing non-targeting and RNF20 targeting shRNA corresponding to Fig. 6a. H2B was used as a loading control. (c) Analysis of H2Bub1 in HEK293T cells expressing RNF20 targeting shRNA as in (b).
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–7 and Supplementary Tables 1–4. (PDF 3448 kb)
Rights and permissions
About this article
Cite this article
Sadeghi, L., Siggens, L., Svensson, J. et al. Centromeric histone H2B monoubiquitination promotes noncoding transcription and chromatin integrity. Nat Struct Mol Biol 21, 236–243 (2014). https://doi.org/10.1038/nsmb.2776
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nsmb.2776
- Springer Nature America, Inc.
This article is cited by
-
The Mis6 inner kinetochore subcomplex maintains CENP-A nucleosomes against centromeric non-coding transcription during mitosis
Communications Biology (2022)
-
The H2B ubiquitin-protein ligase RNF40 is required for somatic cell reprogramming
Cell Death & Disease (2020)
-
Intricate regulation on epigenetic stability of the subtelomeric heterochromatin and the centromeric chromatin in fission yeast
Current Genetics (2019)
-
Histone H3K9 and H4 Acetylations and Transcription Facilitate the Initial CENP-AHCP−3 Deposition and De Novo Centromere Establishment in Caenorhabditis elegans Artificial Chromosomes
Epigenetics & Chromatin (2018)
-
Identification of a histone family gene signature for predicting the prognosis of cervical cancer patients
Scientific Reports (2017)