
Abstract
Tree hyraxes (Dendrohyrax spp.) are highly vocal, elusive nocturnal mammals that live in forests across much of Sub-Saharan Africa. Here, we studied how tree hyrax acoustic communication activity varies during the year in a moist montane forest in Taita Hills, Kenya. Our study increases the knowledge about the seasonal and diel variation in tree hyrax calling activity and on the impact of exogenous factors on the calling behavior of the species. A group of tree hyraxes called 700–2000 times during the night, and calling rate was highest during dry season months. Calling activity was bimodal, with peaks between 19.00 and 20.00 h in the evening and between 04.00 and 05.00 h in the morning; however, counter-calling bouts occurred throughout the night. Singing peaked in May, possibly coinciding with the species’ mating season. Calling activity decreased during full moon phases, probably because the animals felt vulnerable to predation. Tree hyrax calling was suppressed by rain, as rainfall is a powerful acoustic competitor. Annual changes in night temperature, the background chorus of vocalizing orthopterans, or the competing calls of nocturnal primates did not reduce tree hyrax calling rates. The latter suggests that sympatric nocturnal mammals have adapted to each other’s calls as an intrinsic feature of the soundscape of their forest habitat.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Seasonal and diel variation in acoustic communication of animals is connected to changes in response to social, intrinsic (nutritional status, reproductive cycle), and environmental factors (Walsh and Inglish 1989; Wilkins et al. 2013; Staicer et al. 2020). Studies on seasonal and diel variations in terrestrial mammals are limited in number, but studies have been done, e.g. on coyotes (Canis latrans) (Walsh and Inglish 1989), fallow deer (Dama dama) (McElligott et al. 1999; Pitcher et al. 2014) and black-and-gold howler monkeys (Alouatta caraya) (Pérez-Granados and Schuchmann 2021a). Long-term acoustic surveys are also commonly used in studies of aquatic mammals such as whales (e.g., Risch et al. 2013; Español-Jiménez and van der Schaar 2018; Narganes Homfeldt et al. 2022) and pinnipeds (Rouget et al. 2007; Kunc and Wolf 2008; Van Opzeeland 2010; Frouin-Mouy et al. 2015). This topic has been studied more in birds (e.g., Kloubec and Čapek 2005; Sandoval et al. 2016; Pérez-Granados and Schuchmann 2020; 2021b; Pérez-Granados et al. 2022) and amphibians (e.g., Pereyra et al. 2016; Nelson et al. 2017; Boquimpani-Freitas et al. 2007). These studies have revealed patterns related to distribution, breeding periods, responses to moon cycle and rain, and also diel differences between species.
Changes in acoustic communication may be related to food availability. For example, loud roars of black-and-gold howler monkey in the Brazilian Pantanal are most common during the time when fruits are most abundant (Pérez-Granados and Schuchmann 2021a). In black-and-gold howler monkeys, this may indicate that major feeding sites are defended by roaring.
Seasonal patterns, with high calling rate during the mating season, are found in some mammalian species. For example, fallow deer bucks are highly vocal during the mating season, when call rates of males may be over 3000 groans per hour (McElligott et al. 1999; Pitcher et al. 2014). Such a high calling rate is observed when both oestrous females and competing bucks are present. Galápagos sea lions (Zalophus wollebaeki) are vocally active during the breeding season, and territorial males vocalize more than non-territorial males (Kunc and Wolf 2008). In this species, territorial vocalizations are used in the context of male–male interactions, and vocal activity is higher if there are several females around. Thus, females may assess males’ quality by their calls. Also, coyotes are more vocal during their reproductive period (Walsh and Inglis 1989).
Behavior, including acoustic communication of some nocturnal animals, is also affected by periodic phases of moonlight (Kronfeld-Schor et al. 2013). Often potential prey species reduce their activities during full moon, to avoid being spotted by predators (Julien-Laferriere 1997; Starr et al. 2012; Studd et al. 2019). Studies have found that activity patterns of rodents, lagomorphs, chiropterans and carnivores are suppressed by moonlight (Prugh and Golden 2014). However, some species increase their activity during bright moonlight, as moonlight may improve the possibility to forage more effectively, and/or increase the chance to detect predators (Nash 2007). For example, small primates, spectral tarsiers (Tarsius spectrum) increase their activity during bright moonlight (Gursky 2003).
Rainfall is one of the most obvious and often dominant geophonic sources of sound in nature (Sánchez-Giraldo et al. 2020). Rainfall causes noise, which during heavy outpours can reach up to 10 kHz, thus reducing or even blocking the acoustic window for most other sounds. However, relatively few studies have focused on rainfall identification in acoustic recordings of terrestrial ecosystems (Ferroudj et al. 2014; Bedoya et al. 2017; Sánchez-Giraldo et al. 2020) . In the tropics, rainfall is known to influence the temporal activity patterns of African elephants (Loxodonta africana) (Birkett et al. 2012), anurans (Gottsberger and Gruber 2004) and birds (Keast and Marshall 1954). Rain may also cause short time-span changes in animal behavior. For example, rain stimulates sexual behavior in Galápagos finches (Lack 1950), and in anurans, rain may interrupt calling, probably because of acoustic interference (Zina and Haddad 2005).
The insect choruses of tropical forests can often cause a significant amount of at times ear-piercing background noise (Luther and Gentry 2013; Hart et al. 2015). In East Africa, the most vocal insects are often orthopterans including bush crickets and crickets (Hemp 2021). In Central America, some birds evade competition with high amplitude cicada choruses by avoiding sounds in the same frequencies or by communicating at times when the insect chorus has died down (Hart et al. 2015; Stanley et al. 2016). When insect noise was studied at different heights up until 21 m, no major differences in signal attenuation were observed along the vertical canopy gradient (Ellinger and Hödl 2003). In unaltered, old growth habitats birds, many mammals and amphibians tend to vocalize in acoustical niches unimpeded by background noise caused by chorusing insects (Krause 1987; Brumm 2006; Egnor et al. 2007; Wilson et al. 2016).
Tree hyraxes (Dendrohyrax spp.) are medium-sized, arboreal, nocturnal mammals that mainly browse on leaves of woody plants (Roberts et al. 2013) (Fig. 1). Trees and woody climbers used by tree hyraxes for feeding and resting in Taita Hills are listed in Rosti et al. (2022).

Taita tree hyrax (Dendrohyrax sp.) in Ngangao Forest, Taita Hills, Kenya. Photo: Hanna Rosti
In the Taita Hills, other vocally active nocturnal callers are two species of galagids (Galagidae), the small-eared greater galago (Otolemur garnettii lasiotis), from here on referred to as ‘Otolemur’, and the local dwarf galago (Paragalago sp.). Both primates have diverse and species-specific vocal repertoires (Bettridge et al. 2019; Butynski et al. 2006; Harcourt and Perkin 2013). Otolemur is common in both forests, farmlands, and villages, whereas the dwarf galago is a very rare and nearly extinct species strictly confined to indigenous montane forests (Rosti et al. 2020b; Pihlström et al. 2021). Especially the relatively large Otolemur is a potential acoustic competitor of tree hyraxes, as it uses the same frequencies in its acoustic communication.
We studied acoustic communication of tree hyraxes, which represent a characteristic and loud component of the diverse and dynamic night-time soundscape of indigenous montane forests of the Taita Hills. We analyze annual, monthly, nightly, and short-term temporal patterns of acoustic communication. Our first aim is to detect possible links between vocal activity and environmental fluctuations, including those caused by the moon cycle and seasonal differences in night-time temperatures. The second aim is to reveal possible effects of acoustic competition on tree hyrax calling activity, with special emphasis on background noise caused by rain, Otolemurs, and the Orthoptera choir. In addition to calling activity, we describe monthly temporal patterns of tree hyrax songs. We predict tree hyrax calling activity to fluctuate with the seasonal fluctuation of dry and wet seasons. We also expect rainfall and full moon to decrease calling activity of tree hyraxes. Regarding the Orthoptera choir and Otolemur calls, we predict that background noise caused by the potential competitors will not have a marked impact on the calling activity of tree hyraxes, as all these animals have lived in the same soundscape for possibly millions of years and should thus have had ample time to adapt to each other.
Methods
Study site
The Taita Hills belong to the Eastern Arc Mountains in south-eastern Kenya, where they rise from the surrounding dry plains to a series of mountain ridges. The Taita Hills are part of the Eastern Afromontane Biodiversity Hotspot (Myers et al. 2000; Burgess et al. 2007; Gereau et al. 2016). Combined natural moist tropical cloud forest cover in the Taita Hills is about 8 km2 (Wekesa et al. 2020; Zhao et al. 2022). The remaining fragments surrounding the peaks of the mountain ridges are one of the most threatened forests worldwide (Burgess et al. 2007; Githuru et al. 2011). Forest cover of these indigenous forest fragments decreased between 2003 and 2018 (Teucher et al. 2020; Wekesa et al. 2020).
The research was conducted in Ngangao Forest (size 120 ha), Taita Hills (− 3.3703 S, 38.342 E) at the altitude 1790 m a.s.l. (Fig. 2a). Ngangao is a remnant patch of most indigenous lower montane forest with a high and multi-layered canopy (Thijs 2015). Examples of tree species include Pouteria adolfi-friedericii, Newtonia buchananii, Tabernaemontana stapfiana, Albizia gummifera, Strombosia schefflera, Syzygium cordatum and Ficus thonnicii, often supporting dense growths of woody climbers. We chose the research site based on the results of our previous study (Rosti et al. 2022), where we found the chosen site to have the highest tree hyrax density anywhere in Taita Hills. We estimate that it may support a group of 20 or more tree hyraxes. However, the animals are rarely seen and impossible to count, as they mostly inhabit the higher canopy layers reaching elevations up to 48 m (Fig. 2b).

© 2015 Google)
The study site in Ngangao Forest, Taita Hills, Kenya. a Location of Taita Hills, Kenya, also showing the locations of remaining indigenous moist montane forests (yellow) and the study site in Ngangao Forest (red triangle) (Map data. b Ground lidar image depicting transect of 25 m in multi-layered and wine-rich canopy structure. The maximum height of the emergent tree is 48 m.
The distance from the study site to the forest edge was 300 m. The night-time temperatures during 2021 inside the forest varied between 10.0 and 19.2 °C (Fig. 3a). Annual rainfall in 2021 was 1140 mm. Longer rains typically occur in March, April, and May, and short rains in November and December (Fig. 3b). The upper slopes of the mountains receive moisture brought by the trade winds, and this is enough to sustain evergreen montane forests.

Climate of the study site in Ngangao Forest, Kenya. a Variation of monthly minimum temperatures in 2021, measured from 15 cm above the ground. b Monthly precipitation in 2021 from Helsinki University weather station at Kishenyi (elev. 1590 m a.s.l., ca. 1 km from study site)
Study species
Tree hyraxes belong to the Afrotheria group and are thus distantly related to elephants and sirenians (Springer et al. 1997; Stanhope et al. 1998; Nishihara et al. 2005; Tabuce et al. 2008). Tree hyraxes have labile body temperatures, and this allows male tree hyraxes to have internal testes (Glover and Sale 1968). As tree hyraxes are difficult to locate and observe, only a few behavioral studies exist. Milner and Harris (1999a, b) studied a population of Southern tree hyrax (Dendrohyrax arboreus) in the Parc National des Volcans, Rwanda. They discovered that at this location, D. arboreus was only using loud calls and nocturnal screaming during the dry season, and vocalizations had different temporal pattern during dry and wet season. Over the 4-month study period, tree hyrax ranges continuously changed (Milner and Harris 1999b). Core areas of females overlapped, with several females occupying male territories, but no evidence of males sharing common habitats was found. Djossa et al. (2012) studied calling activity and patterns of what they considered to represent a Western tree hyrax (Dendrohyrax dorsalis sylvestris) with passive acoustic monitoring (PAM) in a forested area of 162 km2 in Benin and reported significantly higher calling rates from interior areas of the forests. However, Oates et al. (2022) recently described a new tree hyrax species from the same area, the Benin tree hyrax (Dendrohyrax interfluvialis). Thus, the species studied by Djossa et al. (2012) was in fact D. interfluvialis. Topp-Jørgensen et al. (2008) estimated tree hyrax density in Udzungwa Mountains, Tanzania, by using circular plot counts of calling individuals, latrine counts, and daytime transects. They concluded that tree hyrax population density decreases with increased human disturbance, hunting and logging. Rosti et al. (2022) estimated tree hyrax (Dendrohyrax sp.) population density in Taita Hills using PAM in five different forests and concluded that tree hyrax density significantly decreases with decreasing forest size. Tree hyrax populations that encounter human disturbance changed their bimodal calling pattern and only called between 02.00 and 04.00 in the morning, possibly to avoid being detected by poachers (Rosti et al. 2022). Other studies have concluded that tree hyrax density is highest in natural intact moist forest, with old hollow trees, woody lianas and closed canopy, as these are elements of safety for tree hyraxes (Kundaeli 1976; Gaylard and Kerley 2001; Roberts et al. 2013; Opperman et al. 2018). However, tree hyraxes may be found in caves and rocky outcrops, where they can also find hideouts (Kingdon 1971; Seibt et al. 1977; Rosti et al. 2023).
Acoustic calls and songs of tree hyraxes in the Taita Hills
In the Taita Hills, acoustic communication of tree hyraxes is very active, with up to 2000 calls recorded from a single forest site during one night (Rosti et al. 2022). Some calls are very loud, exceeding 100 dB (Rosti et al. 2020a). Tree hyrax calls are short, and typically repeated 1–100 times by the same individual (depending on the call type). The vocalizations cover a lot of acoustic space with frequency ranges between 200 and 20 000 Hz (Rosti et al. 2020a). The dominant frequency range is from close to 0 to 6500 Hz, and harmonics are rare. Many vocalizations are counter-calls between different individuals, and often used as calling sequences. Most common calls used by tree hyraxes in the Taita Hills are strangled thwack and hac calls (Fig. S1) (Rosti et al. 2020a, b). However, tree hyraxes in the Taita Hills have a wide repertoire, and different calls are consistently modified and recombined. Most likely tree hyraxes have individual batches in their calls, making them individually different. This makes description of the calls challenging. Calls may exceed over 100 dB, or they may be quiet. The animals may call from all heights, and from the ground. Typically, 4–6 individuals take part in counter-calling sequences from different parts of the group’s habitat. Many individuals from the group keep on resting or feeding, and do not participate in counter-calling. Important and interesting elements of acoustic signals of tree hyrax in Taita Hills are songs that were found only recently (Rosti et al. 2020a, b). Songs are by definition long, and complex vocalizations, and usually, but not always, produced by males (Catchpole and Slater 2008). Songs have been studied extensively in rock hyraxes (Procavia capensis) (Koren 2006; Koren and Geffren 2011; Demartsev et al. 2014; 2017; 2022). Tree hyrax songs from Taita Hills and rock hyrax songs have similar vocal elements that suggest shared evolution. Wheeze from Taita Hills tree hyrax is comparable to rock hyrax wail, and both species have chuck calls (Demartsev et al. 2017; Rosti et al. 2020a; 2022). In rock hyraxes, the singers are mostly males, and songs have individual signatures (Demartsev et al. 2022). High-ranking rock hyrax males have better rhythmic stability in their songs and better reproductive success. Tree hyrax songs have been recorded from the forests of Ngangao, Mbololo and Vuria in Taita Hills (Rosti et al. 2022). We have categorized a calling sequence as a song if it has at least three continuous vocal sequences each lasting more than 5 s; between these song periods there are periods of silence (Fig. S2). Typically, tree hyrax singing continues from 20 min to3 h. If more than one individual is singing, songs will last longer. Songs of one individual tree hyrax may continue well over 20 min, with some pauses between sequences. When several individuals are singing, the singing bout will last longer.
Data collection
Recordings
Passive acoustic monitoring (PAM), offers an effective, and increasingly more common method for documenting and analysing trends in biological communities (Thompson et al. 2009; Marques et al. 2013; Hill et al. 2018; Lawson 2022). For this study, recordings were made with passive recorders, AudioMoths (v1.1.0 Open Acoustics Devices, Southampton, UK), between 19.00 and 06.00 for a week in each month from 1 February 2021 to 31 May 2022. Taita tree hyraxes call almost exclusively during the night between 19.00 and 06.00 (Rosti et al. 2022). A recorder was placed at four meters above ground level on a very large Pouteria adolfi-friedericii trunk providing support to a sizeable strangler fig (Ficus thonningii). Sample rate was 44,100 Hz, later (as AudioMoths were updated) 48,000 Hz, period of recording 60 s. Due to failed recordings or timestamps and other errors, data collection was not successful in April and May 2021. These months were added to the data from 2022.
Climate data
Rainfall was measured at 1.5 m height using a tipping bucket rain gauge (Campbell ARG100) at the weather station in Kishenyi valley (elev. 1590 m a.s.l.). The automatic weather station was located next to an agricultural field at a distance of 1 km from the edge of Ngangao Forest. The rainfall sum was measured every 1 min and recorded every 30 min.
Within-canopy temperature was measured using a TMS-4 sensor produced by TOMST (Prague, Czechia). The TMS-4 monitors soil moisture, soil temperature (at 15 cm depth), surface and near-surface air (15 cm above ground) temperatures (Wild et al. 2019). In this study, only the soil moisture and air temperature at 15 cm above ground were used. Measurements were taken every 15 min from February 2021 to May 2022. Within-canopy temperatures (i.e., microclimate) can differ from open-air conditions (i.e., macroclimate) by several degrees. Hence, this information can provide a better indicator of the conditions experienced by organisms inside the forest.
Automatic sound detection
The response variable, i.e., the call rate per hour, was calculated using automatic detection in Raven Pro 1.6 (Cornell University, Ithaca, NY, USA). The total number of analyzed hours was 539. A band limited interactive detector was used to calculate calls per hour with the following settings: min frequency 1000 Hz, max frequency 4500 Hz, min duration 0.01 s, max duration 2 s, min separation 0.23 s. Signal to noise ratio (SNR) 70%, threshold above 10. These settings were chosen after experimenting with different settings. All recordings were observed visually, and false positives were removed. False positives were typically due to calls by either Otolemurs or nocturnal birds. It is important to note that these automatic detection settings did not calculate songs. This is because the maximum duration was set to 2 s, and the duration of a song is always longer. Songs were marked as present or absent during the hourly data by visual and auditory inspection.
Variables used for analysis
The analysis intended to identify behavioral differences between different months, at night from 19.00 and 06.00. All variables used in the statistical analysis are given in Table 1.
Rainfall was classified with two levels, 1—no rain or light rain under 4 kHz, or 2 heavy rainfall exceeding 4 kHz (Fig. S3a, b, c). Orthoptera choir was classified with three levels, 1–no Orthoptera choir, 2–non-constant Orthoptera choir and 3–constant Orthoptera choir (Fig. S3d, e, f). Otolemur loud calls, trailing call and cluster squawks were counted visually from spectrograms (Fig. S4).
Statistical analysis
Statistical analyses were performed in RStudio 2022.07.01 with R version 4.1.2 (R Core Team 2020; Rstudio Team 2020). Data exploration was done according to Zuur and Ieno (2016). This included the detection of outliers, the exclusion of multi-collinearities (via variance inflation factors VIF), and various visualizations to reveal patterns of collinearity, non-linearity, and interactions. Collinear variables (with VIF > 2) were removed from the analysis. For the models, continuous covariates were standardized using the mySTD function in RStudio. The package glmmTMB (Brooks et al. 2017) was used to fit the generalized linear mixed-effects (GLMM) models. To ensure that GLMM model fits were good, we checked residual patterns by using the DHARMa package (Hartig 2022). Model performance was assessed with package performance (Lüdecke et al. 2021). The package ggplot2 was used to create graphs (Wickham 2016). Mixed models for nested data and zero inflated models (model 2) were built, simulated and validated according to Zuur et al. (2012). We constructed two models using the response variable ‘call rate per hour’.
Model 1 analyzed hourly and monthly differences in calling activity and interrelations with temperature, humidity, and moonlight. Model 1 was a GLMM with negative binomial distribution from 539 h of data from all the months during the year (Table 2). Zero inflation was 5.2%. Model assumptions were verified by plotting residuals versus fitted values, versus each covariate in the model and versus each covariate not in the model (Zuur et al. 2012).
Model 2 analyzed interrelations and acoustic competition between calls, rain, Orthoptera choir and Otolemurs. Calls were calculated visually and with an interactive detector (with same settings as above) from spectrograms in Raven Pro. All variables were measures from the same minute. This analysis was done using a GLMM for zero inflated negative binomial data (Table 3). 1-min recordings were analyzed every 15 min between 19.00 and 06.00 from June to December, 2 nights in each month. A total of 600 min was included in the analysis. Data had 52% zeros. Analyses based on GLMM for negative binomial data showed that the model was able to cope with zeros, however, GLMM for zero inflated data had a lower AIC and was thus used as a final model. Model assumptions were always verified by plotting residuals versus fitted values, versus each covariate in the model and versus each covariate not in the model (Zuur and Ieno 2016).
Results
Monthly differences in calling activity
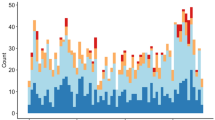
There were significant differences in acoustic communication activity of tree hyraxes between different months (Fig. 4, Table 2). Tree hyrax calling rate per hour was high during the dry months from January to February, and from June to September, and low during rainy season months. Average calling rates ranged from 700 calls per night in December to 2000 calls per night in February. However, there was considerable variation in calling activity between different nights. The vocal activity has a bimodal pattern, with a high number of calls during early evening and again in the morning (Fig. 5). Highest call rates were typically recorded from 19.00 and 20.00 in the evening. After this, the rate of calling decreased, and typically peaked again at 04.00 and 05.00 in the morning. During the night, intensive bouts of counter-calling could occur at any time.

Tree hyrax total calling activity during nights in different months of the year

Tree hyrax calling activity in different months and hours (between 19.00 and 06.00) in the Ngangao Forest, Taita Hills during 2021–2022. There are statistically significant differences in calling activity between months and hours. The line indicates the average call rate per hour and the smoother indicates conditional means
Tree hyraxes tended to call less during full moon (Fig. 6), missing the level of significance (P = 0.045). During heavy rain, tree hyraxes called significantly less. However, temperature was not significant. The overall model including the random factors explained 60% of the variance (conditional R2 = 0.60), and 45% of the variance without the random effects (marginal R2 = 0.45).

Effect of moon phases on tree hyrax calling activity. The number of tree hyrax calls per hour during (1) moonless nights, (2) half-moon, and (3) full moon
Response to acoustic competition
The analysis of acoustic competition from one-minute recordings (model 2) revealed that rain was the only covariate with significant effect (Table 3, Fig. 7a). Rain was present in about 50% of the recorded hours. Heavy rain was a strong acoustic competitor and reduced calling significantly. Neither Otolemur calls nor the Orthopteran choir influenced tree hyrax calling activity (Fig. 7b). Model 2 showed an intermediately strong explained variance (conditional R2 = 0.40, marginal R2 = 0.33) although a low repeatability across consecutive nights.

Effect of background noise caused by rain and chorusing orthopterans on tree hyrax calling activity. a Rainfall Level 1 and 2. b Orthoptera chorus Levels 1–3
Singing
Tree hyraxes sang throughout the year (Fig. 8). Within the total of 539 h of recordings, 55 h (10.2% of recorded hours) contained singing. Singing was most frequent in May, February, January, and September. This descriptive approach points out that rain tended to reduce singing, as during 75% of all hours with singing it was not raining.

Variation in singing activity of tree hyraxes during the year. The bars show the percentage of hours with singing of the total recorded hours per month
Discussion
Bioacoustics may be used for estimating the population size and different aspects of behavioral ecology of vocally active nocturnal mammals. However, this requires some previous knowledge of the temporal patterns of the species’ natural calling activity and responses to various sources of background noise. Our findings revealed that in Taita Hills, tree hyraxes were acoustically most active in January and February. These are dry season months after the short rains, when the forest canopy is dense with young leaves, which is the preferred food of tree hyraxes. Tree hyrax calling activity was bimodal with highest calling rates between 19.00 and 20.00 in the evening and between 04.00 and 05.00 in the morning. A similar bimodal calling pattern has previously been described for tree hyrax populations in Rwanda and Benin (Milner and Harris 1999a; Djossa et al. 2012).
Tree hyraxes seem to decrease their calling activity during full moon, which may represent a response to avoid predators during well-lit nights. This is in accordance to the findings on several other nocturnal mammal species, which are known to change to more sheltered habitats or reduce their activity during full moon nights (e.g., Kronfeld-Schor et al. 2013; Gutman et al. 2011; Pratas-Santiago et al. 2017). Owl monkeys (Aotus azarai azarai) are primates that are cathemeral, or active both diurnally and nocturnally (Fernandez-Duque 2003). On average, owl monkeys are active for five hours during the day, and four hours during the night. During full moon, owl monkeys are more active during the night and less active during the following day, meaning that the preceding night has an impact on the following day’s activity. It is possible that many or some tree hyrax populations are also cathemeral in their activity patterns, even though almost all vocalizations are emitted during the night.
Temperature may also affect the vocalizing behavior of tree hyraxes, as their relatives, rock hyraxes have a labile internal temperature regulation system (Hoeck 1975; Brown and Downs 2006), and their fibrous energy-poor diet requires the use of energy conservation strategies like sun basking, huddling, and heaping, i.e., being close to each other and even on top of each other (Fey 1960; Sale 1970; Bartholomew and Rainy 1971; Brown and Downs 2007). During colder temperatures, rock hyraxes are heaping and huddling with each other in their holes and when the sun comes out, they rest in the sun, basking in it (Sale 1970). In the sun, rock hyraxes keep at least a small distance to conspecifics, suggesting that distance to others is mainly dependent on the outside temperature and used as a method of supporting body temperature. For such energy conserving, temperature regulation related reasons, we hypothesized that tree hyraxes living on the mountains with cold nights might conserve energy during cold nights by calling less. However, nightly temperatures did not have a detectable effect on tree hyrax calling activity. In the Taita Hills, temperatures may fall between 10 and 13 °C during the night. A similar observation was made by Milner and Harris (1999a), who, while studying activity patterns of Dendrohyrax arboreus, found no significant correlation between temperature and the number of calls per night. Possibly, the tree hyraxes, and in particular tree hyrax populations living on higher altitudes in the mountains, have a faster metabolic rate than their diurnal relatives, the rock hyraxes.
Social situations, including the presence of males, can influence vocalization patterns in some species. For example, male simakobu monkeys (Simias concolor) from Siberut Island, Indonesia, give their longest calling bouts at dawn (Erb et al. 2016). This pattern cannot be explained by temperature or fruit availability, and it seems to be mainly influenced by chorusing behavior of other males. Thus, male simakobus motivate each other to call longer, and calling does not seem to be energetically costly. This group stimulation of counter-calling behavior seems to have a similar impact in tree hyraxes. In eastern tree hyrax, counter-calling will last longer, and animals use more variable calls when more animals are calling together. Whether calls differ between sexes in tree hyraxes is not known. In Kenya and Tanzania, isolated D. validus populations have different calling cultures and dialects (Roberts et al. 2013). These different calling cultures may be the consequence of isolation; however, the phylogeography of these different populations has not been studied so far. Some groups have counter-calling bouts during the day. This species, or possibly cluster of different species, has a wide repertoire of calls. These aspects, and species’ unresolved taxonomy, calling cultures, meaning of the calls, and possible differences between sexes should be studied further.
Background noise caused by heavy rainfall is likely to modify the calling behavior of many animals (Lengagne and Slater 2002). For example, howler monkeys were acoustically most active during the wet season during the period of maximum fruit production period (Pérez-Granados and Schuchmann 2021a). Vocal activity decreased during wet and rainy days, while temperature had no impact on their vocalization activity. Rain during the previous night delays the beginning of male Bornean gibbons’ (Hylobates muelleri) solo singing bouts (Clink et al. 2020). In the Danum Valley Conservation Area, they sing for 30 min on average. However, rain on the previous night had no impact on duet events, whereas social aspects may be more important. Temperature or lunar cycle also had no impact on the singing in this gibbon species. We found that tree hyraxes in the Taita Hills called significantly less during heavy rain, most probably as heavy rain reduces the acoustic space of tree hyraxes’ vocalizations almost completely.
Animals adjust to background noise in their acoustic signaling in different ways. They may respond to noise by changing signal amplitude, frequency, duration, complexity, or call rate (Kunc and Schmidt 2021). Background noise caused by insects could potentially lead to signal divergence (Luther and Gentry 2013). For example, little greenbul (Andropadus virens) songs occupy different frequencies depending on background noise in different habitats (Slabbekoorn and Smith 2002). In this study we only studied how call rate per minute changed when the Orthoptera choir was active. The Orthoptera choir in Taita Hills is very loud, with frequencies of 5–12 kHz, whereas tree hyrax vocalizations in the Taita Hills use frequencies between 0.5 and 20 kHz. Thus, we expected that tree hyraxes would choose to vocalize more when the orthopterans are silent. In the Taita Hills, orthopterans cease calling after 3am in the morning, leaving free acoustic space. However, we found no evidence that tree hyraxes called more during times when orthopterans were quiet. As these insects use the acoustic space from 5 to up to 12 kHz, the acoustic space under 5 kHz is free for tree hyraxes. The loudest and most complex, and presumably the most important parts of tree hyrax calls are usually between 1000 and 4500 kHz. Thus, their calls are clearly audible to listeners even during a consistent Orthoptera chorus. However, it is possible that tree hyraxes change their call type according to the strength of the Orthoptera choir. We speculate that tree hyraxes could for example increasingly use smaller, and acoustically simpler calls during loud Orthoptera chorus, or they may change the duration of their calls. This needs to be tested in further studies.
According to our observations, Otolemur calls sometimes seem to “wake up” tree hyraxes and activate them to start counter-calling sequences. On the other hand, if Otolemurs are calling very actively, tree hyraxes may be quiet. However, in our study, these two species did not appear to have an obvious acoustic relationship. Both nocturnal mammal species use the same frequencies, but the pitch, and in trailing calls the length, of the calls are very different, and the two species are easy to distinguish acoustically. Moreover, Otolemurs use the same acoustic space, (same frequency at the same time) only occasionally.
In the rock hyrax, singing peaks during the mating season, even though the animals do sing throughout the year (Demartsev et al. 2022). In the Taita Hills, tree hyraxes also sing throughout the year, however, singing is most common in May. Based on this, we expect that May is the primary tree hyrax mating season in the Taita Hills. In May, over 30% of the hours contained singing. The gestation period in tree hyraxes is 7.5 to 8 months (Kingdon 1971), which coincides well with that of the morphologically very similar rock hyraxes (Millar 1971). If the main mating season of the tree hyrax were in May, the births would thus occur in January, which is the summer in Taita Hills, with favorable conditions with respect to food availability and weather stability. Tree hyraxes also sing during rainy weather if the rain is not too heavy. We suggest that singing may be considered as a sign of general well-being, as in the Taita Hills, disturbed tree hyrax populations of small forest fragments rarely sing (Rosti et al. 2022). Similarities in songs of rock hyraxes and tree hyraxes from Taita Hills should be studied further, as such similarities suggest a shared evolutionary path. Other tree hyrax species, the Southern tree hyrax (Dendrohyrax arboreus), the Benin tree hyrax (D. interfluvialis), and the Western tree hyrax (D. dorsalis) do not sing.
Tree hyraxes have been previously described as solitary animals (Kundaeli 1976; Roberts et al. 2013). However, all studied tree hyrax populations in the Taita Hills and at the coast of Kenya have been found to live in groups (Rosti et al. 2020a, b, 2022). Thus, we suggest that the social structure of tree hyrax groups should be studied further. Perhaps the social structure of the tree hyrax groups, at least in the Taita Hills resembles more the social structure of rock hyraxes (Barocas et al. 2011) than previously known.
Summary and conclusions
Acoustic communication of tree hyraxes in the Taita Hills, Kenya, was studied throughout one year using passive acoustic monitoring (PAM). Tree hyraxes vocalized most actively during the relatively dry and warm months of January and February. Calling had a typical, bimodal pattern, with most active calling hours in the early evening between 19.00 and 21.00 and close to dawn between 04.00 and 06.00. Singing activity was highest in the month of May, which may be the main mating season of tree hyraxes in the Taita Hills. We recommend that for estimates of tree hyrax population size using PAM, data from different locations should be collected during the same months, with hours with heavy rain removed, moon phase noted, and its potential effects considered. Results of this study may be used as a baseline for studying other tree hyrax populations, which in many parts of Africa are isolated and decreasing and suffer from habitat loss and poaching.
Data availability
The datasets generated and analyzed during the current study are available in the Zenodo repository. https://doi.org/10.5281/zenodo.7408368.
References
Barocas A, Ilany A, Koren L, Kam M, Geffen E (2011) Variance in centrality within rock hyrax social networks predicts adult longevity. PLoS ONE 6(7):e22375. https://doi.org/10.1371/journal.pone.0022375
Bartholomew GA, Rainy M (1971) Regulation of body temperature in the rock hyrax, Heterohyrax brucei. J Mammal 52:81–95. https://doi.org/10.2307/1378434
Bedoya C, Isaza C, Daza JM, López JD (2017) Automatic identification of rainfall in acoustic recordings. Ecol Indic 75:95–100. https://doi.org/10.1016/j.ecolind.2016.12.018
Bettridge CM, Kenworthy SP, Butynski TM, De Jong YA, de Kort SR (2019) Vocal repertoire and intraspecific variation within two loud calls of the small-eared greater galago (Otolemur garnettii) in Tanzania and Kenya. Folia Primatol 90:319–335. https://doi.org/10.1159/000500260
Birkett PJ, Vanak AT, Muggeo VMR, Ferreira SM, Slotow R (2012) Animal perception of seasonal thresholds: changes in elephant movement in relation to rainfall patterns. PLoS ONE 7(6):e38363. https://doi.org/10.1371/journal.pone.0038363
Boquimpani-Freitas L, Marra RV, Sluys MV, Rocha CFD (2007) Temporal niche of acoustic activity in anurans: interspecific and seasonal variation in a neotropical assemblage from south-eastern Brazil. Amphibia-Reptilia 28:269–276. https://doi.org/10.1163/156853807780202422
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Mächler M, Bolker BM (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9(2):378. https://doi.org/10.32614/RJ-2017-066
Brown KJ, Downs CT (2006) Basking behaviour in the rock hyrax (Procavia capensis) during winter. Afr Zool 42:70–79. https://doi.org/10.1080/15627020.2007.11407379
Brown KJ, Downs CT (2007) Seasonal patterns in body temperature of free-living rock hyrax (Procavia capensis). Comp Biochem Physiol A 143:42–49. https://doi.org/10.1016/j.cbpa.2005.10.020
Brumm H (2006) Signalling through acoustic windows: nightingales avoid interspecific competition by short-term adjustment of song timing. J Comp Physiol A 192:1279–1285. https://doi.org/10.1007/s00359-006-0158-x
Burgess ND, Butynski TM, Cordeiro NJ, Doggart NH, Fjeldså J, Howell KM, Kilahama FB, Loader SP, Lovett JC, Mbilinyi B, Menegon M, Moyer DC, Nashanda E, Perkin AW, Rovero F, Stanley WT, Stuart SN (2007) The biological importance of the Eastern Arc Mountains of Tanzania and Kenya. Biol Conserv 134:209–231. https://doi.org/10.1016/j.biocon.2006.08.015
Butynski TM, De Jong YA, Perkin AW, Bearder SK, Honess PE (2006) Taxonomy, distribution, and conservation status of three species of dwarf galagos (Galagoides) in eastern Africa. Primate Conserv 21:63–79. https://doi.org/10.1896/0898-6207.21.1.63
Catchpole C, Slater PJB (2008) Bird song: biological themes and variations, 2nd edn. Cambridge University Press, Cambridge and New York
Clink DJ, Hamid Ahmad A, Klinck H (2020) Gibbons aren’t singing in the rain: presence and amount of rainfall influences ape calling behavior in Sabah. Malaysia Sci Rep 10(1):1282. https://doi.org/10.1038/s41598-020-57976-x
Demartsev V, Kershenbaum A, Ilany A, Barocas A, Bar Ziv E, Koren L, Geffen E (2014) Male hyraxes increase song complexity and duration in the presence of alert individuals. Behav Ecol 25:1451–1458. https://doi.org/10.1093/beheco/aru155
Demartsev V, Ilany A, Kershenbaum A, Geva Y, Margalit O, Schnitzer I, Barocas A, Bar-Ziv E, Koren L, Geffen E (2017) The progression pattern of male hyrax songs and the role of climactic ending. Sci Rep 7(1):2794. https://doi.org/10.1038/s41598-017-03035-x
Demartsev V, Haddas-Sasson M, Ilany A, Koren L, Geffen E (2022) Male rock hyraxes that maintain an isochronous song rhythm achieve higher reproductive success. J Anim Ecol 1365–2656:13801. https://doi.org/10.1111/1365-2656.13801
Djossa B, Zachee B, Sinzin B (2012) Activity patterns and habitat use of the western tree hyrax (Dendrohyrax dorsalis), within forest patches and implications for conservation. Ecotropica 18:65–72
Egnor SER, Wickelgren JG, Hauser MD (2007) Tracking silence: adjusting vocal production to avoid acoustic interference. J Comp Physiol A 193:477–483. https://doi.org/10.1007/s00359-006-0205-7
Ellinger N, Hödl W (2003) Habitat acoustics of a neotropical lowland rainforest. Bioacoustics 13:297–321. https://doi.org/10.1080/09524622.2003.9753503
Erb WM, Ziegler T, Lestari NS, Hammerschmidt K (2016) Are simakobu (Simias concolor) loud calls energetically costly signals? Am J Phys Anthropol 161:44–52. https://doi.org/10.1002/ajpa.23005
Español-Jiménez S, van der Schaar M (2018) First record of humpback whale songs in southern Chile: analysis of seasonal and diel variation. Mar Mamm Sci 34:718–733. https://doi.org/10.1111/mms.12477
Fernandez-Duque E (2003) Influences of moonlight, ambient temperature, and food availability on the diurnal and nocturnal activity of owl monkeys (Aotus azarai). Behav Ecol Sociobiol 54:431–440. https://doi.org/10.1007/s00265-003-0637-9
Ferroudj M, Truskinger A, Towsey M, Zhang L, Zhang J, Roe P (2014) Detection of rain in acoustic recordings of the environment. In: Pham DN, Park SB (eds) PRICAI 2014: Trends in Artificial Intelligence. PRICAI 2014. Lecture Notes in Computer Science, vol 8862. Springer, Cham, pp 104–116. https://doi.org/10.1007/978-3-319-13560-1_9
Fey V (1960) A note on behavior of the tree hyrax. J East Afr Nat Hist Soc 23:244–246
Frouin-Mouy H, Mouy X, Martin B, Hannay D (2016) Underwater acoustic behavior of bearded seals (Erignathus barbatus) in the northeastern Chukchi Sea, 2007–2010. Mar Mamm Sci 32:141–160. https://doi.org/10.1111/mms.12246
Gaylard A, Kerley GIH (2001) Habitat assessment for a rare, arboreal forest mammal, the tree hyrax (Dendrohyrax arboreus). Afr J Ecol 39:205–212. https://doi.org/10.1046/j.0141-6707.2000.301.x
Gereau RE, Cumberlidge N, Hemp C, Hochkirch A, Jones T, Kariuki M, Lange CN, Loader SP, Malonza PK, Menegon M, Ndang’ang’a PK, Rovero F, Shirk P (2016) Globally threatened biodiversity of the Eastern Arc Mountains and coastal forests of Kenya and Tanzania. J East Afr Nat Hist 105:115–201. https://doi.org/10.2982/028.105.0104
Githiru M, Lens L, Adriaensen F, Mwang’ombe J, Matthysen E, (2011) Using science to guide conservation: From landscape modelling to increased connectivity in the Taita Hills. SE Kenya J Nat Conserv 19(5):263–268. https://doi.org/10.1016/j.jnc.2011.03.002
Glover TD, Sale JB (1968) The reproductive system of male rock hyrax (Procavia and Heterohyrax). J Zool 156(3):351–362. https://doi.org/10.1111/j.1469-7998.1968.tb04358.x
Gottsberger B, Gruber E (2004) Temporal partitioning of reproductive activity in a neotropical anuran community. J Trop Ecol 20:271–280. https://doi.org/10.1017/S0266467403001172
Gursky S (2003) Lunar philia in a nocturnal primate. Int J Primatol 24:351–367. https://doi.org/10.1023/A:1023053301059
Gutman R, Dayan T, Levy O, Schubert I, Kronfeld-Schor N (2011) The effect of the lunar cycle on fecal cortisol metabolite levels and foraging ecology of nocturnally and diurnally active spiny mice. PLoS ONE 6(8):e23446. https://doi.org/10.1371/journal.pone.0023446
Harcourt CS, Perkin AW (2013) Galagoides cocos – Kenya coast dwarf galago (Diani galago). In: Butynski TM, Kingdon J, Kalina J (eds) Mammals of Africa. Volume II Primates. Bloomsbury Publishing, London, pp 457–459
Hart PJ, Hall R, Ray W, Beck A, Zook J (2015) Cicadas impact bird communication in a noisy tropical rainforest. Behav Ecol 26:839–842. https://doi.org/10.1093/beheco/arv018
Hartig F (2022) DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. R package ver. 0.4.6 http://florianhartig.github.io/DHARMa/
Hemp C (2021) A field guide to the bushcrickets, wetas and raspy crickets of Tanzania and Kenya. Senckenberg Gesellschaft für Naturforschung, Frankfurt am Main
Hill AP, Prince P, Piña Covarrubias E, Doncaster CP, Snaddon JL, Rogers A (2018) AudioMoth: evaluation of a smart open acoustic device for monitoring biodiversity and the environment. Methods Ecol Evol 9:1199–1211. https://doi.org/10.1111/2041-210X.12955
Hoeck HN (1975) Differential feeding behaviour of the sympatric hyrax Procavia johnstoni and Heterohyrax brucei. Oecologia 22:15–47. https://doi.org/10.1007/BF00345256
Julien-Laferriere D (1997) The influence of moonlight on activity of woolly opossums (Caluromys philander). J Mammal 78:251–255. https://doi.org/10.2307/1382659
Keast JA, Marshall AJ (1954) The influence of drought and rainfall on reproduction in Australian desert birds. Proc Zool Soc Lond 124:493–499. https://doi.org/10.1111/j.14697998.1954.tb07790.x
Kingdon J (1971) East African mammals: an atlas of evolution in Africa, vol I. Academic Press, London and New York
Kloubec B, Čapek M (2005) Seasonal and diel budgets of song: a study of Savi’s warbler (Locustella luscinioides). J Ornithol 146:206–214. https://doi.org/10.1007/s10336-005-0081-x
Koren L (2006) Vocalization as an indicator of individual quality in the rock hyrax. Dissertation, Tel-Aviv University
Koren L, Geffen E (2011) Individual identity is communicated through multiple pathways in male rock hyrax (Procavia capensis) songs. Behav Ecol Sociobiol 65:675–684. https://doi.org/10.1007/s00265-010-1069-y
Krause B (1987) The niche hypothesis: how animals taught us to dance and sing. Whole Earth Rev 57(57):14–16
Kronfeld-Schor N, Dominoni D, de la Iglesia H, Levy O, Herzog ED, Dayan T, Helfrich-Forster C (2013) Chronobiology by moonlight. Proc R Soc Lond B 80:20123088. https://doi.org/10.1098/rspb.2012.3088
Kunc HP, Wolf JBW (2008) Seasonal changes of vocal rates and their relation to territorial status in male Galápagos sea lions (Zalophus wollebaeki). Ethology 114:381–388. https://doi.org/10.1111/j.1439-0310.2008.01484.x
Kunc HP, Schmidt R (2021) Species sensitivities to a global pollutant: a meta-analysis on acoustic signals in response to anthropogenic noise. Glob Change Biol 27:675–688. https://doi.org/10.1111/gcb.15428
Kundaeli JN (1976) Distribution of tree hyrax (Dendrohyrax validus validus True) on Mt Kilimanjaro, Tanzania. Afr J Ecol 14:253–264. https://doi.org/10.1111/j.1365-2028.1976.tb00241.x
Lack D (1950) Breeding seasons in the Galapagos. Ibis 92:268–278. https://doi.org/10.1111/j.1474-919X.1950.tb01751.x
Lawson JL (2022) Listening in on the forest: use of bioacoustics to preserve soundscapes and rare species. Dissertation, Imperial College London. https://doi.org/10.25560/96746
Lengagne T, Slater PJB (2002) The effects of rain on acoustic communication: tawny owls have good reason for calling less in wet weather. Proc R Soc Lond B 269:2121–2125. https://doi.org/10.1098/rspb.2002.2115
Lüdecke D, Ben-Shachar M, Patil I, Waggoner P, Makowski D (2021) Performance: an R package for assessment, comparison and testing of statistical models. JOSS. 6:3139. https://doi.org/10.21105/joss.03139
Luther D, Gentry K (2013) Sources of background noise and their influence on vertebrate acoustic communication. Behaviour 150:1045–1068. https://doi.org/10.1163/1568539X-00003054
Marques TA, Thomas L, Martin SW, Mellinger DK, Ward JA, Moretti DJ, Harris D, Tyack PL (2013) Estimating animal population density using passive acoustics. Biol Rev 88:287–309. https://doi.org/10.1111/brv.12001
McElligott AG, O’Neill KP, Hayden TJ (1999) Cumulative long-term investment in vocalization and mating success of fallow bucks, Dama dama. Anim Behav 57:1159–1167. https://doi.org/10.1006/anbe.1999.1076
Millar RP (1971) Reproduction in the rock hyrax Procavia capensis. Afr Zool 6:243– 261. https://doi.org/10.10520/AJA00445096_203
Milner JM, Harris S (1999a) Activity patterns and feeding behaviour of the tree hyrax, Dendrohyrax arboreus, in the Parc National des Volcans, Rwanda. Afr J Ecol 37:267–280. https://doi.org/10.1046/j.1365-2028.1999.00184.x
Milner JM, Harris S (1999b) Habitat use and ranging behaviour of tree hyrax, Dendrohyrax arboreus, in the Virunga Volcanoes, Rwanda: Habitat use by tree hyrax. Afr J Ecol 37:281–294. https://doi.org/10.1046/j.1365-2028.1999.00185.x
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858. https://doi.org/10.1038/35002501
Narganes Homfeldt T, Risch D, Stevenson A, Henry L-A (2022) Seasonal and diel patterns in singing activity of humpback whales migrating through Bermuda. Front Mar Sci 9:941793. https://doi.org/10.3389/fmars.2022.941793
Nash LT (2007) Moonlight and behavior in nocturnal and cathemeral primates, especially lepilemur leucopus: illuminating possible anti-predator efforts. In: Gursky SL, Nekaris KAI (eds) Primate Anti-predator strategies. Springer US, Boston, MA, pp 173–205. https://doi.org/10.1007/978-0-387-34810-0_8
Nelson DV, Garcia TS, Klinck H (2017) Seasonal and diel vocal behavior of the northern red-legged frog, Rana aurora. Northwest Nat 98:33–38. https://doi.org/10.1898/NWN16-06.1
Nishihara H, Satta Y, Nikaido M, Thewissen JGM, Stanhope MJ, Okada N (2005) A retroposon analysis of afrotherian phylogeny. Mol Biol Evol 22:1823–1833
Oates JF, Woodman N, Gaubert P, Sargis EJ, Wiafe ED, Lecompte E, Dowsett-Lemaire F, Dowsett RJ, Gonedelé Bi S, Ikemeh RA, Djagoun CA, Tomsett L, Bearder SK (2022) A new species of tree hyrax (Procaviidae: Dendrohyrax) from West Africa and the significance of the Niger-Volta interfluvium in mammalian biogeography. Zool J Linn Soc 194:527–552. https://doi.org/10.1093/zoolinnean/zlab029
Opperman EJ, Cherry MI, Makunga NP (2018) Community harvesting of trees used as dens and for food by the tree hyrax (Dendrohyrax arboreus) in the Pirie forest. South Africa. Koedoe 60:a1481. https://doi.org/10.4102/koedoe.v60i1.1481
Pereyra LC, Akmentins MS, Sanabria EA, Vaira M (2016) Diurnal? Calling activity patterns reveal nocturnal habits in the aposematic toad Melanophryniscus rubriventris. Can J Zool 94:497–503. https://doi.org/10.1139/cjz-2015-0197
Pérez-Granados C, Schuchmann K-L (2020) Diel and seasonal variations of vocal behavior of the neotropical white-tipped dove (Leptotila verreauxi). Diversity 12(10):402. https://doi.org/10.3390/d12100402
Pérez-Granados C, Schuchmann K-L (2021a) Passive acoustic monitoring of chaco chachalaca (Ortalis canicollis) over a year: vocal activity pattern and monitoring recommendations. Trop Conserv Sci 14:194008292110582. https://doi.org/10.1177/19400829211058295
Pérez-Granados C, Schuchmann K (2021b) Passive acoustic monitoring of the diel and annual vocal behavior of the black and gold howler monkey. Am J Primatol 83:e23241. https://doi.org/10.1002/ajp.23241
Pérez-Granados C, Schuchmann K-L, Marques MI (2022) Addicted to the moon: vocal output and diel pattern of vocal activity in two Neotropical nightjars is related to moon phase. Ethol Ecol Evol 34:66–81. https://doi.org/10.1080/03949370.2021.1886182
Pihlström H, Rosti H, Lombo B, Pellikka P (2021) Domestic dog predation on white-tailed small-eared galago (Otolemur garnettii lasiotis) in the Taita Hills, Kenya. Afr Primates 15:31–38
Pitcher BJ, Briefer EF, Vannoni E, McElligott AG (2014) Fallow bucks attend to vocal cues of motivation and fatigue. Behav Ecol 25:392–401. https://doi.org/10.1093/beheco/art131
Pratas-Santiago LP, Gonçalves ALS, Nogueira AJA, Spironello WR (2017) Dodging the moon: the moon effect on activity allocation of prey in the presence of predators. Ethology 123:467–474. https://doi.org/10.1111/eth.12617
Prugh LR, Golden CD (2014) Does moonlight increase predation risk? Meta-analysis reveals divergent responses of nocturnal mammals to lunar cycles. J Anim Ecol 83(2):504–514. https://doi.org/10.1111/1365-2656.12148
R Core Team (2020) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. URL https://www.R-project.org/.
Risch D, Clark C, Dugan P, Popescu M, Siebert U, Van Parijs S (2013) Minke whale acoustic behavior and multi-year seasonal and diel vocalization patterns in Massachusetts Bay, USA. Mar Ecol Prog Ser 489:279–295. https://doi.org/10.3354/meps10426
Roberts D, Topp-Jørgensen E, Moyer D (2013) Dendrohyrax validus – Eastern tree hyrax. In: Kingdon J, Happold D, Hoffmann M, Butynski T, Happold M, Kalina J (eds) Mammals of Africa. Volume 1: introductory chapters and Afrotheria. Bloomsbury Publishing, London, pp 158–161
Rosti H, Pihlström H, Bearder S, Pellikka P, Rikkinen J (2020a) Vocalization analyses of nocturnal arboreal mammals of the Taita Hills, Kenya. Diversity 12(12):473. https://doi.org/10.3390/d12120473
Rosti H, Rikkinen J, Pellikka P, Bearder S, Mwamodenyi JM (2020b) Taita Mountain dwarf galago is extant in the Taita Hills of Kenya. Oryx 54:152–153. https://doi.org/10.1017/S003060531900142X
Rosti H, Heiskanen J, Loehr J, Pihlström H, Bearder S, Mwangala L, Maghenda M, Pellikka P, Rikkinen J (2022) Habitat preferences, estimated abundance and behavior of tree hyrax (Dendrohyrax sp.) in fragmented montane forests of Taita Hills, Kenya. Sci Rep 12:6331. https://doi.org/10.1038/s41598-022-10235-7
Rosti H, Pihlström H, Rottcher N, Bearder S, Mwangala L, Maghenda M, Rikkinen J (2023) Species identity and behavior of cave-dwelling tree hyraxes of the Kenyan coast. Ecol Evol 13:e9693. https://doi.org/10.1002/ece3.9693
Rouget PA, Terhune JM, Burton HR (2007) Weddell seal underwater calling rates during the winter and spring near Mawson Station, Antarctica. Mar Mamm Sci 23:508–523. https://doi.org/10.1111/j.1748-7692.2007.00129.x
Rstudio Team (2020) Rstudio: Integrated Development for R. Rstudio. PBC, Boston. http://www.rstudio.com
Sale J (1970) The behaviour of the resting rock hyrax in relation to its environment. Zool Afr 5:87–99
Sánchez-Giraldo C, Bedoya CL, Morán-Vásquez RA, Isaza CV, Daza JM (2020) Ecoacoustics in the rain: understanding acoustic indices under the most common geophonic source in tropical rainforests. Remote Sens Ecol Conserv 6(248):261. https://doi.org/10.1002/rse2.162
Sandoval L, Méndez C, Mennill DJ (2016) Vocal behaviour of white-eared ground-sparrows (Melozone leucotis) during the breeding season: repertoires, diel variation, behavioural contexts, and individual distinctiveness. J Ornithol 157:1–12. https://doi.org/10.1007/s10336-015-1237-y
Seibt U, Hoeck H, Wickler W (1977) Dendrohyrax validus True, 1890 in Kenia. Zeitschr Säugetierkd 42:115–118
Slabbekoorn H, Smith TB (2002) Habitat-dependent song divergence in the little greenbul: an analysis of environmental selection pressures on acoustic signals. Evolution 56:1849–1858
Springer MS, Cleven GC, Madsen O, de Jong WW, Waddell VG, Amrine HM, Stanhope MJ (1997) Endemic African mammals shake the phylogenetic tree. Nature 388:61–64
Staicer CA, Spector DA, Horn AG (2020) The dawn chorus and other diel patterns in acoustic signaling. In: Kroodsma DE, Miller EH (eds) Ecology and evolution of acoustic communication in birds. Cornell University Press, Ithaca, pp 426–453. https://doi.org/10.7591/9781501736957-033
Stanhope MJ, Madsen O, Waddell VG, Cleven GC, de Jong WW, Springer MS (1998) Highly congruent molecular support for a diverse superordinal clade of endemic African mammals. Mol Phylogenet Evol 9:501–508
Stanley CQ, Walter MH, Venkatraman MX, Wilkinson GS (2016) Insect noise avoidance in the dawn chorus of Neotropical birds. Anim Behav 112:255–265. https://doi.org/10.1016/j.anbehav.2015.12.003
Starr C, Nekaris KAI, Leung L (2012) Hiding from the moonlight: luminosity and temperature affect activity of Asian nocturnal primates in a highly seasonal forest. PLoS ONE 7(4):e36396. https://doi.org/10.1371/journal.pone.0036396
Studd EK, Boudreau MR, Majchrzak YN, Menzies AK, Peers MJL, Seguin JL, Lavergne SG, Boonstra R, Murray DL, Boutin S, Humphries MM (2019) Use of acceleration and acoustics to classify behavior, generate time budgets, and evaluate responses to moonlight in free-ranging snowshoe hares. Front Ecol Evol 7:154. https://doi.org/10.3389/fevo.2019.00154
Tabuce R, Asher RJ, Lehmann T (2008) Afrotherian mammals: a review of current data. Mammalia 72:2–14
Teucher M, Schmitt CB, Wiese A, Apfelbeck B, Maghenda M, Pellikka P, Lens L, Habel JC (2020) Behind the fog: forest degradation despite logging bans in an East African cloud forest. Global Ecol Conserv 22:e01024. https://doi.org/10.1016/j.gecco.2020.e01024
Thijs KW (2015) Tree community dynamics and ecosystem function in a tropical landscape under deforestation pressure. Dissertation, KU Leuven.
Thompson ME, Schwager SJ, Payne KB, Turkalo AK (2009) Acoustic estimation of wildlife abundance: methodology for vocal mammals in forested habitats. Afr J Ecol 48:654–661. https://doi.org/10.1111/j.1365-2028.2009.01161.x
Topp-Jørgensen JE, Marshal AR, Brink H, Pedersen UB (2008) Quantifying the response of tree hyraxes (Dendrohyrax validus) to human disturbance in the Udzungwa Mountains, Tanzania. Trop Conserv Sci 1:63–74. https://doi.org/10.1177/194008290800100106
Van Opzeeland I, Van Parijs S, Bornemann H, Frickenhaus S, Kindermann L, Klinck H, Plötz J, Boebel O (2010) Acoustic ecology of Antarctic pinnipeds. Mar Ecol Prog Ser 414:267–291. https://doi.org/10.3354/meps08683
Walsh PB, Inglis JM (1989) Seasonal and diel rate of spontaneous vocalization in coyotes in South Texas. J Mammal 70:169–171. https://doi.org/10.2307/1381681
Wekesa C, Kirui BK, Maranga EK, Muturi GM (2020) The fate of Taita Hills forest fragments: evaluation of forest cover change between 1973 and 2016 using Landsat imagery. Open J for 10:22–38
Wickham H (2016) ggplot2: elegant graphics for data analysis, 2nd edn. Springer, Cham. https://doi.org/10.1007/978-3-319-24277-4
Wild J, Kopecký M, Macek M, Šanda M, Jankovec J, Haase T (2019) Climate at ecologically relevant scales: a new temperature and soil moisture logger for long-term microclimate measurement. Agricult Forest Meteorol 268:40–47. https://doi.org/10.1016/j.agrformet.2018.12.018
Wilkins MR, Seddon N, Safran RJ (2013) Evolutionary divergence in acoustic signals: causes and consequences. Trends Ecol Evol 28:156–166. https://doi.org/10.1016/j.tree.2012.10.002
Wilson DR, Ratcliffe LM, Mennill DJ (2016) Black-capped chickadees, Poecile atricapillus, avoid song overlapping: evidence for the acoustic interference hypothesis. Anim Behav 114:219–229. https://doi.org/10.1016/j.anbehav.2016.02.002
Zhao H, Zhong Y, Wang X, Hu X, Luo C, Boitt M, Piiroinen R, Zhang L, Heiskanen J, Pellikka P (2022) Mapping the distribution of invasive tree species using deep one-class classification in the tropical montane landscape of Kenya. ISPRS J Photogramm Remote Sens 187:328–344. https://doi.org/10.1016/j.isprsjprs.2022.03.005
Zina J, Haddad CFB (2005) Reproductive activity and vocalizations of Leptodactylus labyrinthicus (Anura: Leptodactylidae) in southeastern Brazil. Biota Neotrop 5:119–129. https://doi.org/10.1590/S1676-06032005000300008
Zuur AF, Ieno EN (2016) A protocol for conducting and presenting results of regression-type analyses. Methods Ecol Evol 7:636–645. https://doi.org/10.1111/2041-21
Zuur AF, Saveliev AA, Ieno EN (2012) Zero inflated models and generalized linear mixed models with R. Reprinted with corrections. Highland Statistics, Newburgh, United Kingdom
Acknowledgements
We wish to thank the helpful staff of the University of Helsinki’s Taita Research Station, and especially field assistant Benson Lombo for their support. The research was kindly permitted by the National Council for Science and Technology of Kenya NACOSTI/P/21/9300 and the Kenya Forest Service KFS/TTFS/7/1/4/Vol.II/14. We are also very grateful to reviewer Hansjoerg Kunc, Editor in Chief Heiko G. Rödel, and one anonymous referee, who greatly improved the manuscript.
Funding
Open Access funding provided by University of Helsinki including Helsinki University Central Hospital. Author HR was supported by Kone Foundation grant 202007604, Tuovinen foundation, Nordenskiöld-samfundet, Otto A Malm Foundation, Ripaco and by the University of Helsinki. Authors PP and MR were supported by ESSA project funded by the DeSIRA program of European Commission, DG International Partnerships (FOOD/2020/418-132). Author HP was supported by grants from the Oskar Öflund Foundation and the Waldemar von Frenckell Foundation.
Author information
Authors and Affiliations
Contributions
Conceptualization: HR, JR, SB; methodology: HR, EM, MR; formal analysis and investigation: HR, JL; SB; JR; writing—original draft preparation: HR, HP, JL; writing—review and editing: HR, HP, MM, LM, JR, SB; funding acquisition: HR; resources: HR, JR, PP, MM, LM; supervision: JR, SB, PP.
Corresponding author
Ethics declarations
Conflict of interest
There are no competing interests with this study.
Ethical approval
The ethical approval was not required for this study, because we did not use any invasive method that could harm or disturb animals.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling editor: Heiko G. Rödel.
Supplementary Information
Below is the link to the electronic supplementary material.
42991_2023_370_MOESM1_ESM.tif
Supplementary file1 Spectrograms of Taita tree hyrax calls. a) Strangled thwack is broadband call; frequency of the call is from 0 to > 20 kHz. Frequency modulated part is between 1-6 kHz, this gives the call striking pitch (Online Resource 1). b) One example variation of strangled thwack (Online Resource 2). c) Counter-calling sequence of strangled thwacks with several individuals (Online Resource 3). d) Hac call is simple broadband call from 0 to > 20 kHz (Online Resource 4). e) One example variation of hac call (Online Resource 5). f) Calling sequence of hac calls by one individual (Online Resource 6). g) Wheeze calls, fundamental frequency in wheeze is typically between 3-4 kHz, and call may last more than one second. Call has several harmonics. This call may be used alone, as a warning, or it may be part of a song (Online Resource 7). h) Calling sequence of wheezes by one animal (Online Resource 8) (TIF 59942 KB)
42991_2023_370_MOESM2_ESM.tif
Supplementary file2 Spectrogram sequence of tree hyrax song showing different syllables (Online Resource 9) (TIF 12742 KB)

42991_2023_370_MOESM3_ESM.jpg
Supplementary file3 Spectrograms illustrating different types and levels of background noise recorded from Ngangao Forest, Kenya. (a) Rainfall Level 1, no or little rain (Online Resource 10). (b) Rainfall Level 2 reaching 4 kHz and above, caused by moderate rain (Online Resource 11). (c) Rainfall Level 2, reaching above 6 kHz, caused by heavy rain (Online Resource 12). (d) Orthoptera Choir Level 1, free acoustic space with no Orthoptera Choir (Online Resource 13). (e) Orthoptera Choir Level 2, discontinuous Orthoptera Choir using acoustic space between 5–12 kHz (Online Resource 14). (f) Orthopteran Chorus Level 3, continuous Orthopteran Choir filling acoustic space between 5–12 kHz (Online Resource 15) (JPG 3645 KB)
42991_2023_370_MOESM4_ESM.tif
Supplementary file4 Spectrograms of Otolemur calls. (a) Trailing call and (b) cluster squawk calls (Online Resource 16). The trailing call is likely an advertisement or contact call, while the cluster squawk probably represents a distress or alarm call (Bettridge et al. 2019). Photo: Hanna Rosti (TIF 31531 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
Cite this article
Rosti, H., Pihlström, H., Bearder, S. et al. Seasonal and diel variations in the acoustic communication of tree hyraxes in Taita Hills, Kenya. Mamm Biol 104, 1–14 (2024). https://doi.org/10.1007/s42991-023-00370-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-023-00370-7