

Abstract
Cellular mechanisms that mediate steatohepatitis, an increasingly prevalent condition in the Western world for which no therapies are available1, are poorly understood. Despite the fact that its synthetic agonists induce fatty liver, the liver X receptor (LXR) transcription factor remains a target of interest because of its anti-atherogenic, cholesterol removal, and anti-inflammatory activities. Here we show that tetratricopeptide repeat domain protein 39B (Ttc39b, C9orf52) (T39), a high-density lipoprotein gene discovered in human genome-wide association studies2, promotes the ubiquitination and degradation of LXR. Chow-fed mice lacking T39 (T39−/−) display increased high-density lipoprotein cholesterol levels associated with increased enterocyte ATP-binding cassette transporter A1 (Abca1) expression and increased LXR protein without change in LXR messenger RNA. When challenged with a high fat/high cholesterol/bile salt diet, T39−/− mice or mice with hepatocyte-specific T39 deficiency show increased hepatic LXR protein and target gene expression, and unexpectedly protection from steatohepatitis and death. Mice fed a Western-type diet and lacking low-density lipoprotein receptor (Ldlr−/−T39−/−) show decreased fatty liver, increased high-density lipoprotein, decreased low-density lipoprotein, and reduced atherosclerosis. In addition to increasing hepatic Abcg5/8 expression and limiting dietary cholesterol absorption, T39 deficiency inhibits hepatic sterol regulatory element-binding protein 1 (SREBP-1, ADD1) processing. This is explained by an increase in microsomal phospholipids containing polyunsaturated fatty acids, linked to an LXRα-dependent increase in expression of enzymes mediating phosphatidylcholine biosynthesis and incorporation of polyunsaturated fatty acids into phospholipids. The preservation of endogenous LXR protein activates a beneficial profile of gene expression that promotes cholesterol removal and inhibits lipogenesis. T39 inhibition could be an effective strategy for reducing both steatohepatitis and atherosclerosis.




Similar content being viewed by others

References
Musso, G., Gambino, R., Cassader, M. & Pagano, G. A meta-analysis of randomized trials for the treatment of nonalcoholic fatty liver disease. Hepatology 52, 79–104 (2010)
Teslovich, T. M. et al. Biological, clinical and population relevance of 95 loci for blood lipids. Nature 466, 707–713 (2010)
Willer, C. J. et al. Discovery and refinement of loci associated with lipid levels. Nature Genet. 45, 1274–1283 (2013)
McNeish, J. et al. High density lipoprotein deficiency and foam cell accumulation in mice with targeted disruption of ATP-binding cassette transporter-1. Proc. Natl Acad. Sci. USA 97, 4245–4250 (2000)
Brunham, L. R. et al. Intestinal ABCA1 directly contributes to HDL biogenesis in vivo. J. Clin. Invest. 116, 1052–1062 (2006)
Wu, A. L. & Windmueller, H. G. Relative contributions by liver and intestine to individual plasma apolipoproteins in the rat. J. Biol. Chem. 254, 7316–7322 (1979)
Costet, P., Luo, Y., Wang, N. & Tall, A. R. Sterol-dependent transactivation of the ABC1 promoter by the liver X receptor/retinoid X receptor. J. Biol. Chem. 275, 28240–28245 (2000)
Zelcer, N., Hong, C., Boyadjian, R. & Tontonoz, P. LXR regulates cholesterol uptake through Idol-dependent ubiquitination of the LDL receptor. Science 325, 100–104 (2009)
Matsuzawa, N. et al. Lipid-induced oxidative stress causes steatohepatitis in mice fed an atherogenic diet. Hepatology 46, 1392–1403 (2007)
Rong, X. et al. LXRs regulate ER stress and inflammation through dynamic modulation of membrane phospholipid composition. Cell Metab. 18, 685–697 (2013)
Bensinger, S. J. et al. LXR signaling couples sterol metabolism to proliferation in the acquired immune response. Cell 134, 97–111 (2008)
Lo Sasso, G. et al. Intestinal specific LXR activation stimulates reverse cholesterol transport and protects from atherosclerosis. Cell Metab. 12, 187–193 (2010)
Zhang, Y. et al. Liver LXRα expression is crucial for whole body cholesterol homeostasis and reverse cholesterol transport in mice. J. Clin. Invest. 122, 1688–1699 (2012)
Yu, L. et al. Overexpression of ABCG5 and ABCG8 promotes biliary cholesterol secretion and reduces fractional absorption of dietary cholesterol. J. Clin. Invest. 110, 671–680 (2002)
Repa, J. J. et al. Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRα and LXRβ. Genes Dev. 14, 2819–2830 (2000)
Hegarty, B. D. et al. Distinct roles of insulin and liver X receptor in the induction and cleavage of sterol regulatory element-binding protein-1c. Proc. Natl Acad. Sci. USA 102, 791–796 (2005)
Huang, Y. et al. A feed-forward loop amplifies nutritional regulation of PNPLA3. Proc. Natl Acad. Sci. USA 107, 7892–7897 (2010)
Romeo, S. et al. Genetic variation in PNPLA3 confers susceptibility to nonalcoholic fatty liver disease. Nature Genet. 40, 1461–1465 (2008)
Vatner, D. F. et al. Insulin-independent regulation of hepatic triglyceride synthesis by fatty acids. Proc. Natl Acad. Sci. USA 112, 1143–1148 (2015)
Walker, A. K. et al. A conserved SREBP-1/phosphatidylcholine feedback circuit regulates lipogenesis in metazoans. Cell 147, 840–852 (2011)
Hishikawa, D. et al. Discovery of a lysophospholipid acyltransferase family essential for membrane asymmetry and diversity. Proc. Natl Acad. Sci. USA 105, 2830–2835 (2008)
Rong, X. et al. Lpcat3-dependent production of arachidonoyl phospholipids is a key determinant of triglyceride secretion. eLife 4, http://dx.doi.org/10.7554/eLife.06557 (2015)
Hannah, V. C., Ou, J., Luong, A., Goldstein, J. L. & Brown, M. S. Unsaturated fatty acids down-regulate srebp isoforms 1a and 1c by two mechanisms in HEK-293 cells. J. Biol. Chem. 276, 4365–4372 (2001)
Kammoun, H. L. et al. GRP78 expression inhibits insulin and ER stress-induced SREBP-1c activation and reduces hepatic steatosis in mice. J. Clin. Invest. 119, 1201–1215 (2009)
Boergesen, M. et al. Genome-wide profiling of liver X receptor, retinoid X receptor, and peroxisome proliferator-activated receptor α in mouse liver reveals extensive sharing of binding sites. Mol. Cell. Biol. 32, 852–867 (2012)
Lehrke, M. et al. Diet-dependent cardiovascular lipid metabolism controlled by hepatic LXRα. Cell Metab. 1, 297–308 (2005)
Perreault, N. & Beaulieu, J. F. Primary cultures of fully differentiated and pure human intestinal epithelial cells. Exp. Cell Res. 245, 34–42 (1998)
Anwar, K., Iqbal, J. & Hussain, M. M. Mechanisms involved in vitamin E transport by primary enterocytes and in vivo absorption. J. Lipid Res. 48, 2028–2038 (2007)
Iqbal, J., Anwar, K. & Hussain, M. M. Multiple, independently regulated pathways of cholesterol transport across the intestinal epithelial cells. J. Biol. Chem. 278, 31610–31620 (2003)
Kleiner, D. E. et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 41, 1313–1321 (2005)
Tomoyori, H. et al. Phytosterol oxidation products are absorbed in the intestinal lymphatics in rats but do not accelerate atherosclerosis in apolipoprotein E-deficient mice. J. Nutr. 134, 1690–1696 (2004)
Bligh, E. G. & Dyer, W. J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917 (1959)
Dzeletovic, S., Breuer, O., Lund, E. & Diczfalusy, U. Determination of cholesterol oxidation products in human plasma by isotope dilution-mass spectrometry. Anal. Biochem. 225, 73–80 (1995)
Spady, D. K. & Dietschy, J. M. Sterol synthesis in vivo in 18 tissues of the squirrel monkey, guinea pig, rabbit, hamster, and rat. J. Lipid Res. 24, 303–315 (1983)
Chan, R. B. et al. Comparative lipidomic analysis of mouse and human brain with Alzheimer disease. J. Biol. Chem. 287, 2678–2688 (2012)
Stary, H. C. et al. A definition of advanced types of atherosclerotic lesions and a histological classification of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Circulation 92, 1355–1374 (1995)
Gijbels, M. J. et al. Progression and regression of atherosclerosis in APOE3-Leiden transgenic mice: an immunohistochemical study. Atherosclerosis 143, 15–25 (1999)
Collins, J. L. et al. Identification of a nonsteroidal liver X receptor agonist through parallel array synthesis of tertiary amines. J. Med. Chem. 45, 1963–1966 (2002)
Acknowledgements
We express our gratitude to F. Matsuura for support, A. Morishita for advice on liver histology, M. Sakurai and T. Yamashita for advice on immunoprecipitation experiments, W. R. Lagor for advice on the reverse cholesterol transport study, M. Ishibashi for advice on animal administration, J. W. Medley for consultation on the coupling reaction, and N. Wang for project discussions. D. J. Gorman and J. So provided technical support, and O. Xu provided technical services for the lipidomics analysis. This work was supported by grants from the Manpei Suzuki Diabetes Foundation (to M.K.), VIDI grant 91715350 from the Netherlands Organization of Sciences (to M.W.), Rosalind Franklin Fellowship from the University Medical Center Groningen (to M.W.), JSPS KAKENHI Grant 15K160203 (to I.I.), and the Fondation Leducq (to A.R.T.). This work was supported by grants from the National Institutes of Health (T32 training program HL007343, M.M.M.; HL087123 and HL119830, to A.R.T.; HL101864 and HL111398, to D.J.R.; DK46900, to M.M.H.).
Author information
Authors and Affiliations
Contributions
J.H. and M.K. generated epitope-tagged constructs, bred mice, performed in vivo and cell culture experiments, collected data, designed the study, interpreted data, and wrote the paper; M.M.M. isolated hepatocytes and performed the chromatin immunoprecipitation and insulin sensitivity experiments; E.Y. and L.T. collected data; I.I. performed oxysterol and plant sterol measurements; M.W., S.A. and C.B.W. performed atherosclerotic lesion analysis; R.B.C and G.D. designed and performed lipidomics analyses; J.I. performed enterocyte ex vivo secretion studies; S.T. performed microarray analysis; J.H.L. performed histopathological analyses of liver sections; D.J.R., M.M.H., and S.Y. were involved in study design; A.R.T. designed the study, interpreted data, and wrote the paper. All authors discussed the results and commented on the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Reviewer Information Nature thanks C. Semenkovich, A. von Eckardstein and the other anonymous reviewer(s) for their contribution to the peer review of this work.
Extended data figures and tables
Extended Data Figure 1 Organ distribution of T39 mRNA expression.
T39 mRNA levels were assessed in chow-fed male WT and whole-body T39 knockout mice. SI, small intestine; n = 4 animals per genotype.
Extended Data Figure 2 Beneficial lipoprotein changes in T39-deficient mice fed HF/HC/BS diet.
a, HDL cholesterol of WT and whole-body T39 knockout mice fed chow or HF/HC/BS diet for 4 weeks, n = 4 per genotype for chow, 5 per genotype for HF/HC/BS diet. b, Serum ApoA-1 as determined by SDS–PAGE and (c) serum VLDL/chylomicron cholesterol of WT and whole-body T39 knockout mice fed HF/HC/BS diet for 4 weeks, n = 5 per genotype. d, Enterocyte mRNA expression, n = 5 per genotype. e, Quantification of enterocyte protein expression normalized to β-actin, n = 4 per genotype. f, Cholesterol secretion profiles of enterocytes collected from WT and whole-body T39 knockout mice fed chow or (g) HF/HC/BS diet. Enterocytes were incubated with taurocholate micelles containing [3H]cholesterol for 2 h, and then secreted lipoproteins were separated by density ultracentrifugation. Lipoprotein fractions are shown with fraction 1 being the most buoyant and fraction 12 the most dense, n = 3 per genotype, and replicated in two different experiments. For all panels, data are represented as mean ± s.e.m., *P < 0.05, **P < 0.01 and ***P < 0.001 by two-tailed Student’s t-test.
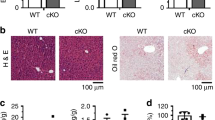
Extended Data Figure 3 Improved features of non-alcoholic steatohepatitis in T39−/− mice fed HF/HC/BS diet.
WT and whole-body T39 knockout mice were fed the HF/HC/BS diet for 18 weeks. a, Serum ALT, (b) liver size, (c) hepatic triglyceride, and (d) hepatic cholesterol content, n = 15–19 per genotype. Grading of (e) inflammatory cell infiltration and (f) hepatocellular ballooning degeneration on the basis of haematoxylin and eosin-stained sections, n = 6 per genotype. g, Ki-67 was immunohistochemically detected with diaminobenzidine (brown) in frozen liver sections with nuclei counterstained with haematoxylin (blue). Quantification of Ki-67-positive nuclei is shown on the right, mean of five fields, n = 6 animals/genotype. Data are represented as mean ± s.e.m., **P < 0.01 and ***P < 0.001 by two-tailed t-test.
Extended Data Figure 4 Improvements in the histological features of non-alcoholic steatohepatitis in liver-specific T39 knockout.
Enterocyte- and hepatocyte-specific T39 knockout and T39fl/fl control mice were fed HF/HC/BS diet for 21 weeks. a, Representative Masson’s trichrome stain of hepatic sections at 100× magnification, showing cytoplasm as red, collagen as blue, and nuclei as dark brown. Green arrowheads point to ballooned hepatocytes. Grading of (b) fibrosis severity, (c) inflammatory foci number and (d) extent of hepatocellular ballooning on the basis of Masson’s trichrome staining, n = 4 per genotype. Data are represented as mean ± s.e.m., **P < 0.01, ***P < 0.001 by one-way ANOVA.
Extended Data Figure 5 Decreased dietary cholesterol absorption in T39-deficient mice fed HF/HC/BS diet.
a, Hepatic Lxrα mRNA expression in mice fed HF/HC/BS diet for 5 weeks, n = 4 per genotype. b, Hepatic oxysterol content of WT and whole-body T39 knockout mice fed HF/HC/BS diet for 18 weeks, n = 5 WT and 4 T39−/−. c, Hepatic content of endogenous LXR ligands in control and liver-specific T39 knockout mice fed HF/HC/BS diet, n = 7 per genotype. d, Absorption of [14C]cholesterol administered by gavage to WT and whole-body T39 knockout with/without LXRα along with Poloxamer-407 injection to inhibit peripheral lipoprotein catabolism, n = 3 Lxrα+/+T39+/+, 4 Lxrα+/+T39−/−, 3 Lxrα−/−T39+/+, and 5 Lxrα−/−T39+/+. e, Enterocyte Npc1l1 mRNA expression in WT and whole-body T39 knockout mice, n = 5 per genotype. f, Hepatic plant sterol content of WT and whole-body T39 knockout mice with/without LXRα fed HF/HC/BS diet for 6 weeks, n = 4 Lxrα+/+T39+/+, 4 Lxrα+/+T39−/−, 4 Lxrα−/−T39+/+, and 6 Lxrα−/−T39+/+. g, Reverse cholesterol transport of WT and whole-body T39 knockout mice fed HF/HC/BS diet. 3H faecal excretion was measured over 3 days after an intravenous injection of [3H]cholesteryl ester-labelled HDL, n = 7 WT and 5 T39−/−. h, Absorption of [14C]cholesterol administered by gavage to tissue-specific T39 knockout mice fed HF/HC/BS diet for 5 weeks and injected with Poloxamer-407, n = 8 T39fl/fl, 7 Villin-Cre+T39fl/fl, and 9 Albumin-Cre+T39fl/fl. Data are represented as mean ± s.e.m., *P < 0.05, **P < 0.01, ***P < 0.001 by two-tailed t-test or two-way ANOVA for absorption studies.
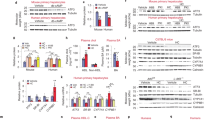
Extended Data Figure 6 Decreased SREBP-1 processing without altered hepatic insulin sensitivity in T39-deficient mice.
a, Hepatic protein expression of precursor SREBP-2 (upper band) and processed SREBP-2 (lower band) of WT and whole-body T39 knockout mice fed HF/HC/BS diet for 5 weeks. b, SREBP-1 processing ratio in WT and whole-body T39 knockout mice with/without LXRα fed HF/HC/BS diet for 6 weeks as based on quantification of an anti-SREBP-1 immunoblot, n = 4 per genotype. c, Hepatic triglyceride and (d) cholesterol content of WT and whole-body T39 knockout mice lacking LDLR fed WTD for 20 weeks, n = 5 per genotype. e, Protein expression of the nuclear form of SREBP-1 (top) and β-actin loading control (bottom) of WTD-fed WT and whole-body T39 knockout mice lacking LDLR fed WTD for 20 weeks. f, Hepatic gene expression in liver-specific T39 knockout and control animals fed HF/HC/BS diet for 18 weeks, following a fasting/refeeding protocol, n = 13 T39fl/fl and 7 Albumin-Cre+T39fl/fl. g, Intraperitoneal glucose tolerance of WT and whole-body T39 knockout mice fed HF/HC/BS diet after a 6 h fast, n = 8 WT and 12 T39−/−. h, Pyruvate tolerance of WT and whole-body T39 knockout mice fed HF/HC/BS diet after an overnight fast, n = 5 WT and 7 T39−/−. i, Hepatic Akt phosphorylation 5 min after portal vein delivery of insulin in WT or whole-body T39 knockout mice fed HF/HC/BS diet. Data are represented as mean ± s.e.m., *P < 0.05, ** P < 0.01, ***P < 0.001 by two-tailed Student’s t-test. See Supplementary Fig. 1 for gel source data.
Extended Data Figure 7 Improved lipoprotein profile and less advanced atherosclerotic lesions in Ldlr−/−T39−/− mice.
Mice on the Ldlr−/− background were fed WTD for 20 weeks. Serum lipoprotein cholesterol levels in (a) male and (b) female mice after 2 weeks on WTD, n = 5 per genotype per gender. c, Proximal aorta atherosclerotic lesion area after 20 weeks on WTD, n = 5 animals per genotype per gender. Data are represented as mean ± s.e.m., *P < 0.05, ***P < 0.001 by two-tailed Student’s t-test. d, Lesion severity as graded by a blinded observer on six sections per animal, n = 5 per genotype per gender. Lesion severity is expressed as number of observations of each complexity category and the difference in the categorical distribution of lesions between the two groups is indicated, *P < 0.05 based on a χ2-test.
Extended Data Figure 8 Increased LXRα protein in T39-deficient hepatocytes has implications for LXR target gene expression.
a, Immunoblots of endogenous LXRα in the nuclear (Nuc) and cytoplasmic (Cyto) fractions with an anti-LXRα antibody (top) 2 and 18 h after hepatocyte isolation and treatment with 2 μM GW3965. Histone H3 (bottom) and Hsp90 (middle) are shown as loading controls for nuclei and cytoplasm, respectively. Normalization of LXRα signal to the appropriate loading control is shown below the LXRα blot. The immunoblot is representative of three different sets. b, RXR occupancy over LXREs of WT and T39 knockout primary hepatocytes treated with GW3965, n = 4 WT and 5 T39−/−. c, Immunoblot showing validation of immobilized GW3965-mediated pulldown of endogenous LXRα from liver lysates. d, Relationship between magnitude of LXR target gene induction in T39 knockout mice and LXR affinity to active regulatory elements in the liver. See Supplementary Fig. 1 for gel source data.
Extended Data Figure 9 LXR protein preservation in T39-deficient gastrointestinal tissues raises HDL and protects from steatohepatitis.
In the absence of T39, LXR assembly into a multiprotein complex that conjugates it to ubiquitin moieties does not occur, and LXR is spared from proteasomal degradation. In enterocytes, the increase in LXRα/β protein upregulates Abca1 mRNA expression and promotes HDL production. In the liver, LXRα protein increase leads to the induction of Abcg5/8, which decreases dietary cholesterol uptake and increases cholesterol excretion, leading to cholesterol lowering. LXRα-mediated Insig2a prevents SREBP-1 processing in the fasted state, while Pcyt1a, and Cept1 induction increases microsomal membrane phosphatidylcholine content that continues to inhibit SREBP-1 processing in postprandial state. LPCAT3 upregulation results in increased incorporation of PUFA into phospholipid species which also contribute to the decrease in SREBP-1 processing. The decrease in nuclear SREBP-1 prevents the induction of lipogenic genes such as Fasn and Pnpla3. Therefore, unlike the gene expression profile that arises from potent synthetic ligands, increasing endogenous LXR protein levels preferentially upregulates cholesterol removal pathways while inhibiting lipogenesis.
Supplementary information
Supplementary Information
This file contains the uncropped blots from the Main and Extended Data Figures. (PDF 1183 kb)
Rights and permissions
About this article

Cite this article
Hsieh, J., Koseki, M., Molusky, M. et al. TTC39B deficiency stabilizes LXR reducing both atherosclerosis and steatohepatitis. Nature 535, 303–307 (2016). https://doi.org/10.1038/nature18628
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nature18628
- Springer Nature Limited
This article is cited by
-
Inhibition of ASGR1 decreases lipid levels by promoting cholesterol excretion
Nature (2022)
-
Proteome-scale profiling reveals MAFF and MAFG as two novel key transcription factors involved in palmitic acid-induced umbilical vein endothelial cell apoptosis
BMC Cardiovascular Disorders (2021)
-
Celastrol induces lipophagy via the LXRα/ABCA1 pathway in clear cell renal cell carcinoma
Acta Pharmacologica Sinica (2021)
-
LncRNA kcnq1ot1 promotes lipid accumulation and accelerates atherosclerosis via functioning as a ceRNA through the miR-452-3p/HDAC3/ABCA1 axis
Cell Death & Disease (2020)
-
The lipogenic LXR-SREBF1 signaling pathway controls cancer cell DNA repair and apoptosis and is a vulnerable point of malignant tumors for cancer therapy
Cell Death & Differentiation (2020)