
Abstract
Allosuckling, the suckling of milk from a non-biological mother, occurs in some species of mammals. Allosuckling has not been quantified in baleen whale calves; therefore, we examine allosuckling in southern right whales (SRWs; Eubalaena australis) off Australia. SRWs are listed as Endangered under the Australian Environment Protection and Biodiversity Conservation Act as their numbers remain below the estimated historical abundance. On a small aggregation area, where there were three mother–calf pairs present, we aimed to quantify the proportion of time that calves allosuckled relative to the time spent filial suckling. To achieve this, we conducted unmanned aerial vehicle focal follows on mother–calf pairs and video recorded all interactions among pairs (n = 22 interactions, 3 h total observation time). During interactions, allosuckling occurred in seven interactions, and filial nursing occurred in 11 interactions. One of the calves performed allosuckling, and it was the largest calf with the largest mother. The calf allosuckled from both of the non-biological mothers present. The average proportion of time allosuckling per interaction was 4% (95% CI = ± 0.01, range = 0–0.25), whilst filial nursing for the same calf was 8% (95% CI = ± 0.02, range = 0–0.37). It is important to understand the frequency of allosuckling, and to quantify the energetic benefits for allosuckling calves and the energetic cost incurred by targeted non-biological mothers. This is particularly pertinent for capital breeders who do not replenish lost energy reserves until they migrate to their feeding grounds.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Allosuckling is a term used for when a lactating mother provides milk to nonfilial offspring, or rather, when young suckle milk from a non-biological mother (Arso Civil et al. 2021). This suckling strategy is highly intriguing in mammals, and occurs in various species of ungulates in the wild (e.g., Ekvall 1998; Zapata et al. 2009; Engelhardt et al. 2014; Saito and Idani 2018; Skok 2022). Marine mammals evolved from a common ancestor to terrestrial ungulates (Ursing and Arnason 1998; Geisler and Uhen 2003), and in marine mammals, allosuckling is fairly well documented in pinnipeds. Allosuckling occurs at higher rates in phocids (true seals) (Le Bœuf et al. 1989; Arso Civil et al. 2021), and at a lesser extent in otariids (eared seals; fur seals and sea lions) (e.g., Lunn 1992; De Bruyn et al. 2010; Pitcher et al. 2011; Sepúlveda and Harcourt 2021). In elephant seals (Mirounga angustirostris), pups may perform allosuckling and become “superweaners”, where the pup nurses from another mother after being weaned from their biological mother (Le Bœuf et al. 1989). The pup effectively gains additional milk from the non-biological mother. There is a large energetic gain for the pups suckling from two lactating females and the pups may weigh twice as much as pups that suckle from their biological mother only (filial nursing). Consequently, superweaners improve their body condition at the end of their parental investment (Le Bœuf et al. 1989), which can have benefits for later survival and reproductive performance (Lindström 1999; McMahon et al. 2000). Allosuckling can also occur prior to weaning, for example, Subantarctic fur seal pups (Arctocephalus tropicalis) attempt to steal milk unnoticed and acquire milk without consent from a non-biological mother across the ~ 10-month nursing season (Roux 1986; Georges and Guinet 2000; De Bruyn et al. 2010).
There are several hypotheses as to why allosuckling may have evolved (Packer et al. 1992; Roulin 2002; Gloneková et al. 2021), and it is commonly explained as ‘milk theft’ (also known as sneak-suckling and milk stealing) as in seal pups (Lunn 1992), or it may be reciprocal as in some populations of sperm whales (Physeter macrocephalus) who perform alloparental care (Gero et al. 2009; Konrad et al. 2019). Allosuckling also occurs in captivity, for example, in beluga whales (Delphinapterus leucas) (Leung et al. 2010), and in the terrestrial relatives of cetaceans, the hippopotamus (Hippopotamus amphibius) (Pluhácek and Bartošová 2011). In baleen whales, there is limited knowledge on allosuckling, as such large whales are not able to be held in captivity and underwater nursing events have been difficult to observe. Off South Africa, there were two cases of “non-offspring nursing” described during aerial flights where a lone southern right whale (Eubalaena australis) calf was observed attempting to suckle from multiple non-biological mothers (Best et al. 2015). However, allosuckling, where the calf has not been abandoned or orphaned and still has a biological mother present, has not been described or quantified in baleen whales.
Using aerial footage from unmanned aerial vehicles (UAVs), the aim of this research was to describe the presence of allosuckling, and quantify the proportion of time in which allosuckling occurs in southern right whale (SRW) calves compared to filial suckling. It is hypothesised that during interactions with mother–calf pairs, calves will allosuckle to a lesser extent compared to filial suckling. It is important to understand the frequency of allosuckling, as the provision of milk to non-offspring from lactating mothers involves a potential cost to themselves and to their own offspring. Lactating mothers carry the highest energetic demands in mammals (Oftedal 1985; Gittleman and Thompson 1988), including baleen whales (Lockyer 1981; Christiansen et al. 2018). Unlike otariids and toothed whales that can balance their energy expenditure with direct energy intake through foraging (income breeders), baleen whales are generally capital breeders and may not feed whilst on the breeding ground and hence are not able to replenish lost energy reserves until they migrate to their feeding ground (i.e., enduring multiple months without feeding) (Lockyer 1987; Stephens et al. 2009; Bierlich et al. 2022).
Methods
Study species and location
Data were collected off Encounter Bay, South Australia (35°32′S, 138°42′E), between August 14 and September 14, 2021. For study area details and map, see Sprogis et al. (2023). Encounter Bay is an aggregation area for SRWs, with an average of four mother–calf pairs residing in the area (Kemper et al. 2022). Mother–calf pairs are present over the cooler months (~ June–November), with an average residency in the area of 50 days (Kemper et al. 2022). On the breeding grounds, SRW mother–calf pairs typically rest and nurse close to shore (Nielsen et al. 2019b), providing a refuge from predation from killer whales (Orcinus orca) (Nielsen et al. 2019a). After the breeding season, Australian SRWs migrate to their offshore Southern Ocean feeding grounds (Mackay et al. 2020). As Australian SRW numbers remain below the estimated historical abundance and range (Department of Sustainability Environment Water Population and Communities 2012), they continue to be listed as Endangered under the Australian Environment Protection and Biodiversity Conservation Act (EPBC 1999).
Data collection: UAV behavioural focal follows
Methodology followed Sprogis et al. (2023). In brief, SRWs were searched for in good weather conditions (Beaufort Sea State, ~ BSS < 3, limited precipitation) along a ~ 15 km stretch of coast. Once whales were sighted, land-based behavioural focal follows of mother–calf pairs began using UAVs (DJI Phantom 4 Advanced, diameter = 350 mm, weight = 1368 g). Consecutive UAV flights were flown to obtain near-continuous focal follow video data (video = 2.7 K, 2720 ″ 1530, 48 fps). Each UAV flight was around 15 min (the duration of one battery). The UAV was flown at > 30 m altitude hovering above the whales, with the camera facing vertically down at a zenithal angle. Higher altitudes (e.g., 35 m) were flown when interactions between two mother–calf pairs occurred to ensure all whales were in-frame. At these altitudes, the noise produced by small quadcopter UAVs is likely close to ambient underwater noise levels in shallow waters (Christiansen et al. 2016). Off Encounter Bay, during days with relatively calm seas (BSS 1–3), the noise level along the surf beach is 74 dB re 1 μPa RMS (2 s; at the NL0.25 kHz band; 95th percentile 87 dB re 1 μPa RMS (2 s)) (Sprogis et al. 2023). During such weather conditions, the whales were unlikely to perceive the UAV noise above the underwater ambient noise level, and the noise from the UAV which penetrated the surface was likely to be above the hearing threshold of SRWs (Christiansen et al. 2016). Furthermore, Christiansen et al. (2020) showed that when hovering a UAV above SRW mother–calf pairs, no behavioural changes were detected when compared to control behaviour. Therefore, this technology was considered non-invasive and appropriate to record undisturbed behaviours. The UAV was flown over the same focal whales which remained in the study area. Individuals were identified from the unique callosity patterns on their heads (Payne et al. 1983; Kraus et al. 1986). The body length and width of each whale was calculated using photogrammetry methods following Christiansen et al. (2018).
Data processing and analyses
The behaviours the whales performed were registered in the UAV videos using the software Solomon Coder v19.08.02 (https://solomon.andraspeter.com/). Behaviours were defined as in the behavioural ethogram (e.g., nursing) in Sprogis et al. (2023). Apparent nursing was defined as when the calf dove below the peduncle of a lactating adult, with its head positioned continuously near the mammary gland area, with its body nearly motionless and its tail parallel to the adult’s tail or approaching adjacent to the adult’s body (following Videsen et al. 2017; Nielsen et al. 2019b). Successful nursing events can be short in duration, e.g., 30 and 23 s (Zoidis and Lomac-MacNair 2017; Tackaberry et al. 2020); therefore, no time limit on nursing was imposed. The head of the calf was out of view from the UAV, and milk was not always confirmed; therefore, the behaviour was considered as apparent nursing (herein nursing). The duration of each nursing event was registered. It is noted that there may be an overestimation of the time spent suckling; however, values were comparable to Nielsen et al. (2019b) and Azizeh et al. (2021). Behaviours of interest for this study were filial nursing and allosuckling. Filial nursing was defined as a calf nursing from its biological mother, and allosuckling was defined as a calf suckling from a non-biological mother.
A close interaction among mother–calf pairs was defined as when pairs were in the frame of the UAV video (e.g., < 50 m distance). During an interaction, whales were typically active and/or in direct contact with one another. The beginning and end of each interaction was registered, so that the total duration of each interaction could be calculated. Since multiple interactions could occur during a single UAV flight (as whales frequently joined, then separated, and then joined again), each occurrence was classified as a separate interaction.
Data where two mother–calves were together in the same frame of the UAV (interaction data) were analysed. The proportion of time allosuckling and filial nursing was calculated from the sum of the total time allosuckling and filial nursing within an interaction, respectively, and was divided by the total duration of the interaction. Each allosuckling event was assessed for the behavioural reaction of the non-biological mother (evasive: turning away and/or swimming away, or neutral: remaining stationary and/or no obvious reaction).
Results
Summary statistics and interaction data
Research effort across a month resulted in UAV videos being collected over 19 days. Effort to search for the whales and track their movements consisted of 138 h, during daylight hours from 07:00 to 17:00. During this period, 168 UAV flights were flown (duration of follows: mean = 65.6 min ± 48.1 SD min, range = 4–192 min, total = 35 h video) (Sprogis et al. 2023). There were three mother–calf pairs present, which were identified from their callosity patterns (Fig. S1), and the mother of each calf was identified by their arrival time into the study area. Mother–calf pair 1 was located in the region for the duration of the study, pair 2 was encountered across three sampling days, and pair 3 arrived in the study area towards the end of the season (Table 1). Pair 1 was longer in length than pair 3 and pair 2 (Table 1). There were 12 flights across five days, where the whales interacted in a group size of four (two mother–calf pairs). During these 12 flights (total = 6.8 h), whales interacted for 45% of the time on 22 occasions with an average interaction time of 8.46 ± 4.31 SD min (range = 1.41–14.14 min, total = 3.1 h video).
Allosuckling and filial nursing
Across the five days where mother–calf pairs interacted, allosuckling was observed across three days. Within the 22 interactions, allosuckling occurred during 7 interactions, and filial nursing occurred during 11 interactions (Fig. 1). Milk was observed in the calves’ mouth after some nursing events (supplementary material video 1). During the seven interactions where allosuckling occurred, there were 15 allosuckling bouts. Allosuckling was performed by one of the three calves, the calf from pair 1 (Calf 1). Calf 1 was the largest calf, who had the largest mother (Table 1; Fig. 2; supplementary material video 2). Calf 1 allosuckled from both of the other mothers, pair 2 and pair 3. The average proportion of time that calf 1 allosuckled per interaction (with the suckling bouts combined in duration) was 4% (95% CI = 0.04 ± 0.01, range = 0–0.25). Calf 1 also filial nursed more often during interactions than the other calves. The average proportion of time filial nursing per interaction (nursing bouts combined) for pair 1 was 8% (n = 10, 95% CI = 0.08 ± 0.02, range = 0–0.37), and pair 3 was 2% (n = 2, 95% CI = 0.02 ± 0.01, range = 0–0.46) (Fig. 3). There was one filial nursing event for pair 2 (n = 1, 95% CI = 0.003 ± 0.002, range = 0–0.07; Fig. 3). Across calves, the average proportion of time filial nursing during interactions (filial nursing bouts combined) was 3% (95% CI = 0.03 ± 0.009, range = 0–0.46).

Photographs representing different nursing positions under the peduncle region of the mother; a whilst the mother was logging on the surface, b whilst the mother was sub-surface logging below the surface and the calf was still visible in good water visibility, and c whilst a mother was slowly swimming and a non-biological calf was allosuckling. Photographs represent cropped still frames from UAV videos

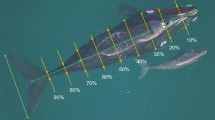
Two mother–calf pairs (pair 1 and pair 3), with allosuckling from the largest calf (calf 1) with the largest mother (pair 1 on the lower side of the photographs). a Directed movement from calf 1 towards a non-biological mother, b allosuckling from the calf below the peduncle region from the non-biological mother, and c calf 1 finished allosuckling and raising its head to take a breath, whilst its biological mother angles herself towards her calf. Photographs represent cropped still frames from UAV videos

Boxplots displaying the total proportion of time allosuckling and filial nursing during interactions of two mother–calf pairs (n = 22). Representing the three pairs present; pair 1 (n = 7 interactions allosuckling and 10 interactions filial nursing), pair 2 (n = 1 interaction filial nursing), and pair 3 (n = 2 interactions filial nursing). Boxplots displaying the median, quartile ranges, and outliers (circles)
The reaction of the non-biological mother to allosuckling was evasive or neutral, with 73% accounts being evasive (n = 11/15; supplementary material video 2), and 27% neutral accounts (n = 4/15; Fig. 4; supplementary material video 3). On one occasion, calf 1 was allosuckling and the targeted (non-biological) mother (mother 3) swum away leaving her calf behind, and the calf 3 was with mother 1 before it was separated from both mothers (Fig. 4a). Calf 3 remained separated from its mother for 60 s at a distance of > 20–50 m before the mother and calf swam towards each other and reunited. On all other occasions, calf 3 was besides the eye of its biological mother (Fig. 4b). When calf 1 was allosuckling, its biological mother was either following slowly behind (Fig. 4a), or stayed beside the trio (Fig. 4b).

Examples of the non-biological mother’s reaction to allosuckling. a Evasive reaction: mother 3 turning away and travelling, whilst calf 1 was allosuckling. In this example, the non-biological mother (mother 3) was travelling, leaving her biological calf (calf 3) behind. b Neutral reaction: mother 3 remaining stationary, whilst calf 1 was allosuckling. In this example, the biological calf (calf 3) stayed closed to its mother (mother 3). Calf 1 was allosuckling, whilst its biological mother (mother 1) remained close by
Discussion
Allosuckling in southern right whale calves
Allosuckling observed in SRWs off Encounter Bay appeared to be a direct and intentional movement from the calf, and the non-biological lactating mother was generally evasive (in 73% of attempts). Although the sample size was low and it was unknown if the allosuckling calf was successful at obtaining milk from the non-biological mother, it is presumed that the attempts were successful and that ‘milk-theft’ was occurring. The largest calf of the three calves in the study area, calf 1, was the only calf observed allosuckling. This could suggest that allosuckling is performed mainly by larger calves. The energetic gain from allosuckling was potentially high, and could have contributed to the large size of calf 1 compared to the other calves during the same time period, although the growth rate of the calf did not necessarily exceed expectation (Christiansen et al. 2018). In elephant seals, “superweaners” that perform milk theft may weigh twice as much compared to pups that do not allosuckle (Le Bœuf et al. 1989). Further research is required to increase the sample size to examine if some SRW calves follow a similar superweaner strategy. Allosuckling is presumed to have large energetic gains for the calf, as such attempts can be risky for the calf. For example, the non-biological mother could be aggressive towards unrelated calves, as is observed in harbour seals (Phoca vitulina) (Arso Civil et al. 2021) and Subantarctic fur seals where the non-biological might bite and toss the pup attempting to allosuckle (De Bruyn et al. 2010). Furthermore, during allosuckling, accidental calf switching between mothers could occur. This was the case in North Atlantic right whales (E. glacialis), where the calves were switched and they remained with their “adopted” mothers (Frasier et al. 2010). The calf swap was most likely an accident due to the close proximity of the mothers prior to the development of mother–calf recognition (Frasier et al. 2010).
Cost on the mother that is being allosuckled from
There is a cost of allosuckling to the targeted lactating mother, in terms of lost energy in the form of milk. Reproducing female baleen whales carry the highest energetic cost compared to other adults (males and non-reproducing females) and juveniles, as they carry the added cost of gestation and lactation (Lockyer 1981; Christiansen et al. 2018, 2022). As a capital breeder, the energy reserves of baleen whale mothers are not replenished during the nursing season, with females losing as much as 25% of their body condition across this time (e.g., ~ three months on the breeding grounds) (Christiansen et al. 2018). With this breeding strategy, the mother has a finite milk reserve, whilst she resides on the breeding ground. This milk has a high-fat content (17–52%) (for review, see McHuron et al. 2023), and consequently, the calf has a growth rate of ~ 3 cm (SD = 0.45 cm) a day in length (Christiansen et al. 2018). There is, therefore, a potentially greater cost to lactating mothers that are being allosuckled from, compared to a lactating mother that is not allosuckled from, as they may be providing milk on occasion for non-biological calves. Christiansen et al. (2018) showed that the growth rate of SRWs calves is directly related to the energy investment of the mother, which is reflected in the rate of loss in her body condition (i.e., body volume). The rate of energy transfer to the calf is in turn dictated by the mother’s absolute body volume, which is determined by her absolute size (i.e., length) and body condition (relative volume given her length) (Christiansen et al. 2018). A reduction in maternal body condition due to allosuckling by a non-biological calf will consequently reduce the mother’s investment in her own offspring, which in turn will reduce its growth rate, and possibly survival.
Phocids (true seals) are also capital breeders, using fat stores accumulated prior to the pupping season to help sustain themselves. Given this high cost for lactating females, allosuckling is therefore a perplexing behaviour to have evolved in capital breeders, as it appears disadvantageous to the lactating females that are being targeted. In phocids, allosuckling can occur at high rates. For example, in harbour seals, up to 47% of pups have been observed performing this behaviour in a given season, targeting up to 37% of lactating females (Arso Civil et al. 2021). In contrast, income breeders adopt a different strategy whereby lactating females continue to forage over the lactation period, and perhaps, this strategy offers itself to differences in allosuckling as in otariids. For example, the highest rate of allosuckling in otariids is only 7–11% for Antarctic fur seals (Arctocephalus gazella) (Lunn 1992; Gemmell 2003). Furthermore, perhaps, an income breeder strategy allows itself to alloparenting in toothed whales (e.g., sperm whales) (Gero et al. 2009), which is not common in baleen whales.
Allosuckling defined in southern right whales
We refer to the suckling of milk from a non-biological mother as allosuckling, which is the term commonly used in pinniped research (e.g., De Bruyn et al. 2010; Pitcher et al. 2011; Arso Civil et al. 2021). Another term commonly used in the scientific literature is allonursing (also nonfilial nursing, non-offspring nursing, and allomothering), which is defined as when milk is provided by the mother to nonfilial offspring (Roulin 2002; McHuron et al. 2023). The terms are similar, however, allosuckling is a term used based on the perspective of the offspring and is suitable in this context to account for the targeted behaviour of the calf. Here, allosuckling behaviour was likely initiated by the calf, as is the case in Subantarctic fur seal pups (Roux 1986; De Bruyn et al. 2010). Fostering, defined as exclusive parental care directed towards another’s young (Riedman 1982), was not the case in these observations as there was no exclusive care of another’s calf, and thus is not a concept touched upon here. The distinction and appropriate use of terminology among allosuckling, allonursing, and fostering behaviour in the literature is essential for future comparative studies, and for the drivers of these behaviours to be investigated in cetaceans.
Filial suckling in southern right whales
It was hypothesised that calves will allosuckle to a lesser extent compared to filial suckling. In alignment with the hypothesis, the calf that allosuckled (calf 1), did so to a lesser extent compared to filial suckling. During interactions, calf 1 allosuckled for 4% of time (± 0.07 SD, range = 0–0.25), whilst filial suckling occurred 8% of the time (± 0.14 SD, range = 0–0.37). With the amount of allosuckling and filial nursing combined, calf 1 potentially nursed for 12% of its time during interactions. In contrast, the smaller calves which did not allosuckle (calves 2 and 3), nursed less often from their biological mother during an interaction. The reduced amount of suckling could be due to the size difference among calves (i.e., calf 1 was larger than calves 2 and 3 and hence might have required more milk) and/or because the smaller calves were disturbed by interactions with calf 1. It remains unknown as to whether this lower rate of filial suckling, and/or the absence of allosuckling behaviour, affected the growth rate of calf 2 and 3.
Future research
This study describes allosuckling and quantifies the frequency of allosuckling compared to filial suckling in a capital breeding baleen whale, the SRW. It is possible that allosuckling occurs in other baleen whale species, such as the critically endangered North Atlantic right whale (Cooke 2020). Genetic analyses show that North Atlantic right whale calves associate with other mothers (Hamilton et al. 2022), and thus, it is possible these non-biological calves could be allosuckling. The use of UAVs has proved beneficial to gain an aerial perspective to observe nursing in clear waters (e.g., Nielsen et al. 2019b; Sprogis et al. 2023). Future research should aim to quantify the energetic benefits of allosuckling for both the calf that is performing the behaviour (added energy intake) and its biological mother (potential energy sparing strategy), as well as the energetic cost for the non-biological mother (reduced body condition) and her biological offspring (reduced milk transfer from biological mother). The effect of external factors, including the density of mother–calf pairs in a nursing area, the size and age of the calves and the mothers, and the timing in the breeding season, also requires investigation. Answering these questions will help researchers to understand how allosuckling might have evolved in SRWs, potential drivers of the behaviour, and its importance for the fitness of mother–calf pairs.
References
Arso Civil M, Hague E, Langley I, Scott-Hayward L (2021) Allo-suckling occurrence and its effect on lactation and nursing duration in harbour seals (Phoca vitulina) in Orkney. Scotland Behav Ecol Sociobiol 75:121. https://doi.org/10.1007/s00265-021-03051-y
Azizeh TR, Sprogis KR, Soley R, Nielsen MLK, Uhart MM, Sironi M, Marón CF, Bejder L, Madsen PT, Christiansen F (2021) Acute and chronic behavioral effects of kelp gull micropredation on southern right whale mother–calf pairs off Península Valdés, Argentina. Mar Ecol Prog Ser 668:133–148. https://doi.org/10.3354/meps13716
Best PB, Elwen SH, Palsbøll PJ, Thornton M, Austin E, Vinding K (2015) Possible non-offspring nursing in the southern right whale, Eubalaena australis. J Mammal 96:405–416. https://doi.org/10.1093/jmammal/gyv042
Bierlich KC, Hewitt J, Schick RS, Pallin L, Dale J, Friedlaender AS, Christiansen F, Sprogis KR, Dawn AH, Bird CN, Larsen GD, Nichols R, Shero MR, Goldbogen J, Read AJ, Johnston DW (2022) Seasonal gain in body condition of foraging humpback whales along the Western Antarctic Peninsula. Front Mar Sci 9:1036860. https://doi.org/10.3389/fmars.2022.1036860
Christiansen F, Rojano-Doñate L, Madsen PT, Bejder L (2016) Noise levels of multi-rotor unmanned aerial vehicles with implications for potential underwater impacts on marine mammals. Front Mar Sci 3:277. https://doi.org/10.3389/fmars.2016.00277
Christiansen F, Vivier F, Charlton C, Ward R, Amerson A, Burnell S, Bejder L (2018) Maternal body size and condition determine calf growth rates in southern right whales. Mar Ecol Prog Ser 592:267–281. https://doi.org/10.3354/meps12522
Christiansen F, Nielsen MLK, Charlton C, Bejder L, Madsen PT (2020) Southern right whales show no behavioural response to low noise levels from a nearby unmanned aerial vehicle. Mar Mamm Sci 36:953–963. https://doi.org/10.1111/mmms.12699
Christiansen F, Uhart MM, Bejder L, Clapham P, Ivashchenko Y, Tormosov D, Lewin N, Sironi M (2022) Foetal growth, birth size and energetic cost of gestation in southern right whales. J Physiol 600:2245–2266. https://doi.org/10.1113/JP282351
Cooke JG (2020) Eubalaena glacialis (errata version published in 2020). The IUCN Red List of Threatened Species 2020: e.T41712A178589687 2022. https://doi.org/10.2305/IUCN.UK.2020-2.RLTS.T41712A178589687.en
De Bruyn P, Cameron EZ, Tosh CA, Oosthuizen WC, Reisinger RR, Mufanadzo NT, Phalanndwa MV, Postma M, Wege M, Van der Merwe DS (2010) Prevalence of allosuckling behaviour in Subantarctic fur seal pups. Mamm Biol 75:555–560. https://doi.org/10.1016/j.mambio.2009.11.004
Department of Sustainability Environment Water Population and Communities (2012) Conservation management plan for the southern right whale. In: Australian Government (Ed) A recovery plan under the Environment Protection and Biodiversity Conservation Act 1999. 2011–2021, Canberra, Australia, p 68
Ekvall K (1998) Effects of social organization, age and aggressive behaviour on allosuckling in wild fallow deer. Anim Behav 56:695–703. https://doi.org/10.1006/anbe.1998.0825
Engelhardt SC, Weladji RB, Holand Ø, de Rioja CM, Ehmann RK, Nieminen M (2014) Allosuckling in reindeer (Rangifer tarandus): milk-theft, mismothering or kin selection? Behav Processes 107:133–141. https://doi.org/10.1016/j.beproc.2014.08.013
EPBC (1999) Species profile and threats database; Eubalaena australis — southern right whale. Environment Protection and Biodiversity Conservation Act. Australian Government 2023. https://www.environment.gov.au/cgi-bin/sprat/public/publicspecies.pl?taxon_id=40. Accessed 14 Aug 2021
Frasier TR, Hamilton PK, Brown MW, Kraus SD, White BN (2010) Reciprocal exchange and subsequent adoption of calves by two North Atlantic right whales (Eubalaena glacialis). Aquat Mamm 36:115–120. https://doi.org/10.1578/AM.36.2.2010.115
Geisler JH, Uhen MD (2003) Morphological support for a close relationship between hippos and whales. J Vert Paleontol 23:991–996
Gemmell NJ (2003) Kin selection may influence fostering behaviour in Antarctic fur seals (Arctocephalus gazella). Proc R Soc Lond B Biol Sci 270:2033–2037. https://doi.org/10.1098/rspb.2003.2467
Georges J-Y, Guinet C (2000) Maternal care in the subantarctic fur seals on Amsterdam Island. Ecology 81:295–308. https://doi.org/10.2307/177427
Gero S, Engelhaupt D, Rendell L, Whitehead H (2009) Who cares? Between-group variation in alloparental caregiving in sperm whales. Behav Ecol 20:838–843. https://doi.org/10.1093/beheco/arp068
Gittleman JL, Thompson SD (1988) Energy allocation in mammalian reproduction. Am Zool 28:863–875. https://doi.org/10.1093/icb/28.3.863
Gloneková M, Brandlová K, Pluháček J (2021) Further behavioural parameters support reciprocity and milk theft as explanations for giraffe allonursing. Sci Rep 11:7024. https://doi.org/10.1038/s41598-021-86499-2
Hamilton PK, Frasier BA, Conger LA, George RC, Jackson KA, Frasier TR (2022) Genetic identifications challenge our assumptions of physical development and mother–calf associations and separation times: a case study of the North Atlantic right whale (Eubalaena glacialis). Mamm Biol 102:1389–1408. https://doi.org/10.1007/s42991-021-00177-4
Kemper C, Steele-Collins E, Al-Humaidhi A, Segawa Fellowes T, Marsh O, Charlton C (2022) Encounter Bay, South Australia, an important aggregation and nursery area for the southern right whale, Eubalaena australis (Balaenidae: Cetacea). Trans R Soc S Aust 146:207–227. https://doi.org/10.1080/03721426.2021.2018759
Konrad CM, Frasier TR, Whitehead H, Gero S (2019) Kin selection and allocare in sperm whales. Behav Ecol 30:194–201. https://doi.org/10.1093/beheco/ary143
Kraus SD, Moore KE, Price CA, Crone MJ, Watkins WA, Winn HE, Prescott JH (1986) The use of photographs to identify individual North Atlantic right whales (Eubalaena glacialis). Report of the International Whaling Commission. Special Issue No. 10, pp 145–151
Le Bœuf BJ, Condit R, Reiter J (1989) Parental investment and the secondary sex ratio in northern elephant seals. Behav Ecol Sociobiol 25:109–117. https://doi.org/10.1007/BF00302927
Leung ES, Vergara V, Barrett-Lennard LG (2010) Allonursing in captive belugas (Delphinapterus leucas). Zoo Biol 29:633–637. https://doi.org/10.1002/zoo.20295
Lindström J (1999) Early development and fitness in birds and mammals. Trends Ecol Evol 14:343–348. https://doi.org/10.1016/S0169-5347(99)01639-0
Lockyer C (1981) Growth and energy budgets of large baleen whales from the Southern Hemisphere. Mammals in the seas. Food and Agriculture Organization of the United Nations, Rome, pp 379–487
Lockyer C (1987) The relationship between body fat, food resource and reproductive energy costs in North Atlantic fin whales (Balaenoptera physalus), Symposia of the Zoological Society of London, pp 343–361
Lunn N (1992) Fostering behaviour and milk stealing in Antarctic fur seals. Can J Zool 70:837–839. https://doi.org/10.1139/z92-119
Mackay AI, Bailleul F, Carroll EL, Andrews-Goff V, Baker CS, Bannister J, Boren L, Carlyon K, Donnelly DM, Double M, Goldsworthy SD, Harcourt R, Holman D, Lowther A, Parra GJ, Childerhouse SJ (2020) Satellite derived offshore migratory movements of southern right whales (Eubalaena australis) from Australian and New Zealand wintering grounds. PLoS ONE 15:e0231577
McHuron EA, Adamczak S, Costa DP, Booth C (2023) Estimating reproductive costs in marine mammal bioenergetic models: a review of current knowledge and data availability. Conserv Physiol 11:coac080. https://doi.org/10.1093/conphys/coac080
McMahon CR, Burton HR, Bester MN (2000) Weaning mass and the future survival of juvenile southern elephant seals, Mirounga leonina, at Macquarie Island. Antarct Sci 12:149–153. https://doi.org/10.1017/S0954102000000195
Nielsen MLK, Bejder L, Videsen SKA, Christiansen F, Madsen PT (2019a) Acoustic crypsis in southern right whale mother–calf pairs: infrequent, low-output calls to avoid predation? J Exp Biol 222:jeb90728. https://doi.org/10.1242/jeb.190728
Nielsen MLK, Sprogis KR, Bejder L, Madsen PT, Christiansen F (2019b) Behavioural development in southern right whale calves. Mar Ecol Prog Ser 629:219–234. https://doi.org/10.3354/meps13125
Oftedal O (1985) Pregnancy and lactation. Bioenergetics of wild herbivores. CRC Press, Boca Raton, pp 215–239
Packer C, Lewis S, Pusey A (1992) A comparative analysis of non-offspring nursing. Anim Behav 43:265–281. https://doi.org/10.1016/S0003-3472(05)80222-2
Payne R, Brazier O, Dorsey E, Perkins J, Rowntree V, Titus A (1983) External features in southern right whales, Eubalaena australis, and their use in identifying individuals. Communication and Behaviour of whales. Westview Press Inc, Boulder, Colorado, pp 371–445
Pitcher BJ, Ahonen H, Charrier I, Harcourt RG (2011) Allosuckling behavior in the Australian sea lion (Neophoca cinerea): an updated understanding. Mar Mamm Sci 27:881–888. https://doi.org/10.1111/j.1748-7692.2010.00440.x
Pluhácek J, Bartošová J (2011) A case of suckling and allosuckling behaviour in captive common hippopotamus. Mamm Biol 76:380–383. https://doi.org/10.1016/j.mambio.2010.06.007
Riedman ML (1982) The evolution of alloparental care and adoption in mammals and birds. Q Rev Biol 57:405–435
Roulin A (2002) Why do lactating females nurse alien offspring? A review of hypotheses and empirical evidence. Anim Behav 63:201–208. https://doi.org/10.1006/anbe.2001.1895
Roux J-P (1986) A successful subantarctic fur seal milk-thief. Mammalia 50:403–405
Saito M, Idani G (2018) Suckling and allosuckling behavior in wild giraffe (Giraffa camelopardalis tippelskirchi). Mamm Biol 93:1–4. https://doi.org/10.1016/j.mambio.2018.07.005
Sepúlveda M, Harcourt RG (2021) Maternal behavior in otariids and the walrus. Ethology and behavioral ecology of otariids and the odobenid. Springer International Publishing, Switzerland, pp 51–61
Skok J (2022) Suckling strategies in terrestrial ungulates (Artiodactyla, Perissodactyla) and the factors influencing them: the offspring perspective. CABI Rev 17:019. https://doi.org/10.1079/cabireviews202217019
Sprogis KR, Holman D, Arranz P, Christiansen F (2023) Effects of whale-watching activities on southern right whales in Encounter Bay, South Australia. Mar Policy 150:105525. https://doi.org/10.1016/j.marpol.2023.105525
Stephens PA, Boyd IL, McNamara JM, Houston AI (2009) Capital breeding and income breeding: their meaning, measurement, and worth. Ecology 90:2057–2067. https://doi.org/10.1890/08-1369.1
Tackaberry JE, Cade DE, Goldbogen JA, Wiley DN, Friedlaender AS, Stimpert AK (2020) From a calf’s perspective: humpback whale nursing behavior on two US feeding grounds. PeerJ 8:e8538. https://doi.org/10.7717/peerj.8538
Ursing BM, Arnason U (1998) Analyses of mitochondrial genomes strongly support a hippopotamus-whale clade. Proc R Soc Lond B Biol Sci 265:2251–2255. https://doi.org/10.1098/rspb.1998.0567
Videsen SKA, Bejder L, Johnson M, Madsen PT (2017) High suckling rates and acoustic crypsis of humpback whale neonates maximise potential for mother–calf energy transfer. Funct Ecol 31:1561–1573. https://doi.org/10.1111/1365-2435.12871
Zapata B, Gaete G, Correa LA, González BA, Ebensperger LA (2009) A case of allosuckling in wild guanacos (Lama guanicoe). J Ethol 27:295–297. https://doi.org/10.1007/s10164-008-0127-7
Zoidis AM, Lomac-MacNair KS (2017) A note on suckling behavior and laterality in nursing humpback whale calves from underwater observations. Animals 7:51. https://doi.org/10.3390/ani7070051
Acknowledgements
The authors especially thank Eyre and Fleurieu Peninsula DEW staff for logistics coordination and on-site field support, to Dirk Holman, and the marine rangers Jon, Darryl, Nikki, Lana and Shelley, and park rangers Seiji, Genki and Amy. Much appreciation to Anna Christie for fieldwork assistance. Thank you to locals in Encounter Bay for their support, including Debbie, Bob, Richard, Lea, Naomi and Maddie. Much appreciation to the anonymous reviewers for strengthening the manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. Thank you to the South Australian Department of Water and Environment (DEW) for support and funding.
Author information
Authors and Affiliations
Contributions
KS conceptualisation, methodology, term, analysis, investigation, resources, project administration, data curation, funding acquisition, and writing—original draft. FC methodology, funding acquisition, investigation, and writing—review and editing.
Corresponding author
Ethics declarations
Conflict of interest
There are no conflicts of interest.
Ethical approval
This research within the Encounter Marine Park was conducted with approval from the Department for Environment and Water Animal Ethics Committee (31/2021) and was licenced by the Department for Environment and Water on a scientific permit (A27080–2). UAV operations were operated under a Remotely piloted aircraft Operator Certificate through Interspacial Aviation Perth (https://interspacialaviation.com.au/), with a remote pilot licence in accordance with the Australian Civil Aviation Safety Authority (CASA) regulations, and UAVs registered with CASA.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling editor: Philip Dammann.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary file2 (MP4 46097 KB)
Supplementary file3 (MP4 96370 KB)
Supplementary file4 (MP4 295234 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sprogis, K.R., Christiansen, F. Allosuckling in southern right whale calves. Mamm Biol 104, 91–99 (2024). https://doi.org/10.1007/s42991-023-00392-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-023-00392-1