
Abstract
During freezing periods, nutrients (carbon and organic matter, etc.) are enriched in the water and sediment of lakes in cold-arid regions, leading to potential algal bloom outbreaks and other health risks to the ecosystem. Particularly, dissolved organic matter (DOM) is a critical component of the nutrients and plays an important role in the global carbon cycle. However, the mechanisms of DOM transfer between ice and water remain unclear. This study analyzed the influence of microbial community on DOM composition using 16 s RNA, 3DEEM, and FT-ICR MS in Daihai Lake and Wuliangsuhai Lake in the Yellow River Basin, China. According to the spectral analysis, the content of endogenous organic matter, such as humus, accounted for 40% of the total DOM in water, while the content of tryptophan and tyrosine accounted for 80% of the total DOM in ice. The results of mass spectrometry showed that lignin was the main component, and the content of organic matter in the ice was less than that in the water. Molecular structures of seven DOM coexisting in the lake ice and water were elucidated with adapted Kendrick-analogous network visualization, which clearly illustrates that long-chain DOM molecules are derived from small molecules, while other heteroatoms are complexed with the side groups. The positive correlations between CHO, CHNO, CHOS, CHOS and Actinomyces indicate that DOM actively interacted with the microbial community. 44% of CHO compounds have the same molecular formula in water, the content of CHOS in the water of the two lakes was closed to 7% higher than that in the ice. Meanwhile, DOM dynamically migrate between ice and water via interstitial water because of the solubility changes under microbial transformation, which has been proved by the decrease in the contents of the humus and tryptophan-like substances in the ice from the bottom to the surface and lower contents of carbohydrate and unsaturated aromatic hydrocarbon in the water than the ice. This study helps to predict the composition and structure of DOM during the migration in lakes and provides a scientific basis for environmental remediation with high concentration of carbon.
Highlights
• Seven DOM molecular structures in ice and water were identified and visualized.
• Tryptophan-like and Tyrosine-like DOM were dynamically migrated from water to ice surface.
• Long-chain DOM were easily buried in sediments under the action of microorganism.
• Bacteroides were significantly affected by salinity and temperature in eutrophic lakes.
AbstractSection Graphical Abstract
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Surface water in the lakes freezes in cold-arid regions during the winter while the nutrients keep migrating vertically, which is one of the important factors contributing to eutrophication once the ice thaws (Backer et al. 2006; Foley et al. 2005; Gomez-Salazar et al. 2012; Paer et al. 2013). Dissolved organic matter (DOM) is a crucial type of nutrient but biologically unstable with heavy metals and other substances in lakes (Zhou et al. 2021). DOM accumulation affects the balance of the ecosystems in the lakes (Wu et al. 2022). Therefore, studying the composition, structure, and bioavailability of DOM in eutrophic lakes in cold-arid regions is vital for maintaining these ecosystems. Currently, water and sediment are mostly studied media for DOM in lakes, with subglacial lakes in Antarctica being the focus of most studies. The total organic carbon (TOC) content of Antarctic lakes is between 0.3–0.4 mg/L, only 4.68%–5.71% of that in lakes in warm regions, because the poor lighting conditions limit the growth of phytoplankton (Chen et al. 2016; Faucher et al. 2021) By contrast, the nutrient content of lakes in the cold-arid regions of the Yellow River Basin plateau is significantly higher than that of lakes in Antarctica, because of climate change and human activities (Yang et al. 2021). Therefore, although the lakes in the Yellow River Basin share some common characteristics (i.e., lack of light, low DO content, and low microbial activity) with the lakes in Antarctica, they are at higher risk of potential eutrophication. The water quality keeps deteriorating and the eutrophication continues aggravating in these lakes in the Yellow River Basin, with the Daihai Lake and Wuliangsuhai Lake as two typical lakes. Ice is an important medium in lakes in the cold-arid regions during ice period. The microbial activity decreases in these lakes during the ice period, which slows down DOM decomposition and causes the DOM accumulation in the lakes, resulting in an abrupt DOM release in the warm ice-melting period and increase in eutrophication risk (Wu et al. 2018a, b). So far, few studies have been focused on the interaction between DOM and microbial community at the ice-water interface. Exploring the composition of DOM in the ice and water can trace the transport of DOM. The changes in DOM composition across different media reflects the environmental behavior of DOM during the ice period. Eutrophication in the lakes can be predicted by analyzing the source, migration and transformation of DOM. At present, studies on DOM in eutrophic lakes mainly focus on the type and structure of DOM (Xu et al. 2022; Hu et al. 2015; Hiriart-Baer et al. 2008; Zhu et al. 2013; Fu et al. 2005), the temporal and spatial distribution of DOM, and the migration and transformation of DOM under specific environmental conditions including microbial community (Jose et al. 2016; Xiong et al. 2002; Simon et al. 2016; Cho et al. 2017). With the analysis of DOM species transformation under microbial activity, the source and distribution of DOM can be analyzed from the biological perspective (Liu et al. 2020; Feng et al. 2023; Young et al. 2022).
As located in the cold-arid regions of northern China, the Daihai lake and the Wuliangsuhai Lake are facing similar problems with other lakes from the cold-arid regions globally, such as water shortage and area shrinkage (Tziotis et al. 2011; Bai et al. 2014; Cui et al. 2021; Yan et al. 2021; Oyewusi et al. 2021). Aiming at the specific environmental factors and microbial communities of these two lakes, the species and structure of DOM were analyzed to find the causes of eutrophication, providing a theoretical basis for solving the environmental problems of lakes. So far, few studies have been conducted on DOM during glaciation in eutrophic lakes. In this study, we studied the Daihai Lake and Wuliangsuhai Lake to investigate the composition, structure and environmental behavior of DOM in the ice and water in the two lakes during ice period, and identify the structure and function of the microbial communities in corresponding compartments. We also conducted a joint redundancy analysis of the microbial community and the environmental factors to predict the sources of DOM. The aims of this study were to improve the understanding of the migration and transformation of DOM, find the common ground of DOM in cold-arid regions and help to establish strategies to prevent or remediate the eutrophication of lakes specially during the freezing period and the year round.
2 Materials and methods
2.1 Study area and sample collection
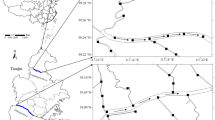
Daihai Lake (40.55–40.60°N, 112.64–112.72°E, Fig. 1a) is a saline lake, located in Inner Mongolia, 323 km to the east of Wuliangsuhai Lake (Song et al. 2019). The glaciation period lasts for 101–142 days. Six seasonal rivers flow into the lake, but the lake is primarily replenished by precipitation in winter (Gao et al. 2022). Nine sampling sites (DH1–DH9) were selected at the lake (Fig. 1b; Wang et al. 2016). DH1-DH8 sites were near the tributary of the lake in order to study the influence of terrigenous organic matter on lake water body, and DH9 was in the center of the lake, which is less disturbed by human activities. The average pH, ORP, DO, and SAL values were 9.42, 105.17 mV, 8.20 mg/L, and 16.08‰, respectively (Table S1). Nine water samples (from DH1-DH9) were collected for 3D-EEM analysis and 2 water samples (from DH 2 and DH3) for FT-ICR-MS analysis and microbial community characterization. Same numbers of samples for corresponding analyses were collected for ice.

Distribution of sampling sites at (a) the Daihai Lake and (b) the Wuliangsuhai Lake
Wuliangsuhai Lake (40.60–41.05°N, 108.72–108.95°E, Fig. 1b) is a typical grassy lake in Inner Mongolia, China. The glaciation lasts for 85–113 days per year (Yang et al. 2021). The lake receives most of the farmland drainage from the Hetao irrigation area and eventually flows into the Yellow River (Yang et al. 2020). During the freezing period, seven sampling sites (W1–W7) were selected in Wuliangsuhai Lake: W1 was near the water outlet, W2 was near the habitat of swan geese, W3 was located in the center of the lake, W4 was located in the tourist area, W5 was located on the eastern shore of the lake, W6 was located near the drainage channel of the lake, and W7 was located in the northern part of the lake, which was less disturbed by human activities and greater in water depth. The average pH, redox potential (ORP), dissolved oxygen (DO), and salinity (SAL) at the sampling sites were 8.85, 184.73 mV, 7.38 mg/L, and 2.17‰, respectively (Table S1). Water samples from Wuliangsuhai Lake (150 mL) were collected from each site using polyethylene bottles and stored in cold condition for 3D-EEM analysis. An additional 1 L water sample was collected respectively from W4 and W6 using polyethylene bottles and stored in cold condition for FT-ICR MS and microbial community analyses. The ice at each site was sampled vertically and divided evenly into three layers (labelled as up [U], middle [M], and down [D]). Approximately 150 mL of ice samples in each layer from Wuliangsuhai Lake were melted and stored in cold condition for 3D-EEM analysis. Approximately, 1 L of additional ice samples was respectively collected at W4 and W6 and stored in cold condition for FT-ICR MS and microbial community analyses.
2.2 3D-EEM fluorescence analysis
The water sample and melt water of the ice sample were filtered through a 0.45-μm membrane. A high-precision fluorescence photometer (HITACHI F-7000, Hitachi, Japan) was used to get the 3D-EEM data of the filtrate using a 150 W xenon arc lamp as the excitation source with a PMT voltage of 700 V, an excitation wavelength (EX) of 200–600 nm and an emission wavelength (EM) of 250–600 nm. The scanning speed of the spectrophotometer was 60,000 nm/min, and the passband width was 5 nm. Ultrapure water (iPurePlus, NeoLab, China) was used as the blank control.
2.3 FT-ICR-MS analysis
FT-ICR MS analysis was performed on the water and the melt water of ice samples. Internal standard methods were used for sample testing and molecular distribution analysis to ensure the reproducibility of the results. The samples were filtered through a 0.22-μm membrane to get the filtrates. 3 mL of the filtrate was, extracted by hydrophilic lipophilic equilibrium (HLB) solid phase extraction, acidified to pH = 2 with 2 mL of hydrochloric acid, and then eluted with 5 mL methanol for FT-ICR MS analysis. Samples after solid phase extraction were analyzed using the Bruker Apex ultra-FT-ICR MS (Apollo II ESI ionization source, a 9.4 T superconductor, and 3,500 V ionization voltage) to get the full spectra of the samples. The Bruker Daltonics software was used to analyze the molecular formula and number of double bonds in the organic matter.
2.3.1 S rRNA high-throughput sequencing
The water and melted ice samples were filtered through a 0.22-μm membrane to get the filtrates. The purity and concentration of DNA in the samples were determined using a NanoDrop 2000 spectrophotometer. DNA integrity was determined by gel electrophoresis on a 1% agarose gel (Aladdin, China). The samples were amplified by PCR and sequentially propagated at 95 °C for 3 min, 55 °C for 30 s, and 72 °C for 45 s. The bacterial 16 S rRNA universal primers 338 F (5 '-ACTCCTACGG-GAGGCAGCAG-3') and 806 R (5 '-GGACTACHVGGGTWTCTAAT-3') were used to amplify the V3–V4 region (Wang et al. 2016). The microbial population was classified according to the results, and the microbial species at the OUT level were counted, the corresponding alpha diversity index was calculated, and the microbial population richness in the sample was analyzed.
2.4 Data analyses
The analysis of 3D-EEM data was conducted using 3D fluorescence data computing software (EFC) “Conversion, Correction, Cognition, Comparison and Calculation of Excitation Emission Matrix” (Wang et al. 2016; He et al. 2015; Zepp et al. 2004; Bro et al. 1997). After blank correction, Raman correction, and PARAFAC, three components (C1–C3) were selected and compared with the EFC database to determine the fluorescence components of DOM in water and ice samples from the Daihai Lake and the Wuliangsuhai Lake.
Scatter plots of the O/C and H/C ratios of DOM were obtained from results of FT-ICR MS analysis, based on which the DOM molecules were classified into groups of lignin, lipids, proteins, carbohydrates, and unsaturated aromatic hydrocarbons. After sifting the FT-ICR MS data, the organic matter mass difference grid was constructed according to the formula and the number of double bonds. Molecular formulas that did not conform to the theory of molecular structure were eliminated (i.e., the number of double bonds was greater than the number of C atoms, and the number of O atoms was greater than the number of C atoms) By comparing with organic molecules with known formula and structure, the unknown structure of organic molecules was inferred, and then the properties and sources of these molecules were analyzed. Consequently, the properties and sources of organic matter molecules in the same group were analyzed. High-throughput sequencing analysis was conducted on the Majorbio cloud platform (www.majorbio.com), and the original data of microbial community were obtained from https://dataview.ncbi.nlm.nih.gov/object/PRJNA888824. All microbial community and environmental data were normalized. The Pearson correlation coefficients between the microbial data and environmental factors were obtained and considered significant at p < 0.01.
3 Results and analyses
3.1 Fluorescence characteristics of DOM
Based on the 3D-EEM fluorescence analysis of the samples from Daihai Lake, the spectrograms of the four types of fluorescent DOM were identified by consensus as humic-like, tryptophan-like, tyrosine-like, and quinone-like (Fig. 2a). Little difference of the four types of DOM was observed in the spatial distribution in the lake; only the contents of humus at DH2 and DH7 differed by approximately 10% from those at the other sites (Fig. 2b). Tryptophan is an important precursor of auxin in plants. Therefore, tryptophan-like substances may be secreted by the plants in the lake and along the coast. The high content of tryptophan-like substances in DH2 and DH7 was likely contributed by plant secretions along the lake. Tyrosine widely exists in microorganisms, animals, and plants; and there was little difference in the percentage of tyrosine-like substances at all sampling sites. The humus mainly comes from lake sediments, and is easy to migrate in water with the combination of metal elements to form organic colloids. The decomposition of humus by microorganisms might have led to difference in the content of humus-like substances at different sites. Water samples from the Wuliangsuhai Lake were analyzed using the 3D-EEM and three components of DOM were identified as humic-like, tryptophan-like and tyrosine-like (Fig. 2c). Except for W1, the spatial distributions of DOM in the water samples from the Wuliangsuhai Lake were not significantly different. W1 was near the replenishment point of the Wuliangsuhai Lake in the winter; therefore, the water quality and DOM content were different from those at the other sites. The humus-like content at W1 was 1.37 times that of the other sites. This indicates that the humus content was high in the reclaimed water flowing into the lake through farmland drainage. In contrast to the Daihai Lake, no quinones were detected in the Wuliangsuhai Lake, which indicates that the aquatic plant species in the lake were different from those in Daihai Lake or specific quinone-secreting plants were absent along the coast of the Wuliangsuhai Lake. Moreover, the tyrosine-like content in the water samples from the Wuliangsuhai Lake (25%–32%) was higher than that in the water samples from Daihai Lake (21%–29%), which may be attributed to the DOM from farmland drainage into the Wuliangsuhai Lake.

3D-EEM fluorescence maps and fluorescent DOM accumulations of ice and water in the Wuliangsuhai Lake and the Daihai Lake. (a) Fluorescence DOM spectrogram; (b) three-dimensional fluorescence percentage content of water samples from Daihai lake; (c) three-dimensional fluorescence percentage content of water samples from Wuliangsuhai Lake; (d) three-dimensional fluorescence percentage content of ice samples from Daihai Lake; (e) three-dimensional fluorescence percentage content of ice samples from Wuliangsuhai Lake
3D-EEM Spectra analysis identified three types of fluorescent DOM in the ice samples as humic-like, tryptophan-like and tyrosine-like in Daihai Lake (Fig. 2d). There was little spatial difference of humic-like substances in the Daihai Lake at different sampling locations, which may be due to the weak migration ability of humic substances as in the organic colloid. The difference in the distribution of tryptophan and tyrosine was mainly vertically, suggesting a vertical migration of the two substances. Tryptophan content in the lower layer of the ice sample was 1.08 times (p = 0.002) that in the upper layer, which may be caused by the secretions of plants growing in the lake. The content of tyrosine-like substances in the upper layer ice was 1.43 times (p = 0.001) that in the lower layer. Three components of DOM were also identified in the ice samples from the Wuliangsuhai Lake using the 3D-EEM as humic-like, tryptophan-like and tyrosine-like (Fig. 2e). The tyrosine content in the ice samples from W1 was the highest (45%–83%), because tyrosine substances mainly flow into the lake from external sources. Humus at other positions except the W1 position showed little difference. The vertical spatial distribution showed that the tryptophan content was the lowest (28%–41%) and the tyrosine content was the highest (52%–66%) in the middle layer of the ice samples. Tryptophan in the lower-layer of ice was mainly from plants in the lake, while it was mainly from plants along the lake in the upper-layer of ice. The lowest content of tryptophan was observed in the middle layers of ice.
3.2 Molecular composition of DOM
Based on the number of atoms (C, H, O, N, S, and P) detected by the FT-ICR MS, the molecular formulas of organic compounds were identified. The soluble organic compounds in the four types of samples (Wuliangsuhai Lake_water, Wuliangsuhai Lake_ice, Daihai Lake_water, and Daihai Lake_ice) were compared using a Venn Diagram (Fig. 3). The proportion of identical organic molecules in different medium samples of the Wuliangsuhai Lake was the highest, accounting for 46.68% of the total molecules. Meanwhile, the samples from two lakes (including ice and water) contained the same number of organic molecules accounting for 36.18% of the total. Richer species of DOM in Wuliangsuhai Lake corresponded to more complex ecosystems than the Dai Lake. The similarity of different DOM molecules was also reflected by the Venn diagram (Fig. 3). The CHO compounds in the water samples of Wuliangsuhai Lake and Daihai Lake were more similar, and 44.3% of them had the same molecular formula. CHOP was only detected in the ice samples from the two lakes, and the detected CHOP compounds in two lake samples were less similar; the identical molecules in the two lakes only accounted for 7.88% of the total detected CHOP compounds (SI. Figure 1). By comparison, the similarity of CHO compounds between different media in lakes was the highest, that is, the same molecules accounted for about 30.88% of the total. This may be because small CHO molecules are easy to migrate upward from water to ice, further confirming the inference of DOM migration process during lake glaciation.

Venn diagram of organic molecule species in the Daihai Lake and the Wuliangsuhai Lake water and ice samples. (a) Venn diagram for all molecular species; (b) Venn diagram for CHO molecules; (c) Venn diagram for CHNO molecules; (d) Venn diagram for CHOS molecules
Based on the length of carbon chain and the number of double bonds in the molecule, a bubble diagram was constructed to compare the degrees of unsaturation and molecular chain lengths of DOM in different samples (Fig. 4). The DOM molecules in the samples from the Daihai Lake contained more double bonds than those from the Wuliangsuhai Lake, indicating that the unsaturation was higher in the samples from the Daihai Lake, especially that the number of carbon atoms of some molecules was approximately 30 (Wang et al. 2016; Hu et al. 2023; Xu et al. 2020). In addition, the dense aggregation of large bubbles in the bubble diagram suggest that the organic molecules are more diverse in water. Moreover, the organic molecules in the water samples were mainly molecules with 10–20 highly saturated carbon atoms and 30–35 highly unsaturated carbon atoms. This indicates that the accumulation of long-chain and highly unsaturated molecules in DOM in water may be due to the microbial metabolism of DOM molecules. DOM with short carbon chains detected in the ice were mainly from the water.

Bubble diagram of the DOM molecular composition of ice and water samples from (a, b) the Daihai Lake and (c, d) the Wuliangsuhai Lake
The organic molecules were divided into lignin, lipid, protein, carbohydrate, and unsaturated aromatic sections according to the ratio of O/C and H/C (Fig. 5; Stedmon et al. 2008). The DOM molecules in the ice samples from the Daihai Lake were mostly lignin and unsaturated aromatic hydrocarbons, and the carbohydrate content was 4.34% higher than that in the water samples, indicating that lignin and unsaturated aromatic hydrocarbons easily migrated from the water to accumulate in the ice. Moreover, the microorganisms to decompose carbohydrates in the ice are lack in the ice, resulting in the accumulation of carbohydrates. The lipid content in the ice samples from the Wuliangsuhai Lake was higher than that from the Daihai Lake, while the unsaturated aromatic hydrocarbon content in the ice samples from the Wuliangsuhai Lake was 4.91% higher than that from the Daihai Lake. There are three possible causes for this result. First, there were fewer microorganisms metabolizing lipids and unsaturated aromatic hydrocarbons in the Daihai Lake than in the Wuliangsuhai Lake. Second, plants in the coastal environment of the Wuliangsuhai Lake secreted unsaturated aromatic hydrocarbons and lipids and flowed into the lake with surface runoff. Third, lipids and unsaturated aromatic hydrocarbons were produced from microbial metabolism in the ice. The contents of protein, lipid, carbohydrate, and other nutrients (sugars and other DOM components that can be used) in the ice samples were higher than those in the water samples in both lakes, indicating that the related metabolic activities of microorganisms in the ice were weak; however, there was still nutrient consumption in the water during winter.

Scatter diagrams of DOM molecular composition and histogram of DOM species in ice and water samples from the Daihai Lake and the Wuliangsuhai Lake. Scatter diagram of DOM molecular composition of water (a) and ice (b) samples from Daihai lake. Scatter diagram of DOM molecular composition of water (c) and ice (d) samples from Wuliangsuhai Lake; (e) Histogram of number of different DOM molecular species
DOM molecules analyzed by FT-ICR MS were classified into CHO, CHOS, and CHNO containing groups (Fig. 6). According to the ratios of H/C and O/C, DOM molecules were divided into saturated components (H/C > 1), unsaturated components (H/C ≤ 1), hypoxic components (O/C ≤ 0.5), and hyperoxic components (O/C > 0.5) (Simon et al. 2016). CHO molecule had the highest content among all DOM molecules, and from ice and water samples from both lakes, most DOM molecules were saturated and had a high oxygen component. The content of CHOS in the water of the two lakes was 6.9% higher than that in the ice. The content of CHOS in the water was 3.1% higher in the Daihai Lake than that in the Wuliangsuhai Lake, indicating that the abundance of microorganisms metabolizing organic sulfur was higher in the Wuliangsuhai Lake. In the CHNO compound from the Wuliangsuhai Lake ice samples, the unsaturated component accounted for 11.85% of the DOM. The CHNO content in the ice was 7.15% higher than that in the water in the Wuliangsuhai Lake, indicating that the content of CHNO aromatic organic molecules in the ice layer is high.

Patterns of DOM molecules in sea ice and water samples from the Daihai Lake and the Wuliangsuhai Lake. The four species in the horizontal coordinate represent the ice and water samples of Daihai Lake and Wuliangsuhai Lake, and the species in the vertical coordinate represents the DOM molecules of CHO, CHOS, and CHNO
3.3 Prediction of chemical structure of DOM
Based on the FT-ICR MS analysis, an intelligent grid of the chemical structures of the DOM molecules was constructed for the ice and water samples from the Wuliangsuhai Lake and Daihai Lake (Fig. 7). In the grid, multiple identical organic matter molecules (nodes) are connected with different bond units (lines) to construct a DOM compound. The molecular structure of the DOM was further analyzed based on its molecular formula and the number of double bonds. Methylene (CH2), amidogen (NH2), hydrogen phosphorus bonds (HP), O, S, and other bond units were used to differentiate the molecules. The number of double bonds of DOM identified by the FT-ICR MS analysis was used to predict the complete structure of DOM and classify different DOMs.

Intelligent grid coupled with the molecular structure of different types of organic matter in lake ice and water samples. The molecular formula and main characteristics of each molecule are shown in Table S2. In the grid, multiple identical organic matter molecules (nodes) are connected with different bond units (lines) to construct a DOM compound
Once the structure of DOM is predicted, it is possible to assess its properties and behaviors in the environment. Molecules with CH2 as the main bond unit of change can easily migrate in lipid rather than in water. Molecules with O as the bond unit change usually have hydroxyl radicals inserted and present good solubility in water; these molecules can also be esterified, which is one of the pathways of DOM transformation and metabolism. The increase of NH units, mainly in the form of amino insertion in the molecular side group, makes the molecule more water-soluble and can undergo condensation reaction with the carboxyl group of other molecules, thus enhancing its migration and transformation. The S units in the form of sulfhydryl groups are fat-soluble and difficult to form hydrogen bonds, resulting in lower mobility in water. The sulfhydryl groups can interact with proteins to form disulfide bonds. Protein molecules formed by sulfhydryl groups are more easily oxidized, thus enhancing the protein's conversion ability, which also leads to the conversion of sulfhydryl DOM molecules into more complex structures. HP units, mainly inserted in the form of phosphate groups, can form phosphate or phosphate ester, which is a potential threat to water eutrophication.
In this study, seven new molecular structures of DOM were identified in the ice and water samples respectively from both lakes with FT-ICR MS (Fig. 7 and Table S2). The newly identified common organic matter molecular structures are: (1) Lauryl sulfuric acid (C12H25O4S); (2) Tetraallylbenzene sulfonic acid (C18H29O3S); (3) Monoethyl 2-adamantyl succinate (C16H21O4); (4) Gallic acid (C15H22O5); (5) Phenyl-1-mercaptan-β-D-galactoside (C12H16O5S); (6) Stearic acid (C18H35O2); and (7) Melilotine (C15H18O8). Monoethyl 2-adamantyl succinate molecules were probably mainly derived from exogenous input according to the analysis. Combined with grid analysis, exogenous DOM might be transformed or combined with small molecules of DOM to form more complex DOM after flowing into the lake. Once the properties of DOM are predicted, it is helpful to understand their environmental behavior and explore lake eutrophication on the molecular level.
3.4 Microbial community analysis
Microbial community was analyzed with 16 S rRNA sequencing and characterized with. The microbial alpha diversity index including Chao index, Shannon index, and Simpson index (Maucourt et al. 2020). The Chao index reflects microbial species richness, Shannon index reflects microbial population diversity, and the Simpson index reflects the differences among microbial populations (Ren et al. 2019). The average Chao index, Shannon index and Simpson index of the water samples from the Wuliangsuhai Lake were 293, 384, and 390, and those of the Daihai Lake water samples were 226, 214, and 210, respectively. The average diversity indices of the water samples from the Wuliangsuhai Lake and the Daihai Lake were 3.6 and 0.04, respectively, and the average diversity index of the ice samples from the Wuliangsuhai Lake and the Daihai Lake were 3.4 and 0.06, respectively. Overall, little difference was observed in microbial community at phylum level between the Wuliangsuhai Lake and the Daihai Lake. The percentage accumulation maps of the first five abundant microbial communities (Proteus, Bacteroides, Actinomyces, Cyanobacteria, Verrucomicrobia) were constructed for the samples (Fig. 8a). Microbial species were most abundant in the sediments, followed by the water and ice samples. Overall, there were more microbial species in the Daihai Lake than in the Wuliangsuhai Lake. Bacteroides and Proteus species were the main microorganisms in the ice samples, and the amount of Bacteroides in the ice sample from the Wuliangsuhai Lake was twice that in the Daihai Lake.

Microbial community analysis. (a) Microbial species percentage content; (b) PCoA analysis of microbial species
Principal coordinate analysis (PCoA) was conducted at the operational taxonomic unit (OTU) level of the microbial community, and clustering was performed among different sampling sites and media (Fig. 8b). The OTU clustering analysis of microorganisms showed that the microbial differences were mainly reflected in the media and the natural condition of different lakes. Redundancy analysis (RDA) was performed on the samples from different sampling sites and media with the 6 identified most abundant microbial species and environmental factors (Fig. 9). According to the correlation from the RDA analysis, the dominant bacteria, such as Bacteroides, Actinomyces, and Cyanobacteria, were significantly affected by salinity and temperature in the lakes. The pH, salinity, and TOC were positively correlated with the abundance of Proteus, Actinomyces, Cyanobacteria, and Green Bay bacteria but negatively correlated with Bacteroides and Gammaproteobacteria. When different molecular formula of DOM (all DOM molecules, CHO, CHNO, CHOS) were used as observed value, there was no significant difference in the correlation between microorganisms (except actinobacteria) and environmental factors, indicating that these microorganisms (Bacteroides, Actinomyces, Alphaproteobacteria Gammaproteobacteria and Cyanobacteria) were mainly affected by environmental factors. The results showed that environmental factors had little effect on Actinomyces during the winter, and other microorganisms were affected by environmental factors and DOM to the same extent.

Redundancy analysis of microbial species and environmental factors. Redundancy analysis of CHO molecules (a), CHOS molecules (b), CHNO molecules (c), CHNOS molecules (d)
4 Discussion
4.1 Effects of lake microorganisms on DOM components during freezing periods
The lack of light during the freezing period reduced the photosynthesis of algae, resulting in less organic matter in water and impacting the microbial community (Cross et al. 2005; Sattler et al. 2004). Except for cyanobacteria affected by specific environmental factors, the microbial community structure in the lakes in the cold-arid regions of northern China during the freezing period was similar to that in the Antarctic, with Actinomyces, Bacteroides, Proteus, and other anaerobic bacteria as the dominant species (Wang et al. 2016). In the symbiosis between plankton and microorganisms, plankton provides necessary nutrients and space for microorganisms and affects their metabolic capacity, thus affecting the cycling of carbon and nitrogen in the lake (Siegert et al. 2001). DOM can also be decomposed by microorganisms in the absence of light as proved in a previous study (Wang et al. 2021). Considering the effect of light reduction on the decomposition of DOM during freezing periods, it was predicted that the loss of DOM in the water samples caused by microbial and photolytic action in the melting process was 3.9%–11.9%. The fluorescent DOM in the sediments of the lake, which were detected primarily as terrestrial DOM, is preferred for preservation in pore water under anoxic conditions (Li et al. 2024). As the transition between sediment and water, pore water plays a role in the upward migration of DOM in lakes. One of the fluorescent DOM components, tryptophan-like DOM in the pore water of sediments was less abundant because of the decomposition of microorganisms (Isabelle et al. 2013), while other fluorescent DOM components (humus-like, tyrosine-like) had similar amounts in pore-water as in water.
DO concentrations were measured in situ to be 8.20 and 11.03 mg/L in the waters of the Daihai Lake and the Wuliangsuhai Lake in early January during the freezing period, respectively similar and higher compared to that of the Keweenaw lake, a freshwater lake in Michigan, USA. Although there was limited gas exchange with the atmosphere during the ice period, the DO level in the water of the Keweenaw lake could still support the aerobic metabolism of microorganisms (Isabelle et al. 2013). This confirms the results of our study that aerobic microorganisms are the dominant microorganism in the freezing period of the two lakes. According to the DOM configuration by the Kendrick grid, the DOM in the two lakes during the freezing period was derived from the metabolic decomposition of sugars and proteins, which may have occurred during the aerobic metabolism (Butler et al. 2019). Moreover, the microbial community in the freezing period is relatively stable compared to that in the non-freezing period, but the microbial community under the ice layer was still growing actively (Li et al. 2021). According to the analysis of chlorophyll in the ice, algae were one type of the active microbial communities (Butler et al. 2019). Water bloom may occur during the freezing period in the Wuliangsuhai Lake and Daihai Lake with the accumulation of DOM and other nutrients. Previous studies have proved that water blooms could occur in lakes during the freezing period (Pernthaler et al. 1998), which provides addition evidence for our speculation.
4.2 Influencing factors affecting DOM molecular composition in grassy and saline lakes
The Wuliangsuhai Lake is a typical agricultural irrigation regressive lake with naturally growing plants. It is greatly affected by the drainage and inflow of surrounding farmland, that is, it is greatly influenced by human activities. The Daihai Lake is gradually salinized due to the large annual evaporation and shrinkage of the water body. The decomposition of macrophytes and algae contributes significantly to the DOM in grassy lakes (Bižić-Ionescu et al. 2014). Therefore, the type of plants is a controlling factor for the molecular composition of DOM in these lakes. Studies have shown that the DOM of macroplants (65.30 mg/kg) and algae (95.32 mg/kg) are main resource of that in the lake water (0.83–2.33 mg/L) (Bižić-Ionescu et al. 2014; Zhou et al. 2018; Huang et al. 2023; Paul et al. 2008; Feng et al. 2020). Therefore, species and content of DOM in lakes are greatly affected by plants and algae. Furthermore, the decomposition of macroplants was more difficult as proved in a previous research (Wang et al. 2016). Therefore, the contribution of macroplants to DOM in lakes should be smaller than that of algae. The main factor influencing DOM in salinized lakes is the salinity, which influences the microbial community causing the difference in DOM molecular composition (Yang et al. 2020). Typical microorganisms in salt lakes include Firmicutes, Actinomycetes, and Alphaproteobacteria (Baker et al. 2001); Firmicutes degrade plant residues and secrete tyrosine-like substances during algal decomposition process of algae (Banda et al. 2021; Kielak et al. 2016). Actinomyces decomposes proteins and lipids, and their secreted enzymes affect the environmental behavior of phosphorus (Wu et al. 2018a, b). These microorganisms have an important impact on the DOM composition of the lake.
In addition to the influence of environmental factors on the endogenous DOM of lakes, exogenous inputs from the surrounding environment also influence DOM composition. The grassy lake in this study, i.e., the Wuliangsuhai Lake, is surrounded by the return water of farmland irrigation, and the exogenous DOM drained from farmland has a significant influence on the composition of DOM (Subramani et al. 2012). Moreover, studies have shown that land-based DOM inputs also play a major role for saline lakes such as the Daihai Lake (Yang et al. 2020). In this study, the antioxidants and supplements used for FT-ICR MS analysis were derived from exogenous DOM inflows. The inflow of plant secretions affects the DOM components in the lake. The plants that secreted quinones around the Daihai Lake contributed quinone components in the fluorescent DOM detected in the water samples during the freezing period. Moreover, in the DOM structure analysis using the Kendricks grid, the organic matter from Melilotus officinalis was identified which could be formed by a combination of plant secretions and various organic matter; therefore, exogenous input is an important source of DOM in lakes during the winter ice period.
4.3 Microbial network and function in ice and water
According to the results of this study, a certain amount of Proteus, Bacteroides and other microorganisms still exist in the ice and water phases during winter, while less autotrophic microorganisms such as cyanobacteria were observed in water. In addition, the microbial community tends to be stable in winter due to low temperature and poor light conditions compared with other seasons (Li et al. 2021). In this study, autotrophic microorganisms such as cyanobacteria were detected in the water, indicating that photosynthesis may occur during winter and affect DOM composition in the lakes. A previous study also proves that photosynthesis by plankton, submerged plants, and epiphytic algae still occur over winter in shallow lakes in mid‐latitudes when there is no snow cover on the ice (Nuen et al. 2020). Moreover, microbial diversity rarely increases in winter due to the reduced migration of microorganisms from the bottom to the top (Tulonen et al. 1994; Techatmann et al. 2016). In addition to the reduced migration ability of microorganisms in winter, dormancy mechanism is also one of the reasons affecting microbial diversity. It has been proven that microbial dormancy is a mechanism for maintaining community diversity at low nutrients (Jones et al., 2010). In certain nutrient-poor systems, approximately 40% of the genes detected was from dormant cells (Li et al. 2021). These results indicated that microorganisms in the dormant period may not participate in the metabolism of nutrients in the environment but still compose high population richness (Jones et al. 2010; Churchill et al. 2004). Therefore, there are microbial metabolic activities in lake ice and water in winter, but the overall activity is low, and the influence on DOM species is less than that in other seasons.
5 Conclusion
In this study, the intricate interplays between DOM, microbes, and environmental factors in the ice and water were investigated in the Wuliangsuhai Lake, a typical grass lake, and the Daihai Lake, a typical saline lake, in the cold-arid regions in China during winter. Important findings were revealed with the application of several advanced techniques, namely 3D-EEM, FT-ICR MS, and 16 S rRNA. The findings suggest that fluorescent DOM in the ice primarily resulted from the upward migration of water. The DOM molecules were difficult to decompose because of the long carbon chains with high unsaturation. Despite being in a relatively inactive mode, microorganisms still play a crucial role in the decomposition of these macromolecular DOM in the ice. The seven common DOM components from the two lakes prove that microbial metabolism, plant secretions, and exogenous inflow were the three main sources of DOMs in the lakes. An important Kendrick-analogous network was also developed to visualize the structure of DOMs and help to predict their environmental behaviors. These results help us to understand the biogeochemical cycle of nutrients between different compartments in the lakes during the freezing period and the possibility of eutrophication throughout the year. This study provides implications to address the issue of eutrophication in water bodies and helps facilitate the investigation of the carbon cycle, carbon storage and other environmental processes in the ecosystems.
Availability of data and materials
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- DOM:
-
Dissolve organic
- TOC:
-
Total organic carbon
- 3D EEM:
-
Three-dimensional fluorescence spectroscopy
- PARAFAC:
-
Parallel factor analysis
- FT ICR MS:
-
Fourier-transform ion cyclotron mass spectrometry
- H/C:
-
Ratios of hydrogen to carbon
- O/C:
-
Ratios of oxygen to carbon
- ORP:
-
Redox potential
- DO:
-
Dissolve oxygen
- SAL:
-
Salinity
- EX:
-
Excitation wavelength
- EM:
-
Emission wavelength
- PCoA:
-
Principal coordinate analysis
- RDA:
-
Redundancy analysis
References
Backer LC, McGillicuddy DJ Jr (2006) Harmful algal blooms at the Interface between coastal oceanography and human health. Oceanography 19:94–106
Baker A (2001) Fluorescence excitation–emission matrix characterization of some sewage-impacted rivers. Environ Sci Technol 35(5):948–953
Banda JF, Zhang Q, Ma LQ, Pei LX, Du ZR, Hao CB, Dong HL (2021) Both PH and salinity shape the microbial communities of the lakes in Badain Jaran Desert. NW China Sci Total Environ 791:148108
Bai YH, Liang JS, Liu RP, Hu CZ (2014) Qu JH (2014) Metagenomic analysis reveals microbial diversity and function in the rhizosphere soil of a constructed wetland. Environ Technol 35(17–20):2521–2527
Bižić-Ionescu M, Amann R, Grossart HP (2014) Massive regime shifts and high activity of heterotrophic bacteria in an ice-covered lake. PLoS ONE 9:113611
Bro R (1997) PARAFAC. Tutorial and Applications Chemometr Intell Lab 38(2):149–171
Butler TM, Wilhelm AC, Dwyer AC, Webb PN, Baldwin AL, Techtmann SM (2019) Microbial community dynamics during lake ice freezing. Sci Rep 9:6231
Chen M, Kim JH, Nam SI, Niessen F, Hong WL, Kang MH, Hur J (2016) Production of fluorescent dissolved organic matter in Arctic Ocean sediments. Sci Rep. 6:39213
Cho YJ, Birdwell JE, Hur M, Lee JH, Kim BJ, Kim SH (2017) Extension of the Analytical Window for Characterizing Aromatic Compounds in Oils Using a Comprehensive Suite of High-Resolution Mass Spectrometry Techniques and Double Bond Equivalence versus Carbon Number Plot. Energy Fuels 31(8):1520–5029
Churchill JH, Kerfoot WC, Auer MT (2004) Exchange of water between the Keweenaw Waterway and Lake Superior: characteristics and forcing mechanisms. J Great Lakes Res 30:55–56
Cross WF, Benstead JP, Frost PC, Thomas SA (2005) Ecological stoichiometry in freshwater benthic systems: recent progress and perspectives. Freshw Biol 50(11):1895–1912
Cui HZ, Zhou WY, Deng Y, Zheng BY, Zhang Q, Zhang ZY, Lu T, Qian HF (2021) Meta-transcriptomic profiling of functional variation of freshwater microbial communities induced by an antidepressant sertraline hydrochloride. Sci Total Environ 10:147434
Faucher B, Lacelle D, Marsh NB, Jasperse L, Clark ID, Andersen DT (2021) Glacial lake outburst floods enhance benthic microbial productivity in perennially ice-covered Lake Untersee (East Antarctica). Commun Earth Environ 2:211
Feng WY, Yang F, Zhang C, Liu J, Song FH, Chen HY, Zhu YR, Liu SS, Giesy JP (2020) Composition characterization and biotransformation of dissolved, particulate and algae organic phosphorus in eutrophic lakes. Environ Pollut 265:114838
Feng W, Wang T, Zhu Y, Sun F, Giesy JP, Wu FC (2023) Chemical composition, sources, and ecological effect of organic phosphorus in water ecosystems: a review. Carbon Res 2:12
Foley JA, DeFries R, Asner PG, Barford C, Bonan G, Carpenter SR, Chapin FS, Coe MT, Daily GC, Gibbs HK, Helkowski JH, Holloway T, Howard EA, Kucharilk CJ, Monfreda C, Patz JA, Prentice IC, Ramankutty N, Snyder PK (2005) Global consequences of land use. Science 309(5734):570–574
Fu PQ, Liu CQ, Wu FC (2005) Three-dimensional excitation emission matrix fluorescence spectroscopic characterization of dissolve organic matter. Spectroscopy Spect Anal 25(12):2024–2028
Gao JY, Feng WY, Yang F, Liu J, Fan WH, Wang Y, Zhang Q, Yang WZ (2022) Effects of water quality and bacterial community composition on dissolved organic matter structure in Daihai lake and the mechanisms. Environ Res 214:114109
Gomez-Salazar C, Coll M, Whitehead H (2012) River dolphins as indicators of ecosystem degradation in large tropical rivers. Ecol Indicat 23:19–26
He W, Hur J (2015) Conservative behavior of fluorescence EEM-PARAFAC components in resin Fractionation processes and its applicability for characterizing dissolved organic matter. Water Res 83:217–226
Hiriart-Baer VP, Diep N, Smith REH (2008) Dissolved organic matter in the great lakes: role and nature of allochthonous material. J Great Lake Res 34(3):383–394
Hu CK, Xu H, Shi SW, Lan JH, Zhou K, Zhang J, Song YP, Wang J, Fu PQ (2023) Sedimentary organic matter molecular composition reveals the eutrophication of the past 500 years in Lake Daihai. Inner Mongolia Environ Res 227:15
Hu QD, Wang HJ, Li J, Hu CM, Zhang Y, Yu T, Zhang Y (2015) Various Inflows to taihu lake in autumn spectroscopy characteristics and DOM Flux. Environ Sci Technol 38(3):152–158
Huang L, Wang WC, Wei GJ, Li SD, Yang MD, Wu Y, Luo QL, Huang ZW, Fang HY, Fan ZY, Zeng FT (2023) Linking the source, molecular composition, and reactivity of dissolved organic matter in major rivers across the pearl river delta. J Clean Prod 420(25):138460
Isabelle L, Natalie M (2013) Dissolved organic matter photolysis in Canadian arctic thaw ponds. Environ Res Lett 8(3):035026
Jones SE, Lennon JT (2010) Dormancy contributes to the maintenance of microbial diversity. Proc Natl Acad Sci USA 107:5881–5886
Jose J, Melendez P, Mónica J, Martínez M, Almas TA, Pedro S, Fadini AA, Mozeto M, Nogueira E (2016) Characterization and comparison of riverine, lacustrine, marine and estuarine dissolved organic matter by ultra-high resolution and accuracy Fourier transform mass spectrometry. Org Geochem. 101:99–107
Kielak AM, Barreto CC, Kowalchuk GA, Veen JA, Kuramae EE (2016) The ecology of acidobacteria: moving beyond genes and genomes. Front Microbiol 7:1–16
Li SD, Lü LF, Wu YF, Zhao ZL, Huang CC, Huang T, Yang H, Ma XH, Jiang QL (2021) Investigation on depth-dependent properties and benthic effluxes of dissolved organic matter (DOM) in pore water from plateau lake sediments. Ecol Ind 125:107500
Li TT, Ruan MQ, Cao YH, Feng WY, Song FH, Bai YC, Zhao XL, Wu FC (2024) Molecular-level insights into the temperature-dependent formation dynamics and mechanism of water-soluble dissolved organic carbon derived from biomass pyrolysis smoke. Water Res. 252:121176
Liu SS, He ZQ, Tang Z, Liu LZ, Hou JW, Li TT, Zhang YH, Shi Q, Giesy JP, Wu FC (2020) Linking the molecular composition of autochthonous dissolved organic matter to source identification for freshwater lake ecosystems by combination of optical spectroscopy and FT-ICR-MS analysis. Sci Total Environ 703:134764
Maucourt B, Vuilleumier S (2020) Bringel F (2020) Transcriptional regulation of organohalide pollutant utilisation in bacteria. FEMS Microbiol Rev 44(2):189–207
Nuen J, Li S, Cheng L, Swan L (2020) Analysis of nonpoint source pollution load in Ulansuhai Nur basin. Arab J Geosci 13:1138
Oyewusi Habeebat A, Wahab Roswanira A, Huyop F (2021) Whole genome strategies and bioremediation insight into dehalogenase-producing bacteria. Mol Biol Rep 48(3):2687–2701
Paerl HW, Otten TG (2013) Harmful cyanobacterial blooms: causes, consequences, and controls. Microb Ecol 65:995–1010
Paul VJ (2008) In: Hudnell, H.K. Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs. Springer New York. 239–257
Pernthaler J, Glöckner FO, Unterholzner S, Alfreider A, Psenner R, Amann R (1998) Seasonal community and population dynamics of pelagic bacteria and archaea in a high mountain lake. Appl Environ Microbiol 64:4299–4306
Ren Z, Martyniuk N, Oleksy IA, Swain A, Hotaling S (2019) Ecological Stoichiometry of the Mountain Cryosphere. Frontiers 7:360
Siegert M, Ellis-Evans J, Tranter M, Mayer C, Petit JR, Salamatin A, Priscu JC (2001) Physical, chemical and biological processes in Lake Vostok and other Antarctic subglacial lakes. Nature 414:603–609
Sattler B, Waldhuber S, Fischer H, Semmler H (2004) Microbial activity and phylogeny in ice cores retrieved from Lake Paula, a newly detected freshwater lake in Antarctica. SPIE, Proc, p 5555
Simon HJ, van Agthoven MA, Lam PY, Floris F, Chiron L, Delsuc MA, Rolando C, Barrow MP (2016) Uncoiling collagen: a multidimensional mass spectrometry study. Analyst 141(1):157–165
Song S, Li C, Shi X, Zhao S, Tian W, Li Z (2019) Under-ice metabolism in a shallow lake (Wuliangsuhai) in Inner Mongolia, in cold and arid climate zone. Freshw Biol 00:1–11
Stedmon CA (2008) Bro R (2008) Characterizing dissolved organic matter fluorescence with parallel factor Analysis: a tutorial. Limnol Oceanogr Methods 6(11):572–579
Subramani R, Aalbersberg W (2012) Marine Actinomycetes: an ongoing source of novel bioactive metabolites. Microbiol Res 167(10):571–580
Techtmann SM, Hazen TC (2016) Metagenomic applications in environmental monitoring and bioremediation. J Ind Microbiol Biotechnol 43:1345–1354
Tulonen T, Kankaala P, Ojala A, Arvola L (1994) Factors Controlling Production of Phytoplankton and Bacteria under-Ice in a Humic. Boreal Lake J Plankton Res 16:1411–1432
Tziotis D, Hertkorn N, Schmitt-Kopplin P (2011) Kendrick-analogous network visualisation of ion cyclotron resonance Fourier transform mass spectra: improved options for the assignment of elemental compositions and the classification of organic molecular complexity. Eur J Mass Spectrom. 17(4):415–421
Wang CH, Jiang HL (2016) Chemicals used for in situ immobilization to reduce the internal phosphorus loading from lake sediments for eutrophication control. Crit Rev Environ Sci Technol 46(10):947–997
Wang Q, Hao Z, Ding RR, Li HB, Tang XM, Chen FZ (2021) Host Dependence of Zooplankton-Associated Microbes and Their Ecological Implications in Freshwater Lakes. Water 13:2949
Wang XW, Yang H, Cai YP, Yu CX, Yue WC (2016) Identification of optimal strategies for agricultural nonpoint source management in Ulansuhai Nur watershed of Inner Mongolia. China Stoch Environ Res Risk Assess 30:137–153
Wu FC, Li F, Zhao X (2022) Meet the challenges in the “Carbon Age.” Carbon Res 1:1
Wu XQ, Yang CB, Chang YW, Duan Q (2018a) Study on Water Quality Monitoring of Daihai Lake Based on Remote Sensing Technology. Web Poceedings 462–467
Wu XJ, Yang H, Cheng R, Sheng Y, Han JG, Li PP (2018b) Bacterial community structures in sediments are affected by different plants in hongze lake wetlands. J Agro-Environ Sci 37(5):984–991
Xiong SX, Pu D, Xin B, Wang GH (2002) Structural characterization of anhydroicaritin glycosides using ESI-FT-ICR mass spectrometry. Chin Chem Lett 2002:1001–8417
Xu W, Gao J, He C, Shi Q, Hou Z, Zhao HZ (2020) Using ESI FT-ICR MS to Characterize Dissolved Organic Matter in Salt Lakes with Different Salinity. Environ Sci Technol 54(20):12929–12937
Xu YY, Gun Z, Zhao JW, Cheng X (2022) Variations in lake water storage over Inner Mongolia during recent three decades based on multi-mission satellites. J Hydrol 609:127719
Yan XG, Li WG, Liang DM, Caiyin Q, Zhao GR, Zhang ZJ, Ma WZ (2021) Qiao JJ (2021) De novo assembly of the Mylia taylorii transcriptome and identification of sesquiterpene synthases. Arch Biochem Biophys 698(15):108742
Yang F, Cen R, Feng W, Zhu Q, Leppa¨ranta M, Yang Y, Wang X, Liao H (2021) Dynamic simulation of nutrient distribution in lakes during ice cover growth and ablation. Chemosphere 281:130781
Yang HY (2020) Brief analysis on water ecological environment problems and control measures of Daihai Lake. Inner Mongolia Water Resource 9(1):33–34
Young RB, Pica NE, Sharifan H, Chen H, Roth HK, Blakney GT, Borch T, Higgins CP, Kornuc JJ, McKenna AM, Blotevogel J (2022) PFAS analysis with ultrahigh resolution 21T FT-ICR MS: suspect and nontargeted screening with unrivaled mass resolving power and accuracy. Environ Sci Technol. 56(4):2455–2465
Zepp RG, Sheldon WM (2004) Moran MA (2004) Dissolved organic Fluorophores in southeastern US coastal waters: correction method for eliminating Rayleigh and Raman scattering peaks in excitation-emission matrices. Mar Chem 89(1–4):15–36
Zhou Y, Davidson TA, Yao X, Zhang Y, Jeppesen E, de Souza JG, Wu H, Shi K, Qin B (2018) How autochthonous dissolved organic matter responds to eutrophication and climate warming: evidence from a cross-continental data analysis and experiments. Earth-Sci Rev 185:928–937
Zhou YQ, Yao XL, ZhouZhaoWangJangTianZhangPodgorskiSpencerKothawalaJeppesenWu LZHXLKSWYLDCRMDNEFC (2021) How hydrology and anthropogenic activity influence the molecular composition and export of dissolved organic matter: Observations along a large river continuum. Limnol 66(5):1730–1742
Zhu P, Hua ZL, Li HM (2013) PARAFAC Method Used for Analysis of Three-Dimensional Fluorescence Spectra of DOM in Taihu Lake. Spectroscopy Spect Anal 33(6):1619–1625
Acknowledgements
Not applicable.
Funding
This research was jointly supported by the National Natural Science Foundation of China (42177400, 41907338, 42207325) and Science and Technology Plan Project of the Inner Mongolia Autonomous Region (2022YFHH0044).
Author information
Authors and Affiliations
Contributions
Tengke Wang analyzed the data, developed the graphical representations and wrote the manuscript. Weiying Feng and Jing Liu modified the manuscript. Wenhong Fan provided experimental platform. Tingting Li and Fanhao Song modified the manuscript. Fang Yang modified the manuscript and provided fund support. Haiqing Liao provided constructive guidance. Matti Leppäranta improved the manuscript language.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Additional information
Handling editor: Baoshan Xing.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wang, T., Feng, W., Liu, J. et al. Eutrophication in cold-arid lakes: molecular characteristics and transformation mechanism of DOM under microbial action at the ice-water interface. Carbon Res. 3, 42 (2024). https://doi.org/10.1007/s44246-024-00126-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s44246-024-00126-z