
Abstract
Biomolecular condensates assembled through phase transitions regulate diverse aspects of plant growth, development, and stress responses. How biomolecular condensates control plant immunity is poorly understood. In Nature Plants, a new study (Zhou et al., Nat Plants 9:289–301, 2023) reveals how plants assemble translational condensates to balance tissue health and disease resistance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
In electrical devices, a rheostat controls the intensity of current and ensures proper circuit functioning by adjusting its resistance. In biological systems, similar mechanisms exist to control the level of gene products and the strength of signaling output, which, if overproduced, can compromise an organism's general fitness. One of the key mechanisms that cells employ to regulate gene expression is translational reprogramming (Spriggs et al. 2010). This process allows cells to rapidly adjust the translation of specific mRNAs in response to changing environmental conditions, developmental cues, or cellular stresses. In a recent article in Nature Plants, Zhou et al. (Zhou et al. 2023) have begun to unravel the molecular mechanisms of this fundamental layer of immune control in the model organism Arabidopsis thaliana (Fig. 1).

ETI-induced HEM1 condensates sequester translational components to inhibit immune gene translation and to promote cell survival. During effector triggered immunity (ETI), plants massively reprogram the translatome to produce immune gene products to restrict pathogen growth through programmed cell death. Zhou et al. found that ETI activation induces HEM1, a low complexity domain-containing protein, to form cytoplasmic biomolecular condensates through biological phase separation that sequester translation factors to promote cell survival and tissue health. This enables plants to fine-tune growth-defense trade-offs upon infectious diseases
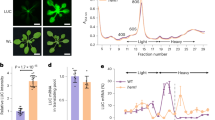
To identify the bona fide regulators of global translational reprograming in plant defense, Zhou and colleagues enlisted a genetic screen employing a translationally controlled luciferase reporter system (Xu et al. 2017) to screen for mutants with enhanced luciferase activity, which served as a proxy for immune activation. The authors delineated functions of the HEM1 gene uncovered from the screen in immune responses and translational control. Pathogen infection assays revealed that HEM1 predominantly contributed to effector-triggered immunity (ETI) but not pattern-triggered immunity (PTI) (Jones and Dangl 2006). Remarkably, ETI-induced hypersensitive response (HR) cell death was greatly accelerated in hem1 loss-of-function mutant. Hence, HEM1 served as a negative regulator of ETI pathway.
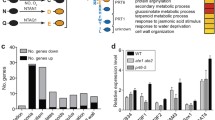
How does HEM1 repress ETI? Earlier work proposed a role for HEM1 in actin nucleation in both plants and animals (Hummel et al. 2000; Brembu et al. 2004). Zhou et al. found that HEM1 has a novel role in regulating translation efficiency. Ribosome footprinting (Ribo-seq) uncovered a global defect in translational reprogramming during immune activation. Closer inspection of Ribo-seq profiles revealed that the translation of pro-death genes was inhibited during ETI. These pro-death genes, when overproduced, cause accelerated cell death as seen for certain autoimmune mutants (van Wersch et al. 2016) and thus are under tight negative regulation in the absence of pathogen infections. To probe the molecular basis for suppression of ETI by HEM1, Zhou and colleagues enlisted protein interaction network mapping and in silico analyses. HEM1 was found to interact extensively with components of the translation machinery. Surprisingly, a plant-specific low-complexity domain (LCD) was detected at the C-terminus of HEM1. Truncation and mutagenesis assays further supported the functional role of the LCD in mediating the interaction of HEM1 with the translation machinery and repressing cell death during ETI.
Recent studies suggest that LCD-mediated liquid–liquid phase separation (LLPS) contributes to the formation of biomolecular condensates (Molliex et al. 2015), which are membrane-less assemblies that can concentrate proteins and nucleic acids to regulate diverse cellular processes (Banani et al. 2017). Zhou and colleagues tested this possibility. Biochemically, recombinant HEM1 protein underwent robust phase separation in vitro due to multivalent interactions mediated by its LCD. In planta, HEM1 cytoplasmic condensates were largely induced by ETI activation as revealed by live cell imaging. The authors further isolated HEM1 condensates from live plants and showed that they sequestered numerous translation factors, likely limiting their availability for the translation of pro-death genes. Phase separation-deficient HEM1 mutants failed to control ETI-induced cell death and resulted in altered translation. Clearly, HEM1 condensates are functionally important in promoting cell survival during infections. Future research should address the molecular mechanism of selectivity in client components and potential RNA involvement of HEM1 condensates during plant defense.
So, what can we learn from this study? First, it reinforces the emerging roles of biomolecular condensates in plant organismal defense (Zavaliev et al. 2020; Huang et al. 2021; Kim et al. 2022). The discovery of a condensation domain within HEM1 opens up new avenues for future research into the mechanisms underlying translational condensate formation and function in plants. Second, it provides new insights into the regulation of immune gene translation in eukaryotes. As biomolecular condensates play a critical role in a wide range of biological processes in other organisms, including neurodegenerative diseases in humans, these findings could have valuable implications beyond plant biology. Third, the discovery of a cell death suppression mechanism at the translational level will also challenge some common beliefs of the roles and regulation of death-promoting genes in disease development. The suppression of cell death may appear to benefit pathogen growth initially, but over time, it could reduce the selection pressure that drives the host–pathogen arms race in evolution.
In summary, Zhou and colleagues’ discovery of a phase separation control mechanism of gene translation provides a major conceptual advance in understanding the complex plant immune system. Similar to a circuit rheostat, plants appear to fine-tune the intensity of immune signaling through HEM1 phase separation to balance cell survival and death. Future research using synthetic approaches to harness HEM1’s phase behavior could have practical applications in both agriculture and human health.
Availability of data and materials
Not applicable.
References
Banani SF, Lee HO, Hyman AA, Rosen MK (2017) Biomolecular condensates: organizers of cellular biochemistry. Nat Rev Mol Cell Biol 18:285–298. https://doi.org/10.1038/nrm.2017.7
Brembu T, Winge P, Seem M, Bones AM (2004) NAPP and PIRP encode subunits of a putative wave regulatory protein complex involved in plant cell morphogenesis. Plant Cell 16:2335–2349. https://doi.org/10.1105/tpc.104.023739
Huang S, Zhu S, Kumar P, MacMicking JD (2021) A phase-separated nuclear GBPL circuit controls immunity in plants. Nature 594:424–429. https://doi.org/10.1038/s41586-021-03572-6
Hummel T, Leifker K, Klambt C (2000) The Drosophila HEM-2/NAP1 homolog KETTE controls axonal pathfinding and cytoskeletal organization. Genes Dev 14:863–873. https://doi.org/10.1101/gad.14.7.863
Jones JD, Dangl JL (2006) The plant immune system. Nature 444:323–329. https://doi.org/10.1038/nature05286
Kim JH, Castroverde CDM, Huang S, Li C, Hilleary R, Seroka A, Sohrabi R, Medina-Yerena D, Huot B, Wang J, Nomura K, Marr SK, Wildermuth MC, Chen T, MacMicking JD, He SY (2022) Increasing the resilience of plant immunity to a warming climate. Nature 607:339–344. https://doi.org/10.1038/s41586-022-04902-y
Molliex A, Temirov J, Lee J, Coughlin M, Kanagaraj AP, Kim HJ, Mittag T, Taylor JP (2015) Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell 163:123–133. https://doi.org/10.1016/j.cell.2015.09.015
Spriggs KA, Bushell M, Willis AE (2010) Translational regulation of gene expression during conditions of cell stress. Mol Cell 40:228–237. https://doi.org/10.1016/j.molcel.2010.09.028
van Wersch R, Li X, Zhang YL (2016) Mighty dwarfs: Arabidopsis autoimmune mutants and their usages in genetic dissection of plant immunity. Front Plant Sci 7:1717. https://doi.org/10.3389/fpls.2016.01717
Xu GY, Uan MY, Ai CR, Liu LJ, Zhuang E, Karapetyan S, Wang S, Dong XN (2017) uORF-mediated translation allows engineered plant disease resistance without fitness costs. Nature 545(7655):491–494. https://doi.org/10.1038/nature22372
Zavaliev R, Mohan R, Chen T, Dong X (2020) Formation of NPR1 condensates promotes cell survival during the plant immune response. Cell 182(1093–1108):e1018. https://doi.org/10.1016/j.cell.2020.07.016
Zhou Y, Niu R, Tang Z, Mou R, Wang Z, Zhu S, Yang H, Ding P, Xu G (2023) Plant HEM1 specifies a condensation domain to control immune gene translation. Nat Plants 9:289–301. https://doi.org/10.1038/s41477-023-01355-7
Acknowledgements
Not applicable.
Funding
S.H. is supported by start-up funds from The Ohio State University and JobsOhio.
Author information
Authors and Affiliations
Contributions
S.H. wrote the paper. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
No competing interests declared.
Additional information
Handling editor: Jin-Rong Xu.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Huang, S. Tuning the rheostat of immune gene translation. Stress Biology 3, 7 (2023). https://doi.org/10.1007/s44154-023-00087-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s44154-023-00087-0