
Abstract
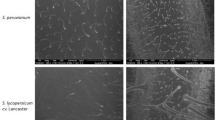
Hyphae of Phyllactinia corylea produce two kinds of special branches on the host surface: adhesion bodies which serve as fungal attachment and stomatopodia which enter the leaf through stomata. Conidial germination on host and non-host surfaces was examined with a scanning electron microscope to explain the stimuli responsible for development of the special branches, and the involvement of host recognition in the process. Conidia germinated within 4 h on host and non-host surfaces, but on non-host surfaces the emergence of the germ tube was not always directed towards the substratum. Adhesion bodies were formed from the tips of germ tubes at the first contact point on host and non-host surfaces. Development of stomatopodia was more specific and they were formed precisely over stomata on the host surface. Stomatopodia-like structures were occasionally formed over finely ridged leaf veins on the host surface and over some fine scratches on synthetic surfaces. The experiments showed that while conidial germination and development of adhesion bodies are in response to contact stimuli, the development of stomatopodia is a response to precise topographical signals, and the directional emergence and attached growth of germ tubes involve host recognition.
Similar content being viewed by others


References
Wheeler BEJ. Fungal parasites of plants. In: Ainsworth GC and Sussman AS, eds. The fungi – an advance treatise, Vol. 3. New York and London: Academic Press, 1968: 179–210.
Alexopoulose CJ, Mims CW. Introductory mycology. New Delhi: Wiley Eastern Limited, 1979: 309–321.
Kumar V, Babu AM, Sharma DD, Datta RK Penetration and infection process of Phyllactinia corylea on mulberry leaf causing powdery mildew – I. Asexual stage. J Phytopathol 1998; 146: 469–472.
Kunoh H, Kohno M, Tashiro S, Ishizaki H. Ultrastructural studies of powdery mildew of mulberry caused by Phyllactinia moricola (P. Henn.) Homma. Fitopatologia Brasileira 1980; 5: 11–20.
Staples RC, Grambow H-J, Hoch HC, Wynn WK. Contact with membrane grooves induces wheat stem rust uredospores germlings to differentiate appressorium but not vesicles, Phytopathol 1983; 73: 1436–1439.
Hoch HC, Staples RC, Whitehead B, Comeau J, Woff ED. Signaling for growth orientation and cell differentiation by surface topography in Uromyces. Science 1987; 235: 1659–1662.
Heath MC, Perumalla CJ. Haustorial mother cell development by Uromyces vignae on collodion membranes. Can J Bot 1988; 66: 736–741.
Read ND, Kellock LJ, Collins TJ, Gundlach AM. Role of topography sensing for infection-structure differentiation in cereal rust fungi. Planta 1997; 202: 163–170.
Bozzola JJ, Russell LD. Electron microscopy: Principles and techniques for biologists. Boston: John and Bartlett Publishers, 1992: 40–62.
Wynn WK. Tropic and taxic responses of pathogens to plants. Ann Rev Phytopathol 1981; 19: 237–255.
Kwon YH, Hoch HC. Temporal and spatial dynamics of appressorium formation in Uromyces appendiculatus. Exp Mycol 1991; 15: 116–131.
Stark-Urnau M, Mendegen K. Differentiation of aecidiosporeand uredospore-derived infection structures on cowpea leaves and on artificial surfaces by Uromyces vignae. Can J Bot 1993; 71: 1236–1242.
Rights and permissions
About this article
Cite this article
Babu, A.M., Kumar, V. & Datta, R.K. Development of special hyphal branches of Phyllactinia corylea on host and non-host surfaces. Mycopathologia 145, 29–33 (1999). https://doi.org/10.1023/A:1007040228477
Issue Date:
DOI: https://doi.org/10.1023/A:1007040228477