
Abstract
Fermentation of quinoa by probiotics provides higher nutritional value and can be considered as a significant source of bioactive compounds and alive probiotics for the human body. Moringa leaves powder (MLP) at the levels of 0.25 and 0.50% were used as an additional prebiotic source to supply quinoa fermentation by Lactobacillus plantaram ATCC 14917 and Lactobacillus delbrueckii ssp. Bulgaricus EMCC 11102 and produce healthier quinoa products. The results indicated that supplementation of fermented quinoa products with MLP at bath levels increased its contents of free phenolics and water extractable arabinoxylans as well as enhanced its antioxidant activity and phytate degradation. Fermented quinoa products with 0.50% MLP showed better chemical properties than fermented quinoa products with 0.25% MLP. Furthermore, supplementation of fermented quinoa products with MLP (0.50%) displayed positive effects on blood glucose levels of the experimental rats. Further studies are required on the test of more kinds of probiotics with different concentration of MLP in quinoa fermentation in the future.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Quinoa (Chenopodium quinoa) is a pseudo-cereal plant domesticated in the Indian region 5000 years ago [1]. It is known for its high nutritional value and high contents of essential amino acids, minerals and dietary fibres. Due to its significant content of phenolic compounds, quinoa showed antioxidant activity which associated with many health benefits [2, 3].
Probiotic bacteria are mainly used in dairy products and show the biggest share of the probiotic food market [4]. However, some disadvantages of dairy products containing probiotics have been recorded for many consumers worldwide such as lactose intolerance; allergy to β-caseine in cow’s milk; high content of cholesterol and saturated fatty acids in dairy products as well as high costs of milk [5]. Probiotic cereal foods may be produced as a good alternative to avoid drawbacks of fermented dairy products [4]. Several studies have evaluated the fermentation of quinoa by probiotics for production of fermented quinoa products with higher healthy benefits [6,7,8].
Moringa oleifera leaves powder (MLP) produce numerous healthy benefits and have good amounts of crude protein, crude fiber, extract ether, carbohydrates, energy, minerals, vitamins, β-carotene and polysaccharides [9, 10]. Also, MLP displayed a prebiotic effect may be due to its significant content of prebiotic compounds such as oligosaccharides [11, 12]. Also, the high antioxidant activity and phenolic compounds of MLP were reported by many previous studies [13,14,15].
This is the first study evaluates the effect of supplementation with MLP as a source of prebiotics to produce healthier fermented quinoa products. The changes in the free phenolic content and antioxidant activity as well as the levels of phytate and water extractable arabinoxylans of fermented quinoa products were determined during storage period. Also, the effects of MLP (0.50%) on the blood glucose levels of the experimental rats were estimated during the feeding period for 30 days.
2 Materials and methods
2.1 Materials
2.1.1 Raw materials and reagents
Quinoa seeds were purchased from Agricultural Research Center, Giza, Egypt. Moringa (Moringa oleifera) leaves were obtained from a local farm located in Albalyana city, Sohag, Egypt, and sugar was purchased from a local market in Assiut city, Egypt. Chemicals used in the analytical methods were produced by sigma chemical co. (St. Louis, MO, USA) and obtained from El-Gomhouria Trading Chemicals and Drugs Company, Assiut city, Egypt. The reagents used in the biological experiment were purchased from Spectrum Company in Assiut city, Egypt.
2.1.2 Probiotic strains
Lactobacillus plantarum ATCC 14917 and L. delbrueckii ssp. Bulgaricus EMCC 11102 were purchased from Microbiological Resources center (Cairo MIRCEN) Ain Shams University, Cairo, Egypt.
2.2 Methods
2.2.1 Preparation of quinoa fermentation
2.2.1.1 Preparation of raw materials and quinoa blends
Grain quinoa seeds were washed and soaking for 24 h in water which was discarded and changed every 3 h. The soaked quinoa seeds was dried at 60 °C for 8 h and milled to get the whole quinoa flour (WQF). The Moringa leaves were dried and milled to produce MLP, which stored in a cool dry place until the experimental work. The blends of WQF, MLP and sugar were prepared and tap water was added to produce the final formulas as follow in Table 1:
In the next step, the mixtures were gelatinized using water bath and autoclaved at 121 °C for 15 min Fig. 1.

Production of fermented quinoa products
2.2.1.2 Inoculant strains and quinoa fermentation conditions
The Lactobacillus bacteria strains were activated by inoculating in sterile MRS broth (9 mL) and incubation at 37 °C for 24 h. The cells were separated from the broth by centrifuging, and re-suspended in sterile saline solution (9 mL) with final concentration 108 CFU/mL [16].
The 24 h activated culture of probiotics (108 CFU/mL) were added to the previous quinoa mixtures by a concentration of 1%. A control sample without inoculation of bacteria was prepared. All treatments were incubated at 37 °C for 24 h for L. plantarum fermentation and 8 h for L. delbrueckii ssp. Bulgaricus fermentation. The fermented quinoa products were stored after fermentation at 4 ± 1 °C for 21 days. Chemical and microbiological properties of fermented quinoa products were estimated at 0, 7, 14 and 21 days.
2.2.2 Determination of titrable acidity
Total acidity was determined by titration method (10 g of fermented products with 90 mL distilled water, using 0.1 N NaOH solution and phenolphthalein as indicator) according to Coda et al. [17].
2.2.3 Determination of free phenolics and antioxidant activity
2.2.3.1 Extract preparation
Free phenolic compounds were extracted from samples using the method of Acosta-Estrada et al. [18]. Briefly, 1 g of fermented quinoa samples were mixed with 10 mL of chilled ethanol/water (80:20 v/v) and shaken at 250 rpm for 10 min at 25 °C, then centrifuged at 3000 g for 10 min. The supernatant was recovered and the extraction repeated once. Supernatants were pooled and evaporated at 50 °C and 20 mbar. The resulting extracts were stored at − 20 °C until use.
2.2.3.2 Determination of free phenolic content
Free phenolic contents were estimated in the pravious ethanolic extracts of fermented quinoa products by Folin-Ciocalteu method according to Jaramillo-Flores et al. [19]. An 100 µL of ethanolic extract was mixed with 900 µL of Folin-Ciocalteu reagent (diluted 1:10 with distilled water) and was allowed to stand for 5 min at room temperature; 0.75 mL of sodium bicarbonate solution (7%) was added to the mixture and vortexed for 30 s, and allowed to stand at room temperature for 90 min. The absorbance was measured at 725 nm using a spectrophotometer (6505 UV/Vis, Jenway LTD., Felsted, Dunmow, UK). A calibration curve of gallic acid (ranging from 0 to 1 mg/mL) was prepared and tested under similar conditions. All values were expressed as mean (mg of Gallic acid equivalents/100 g wet weight fermented quinoa).
2.2.3.3 Determination of DPPH radical scavenging activity
Free radical scavenging activity was determined by the 1,1-diphenyl-2 picrylhydrazyl (DPPH) assay according to Elfahri et al. [20] with slight modification. Briefly, 900 μL of the DPPH reagent (0.1 mM DPPH dissolved in 95% methanol) was added to 100 μL of fermented product extracts in glass test tubes. The samples were shaken vigorously and incubated for 30 min in the dark at room temperature. The absorbance of the incubated samples was measured at 517 nm. The percentage of radical scavenging activity was expressed as scavenging (%) using Eq.:
2.2.4 Determination of phytate content
The phytic acid (IP6) was determined in terms of its phosphorous content, using the method described by [21]. Phytic acid was calculated from phytate phosphorus from the weight ratio of phosphorus atoms per molecule of IP6 (1:3.52).
2.2.5 Quantification of water extractable arabinoxylans (WEAX)
The WEAX of fermented products was determined according to [22], with slight modification. Samples of fermented quinoa products (1 g) were extracted with of distilled water (20 mL) under constant agitation and centrifuged for 10 min at 4000 rpm. Then, 100 µL of supernatant, 100 µL of distilled water and 2 mL of daily prepared working reaction solution (1 g of phloroglucinol dissolved in 5 mL of pure ethanol, 2 mL of hydrochloric acid, 110 mL of glacial acetic acid and 1 mL of a 17.5 g L−1 glucose solution) were added to stoppered glass tubes (12 mL, 16 × 100 mm). The tubes were located for 25 min in a boiling water bath, next immediately cooled in the ice. The absorbance was measured using an UV–Vis spectrophotometer at 552 nm and 510 nm. D-(+)-Xylose was used as standard for the calibration curve (0.05–30 mg Kg−1). Finally, the WEAX content was calculated subtracting the absorbance value at 510 nm, which corresponds to hexose interferences, from the absorbance value at 552 nm and the obtained value was compared with the regression equation.
2.2.6 Biological experiment
2.2.6.1 Adaptation and distributing of experimental animals
The animals were housed as groups in wire cages under the normal laboratory conditions and fed on basal diets for 10 days as adaptation period. The rats were distributed to 4 groups containing control group and fed during experimental period (30 days).
2.2.6.2 Experimental design
The rats were randomly divided into 4 groups (1, 2, 3 and 4), each group was contained 10 rats. Group 1 (control): fed on basal diet, Group 2: fed on basal diet containing of 30% of non-fermented quinoa, Group 3: fed on basal diet containing of 30% of FQP2, Group 4: fed on basal diet containing of 30% of FQD2.
2.2.6.3 Blood sampling
At the day of blood sampling collection, rats were fasted overnight and anesthetized. Blood samples were collected from the retro-orbital plexus from all animals of each group into clean, dry and labeled tubes. The tubes were centrifuged at 3500 rpm for 15 min to separate the blood serum, which used to estimation of biochemical parameters [23].
2.2.6.4 Determination of fasting and postprandial blood glucose
The concentrations of fasting and postprandial blood glucose were determined according to Lopes et al. [24] using enzymatic kits (SPECTRUM diagnosis, Germany).
2.2.7 Statistical analysis
Basic statistics and analysis of variance (ANOVA) were performed to data analysis and test the significance within replications and between treatments by using IBM SPSS software version 22. Duncan test was used to determine the differences among the means at the significance level of 0.05%.
3 Results and discussions
3.1 Changes in titrable acidity of fermented quinoa products
Variations in titrable acidity % of fermented quinoa products during storage at 4 ºC ± 1 for 21 days were illustrated in Table 2. By fermentation process using L. plantaram and L. delbrueckii ssp. Bulgaricus, the titrable acidity increased from 0.27% to values ranged between 0.38 and 0.43%. The treatment of FQD2 showed the highest acidity value (0.43%) (P > 0.05) followed by FQD1 (0.42%) may be due to the high count of probiotic bacteria in these treatments, while FQP treatment presented the lowest acidity value (0.38%) after control sample. Lactate produced by lactic acid bacteria is responsible for the acidity increasing in the fermented quinoa products [25, 26]. Furthermore, the acidity values were continuously increased during storage period and the highest values were recorded at the end of storage for all treatments.
3.2 Changes in free phenolic content of fermented quinoa products
Generally, fermentation of quinoa by probiotic bacteria increased the free phenolic content of fermented quinoa products in all treatments compared to non-fermented quinoa products (Table 3). Also, the results showed that the adding of MLP at the two levels (0.25 and 0.50%) significantly enhanced the free phenolic content of fermented quinoa products more than fermented quinoa products without MLP, and the samples containing the higher level of MLP (0.50%) displayed the highest values of free phenolic content (34.24 and 32.85 for FQD2 and FQP2, respectively). Many previous studies indicated to the high antioxidant activity and phenolic compounds of MLP, this may explain the higher phenolic content of fermented quinoa samples supplemented with MLP [13,14,15].
During the first and second weeks, the free phenolic content of fermented quinoa continuously increased through the storage period may be because of the activity of probiotic bacteria. Călinoiu et al. and Hole et al. [27, 28], reported that the increase of antioxidants such as phenolic compounds after fermentation may be due to its increased release or synthesis by some possible enzymes, such as β-glucosidases, esterase, cellulose, β-glucosidases and glycoside hydrolase that produced by some probiotic strains. The production of these enzymes during fermentation could release esterified and insoluble-bound phenolcs in a time-dependent manner. These results are in line with those obtained from [29, 30], they stated that quinoa fermentation by probiotics improved the free phenolic contents and antioxidant activity.
3.3 Changes in free DPPH radical scavenging activity of fermented quinoa products
Changes in free DPPH of fermented quinoa during storage at 4 ºC ± 1 for 21 days were presented in Table 4. Significant differences were recorded in the free antioxidant activity of fermented quinoa supplemented and non-supplemented with MLP. Quinoa products contained MLP at the level of 0.50% and fermented by L. delbrueckii ssp. Bulgaricus and L. plantaram showed the highest antioxidant activity (7.75 and 6.62%, respectively) followed by fermented quinoa products contained MLP at the level of 0.25% (6.29 and 5.48%, respectively). The increase of free phenolic content results in improving the antioxidant activity of fermented products. The correlation between the phenolic content and the antioxidant activity was reported by many previous studies [27, 31]. Similar results were recorded by [29, 30], they indicated that the antioxidant activity of quinoa was increased after fermentation process by probiotics.
3.4 Changes in phytate content (mg/100 ww) of fermented quinoa products
Phytate is the primary storage form of phosphate in many cereals and it has anti-nutritional effect resulting in malabsorption of some important dietary minerals such as Fe2+ and Ca2+ as well as decreasing of protein solubility and amino acids bioavailability [32]. From the nutritional point of view, reducing of phytate levels is desirable and necessary to enhance the bioavailability of mineral and amino acids in foods [33]. Several methods have been applied to decrease the phytate concentration in foods such as germination, malting and soaking as well as fermentation, which considers the most common process, used to decrease the levels of phytate in food matrix [34].
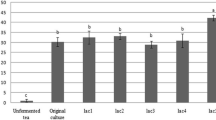
Our results indicated that, the fermentation process by probiotics resulted in significant decrease (P > 0.05) in phytate concentrations of fermented quinoa products, and fermentation by L. plantaram (FQP) presented higher activity for phytate degradation than fermentation by L. delbrueckii ssp. Bulgaricus (FQD) (Table 5). Phytate levels were recorded as 67.06 and 71.88 mg/100 g ww for FQP and FQD, respectively compared with control sample, which had the highest phytate content 94.38 mg/100 g ww. Fermented quinoa products supplemented with MLP showed lower phytate concentrations than fermented quinoa products without MLP, and fermented quinoa containing the highest level of MLP showed the lowest phytate concentrations (51.82 and 57.21 mg/100 g ww for FQP2 and FQD2, respectively). All fermented quinoa products exhibited insignificant decreases (P > 0.05) in phytate levels through storage period for 21 days at 4 ºC ± 1, and FQP2 as well as FQD2 had the lowest phytate contents (44.47 and 47.59, mg/100 g ww respectively) at the end of storage.
Phytate degradation in the fermented quinoa products may be due to the exogenous phytase activity of Lactobacillus bacteria rather than endogenous phytase activity of quinoa because of the activity of quinoa endogenous phytase may be reduced by the thermal processes before fermentation [2] as we can observe in the control sample. These results are agree with those obtained from [2], they stated that the fermentation by Lactobacillus strains degraded phytate and reduced it in the quinoa flour.
3.5 Changes in water extractable arabinoxylans (WEAX) (mg/100 g ww) of fermented quinoa products
WEAX have antioxidant activity against free radicals in gastrointestinal tract and food matrix and may help in inhibition of colorectal cancers by mitigating the toxicity of free radicals [35]. Also, Malunga and coworkers indicated to the potential antiglycemic effect of WEAX may be due to the direct inhibition of α-glucosidase enzyme activity [36].
The effect of fermentation process on the content of WEAX of fermented quinoa during storage at 4 ºC ± 1 for 21 days was shown in Table 6. The content of WEAX increased from 12.18 for control sample to values ranged between 18.42 to 29.34 mg/100 g ww for fermented quinoa products. The results indicated that quinoa products fermented by L. plantaram showed higher content of WEAX than quinoa products fermented by L. delbrueckii ssp. Bulgaricus. Fermented quinoa by L. plantaram recorded 29.34, 24.06 and 21.30 mg/100 g ww for FQP2, FQP1 and FQP, respectively, while fermented quinoa by L. delbrueckii ssp. Bulgaricus recorded 24.54, 21.96 and 18.42 mg/100 g ww for FQD2, FQD1 and FQD, respectively. Interestingly, fermented quinoa containing higher content of MLP (0.50%) had the highest contents of WEAX and displayed more positive effect on improving WEAX content than fermented quinoa containing lower content of MLP (0.25%) for both probiotic bacteria may be the enhancing effect of MLP on the growth and survival of probiotics which responsible for the quinoa fermentation and increasing of quinoa WEAX. At the end of storage period, WEAX content reached to the highest values for all fermented quinoa products and FQP2 as well as FQP1 showed the highest contents as shown in (Table 6).
These results were in agreement with [33, 37], they stated that the content of WEAX was increased in fermented cereals after fermentation by lactic acid bacteria. Acidification and specific enzymes, such as xylan-degrading enzymes can hydrolyse the backbone of high molecular weight arabinoxylans and enhance its solubilization [33, 38].
3.6 Determination of fasting (FG) and postprandial (PP) blood glucose
Type 2 diabetes mellitus (T2DM) is one of the most prevalent chronic diseases around the world. It is be characterized by high level of blood glucose, resistance to insulin and relative insulin deficiency, accompanying with multiple systemic complications. According to International Diabetes Federation (IDF), T2DM has affected 415 million adults, and this number can increase to more than 600 million up to 2040 [39]. Data showed in Table 7 illustrated the effect of feeding with fermented quinoa products by probiotics on fasting and postprandial blood glucose levels of experimental rats. The results indicated that the feeding with fermented quinoa products by probiotics significantly decreased (P > 0.05) fasting blood glucose levels. Group 3 (Fed with FQP2) showed the highest decrement of fasting blood glucose level (10.37%). Moreover, group 2 (fed with non-fermented quinoa) and 3 (Fed with FQD2) exhibited insignificant decrease (P > 0.05) in the fasting blood glucose levels. Also, the results indicated that groups 3 and 4 presented higher decrement (P > 0.05) in postprandial glucose levels (13.82 and 10.85%, respectively) than group 2 (7.78%). The higher activity of reducing blood glucose which was recorded in the groups 3 and 4 may be due to the presence of probiotics and prebiotic effect of MLP. These results are in line with [40, 41], they reported that probiotics showed reducing blood glucose activity in bath in vitro and in vivo studies.
4 Conclusion
Symbiotic foods based on quinoa fermented by probiotics were produced as a significant source of bioactive compounds and alive probiotics for the human body. Moringa leaves powder (MLP) was added at the concentrations of 0.25 and 0.50% as additional prebiotic source to quinoa fermented by Lactobacillus plantaram ATCC 14917 and Lactobacillus delbrueckii ssp. Bulgaricus EMCC 11102. The results indicated that supplementation of fermented quinoa products with MLP at bath levels increased its contents of free phenolics and water extractable arabinoxylans as well as enhanced its antioxidant activity and phytate degradation. Fermented quinoa products with 0.50% MLP showed better chemical properties than fermented quinoa products with 0.25% MLP with positive effect on the blood glucose level of experimental rats during feeding period. Supplementation of fermented quinoa products with MLP resulted in improving its nutritional value and healthy benefits. More studies are needed in the future to test more kinds of probiotics with different concentration of MLP in quinoa fermentation.
Data availability
All data generated or analysed during this study are included in this article.
Code availability
Not applicable.
References
Bazile D, Jacobsen S, Verniau A. The global expansion of quinoa: trends and limits. Front Plant Sci. 2016;7:622.
Castro-Alba V, Lazarte CE, Perez-Rea D, Sandberg AS, Carlsson NG, Almgren A, Bergenståhl B. Effect of fermentation and dry roasting on the nutritional quality and sensory attributes of quinoa. Food Sci Nutr. 2019;7(12):3902–11.
Obaroakpo JU, Liu L, Zhang S, Lu J, Pang X, Lv J. α-Glucosidase and ACE dual inhibitory protein hydrolysates and peptide fractions of sprouted quinoa yoghurt beverages inoculated with Lactobacillus casei. Food Chem. 2019;299:124985.
Mäkinen OE, Wanhalinna V, Zannini E, Arendt EK. Foods for special dietary needs: non-dairy plant-based milk substitutes and fermented dairy-type products. Crit Rev Food Sci Nutr. 2016;56(3):339–49.
Singhal S, Baker RD, Baker SS. A comparison of the nutritional value of cow’s milk and nondairy beverages. J Pediatr Gastroenterol Nutr. 2017;64(5):799–805.
Ludena Urquizo FE, García Torres SM, Tolonen T, Jaakkola M, Pena-Niebuhr MG, von Wright A, Repo-Carrasco-Valencia R, Korhonen H, Plumed-Ferrer C. Development of a fermented quinoa-based beverage. Food Sci Nutr. 2017;5(3):602–8.
Paz PC, Janny RJ, Håkansson ÅJ. Safeguarding of quinoa beverage production by fermentation with Lactobacillus plantarum DSM 9843. Int J Food Microbiol. 2020;324:108630.
Canaviri-Paz P, Oscarsson E, Håkansson ÅJ. Autochthonous microorganisms of white quinoa grains with special attention to novel functional properties of Lactobacilli strains. J Funct Foods. 2021;84:104586.
Abu Hafsa SH, Ibrahim SA, Eid YZ, Hassan AA. Effect of dietary Moringa oleifera leaves on the performance, ileal microbiota and antioxidative status of broiler chickens. J Anim Physiol Anim Nutr (Berl). 2020;104(2):529–38.
Umar S, Mohammed Z, Nuhu A, Musa K, Tanko Y. Evaluation of hypoglycaemic and antioxidant activity of moringa oleifera root in normal and alloxan-induced diabetic rats. Trop J Nat Prod Res. 2018;2(8):401–8.
Saucedo-Pompa S, Torres-Castillo JA, Castro-López C, Rojas R, Sánchez-Alejo EJ, Ngangyo-Heya M, Martínez-Ávila G. Moringa plants: bioactive compounds and promising applications in food products. Food Res Int. 2018;111:438–50.
Cao P, Wu L, Wu Z, Pan D, Zeng X, Guo Y, Lian L. Effects of oligosaccharides on the fermentation properties of Lactobacillus plantarum. J Dairy Sci. 2019;102(4):2863–72.
Hassan MA, Xu T, Tian Y, Zhong Y, Ali FAZ, Yang X, Lu B. Health benefits and phenolic compounds of moringa oleifera leaves: a comprehensive review. Phytomedicine. 2021;93:153771.
El-Hadary AE, Ramadan MF. Antioxidant traits and protective impact of Moringa oleifera leaf extract against diclofenac sodium-induced liver toxicity in rats. J Food Biochem. 2019;43(2):e12704.
Prabakaran M, Kim SH, Sasireka A, Chandrasekaran M, Chung IM. Polyphenol composition and antimicrobial activity of various solvent extracts from different plant parts of Moringa oleifera. Food Biosci. 2018;26:23–9.
Ujiroghene OJ, Liu L, Zhang S, Lu J, Zhang C, Pang X, Lv J. Potent α-amylase inhibitory activity of sprouted quinoa-based yoghurt beverages fermented with selected anti-diabetic strains of lactic acid bacteria. RSC Adv. 2019;9(17):9486–93.
Coda R, Rizzello CG, Trani A, Gobbetti M. Manufacture and characterization of functional emmer beverages fermented by selected lactic acid bacteria. Food Microbiol. 2011;28(3):526–36.
Acosta-Estrada BA, Serna-Saldívar SO, Gutiérrez-Uribe JA. Chemopreventive effects of feruloyl putrescines from wastewater (nejayote) of lime-cooked white maize (Zea mays). J Cereal Sci. 2015;64:23–8.
Jaramillo-Flores ME, González-Cruz L, Cornejo-Mazón M, Dorantes-Alvarez L, Gutiérrez-López GF, Hernández-Sánchez H. Effect of thermal treatment on the antioxidant activity and content of carotenoids and phenolic compounds of cactus pear cladodes (Opuntia ficus-indica). Food Sci Technol Int. 2003;9(4):271–8.
Elfahri KR, Vasiljevic T, Yeager T, Donkor ON. Anti-colon cancer and antioxidant activities of bovine skim milk fermented by selected Lactobacillus helveticus strains. J Dairy Sci. 2016;99(1):31–40.
Kent-Jones D, Amos AJ. Modern cereal chemistry. Liverpool: Northern Pub; 1957.
Kiszonas AM, Courtin CM, Morris CFJCc. A critical assessment of the quantification of wheat grain arabinoxylans using a phloroglucinol colorimetric assay. Cereal chem. 2012;89(3):143–50.
Ilwy Y, The effect of some kinds of sea food (fish) on blood lipid profile in rats. (Doctoral dissertation, Ph. D. Thesis, faculty of specific education, Ain Shams University; 2003.
Lopes C, Barcelos M, Vieira C, de Abreu W, Ferreira EB, Pereira RC, de Angelis-Pereira MC. Effects of sprouted and fermented quinoa (Chenopodium quinoa) on glycemic index of diet and biochemical parameters of blood of Wistar rats fed high carbohydrate diet. J Food Sci Technol. 2019;56(1):40–8.
Gallo M, Nigro F, Passannanti F, Salameh D, Schiattarella P, Budelli A, Nigro R. Lactic fermentation of cereal flour: feasibility tests on rice, oat and wheat. Appl Food Biotechnol. 2019;6(3):165–72.
Chen L, Wu D, Schlundt J, Conway PL. Development of a dairy-free fermented oat-based beverage with enhanced probiotic and bioactive properties. Front Microbiol. 2020;11:609734.
Călinoiu LF, Cătoi AF, Vodnar DC. Solid-state yeast fermented wheat and oat bran as a route for delivery of antioxidants. Antioxidants. 2019;8(9):372.
Hole AS, Rud I, Grimmer S, Sigl S, Narvhus J, Sahlstrøm S. Improved bioavailability of dietary phenolic acids in whole grain barley and oat groat following fermentation with probiotic Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus reuteri. J Agric Food Chem. 2012;60(25):6369–75.
Ayyash M, Johnson SK, Liu SQ, Al-Mheiri A, Abushelaibi A. Cytotoxicity, antihypertensive, antidiabetic and antioxidant activities of solid-state fermented lupin, quinoa and wheat by Bifidobacterium species: in-vitro investigations. LWT. 2018;95:295–302.
Li S, Chen C, Ji Y, Lin J, Chen X, Qi B. Improvement of nutritional value, bioactivity and volatile constituents of quinoa seeds by fermentation with Lactobacillus casei. J Cereal Sci. 2018;84:83–9.
Bei Q, Wu Z, Chen G. Dynamic changes in the phenolic composition and antioxidant activity of oats during simultaneous hydrolysis and fermentation. Food Chem. 2020;305:125269.
Carrizo SL, de Oca CEM, Laiño JE, Suarez NE, Vignolo G, LeBlanc JG, Rollán GJ. Ancestral Andean grain quinoa as source of lactic acid bacteria capable to degrade phytate and produce B-group vitamins. Food Res Int. 2016;89:488–94.
Spaggiari M, Ricci A, Calani L, Bresciani L, Neviani E, Dall’Asta C, Lazzi C, Galaverna G. Solid state lactic acid fermentation: a strategy to improve wheat bran functionality. LWT. 2020;118:108668.
Kumar V, Sinha AK, Makkar HP, Becker KJ. Dietary roles of phytate and phytase in human nutrition: a review. Food Chem. 2010;120(4):945–59.
Malunga LN, Beta TJ. Antioxidant capacity of water-extractable arabinoxylan from commercial barley, wheat, and wheat fractions. Cereal Chem. 2015;92(1):29–36.
Malunga LN, Izydorczyk MJ. Antiglycemic effect of water extractable arabinoxylan from wheat aleurone and bran. J Nutr Metabolism. 2017;2017:1–6.
Zhao HM, Guo XN, Zhu KX. Impact of solid state fermentation on nutritional, physical and flavor properties of wheat bran. Food Chem. 2017;217:28–36.
Hansen HB, Andreasen M, Nielsen M, Larsen L, Knudsen BK, Meyer A, Christensen L, Hansen ÅJ. Changes in dietary fibre, phenolic acids and activity of endogenous enzymes during rye bread-making. Eur Food Res Technol. 2002;214(1):33–42.
Bunyan G, practice C. World Diabetes Congress 2015: living with diabetes stream: triumphing over complexity and flourishing with diabetes. Diabetes Res Clin Pract. 2015;108(3):514–5.
Muganga L, Liu X, Tian F, Zhao J, Zhang H, Chen W. Screening for lactic acid bacteria based on antihyperglycaemic and probiotic potential and application in synbiotic set yoghurt. J Funct Foods. 2015;16:125–36.
Hu Y, Zhou F, Yuan Y, Xu Y. Effects of probiotics supplement in patients with type 2 diabetes mellitus: a meta-analysis of randomized trials. Med Clín. 2017;148(8):362–70.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
AMA: conceptualization, data curation, formal analysis, investigation, methodology, resources, software, visualization and roles/writing—original draft. EAEl-N: supervision and validation. MNK: supervision, validation and writing—review and editing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
No competing interest exists in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abdelshafy, A.M., El-Naggar, E.A. & Kenawi, M.N. Enhancing of nutritional properties of quinoa fermented by probiotics. Discov Food 2, 21 (2022). https://doi.org/10.1007/s44187-022-00022-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s44187-022-00022-8