
Abstract
Surfactants represent a large group of industrial relevant substances and can be generated chemically, but also biologically. Important microbial producers are several Pseudomonas species, mainly known for their ability to produce rhamnolipids. For safety reasons, recombinant Pseudomonas putida is preferred for heterologous production of rhamnolipids as well as its precursor 3-(3-hydroxyalkanoyloxy) alkanoic acid (HAA), which is a promising platform substance. The surface-active properties of HAA combined with classical submerged aeration cause strong foam formation making biotechnological production in stirred tank bioreactors challenging. Therefore, a foam-free bioprocess for HAA production was developed in this study. The combination of headspace aeration, overpressure up to 8 bar, and temperature reduction prevented oxygen limitation, shortened process time, and enhanced the maximum HAA concentration to 1.71 g l− 1 with a space-time yield of 0.08 g l− 1 h− 1.
Graphical Abstract

Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
Biobased and biodegradable materials attracted strong interest in recent years since environmental pollution increases steadily and petrochemical resources are limited [1,2,3]. Biosurfactants represent a group of surface-active substances produced by microorganisms. Among them, rhamnolipids are one example of glycolipidic biosurfactants [4] and were firstly found in Pseudomonas aeruginosa (P. aeruginosa) [5, 6]. They usually consist of one or two rhamnose units connected via β-glycosidic bond to one or two β-hydroxy fatty acids of various lengths [6, 7]. The dimer of two activated β-hydroxy fatty acids, which originate from de novo fatty acid synthesis, linked by the acyltransferase RhlA is called 3-(3-hydroxyalkanoyloxy) alkanoic acid (HAA) [8, 9]. Typically, different congeners of HAA, varying in amount and length of fatty acids, are secreted by an organism [10]. Even without the hydrophilic rhamnose part, HAA shows surface-active properties due to their ester, carboxyl, and hydroxyl groups [8, 11]. Besides the potential application as biosurfactants [11], HAA presents a suitable platform substance for biofuels and biobased plastics production [12, 13].
Recently, Tiso et al. engineered the non-pathogenic Pseudomonas putida (P. putida) strain to produce HAA [11]. Since the production host is an obligate aerobic organism, the surface-active nature of the product promotes strong foam formation during submerged aeration [14, 15]. The foam can enter and block the exhaust gas filters leading to undesired pressure increases and the foam layer can delay the dosage of acid, base or feeding solutions into the culture broth. Moreover, space-time yield of the process is reduced, because biomass is transferred out of the culture broth by attaching to the foam with the hydrophobic cell surface structures [16,17,18].
The conventional use of antifoaming agents has several drawbacks. Depending on the antifoaming agent, the oxygen transfer can be negatively affected, downstream processing can become problematic, and process costs increase [19,20,21]. As the industrial application of such biosurfactants has become increasingly interesting, various strategies have been developed in recent years to address foaming and related problems more advantageously. From a biological point of view, cell surface modification was developed to avoid cell attachment to the foam [22], and switching to anaerobic mode was shown to realize a foam-free bioprocess [23]. Process engineering methods like the application of in situ liquid-liquid extraction reduced foaming by product removal into an organic phase [24]. In several studies, foam fractionation strategies were applied with the additional benefit of an in situ product removal [22, 25,26,27,28]. On the technical side, membrane aerators facilitate bubble-free aeration [14, 29, 30]. Kuo and Lanser applied partial headspace aeration in P. putida bioprocess to avoid excessive foaming [31]. However, in comparison to the gas-liquid surface provided by air bubbles of submerged aeration, headspace aeration reduces the gas exchange area. Therefore, oxygen transfer to the liquid phase is tremendously reduced.

Application of headspace aeration, overpressure, and temperature reduction for the foam-free production of HAA by Pseudomonas putida
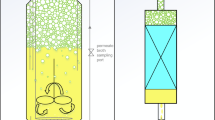
To overcome gas transfer limitations, overpressure can be applied to increase gas solubility [32, 33] and was used in several studies to enhance the oxygen supply [34,35,36,37,38,39,40]. The scalability of pressurized cultivations was discussed in detail by Knoll et al. and Kreyenschulte et al. [41, 42]. In this study, the combination of headspace aeration and overpressure was utilized to overcome the contrary bioprocess requirements originating from a strongly foaming product and a highly oxygen-demanding production host (Fig. 1). Additionally, cultivation temperature was reduced to slow down the microbial metabolic activity to prevent oxygen limitations. The developed bioprocess provides an alternative strategy for scalable production of HAA with little technical effort.
2 Methods and materials
2.1 Microbial strain
Recombinant Pseudomonas putida KT2440 KS3 (attTn7::Pffg-rhlA) [22] was used for heterologous HAA production. The cryogenic cultures were stored at − 80 °C.
2.2 Shaken precultures and medium composition
For the first preculture, a 500 ml unbaffled Erlenmeyer flask containing 50 ml Luria-Bertani (LB) medium was inoculated with 1 ml cryogenic culture. The preculture was incubated at 30 °C and 300 rpm with a 50 mm shaking diameter (ISF1-X, Adolf Kuhner AG, Birsfelden, Switzerland) until a minimal optical density at 600 nm (OD600) of 2 was reached during the exponential growth phase. Yeast extract SERVABACTER® and tryptone/peptone ex casein for LB medium were supplied by SERVA Electrophoresis GmbH (Heidelberg, Germany) and Carl Roth GmbH & Co. KG (Karlsruhe, Germany). For the second preculture, a mineral salt medium (MSM) according to Hartmans et al. [43] with a modified phosphate buffer of Blesken et al. [22] was used and contained: 20 g l− 1 glucose, 11.64 g l− 1 K2HPO4, 4.89 g l− 1 NaH2PO4, 2.00 g l− 1 (NH4)2SO4, 10 mg l− 1 EDTA, 100 mg l− 1 MgCl2 × 6 H2O, 2 mg l− 1 ZnSO4 × 7 H2O, 1 mg l− 1 CaCl2 × 2 H2O, 5 mg l− 1 FeSO4 × 7 H2O, 0.2 mg l− 1 Na2MoO4 × 2 H2O, 0.2 mg l− 1 CuSO4 × 5 H2O, 0.4 mg l− 1 CoCl2 × 6 H2O and 1 mg l− 1 MnCl2 × 2 H2O. The second preculture was cultivated at pH 7.00 in 1 l unbaffled Erlenmeyer flasks with 50 ml MSM and inoculated to a start OD600 of 0.1. Cultures were incubated at 300 rpm with a 50 mm shaking diameter (ISF1-X, Adolf Kuhner AG, Birsfelden, Switzerland) until a minimal OD600 of 6 was reached. In difference to the first preculture, the incubation temperature was decreased to 28 °C to delay growth (Supplementary Information, Fig. S2) and temporally enable harvest in late exponential growth phase.
2.3 Cultivations and online measurements in stirred tank bioreactors
Cultivations in stirred tank bioreactors were conducted in batch mode using MSM with reduced phosphate buffer concentration of 3.88 g l− 1 K2HPO4 and 1.63 g l− 1 NaH2PO4 according to Blesken et al. [22]. The pH was maintained at 7.00 ± 0.05 with NH4OH. The bioreactors were inoculated with an initial OD600 of 0.2. For cultivations with an overpressure ≤ 0.9 bar, a 30 l baffled stainless steel bioreactor (BIOSTAT® D-DCU, Sartorius AG, Goettingen, Germany) equipped with a pH-probe (EasyFerm® Plus, Hamilton Bonaduz AG, Bonaduz, Switzerland), a dissolved oxygen (DO) probe (OxyFerm® FDA, Hamilton Bonaduz AG, Bonaduz, Switzerland) installed within the lower half of the culture broth and two six-bladed Rushton turbines was used. The filling volume was 15 l and aeration was realized only via headspace with 20 l min− 1. The stirring rate was kept constant at 500 rpm. The cultivation temperature was set to 23 or 30 °C as stated in the result section. For pressurized mode, the cultivation was conducted in a 7.5 l baffled pressure stirred tank bioreactor (Proreact 3P, Heinrich Frings GmbH & Co. KG, Rheinbach, Germany) with a filling volume of 4.5 l. The bioreactor was equipped with probes for pH (Polilyte® Plus H, Hamilton Bonaduz AG, Bonaduz, Switzerland), DO (VisiFerm® mA, Hamilton Bonaduz AG, Bonaduz, Switzerland) and dissolved carbon dioxide (CO2NTROL, Hamilton Bonaduz AG, Bonaduz, Switzerland), both installed within the lower third of the culture broth, as well as with two six-bladed Rushton turbines. The dissolved oxygen was manually controlled to > 0% by increasing the stirring rate (400–850 rpm), decreasing cultivation temperature (30−23 °C), and increasing the overpressure (7.5–8.0 bar). The headspace aeration was initially set to 4 l min− 1 and adjusted to 12 l min− 1 after 6.4 h to ensure functional pressure control. Continuous gas analyzers (Rosemount™ X-STREAM Enhanced XEGK Continuous Gas Analyzer, Emerson Electric Co., Saint Louis Missouri, USA or BioPAT® Xgas, Sartorius AG, Goettingen, Germany) were connected to the exhaust gas line to monitor off-gas CO2 and O2. All cultivations were performed under turbulent flow conditions (Reynolds number > > 10,000). To ensure that no foaming occurred, it was visually monitored using the sight glasses of the reactors. In addition, foam probes (conductivity measurement) were installed in the headspace of all reactors.
2.4 Shake flask experiments
Cultivations were conducted in 250 ml unbaffled shake flasks with 4% filling volume. The cultures were inoculated to an initial OD600 of 0.4 and incubated at 30 °C and 300 rpm with 50 mm shaking diameter (ISF1-X, Adolf Kuhner AG, Birsfelden, Switzerland). The incubator was equipped with the Kuhner TOM system (Adolf Kuhner AG, Birsfelden, Switzerland) for online exhaust gas analysis and determination of oxygen transfer rate (OTR) and carbon dioxide transfer rate (CTR). The cultures were aerated with 20 ml min−1 standard compressed air or a synthetic gas mixture (10% CO2, 25% O2, 65% N2). One measuring cycle of 10 min included 3 min measuring time.
2.5 Offline analytics and data analysis
The OD600 was measured by an UV/Vis spectrophotometer V-630 (Jasco Deutschland GmbH, Pfungstadt, Germany). For the determination of cell dry weight (CDW), 10 ml of sample was centrifuged for 15 min at 10,000 g and 4 °C (Sigma 3–18 K Centrifuge, Sigma Laborzentrifugen GmbH, Osterode am Harz, Germany). The pellet was dried and weighted using a MA160 moisture analyzer (Sartorius AG, Goettingen, Germany). The glucose concentration was determined in the supernatant using a YSI 2950 Biochemistry Analyzer (YSI Inc., Yellow Springs, USA). The CTR was calculated according to Peña-Ortiz et al. from exhaust gas data [44]. Calculation of yields was based on HAA maxima. HAA was quantified according to a method developed by Behrens et al. and Tiso et al. [45, 46]. For HAA analytics, the culture filtrate was diluted 1:2 with acetonitrile and centrifuged at 4 °C and 15,000×g for 10 min (Prism™ R Refrigerated MicroCentrifuge, Labnet International Inc., Edison, USA). HAA was analyzed by reversed-phase chromatography in a HPLC system (JASCO International Co., Ltd., Tokyo, Japan) equipped with a YMC-Pack Pro C18 column (150 × 4.6 mm, 3 μm, YMC Europe GmbH, Dinslaken, Germany) and an evaporative light scattering detector (ELSD) (evaporator 80 °C, nebulizer 30 °C, LED 50%, gas flow 1 slm). The mobile phase was formed with 0.2% formic acid in water (A) and acetonitrile (B). The flow was set to 1 ml min− 1 and the oven temperature was 35 °C. Initially, 70% B was held for 1 min, afterwards, B was increased to 80% until 8 min. From 9 to 10 min, B was linearly increased to 100% and from 15 to 18 min, B was reduced to 70% again. The injection volume was 20 µl.
3 Results and discussion
3.1 Impact of headspace aeration and temperature decrease on HAA formation
Foam formation, caused by HAA production, is a consequence of submerged aeration in conventional stirred tank reactor cultivations. Switching to headspace aeration provides a simple technical solution but also reduces gas transfer area tremendously. Thus, the oxygen supply of P. putida as well as the removal of the generated CO2 are limited. Therefore, the impact of headspace aeration on HAA formation was investigated (Fig. 2a, b). P. putida was oxygen-limited from 2 h until the end of the cultivation, indicated by a dissolved oxygen tension (DOT) value of 0% (Fig. 2a).

Batch culture of P. putida with headspace aeration in 30 l stirred tank reactor a and b—cultivation at 30 °C; c and d—cultivation at 23 °C; a and c—online data for CTR and DOT; b and d—offline data of glucose concentration, CDW and HAA concentrations; Experimental conditions: 15 l filling volume; 500 rpm; 20 l min− 1 gas flow rate via headspace; pH 7.00 ± 0.05 (25% (v/v) NH4OH); 0.9 bar overpressure
The maximum achieved gas transfer rates were limited to 7 mmol l− 1 h− 1 (Supplementary Information, Fig. S1), which is approximately 20 times lower than the maximum gas transfer rates of this reactor system with submerged aeration. The process was ended after 32 h when cell dry weight and HAA concentration started to decrease (Fig. 2b). In accordance with the observations of Demling et al. biomass grew linearly during the oxygen-limited bioprocess [47] and the glucose concentration also showed a linear decrease. Interestingly, biomass growth stopped although 5.5 g l− 1 of glucose was still available. Similar behavior was observed for HAA formation. HAA concentration increased almost linearly and reached a maximum of 0.58 g l− 1 between 25 and 29 h (Fig. 2b). When the biomass decreased, no further HAA formation could be observed, which suggests a growth-related HAA production. Besides the limited HAA formation, the product concentration decreased between 27 and 32 h, which is a critical issue. One explanation could be the uptake and cleavage of HAA by P. putida [48] and degradation of the 3-hydroxyalkanoates via β-oxidation to form acetyl-CoA. This can be metabolized via the tricarboxylic acid cycle for ATP generation since decreased intracellular ATP levels were observed under oxygen starvation [47]. This degradation pathway was already found for intracellular polyhydroxyalkanoates (PHAs) [49] that share the same precursor substances as for HAA biosynthesis in P. aeruginosa [50].
The reduction of the cultivation temperature is a conventional measure to decelerate the metabolic activity and consequently provide the necessary oxygen transfer rate to avoid an oxygen limitation of the microbial system. To improve oxygen availability in the culture, the impact of a temperature reduction by 7 °C, from 30 to 23 °C, on HAA formation was investigated (Fig. 2c, d). The beginning of the CTR plateau (Fig. 2c), reflecting the metabolic activity, was delayed by approx. 7 h compared to the previous experiment shown in Fig. 2a. Besides the consequence of an extended process time by 4 h, only positive effects were observed from temperature reduction. The HAA concentration was doubled and the entire glucose was converted before product degradation (Fig. 2d). The maximum HAA concentration of 1.28 g l− 1 was achieved at 34 h. In comparison to the cultivation at 30 °C, the hypothesized ATP shortage by oxygen limitation should be reduced at 23 °C resulting in a higher conversion of glucose into HAA. Interestingly, the rates for glucose consumption, biomass formation and HAA formation were higher under oxygen-limited conditions at 23 °C in comparison to 30 °C (Table 1). A possible explanation could be the elevated oxygen solubility at lower temperatures.
Another suitable process parameter that could result in similar effects as observed for the temperature could be the pH value. By lowering the pH to 6.0, Demling et al. observed a reduced growth rate during rhamnolipid production with P. putida [24]. For the presented bioprocess for HAA production, shifting pH would be an additional possibility to reduce the negative impact of an oxygen limitation and enhance HAA formation.
3.2 Effect of overpressure up to 8 bar on HAA formation
Since headspace aeration tremendously reduces the oxygen transfer rate, an increase of the headspace pressure is an option to overcome this limited oxygen supply. The elevated pressure increases gas solubility (oxygen and carbon dioxide) linearly. Therefore, it was aimed to apply overpressure to the HAA-producing bioprocess to demonstrate a positive effect of an improved oxygen supply on product formation. Furthermore, the impact of the CO2 accumulated in the liquid phase on metabolic activity was observed.
The cultivation was conducted with an overpressure between 7.5 and 8.0 bar (Fig. 3a). Since the experiment aimed to increase the final HAA titre and to maximize the space-time yield (STY), the cultivation was started at 30 °C. The temperature was gradually reduced to 23 °C (Fig. 3a) after DO had fallen below 30% at 15 h (Fig. 3b) to reduce metabolic activity and consequently oxygen demand.To ensure the functionality of the pressure control, the headspace aeration had to be increased from 4 to 12 l min− 1 at 6.4 h after spontaneous pressure losses at 2 h, 4 h, and 6 h, indicated by sharp drops in the DO signal (Fig. 3b).

Batch culture of P. putida with headspace aeration in 7.5 l pressure reactor. a Online data of overpressure, temperature, and stirring rate; b Online data of pCO2, CTR, and DOT; c Offline data of glucose concentration, CDW, and HAA concentration; Experimental conditions: 4.5 l filling volume; gas flow rate via headspace 4 l min− 1 (0-6.4 h) ; pH 7.00 ± 0.05 (12.5% (v/v) NH4OH).
Under these conditions, an oxygen-unlimited process was realized until glucose was consumed at 20.9 h (Fig. 3c). With a final value of 1.71 g l− 1 after 21 h (Fig. 3c), the highest HAA concentration was obtained within this study, which was 190% higher than in the unpressurized bioprocess at 30 °C and 120% higher compared with the unpressurized bioprocess at 23 °C (Table 1). Moreover, concerning STY and HAA-glucose yield, the pressurized process was the superior one with 0.08 gHAA l− 1 h− 1 and 0.09 gHAA gGlucose−1 (Table 1). In literature, lower STY of 0.06 gHAA l− 1 h− 1 [11] and 0.07 g l− 1 h− 1 [22] with P. putida and 0.07 gHAA l− 1 h− 1 with recombinant E. coli [51] were achieved on shake flask scale. Using foam fractionation with biomass recovery on bioreactor scale, a STY of 0.04 gHAA l− 1 h− 1 was reported by Blesken et al. [28], which is comparable to the STY of the unpressurized process at 23 °C presented in this study. Advantageously, temperature reduction can be realized without any technical effort compared to foam fractionation technology.
Nevertheless, a combination of headspace aeration and elevated reactor pressure needs to be critically discussed since accumulated CO2 often inhibits biomass formation [52, 53]. During the pressurized bioprocess, CO2 accumulated up to around 0.15 bar after 11.5 h in the culture broth and then further increased with the reduction of the cultivation temperature to its maximum value of 0.23 bar at 20.8 h (Fig. 3b). Although biomass formation was not exponential after 12 h, it still increased linearly in the presence of the high CO2 partial pressure (pCO2) (Fig. 3c) and did not show any declines in comparison to the unpressurized bioprocesses (0.9 bar, Fig. 2b and d). However, both unpressurized bioprocesses were oxygen-limited; therefore the microbes did not grow at the maximum growth rate. Hence, the comparable biomass concentrations could also result from the combination of the positive impact of oxygen availability and the negative effect of inhibiting CO2 during pressurized conditions making a direct deduction between growth and enhanced CO2 levels impossible. For P. putida, Follonier et al. found pCO2 > 0.015 bar to be growth-inhibiting without influencing productivity [39] and Knoll et al. reported 40% growth reduction at 0.2 bar pCO2 [41]. For Corynebacterium glutamicum, a constant growth rate was found until 0.26 bar pCO2 on glucose, but different behavior was observed on lactate [54]. Also, strong resistance to pCO2 of 0.6 bar and 0.8 bar of Corynebacterium glutamicum and Escherichia coli were found in pressurized carbon-limited fed-batch cultures [38]. From literature, one can conclude that the impact of dissolved CO2 on growth and productivity depends not only on the microorganism but also on growth conditions like carbon source and the medium itself as well as other primary nutrient limitations whose growth-limiting effect could be much larger. Although P. putida can tolerate pCO2 values up to 0.23 bar as demonstrated in the pressurized cultivation (Fig. 3c), the influence of the elevated pCO2 needs to be investigated in detail. In the special case of HAA formation, a high CO2 concentration might be even beneficial. In the first step of fatty acid de novo synthesis, catalyzed by the acetyl-CoA-carboxylase, which provides the precursors for HAA synthesis, CO2 is a key reactant. Increasing the pCO2 might thus shift the equilibrium towards the products, therefore, increasing the metabolic flow towards HAA. To elucidate a possible effect of an elevated CO2 concentration on the metabolic activity, a shake flask cultivation was aerated with a synthetic gas mixture containing 10% CO2, 25% O2 and 65% N2. In parallel, a reference cultivation was aerated with standard compressed air (0.045% CO2, 20.95% O2). For both growth conditions, nearly identical OTR trend lines (Fig. 4a) and similar cell dry weights and glucose concentrations (Fig. 4b) were measured. Without additional CO2, a slightly higher maximum HAA concentration of 1.06 g l− 1 was analyzed (0.88 g l− 1 for elevated CO2). This experiment proved that at least up to 10% there is no inhibitory effect of CO2. Furthermore, no positive impact on the HAA formation could be observed by an increased CO2 concentration.

Impact of an elevated CO2 concentration on growth and HAA formation in P. putida. a Online data of OTR; b Offline data of glucose concentration, CDW, and HAA concentration; Experimental conditions: Kuhner TOM system; 250 ml unbaffled Erlenmeyer flasks, 30 °C, 10 ml filling volume; aeration rate 20 ml min− 1, 300 rpm; gap between 5.5 and 6.0 h in a due to failure of data recording
3.3 Scalability
The consequence of headspace aeration is a reduced gas transfer between liquid and gaseous phases due to the limited surface area. To maintain this gas transfer at a constant level on different scales, the surface/filling volume ratio needs to be chosen as the scale-up criterion. This would result in low filling volumes in commercially relevant large-scale reactors making such bioprocesses uneconomical. A scalable game changer to realize headspace aeration on an industrial scale can be overpressure. The number of pressurizable large-scale bioreactor systems is increasing since it is an efficient technical solution to convert gaseous substrates into valuable products [55] and to increase the gas transfer in viscous microbial systems [56]. Pressure reactors can also be used for any unpressurized bioprocess, resulting in smaller space requirement compared to additional dedicated equipment such as foam fractionation columns. In contrast to in situ liquid-liquid extraction processes, the entire working volume can serve as reaction space to build up actively producing biomass. The additional application of membrane modules for bubble-free aeration would be an option to realize even higher gas transfer rates compared to the pressurized bioprocess. However, membrane modules as described in Bongartz et al. [30] are currently not available for all bioreactor scales. Finally, the reduction of temperature is an additional fully scalable and absolute simple approach to make headspace aeration applicable to handle foaming bioprocesses.
4 Conclusion
In this study, an alternative strategy for the foam-free production of HAA based on headspace aeration was developed. However, it became obvious that sufficient oxygen supply is crucial for HAA production in P. putida, which is challenging under headspace-aerated conditions. To reduce the oxygen uptake rate of P. putida, the cultivation temperature was decreased by 7 °C, which prolonged the process time but doubled the final HAA concentration up to 1.28 g l− 1. For further process intensification, the reactor system was pressurized up to 8 bar overpressure, ensuring oxygen-unlimited conditions over the total process time. Based on this process, it was demonstrated that the combination of headspace aeration, temperature reduction, and overpressure is a scalable strategy for foam-free production of bipolar substances with low technical effort.
Data availability
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
References
Garcia-Depraect O, Bordel S, Lebrero R, Santos-Beneit F, Borner RA, Borner T, et al. Inspired by nature: microbial production, degradation and valorization of biodegradable bioplastics for life-cycle-engineered products. Biotechnol Adv. 2021;53:107772.
Geyer R, Jambeck JR, Law KL. Production, use, and fate of all plastics ever made. Sci Adv. 2017;3:e1700782.
Mazhandu ZS, Muzenda E, Mamvura TA, Belaid M, Nhubu T. Integrated and consolidated review of plastic waste management and bio-based biodegradable plastics: challenges and opportunities. Sustainability. 2020;12:8360.
Lang S, Trowtizsch-Kienast W. Biotenside. 1 ed. Wiesbaden: Vieweg + Teubner Verlag Wiesbaden; 2002.
Bergström S, Theorell H, Davide H. Pyolipic acid, a metabolic product of Pseudomonas pyocyanea, active against Mycobacterium tuberculosis. Arch Biochem. 1946;10(1):165–6.
Jarvis FG, Johnson MJ. A glyco-lipide produced by Pseudomonas aeruginosa. J Am Chem Sci. 1949;71:4124–6.
Deziel E, Lepine F, Dennie D, Boismenu D, Mamer OA, Villemur R. Liquid chromatography/mass spectrometry analysis of mixtures of rhamnolipids produced by Pseudomonas aeruginosa strain 57RP grown on mannitol or naphthalene. Biochim Biophys Acta. 1999;1440:244–52.
Deziel E, Lepine F, Milot S, Villemur R. rhlA is required for the production of a novel biosurfactant promoting swarming motility in Pseudomonas aeruginosa: 3-(3-hydroxyalkanoyloxy)alkanoic acids (HAAs), the precursors of rhamnolipids. Microbiol (Reading). 2003;149:2005–13.
Zhu K, Rock CO. RhlA converts beta-hydroxyacyl-acyl carrier protein intermediates in fatty acid synthesis to the beta-hydroxydecanoyl-beta-hydroxydecanoate component of rhamnolipids in Pseudomonas aeruginosa. J Bacteriol. 2008;190:3147–54.
Abdel-Mawgoud AM, Lepine F, Deziel E. Rhamnolipids: diversity of structures, microbial origins and roles. Appl Microbiol Biotechnol. 2010;86:1323–36.
Tiso T, Zauter R, Tulke H, Leuchtle B, Li WJ, Behrens B, et al. Designer rhamnolipids by reduction of congener diversity: production and characterization. Microb Cell Fact. 2017;16:1–14.
Meyers J, Mensah JB, Holzhäuser FJ, Omari A, Blesken CC, Tiso T, et al. Electrochemical conversion of a bio-derivable hydroxy acid to a drop-in oxygenate diesel fuel. Energy Environ Sci. 2019;12:2406–11.
Tiso T, Narancic T, Wei R, Pollet E, Beagan N, Schröder K, et al. Bio-upcycling of polyethylene terephthalate. bioRxiv preprint. 2020.
de Kronemberger FA, Anna LMMS, Fernandes ACLB, de Menezes RR, Borges CP, Freire DMG. Oxygen-controlled biosurfactant production in a bench scale bioreactor. Appl Biochem Biotechnol. 2008;147:33–45.
Dolman BM, Wang F, Winterburn JB. Integrated production and separation of biosurfactants. Process Biochem. 2019;83:1–8.
Krieger N, Neto DC, Mitchell DA. Production of microbial biosurfactants by solid-state cultivation. In: Sen R, editor. Biosurfactants. New York: Springer New York; 2010. pp. 203–10.
Vardar-Sukan F. Efficiency of natural oils as antifoaming agents in bioprocesses. J Chem Technol Biotechnol. 1988;43:39–47.
Vardar-Sukan F. Foaming: consequences, prevention and destruction. Biotechnol Adv. 1998;16:913–48.
Routledge SJ. Beyond de-foaming: the effects of antifoams on bioprocess productivity. Comput Struct Biotechnol J. 2012;3:e201210014.
Varley J, Brown AK, Boyd JWR, Dodd PW, Gallagher S. Dynamic multi-point measurement of foam behaviour for a continuous fermentation over a range of key process variables. Biochem Eng J. 2004;20:61–72.
Stein J, Schlosser N, Bardl B, Peschel G, Meyer F, Kloss F, et al. Scalable downstream method for the cyclic lipopetide jagaricin. Eng Life Sci. 2021;1–7.
Blesken CC, Bator I, Eberlein C, Heipieper HJ, Tiso T, Blank LM. Genetic cell-surface modification for optimized foam fractionation. Front Bioeng Biotechnol. 2020;8: 572872.
Willenbacher J, Rau JT, Rogalla J, Syldatk C, Hausmann R. Foam-free production of surfactin via anaerobic fermentation of Bacillus subtilis DSM 10(T). AMB Express. 2015;5:21.
Demling P, von Campenhausen M, Grütering C, Tiso T, Jupke A, Blank LM. Selection of a recyclable in situ liquid–liquid extraction solvent for foam-free synthesis of rhamnolipids in a two-phase fermentation. Green Chem. 2020;22:8495–510.
Díaz De Rienzo MA, Kamalanathan ID, Martin PJ. Comparative study of the production of rhamnolipid biosurfactants by B. thailandensis E264 and P. aeruginosa ATCC 9027 using foam fractionation. Process Biochem. 2016;51:820–7.
Beuker J, Steier A, Wittgens A, Rosenau F, Henkel M, Hausmann R. Integrated foam fractionation for heterologous rhamnolipid production with recombinant Pseudomonas putida in a bioreactor. AMB Expr. 2016;6:11.
Guez JS, Vassaux A, Larroche C, Jacques P, Coutte F. New continuous process for the production of lipopeptide biosurfactants in foam overflowing bioreactor. Front Bioeng Biotechnol. 2021;9:678469.
Blesken CC, Strumpfler T, Tiso T, Blank LM. Uncoupling foam fractionation and foam adsorption for enhanced biosurfactant synthesis and recovery. Microorganisms. 2020;8:2029.
Coutte F, Lecouturier D, Yahia SA, Leclere V, Bechet M, Jacques P, et al. Production of surfactin and fengycin by Bacillus subtilis in a bubbleless membrane bioreactor. Appl Microbiol Biotechnol. 2010;87:499–507.
Bongartz P, Bator I, Baitalow K, Keller R, Tiso T, Blank LM, et al. A scalable bubble-free membrane aerator for biosurfactant production. Biotechnol Bioeng. 2021;118:3545–58.
Kuo TM, Lanser AC. Factors influencing the production of a novel compound, 7,10-dihydroxy-8(E)-octadecenoic acid, by Pseudomonas aeruginosa PR3 (NRRL B-18602) in batch cultures. Curr Microbiol. 2003;47:186–91.
Lopes M, Mota M, Belo I. Oxygen mass transfer rate in a pressurized lab-scale stirred bioreactor. Chem Eng Technol. 2013;36:1779–84.
Noorman HJ, van Winden W, Heijnen JJ, van der Lans RGJM. Intensification of biobased processes: RSC; 2018. pp. 1–41.
Belo I, Pinheiro R, Mota M. Fed-batch cultivation of Saccharomyces cerevisiae in a hyperbaric bioreactor. Biotechnol Prog. 2003;19:665–71.
Campani G, Ribeiro MPA, Horta ACL, Giordano RC, Badino AC, Zangirolami TC. Oxygen transfer in a pressurized airlift bioreactor. Bioprocess Biosyst Eng. 2015;38:1559–67.
Knabben I, Regestein L, Marquering F, Steinbusch S, Lara AR, Büchs J. High cell-density processes in batch mode of a genetically engineered Escherichia coli strain with minimized overflow metabolism using a pressurized bioreactor. J Biotechnol. 2010;150:73–9.
Knabben I, Regestein L, Schauf J, Steinbusch S, Büchs J. Linear correlation between online capacitance and offline biomass measurement up to high cell densities in Escherichia coli fermentations in a pilot-scale pressurized bioreactor. J Microbiol Biotechnol. 2011;21:204–11.
Knoll A, Bartsch S, Husemann B, Engel P, Schroer K, Ribeiro B, et al. High cell density cultivation of recombinant yeasts and bacteria under non-pressurized and pressurized conditions in stirred tank bioreactors. J Biotechnol. 2007;132:167–79.
Follonier S, Henes B, Panke S, Zinn M. Putting cells under pressure: a simple and efficient way to enhance the productivity of medium-chain-length polyhydroxyalkanoate in processes with Pseudomonas putida KT2440. Biotechnol Bioeng. 2012;109:451–61.
Knabben I, Regestein L, Grumbach C, Steinbusch S, Kunze G, Büchs J. Online determination of viable biomass up to very high cell densities in Arxula adeninivorans fermentations using an impedance signal. J Biotechnol. 2010;149:60–6.
Knoll A, Maier B, Tscherrig H, Büchs J. The oxygen mass transfer, carbon dioxide inhibition, heat removal, and the energy and cost efficiencies of high pressure fermentation. Adv Biochem Eng Biotechnol. 2005;92:77–99.
Kreyenschulte D, Emde F, Regestein L, Büchs J. Computational minimization of the specific energy demand of large-scale aerobic fermentation processes based on small-scale data. Chem Eng Sci. 2016;153:270–83.
Hartmans S, Smits JP, van der Werf MJ, Volkering F, de Bont JA. Metabolism of styrene oxide and 2-phenylethanol in the styrene-degrading Xanthobacter strain 124X. Appl Environ Microbiol. 1989;55:2850–5.
Peña-Ortiz L, Schlembach I, Lackner G, Regestein L. Impact of oxygen supply and scale up on Mycobacterium smegmatis cultivation and mycofactocin formation. Front Bioeng Biotechnol. 2020;8:593781.
Tiso T, Germer A, Küpper B, Wichmann R, Blank LM. Methods for recombinant rhamnolipid production. In: McGenity TJ, Timmis KN, Nogales Fernández B, editors. Hydrocarbon and lipid microbiology protocols. Berlin: Springer Berlin Heidelberg; 2016. p. 65–94.
Behrens B, Baune M, Jungkeit J, Tiso T, Blank LM, Hayen H. High performance liquid chromatography-charged aerosol detection applying an inverse gradient for quantification of rhamnolipid biosurfactants. J Chromatogr A. 2016;1455:125–32.
Demling P, Ankenbauer A, Klein B, Noack S, Tiso T, Takors R, et al. Pseudomonas putida KT2440 endures temporary oxygen limitations. Biotechnol Bioeng. 2021;118:4735–50.
Wittgens A, Kovacic F, Müller MM, Gerlitzki M, Santiago-Schübel B, Hofmann D, et al. Novel insights into biosynthesis and uptake of rhamnolipids and their precursors. Appl Microbiol Biotechnol. 2017;101:2865–78.
Ankenbauer A, Schafer RA, Viegas SC, Pobre V, Voss B, Arraiano CM, et al. Pseudomonas putida KT2440 is naturally endowed to withstand industrial-scale stress conditions. Microb Biotechnol. 2020;13:1145–61.
Abdel-Mawgoud AM, Lepine F, Deziel E. A stereospecific pathway diverts beta-oxidation intermediates to the biosynthesis of rhamnolipid biosurfactants. Chem Biol. 2014;21:156–64.
Germer A, Tiso T, Müller C, Behrens B, Vosse C, Scholz K, et al. Exploiting the natural diversity of RhlA acyltransferases for the synthesis of the rhamnolipid precursor 3-(3-hydroxyalkanoyloxy)alkanoic acid. Appl Environ Microbiol. 2020;86:e02317-19.
Dixon NM, Kell DB. The inhibition by CO2 of the growth and metabolism of micro-organisms. J Appl Bacteriol. 1989;67:109–36.
Blombach B, Takors R. CO2 - intrinsic product, essential substrate, and regulatory trigger of microbial and mammalian production processes. Front Bioeng Biotechnol. 2015;3:108.
Bäumchen C, Knoll A, Husemann B, Seletzky J, Maier B, Dietrich C, et al. Effect of elevated dissolved carbon dioxide concentrations on growth of Corynebacterium glutamicum on D-glucose and L-lactate. J Biotech. 2007;128:868–74.
Van Hecke W, Bockrath R, De Wever H. Effects of moderately elevated pressure on gas fermentation processes. Bioresour Technol. 2019;293:122129.
Schelden M, Lima W, Doerr EW, Wunderlich M, Rehmann L, Büchs J, et al. Online measurement of viscosity for biological systems in stirred tank bioreactors. Biotechnol Bioeng. 2017;114:990–7.
Acknowledgements
This work was supported by a grant of the European Social Fund ESF “Europe for Thuringia” project HoWi (2019FGR0079, LR, BB and SW). TT and LMB acknowledge additional funding by the German Research Foundation (DFG) under Germany’s Excellence Strategy within the Cluster of Excellence FSC 2186 “The Fuel Science Center”. The research was partially funded by the ministry of economic affairs, industry, climate action, and energy of the state of North Rhine-Westphalia within the framework of the „Exzellenz Start-up Center.NRW” in the program “Innovation Sprint” of the RWTH Aachen University. The authors thank Gudrun Krauter, Patrick Berthel, and Michael Meyer for their helpful technical support during fermentations and analytics. The biosurfactants team at the iAMB (Philipp Demling, Melanie Filbig, Tobias Karmaninski, Carina Michel, Conrad Müller, Gina Welsing) is acknowledged for many fruitful discussions.
Funding
Open Access funding enabled and organized by Projekt DEAL. This work was supported by a grant of the European Social Fund ESF “Europe for Thuringia” project HoWi (2019FGR0079, LR and SW). TT and LMB acknowledge additional funding by the German Research Foundation (DFG) under Germany’s Excellence Strategy within the Cluster of Excellence FSC 2186 “The Fuel Science Center”. The research was partially funded by the ministry of economic affairs, industry, climate action, and energy of the state of North Rhine-Westphalia within the framework of the „Exzellenz Start-up Center.NRW” in the program “Innovation Sprint” of the RWTH Aachen University.
Author information
Authors and Affiliations
Contributions
SW, LMB, TT, and LR have written the manuscript. SW, KW, and LE conducted all cultivations. BB developed and performed all analytical methods. TT, LMB, and LR supervised the study. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Weiser, S., Tiso, T., Willing, K. et al. Foam-free production of the rhamnolipid precursor 3-(3-hydroxyalkanoyloxy) alkanoic acid (HAA) by Pseudomonas putida. Discov Chem Eng 2, 8 (2022). https://doi.org/10.1007/s43938-022-00017-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s43938-022-00017-y