
Abstract
The risk of large carnivore mortality from anthropogenic sources is increased in areas where frequent human-carnivore encounters are high. Translocation is a potential tool for reducing these conflicts and re-establishing wild populations. Determining proper translocation sites for tigers is important for securing their future. In this research, we quantitatively evaluated the habitat suitability of a potential tiger translocation area on the Kampar Peninsula, a large peatland landscape in Sumatra. This landscape is essential for the conservation of the local tiger population, but its role as a tiger habitat has rarely been investigated. We conducted this research in a region with mosaicked land-use types covering an area of 7,045.89 km2. We applied species ecological niche factor analysis (ENFA) to model habitat suitability using species presence and ecogeographical variables as the bases. The model predicted that approximately 60% of the study area is tiger-suitable habitat with varying suitability levels. The tiger habitat characteristics in the study region are dissimilar to the global tiger habitat characteristics, and the high specialization value obtained indicates that tigers inhabit very narrow niches. Considering the low density of tigers in peatlands, we estimate that the carrying capacity of the whole Kampar landscape is approximately 13 tigers. Therefore, to conserve a viable tiger population, it is necessary to maintain connectivity between the Kampar peatlands and the Kerumutan landscape situated to the south of the Kampar Peninsula.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Large carnivore translocation is an essential tool for mitigating human–wildlife conflicts caused by livestock depredation or carnivores entering human settlements or agriculture areas, leading to animal mortality (Priatna et al. 2012). With the expansion of agricultural lands and the high rate of primary forest loss in Indonesia (Brun et al. 2015; Margono et al. 2014), the competition between carnivores and humans over space and food are expected to intensify and further engender human-animal conflicts (Karanth and Chellam 2009). In Sumatra, conflicts between humans and endangered Sumatran tigers (Panthera tigris sumatrae) are more frequent in intermediate disturbed landscapes such as secondary forests, along the boundaries of protected areas, and in multiple-use forestlands where humans and tigers coexist; in contrast, these conflicts are less common in protected areas (Nyhus and Tilson 2004). To prevent the killing of tigers by local people, animals living in small and fragmented habitats should be translocated to safe habitats. Recently, the urgency with which tiger translocation should be performed has also been triggered by the need to release a tiger that was rescued from a wire snare set by poachers. The official website of the Ministry of Environment and Forestry (2020) published a press conference about the release of a snared female tiger named Corina in Kampar Forest after 8 months in care in a tiger rehabilitation centre (http://ppid.menlhk.go.id/berita/siaran-pers/5752/harimau-sumatera-corina-pulang-kampung). Recent data have shown that, due to impressive economic growth in Asia, the market demand for endangered species such as the Sunda pangolin and tiger has increased. The annual poaching rate is strongly and positively correlated with the increase in the price of tiger skin at local markets (Linkie et al. 2018).
Translocating tigers would also support the goal of doubling the global tiger population by 2022; this goal was set by 13 countries with wild tigers, including Indonesia (The World Bank and BPS 2019). In the last survey, it was found that Sumatran tigers are present in 27 habitat patches larger than 250 km2 and possibly in two other habitat patches (Wibisono and Pusparini 2010). The island-wide tiger population size declined from the year 2000 (742 individuals) to 2012 (618 individuals), exhibiting an approximately 16.6% decline in Sumatra. Tiger occupancy varies substantially among different landscapes. The highest occupancy was recorded in the large Kerinci Seblat-Batang Hari landscape (13,971 km2), while lower occupancies have been observed in smaller landscapes such as Way Kambas National Park (1300 km2). The lowest occupancy has been found in the Northern Riau landscape, which has undergone a high deforestation rate and is dominated by peatland ecosystems (Wibisono et al. 2011). The Sumatran tiger is still facing a high risk of extinction, as indicated by the presence of only two robust populations with more than 30 breeding females in Kerinci Seblat and Leuser National Park (Luskin et al. 2017).
The success of animal translocation depends on many factors, such as the mortality of animals due to natural predators, poaching, intra- and interspecific competition, and the habitat quality of the release sites (Cheyne 2006). Sufficient knowledge of the factors determining “good” habitat quality is a prerequisite for translocation success, especially for endangered species (Cook et al. 2010; Hirzel et al. 2004). The survival rate of translocated cheetahs (Acynonyx jubatus) in Botswana was low due to their homing behaviours and wide-ranging movements post release (Boast et al. 2016). A similar result was also reported following the translocation of Amur tigers (Goodrich and Miquelle 2005). In these cases, conflict mitigation measures that promote the coexistence of predators and humans might have been a more effective tool than translocation. Actively involving the local community in human–wildlife conservation aids in the development of strong social capital, which is helpful for managing large carnivore species (Morehouse et al. 2020). Soft release protocols result in better translocation outcomes than hard release programmes (Resende et al. 2020).
Constructing a reliable predictive model that explains the relationships between tigers and habitat characteristics is an essential step before a translocation programme can be implemented. However, researching large carnivores such as tigers, which are rare, exhibit elusive behaviours and occupy extensive home ranges, is a demanding task. Previous studies have shown that, at the landscape level, Sumatran tigers prefer forests over acacia and oil palm plantations, and the probability of tiger occupancy is positively correlated with the elevation and forest area and negatively correlated with the distance from the forest centroid (Sunarto et al. 2012). The tiger population density is 47% higher in primary forests than in degraded forests and 49.9% lower in peatlands and montane regions than in lowlands and hilly forests (Luskin et al. 2017). Although peatland landscapes occupy a vast area in Sumatra (approximately 15% of the total area; 61,400 km2) (Miettinen and Liew 2010) and some peatland areas cover the Tiger Conservation Landscape (TCL), only very few studies on tigers have been conducted in this ecosystem type (Sunarto et al. 2012, 2013, 2015).
Sumatra peatlands have experienced rapid land cover changes over recent decades as they have been converted from forests to managed land cover types, such as industrial plantations and smallholder agricultural lands (Miettinen et al. 2016). This study was conducted in a large peatland landscape on the Kampar Peninsula in Riau Province, where a tiger population still exists (Luskin et al. 2017; Sunarto et al. 2013, 2015). The study area is a mosaic of peatland landscapes that constitute protected areas, industrial and smallholder plantations, and peatland ecosystem restoration concessions (Restorasi Ekosistem Riau/RER) managed by plantation companies. The RER was established with the aim of protecting, restoring and conserving ecologically important peatland ecosystems as well as to fulfil the commitments of the plantation management companies to restore and conserve one-hectare of natural forest for every hectare of fibre plantation.
The Kampar Peninsula might be the largest peat swamp landscape remaining in Sumatra (Goenarto and Gunaryadi 2018) and has been regarded as a regional priority landscape for tiger conservation with a moderate probability of long-term tiger population persistence (Sanderson et al. 2006). However, no accurate estimation of the tiger density in this area has been obtained. A previous study conducted with camera traps from July to November 2007 failed to capture a single picture of a tiger, suggesting that human disturbances are likely very high in this area, as indicated by the high number of camera traps lost: among 20 camera trap stations, 16 (80%) disappeared (Sunarto et al. 2013). The tiger occupancy probability determined based on the tiger presence signs varied greatly among the surveyed grids. Most grids had zero values, while others had values ranging from 0.18 to 0.56 (Sunarto et al. 2012).
In this study, we aimed to assess the suitability of the Kampar Peninsula peatland landscape for a tiger population. The results of this study will answer the question of whether the relatively low tiger occupancy probability determined in previous studies is related to habitat suitability (Luskin et al. 2017). Furthermore, the findings of this work will also provide a scientific basis for whether the study area is suitable for tiger translocation or reintroduction if human activities such as poaching and illegal logging can be controlled. Evaluations of proposed tiger restoration sites constitute an urgent need in Sumatra, considering the high rate of human-tiger conflicts, poaching with wire snares and the increasing number of problem tigers that must be rescued and relocated (Priatna et al. 2012).
Study site
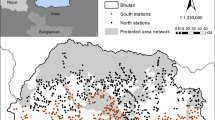
This study was conducted in the peatland landscape on the Kampar Peninsula along the eastern coastline of Sumatra Island, Indonesia; this region encompasses a 7045.89 km2 area that includes the Siak and Kampar Rivers and the Panjang Strait (Fig. 1). The central part of the area is covered by lowland peat swamp forests with some lakes upstream of the small inland rivers. Although inland rivers drain the area, most of the Kampar Peninsula is constantly saturated with freshwater due to the flat terrain and tropical rainfall. Dry seasons commonly occur from late January to mid-March and again from June to September. The average annual precipitation total in this area is 2183 mm.

Map of the study area showing a variety of land-use types, such as protected areas, plantations and Ecosystem Restoration Areas
The landscape in the study region is composed of a mosaic of habitats, including 2985.46 km2 of peat swamp natural forests (42.4% of the study area), 1967.32 km2 of industrial plantations (27.9%), and 2093.11 km2 of other habitat types such as small-scale agriculture and rubber plantations, settlements, and industrial infrastructures (29.7%). There are four protected areas within the study area: Zamrud National Park (314.80 km2) and the Tasik Serkap (66.37 km2), Tasik Besar Serkap (49.79 km2), and Tasik Belat (25.29 km2) Wildlife Sanctuaries, in addition to the Riau Ecosystem Restoration (RER) area of 1387.32 km2.
The RER region is a biodiversity-rich peatland ecosystem with at least 76 mammals, 307 birds, 107 herpetofauna, and 190 plant species (RER-FFI 2016, 2020). In addition to the Sumatran tiger, five wildcat species occupy this area (Goenarto and Gunaryadi 2018): the Sunda clouded leopard (Neofelis nebulosa), leopard cat (Prionailurus bengalensis), marbled cat (Pardofelis marmorata), fishing cat (Prionailurus viverrinus), and flat-headed cat (Prionailurus planiceps). The Sunda clouded leopard and flat-headed cat are classified as endangered species; the fishing cat is classified as vulnerable; and the marbled cat is classified as a near-threatened species.
In addition, signs of prey such as sambar deer, wild pigs, barking deer, and primates have been detected throughout the study area.
Research methods
We used a statistical habitat modelling method, ecological niche factor analysis (ENFA), to reveal the ecogeographical variables that determine the spatial distribution of the Sumatran tiger on the Kampar Peninsula. ENFA is a multivariate statistical approach used to study the habitat suitability of a given species at large spatial scales (Basille et al. 2008; Hirzel et al. 2002). The advantage of this approach over other methods is that it requires only presence data, as accurate absence data are particularly difficult to obtain (Cianfrani et al. 2010). The model uses two types of data variables, namely, ecogeographical variables (EGVs), which included biogeographical and human disturbance variables, as the predictor variables and species presence information as the dependent variable. In this method, the study area map must be converted into a raster map consisting of N adjacent isometric cells. In ENFA, the suitability functions are calculated by comparing the species distribution within the EGV spaces to that of the whole-set cells. The mathematical procedures and implementation process in the Biomapper Software were explained in detail by Hirzel et al. 2002).
To compare the specific environmental needs of the studied species and the general environmental conditions (global environment) of the study area, which provide the whole-cell environmental variable values in the research area, three conceptions were developed during the ENFA process: marginality (M), specialization (S), and tolerance (T) (Hirzel et al. 2002; Hirzel and Gwenaelle 2008). M represents the ecological distance between the species mean and the global mean. A higher absolute value of M indicates that the ecological distance of the species optimum is far from the mean of the available habitat. A positive M value signifies that the focal species prefer a higher value than the mean value throughout the whole analysed area. S provides information about the selectivity of the studied species by comparing the species distribution to the available range of specified environmental conditions. S is the ratio of the standard deviation of the global distribution to that of the focal species. S values range from 1 to infinity. T is a more interpretable metric; it is the inverse of specialization. A T value close to 0 means that the species inhabits a very narrow niche, while a T value close to 1 means that the species occupies a wide area (Hirzel et al. 2002; Xuezhi et al. 2008).
Species data
The species data utilized herein were collected through regular biodiversity surveys and monitoring projects conducted by the RER programme from 2015 to 2020. The data collection methods included transect surveys and camera traps. In the inventory survey, the RER teams established 32 transects and 350 camera traps systematically set up within 2 × 2 km grid cells using both single and paired setting methods. Tiger sighting data (visual sightings, tracks and markings) were also recorded and collected by regular forest protection patrol teams, such as RER forest rangers, Riau Andalan Pulp and Paper (RAPP) forest protection officers and Balai Konservasi Sumber Daya Alam/Government Agency for Natural Resources Conservation (BKSDA) personnel, in joint operation.
In total, 43 Sumatran tiger presence points were used in the model (Fig. 2). The utilized data included 11 tigers identified by camera traps, 10 identified by footprints, and 22 identified by direct sightings in landscapes with different features, e.g., protected areas, RER regions, and industrial plantations (Figs. 3 and 4). The species data collection period spanned from 2018 to 2020. We used only recent species data to ensure that the environmental conditions at the time the species data were collected did not differ from the conditions captured by a remote sensing image acquired on 28 August 2020 as a source for the thematic raster maps and land cover classification maps. This short temporal interval between the species data and remote sensing data is important for this study, because land cover types in the study area, especially forest plantations, are highly dynamic. The Sumatran tiger presence dataset containing 43 data points was then converted into a Boolean raster map with cell sizes of 20 × 20 m, matching the spatial resolution of the Sentinel 2 level-2A data used to construct the thematic map and land cover classification map. The whole Kampar Peninsula area was modelled as a raster map based on a Universal Transverse Mercator (UTM)-48N projection comprising 15,944,993 cells. Cells with a value of 1 indicate species presence in the underlying area, and cells with a value equal to 0 indicate locations where the presence of a tiger could not be proven.

Map of the locations at which tigers were detected by camera traps, footprints, or direct sighting

Direct tiger sightings within cultivated areas near the Metas riparian forest (photograph provided by APRIL)

Visual sighting with a camera trap near the Tasik Belat Wildlife Sanctuary in January 2018
Environmental data
Based on the principle of ENFA, the number of EGVs must be less than the number of species presence points (43 in this study), and it is best to have at least three times as many presence points as EGVs (Hirzel et al. 2002; Kywe 2012). We selected ten EGVs, including land cover-related features (vegetation, water, and land) and human disturbance-related features (roads, forest plantations, and working camps), that theoretically influence the presence of the Sumatran tiger. Distance data were obtained to describe the distance between the focal cell and the nearest cell belonging to a specific category. The vegetation index (VI) was assessed as an indicator of the spatial vegetation variations throughout the landscape (Meng et al. 2019). The VI is defined as an arithmetic combination of two or more bands related to the spectral characteristics of vegetation (Xue and Su 2017). Because the normalized difference vegetation index (NDVI) has some deficiencies when describing the spectral behaviour of vegetation with regards to the soil background and soil–atmosphere interactions, both the soil-adjusted vegetation index (SAVI) and enhanced vegetation index (EVI) were used in this study to improve the detection sensitivity of green vegetation (Xue and Su 2017).
We used a combination of vegetation reflectance measurements, including the NDVI, EVI, and SAVI. The EVI can be used to correct the NDVI under some atmospheric circumstances and canopy background noise and is most sensitive in areas with dense vegetation. The SAVI is used to adjust the NDVI for soil brightness impacts in areas where the vegetation cover is low (European Space Agency spatiale europeenne 2015; Xue and Su 2017). The VI carries accurate information regarding aboveground biomass, primary production and spatial changes in vegetation, including both positive (“greening”) and negative (“browning”) changes.
Because this research was conducted in peatlands, where water is a significant environmental factor, we measured water parameters using the normalized difference water index (NDWI) and land/water index as variables. We used the NDWI to obtain information on the spatial distribution of water stress in vegetation. A higher NDWI value indicates sufficient moisture, while a low value indicates water stress. The land and water index distinguishes water from land and measures the quantity of water on the land surface. Thus, this index is helpful for monitoring vegetation health and soil moisture (https://eos.com/land-water/).
Maps of undisturbed, degraded, and heavily degraded forests were provided by the RER programme (Fig. 5). In addition, we used Sentinel-2A multispectral imagery acquired on 28 August 2020 as a data source to define several spectral indices with arithmetic combinations derived from the reflectance levels of two or more bands in the Saga geographic information system (GIS) 7.5.0 (European Space Agency spatiale europeenne 2015; Olaya 2004). The Sentinel-2 multispectral instrument (MSI) level-2A product was downloaded from the Earth Observing System Data Analytics website (http://www.eos/landviewer/).

EGV maps of distance and thematic variables, representing a visualization of the vector maps and the raster maps used as predictor variables in the ENFA model. The values of all cells were normalized from 0 to 1. The maps present the a UNDISF_DIST (distance to undisturbed forest), b DEGF_DIST (distance to degraded forest), c HVDEGF_DIST (distance to heavily degraded forest), d LST_DIST (distance to lake or stream), and e RPL_DIST (distance to road) f PLANTATION_IND (agriculture index: measure of the presence of forest plantations), g MULVI_IND (vegetation index: measure of vegetation greenness with NDVI, EVI and SAVI), h NDWI_IND (water index), i LANDWATER_IND (land and water index: differentiation of water from land in the landscape, vegetation health and soil moisture), j URBAN_IND (urban index: measure of the presence of working camps)
Data analysis
The ENFA calculations were performed in Biomapper 4.0 (Hirzel et al. 2002, 2004). The first four factors obtained from the eigenvalues were selected and retained to map habitat suitability, because these eigenvalues were higher than those predicted by McArthur’s broken-stick distribution (Zhenxin et al. 2010). The habitat suitability index (HSI) values ranged from 0 to 1; a value of 0 indicates an utterly unsuitable habitat, while a value of 1 denotes the most suitable habitat possible. These values were then categorized into 5 classes: unsuitable, marginally suitable, moderately suitable, suitable, and most suitable habitats.
Model evaluation
A model evaluation was conducted to quantitatively evaluate the accuracy of the derived maps when predicting the presence or absence of the target species. For this task, we used k-fold cross-validation based on Huberty’s rule in Biomapper 4 (Hirzel et al. 2002; Kywe 2012). In the cross-validation procedures, the datasets were randomly divided into k independent partitions, k-1 partitions were used to calibrate the model, and the remaining partitions were used to evaluate the model (Hirzel and Arlettaz 2003; Hirzel et al. 2006; Zhenxin et al. 2010). This procedure was repeated k times, and three presence-only evaluation measures were computed. First, the absolute validation index (AVI) was obtained to represent the proportion of evaluation partitions with habitat suitability levels greater than 0.5. The AVI values ranged from 0 to 1. Thus, this index indicates how well the model discriminated high-suitability areas from low-suitability areas (Hirzel and Arlettaz 2003; Hirzel et al. 2006; Zhenxin et al. 2010). Second, the contrast validation index (CVI), calculated by subtracting the AVI of the null model from the AVI of constructed model, indicates the extent to which the AVI differs from that obtained with a random model; this index varies from 0 to 0.5. A high CVI value means that the model performed relatively well and is both more accurate and more “contrasted” (Fielding and Bell 1997; Hirzel et al. 2006; Hirzel and Arlettaz 2003). Third, the continuous Boyce index (BI) and smooth ratio predicted frequency (P)—expected frequency (E) curve (P/E curve) were used to obtain a more continuous assessment of the model’s predictive power; these values vary from − 1 to 1 (Hirzel et al. 2006). Positive values indicate a model whose predictions are consistent with the species presence distribution in the evaluation dataset; values close to zero suggest that the model is not different from a random model; and negative values indicate an incorrect model that predicts frequent species presence in poor-quality areas (Hirzel and Arlettaz 2003; Hirzel et al. 2006).
Results
Influence of environmental factors on tiger presence
The moderate global M value obtained (0.51) suggests that the tiger habitat characteristics in the study area differ from the global habitat characteristics; the high S value (6.84) and low T value (0.15) indicate that the tigers in the study area occupy a very narrow niche. The contributions of environmental variables to M and S are shown by the coefficient values (Table 1). The first factor represents the M, which describes the distance of the species optimum from the mean habitat in the study area. The score matrix coefficients related to the first factor indicate a correlation between each EGV and M. The greater the absolute value of the coefficient is, the more the corresponding EGV contributes to M.
The subsequent factors are S factors and are sorted by decreasing amounts of explained variance. These factors describe how specialized the target species is, as determined by referring to the available range of habitats in the study area (Hirzel et al. 2002). Therefore, only a few of the first factors explain the majority of the overall information (Santos et al. 2006). The first four factors retained (out of the ten factors computed) in the score matrix is ENFA outcomes, as this analysis process extracts mutually independent factors (Hirzel et al. 2002, 2004). The four significant factors (as determined through comparisons with the MacArthur broken-stick distribution) accounted for 83.5% of the total number of eigenvalues (i.e., the total percent of the explained S) (Hirzel and Arlettaz 2003; Hirzel et al. 2002). The first factor accounts for 32% of the species S and contributes the most to species niche S; the largest specialization number was observed in the distance to heavily degraded forest, at − 0.528. This finding indicates that the “distance to heavily degraded forest” variable strongly contributes to the overall species S value.
Among the analysed EGV, the “distance to heavily degraded forest” and “distance to lake or stream” variables had the largest absolute coefficient values (− 0.528 and − 0.497, respectively), suggesting that these two variables very strongly influence tigers in the study area. The M coefficient showed that tigers essentially like to use habitats near roads (with a value lower than the average distance, − 0.25), habitats near undisturbed forests (lower than average distance, − 0.161), habitats near degraded forests (lower than the average distance, − 0.345) and habitats with sufficient atmospheric moisture (higher than average, 0.294). Tigers very much like to use habitats near heavily degraded forest (lower than average distance, − 0.5480) and near lakes and streams (lower than average distance, − 0.497). In contrast, the tigers in the study area tend to avoid plantations (lower than average value, − 0.20), dense habitats (lower-than-average VI, − 0.307), and dry vegetation and land areas (lower than average value, − 0.203). Tigers might not be influenced by working camps (the result was a value lower but very close to the average value, − 0.06).
Habitat suitability map
The habitat suitability map is provided in Fig. 6. The habitat suitability levels were categorized into five classes based on the habitat suitability values as follows: (1) unsuitable habitats (0–36); (2) marginally suitable habitats (37–52); (3) moderately suitable habitats (53–68); (4) suitable habitats (67–84) and (5) most suitable habitats (85–100). Based on the results of the habitat suitability classification method described above, the unsuitable habitat area covers 39.31% of the study area (2505.21 km2), the marginally suitable habitat area covers 25.72% (1638.99 km2), the moderately suitable habitat area covers 13.92% (887.45 km2), the suitable habitat area covers 13.90% (888.41 km2) and the most suitable habitat area covers 7.17% (457.05 km2). The spatial distribution of these habitats showed that good connections still exist among marginally suitable, moderately suitable, suitable, and most suitable habitats. Together, these classes compose approximately 60% of the total landscape or approximately 3870 km2.

Habitat suitability map derived for the Sumatran tiger on the Kampar Peninsula. The study area was classified into five classes, i.e., unsuitable, marginally suitable, moderately suitable, suitable, and most suitable habitats. The habitat suitability map is overlaid with the land-use map
Evaluation of the model
In general, the model performance was moderate, with a BI value of 0.622 ± 0.039; the AVI and CVI values were 0.611 and 0.272, respectively. The P/E curve showed a monotonic increase with increasing habitat suitability (Fig. 7). This is a sign of a good model prediction and expresses that the HSI is proportional to the probability of use by tigers. Based on this curve, the number of habitat suitability classes and their interval values were determined. The variances fluctuated along the curve. The narrowness of the confidential interval reflects its sensitivity to particular calibration points. From the curve, the confidence interval can be seen to be narrow in unsuitable (10–22) and suitable habitats (70–76) and wide in other regions. The model predictions are relatively poor for the moderately suitable and most suitable habitats, as indicated by the wide confidential intervals in these classes. Although the number of actual tiger records in this study is adequate for a habitat suitability analysis (0.006 tigers/km2) and the value is higher than those used in other research (Kanagaraj et al. 2011; Zhenxin et al. 2010), it is likely that additional animal records are necessary. Furthermore, to improve the robustness of the model, i.e., to obtain smaller variance fluctuations or narrower confidence intervals, more ground truth data should be collected, and land-use maps should be generated using higher resolution satellite imagery (Hirzel et al. 2006; Kywe 2012).

P/E curve produced by a cross-validation process: the solid line represents the mean model result obtained from the cross-validation process, and the dashed lines show the standard deviation. The red dashed line Fi = 1 represents a random model
Discussion
Our findings show that tigers have meagre tolerance values and very narrow niches in the studied peat swamp habitats. The most vital environmental variables found to influence tiger habitat preference were the distance to heavily degraded forests and distance to lakes and streams. While the results show that tigers like to use habitats near heavily degraded forests, there are no tiger presence records in these habitats. Most tigers’ sightings occurred on the boundaries between undisturbed and degraded forests, nearby heavily degraded forests, or near streams or lakes. Degraded forests do not necessarily indicate low-quality habitats for tigers per se (Linkie et al. 2008). Most likely, degraded forests with relatively few trees support more prey species than undisturbed forests (Rayan and Mohamad 2009). These degraded sites, in combination with undisturbed forests, riparian forests, and plantations, form a mixed forest that provides enough food and undergrowth to support a sufficient density of tigers. The success of tiger hunting is also higher in areas with low tree cover (Pakpien et al. 2017). However, degraded sites are more accessible for hunters; thus, ensuring patrols throughout these areas is vital for securing the tiger population, along with tiger prey, from poaching.
Tigers also like to use riparian peat swamp forests and the surrounding areas. These areas are more fertile than other forest regions because they regularly receive nutrients from rivers during flooding events (Wahyunto et al. 2013). Moreover, terrestrial and boreal tiger prey species prefer this habitat type due to the abundant food sources, such as vines, fruit tree fruits and young leaves for primates and forage for ungulates (Wantzen et al. 2008). Former research conducted in Malaysian peat swamps showed that poor habitats located away from rivers contain fewer species, lower individual densities, and smaller group sizes (Marsh and Wilson 1981). Furthermore, streams might form natural boundaries for tiger home ranges. Thus, it is necessary to perform intensive routine patrols near rivers (Naha et al. 2016).
The impacts of logging roads on tigers are unclear. For example, Linkie et al. (Linkie et al. 2008) found that tigers do not like to stay in habitats near public roads or logging roads. However, another study discovered that more tiger scat was observed along open trails and logging roads than in animal trails in forests. (Sunarto et al. 2012) found that, at the landscape scale, the distance to a road was not an important factor affecting the tiger occupancy of the study area. This finding indicated that tigers, at least, do not avoid man-made trails or logging roads (Smith et al. 2018). On the Kampar Peninsula, tigers prefer to travel and be active near logging roads. Logging roads in wet ecosystems and nearby areas are probably more accessible to tigers, because these routes are almost always dry.
On the other hand, in peat swamp forests, tigers tend to prefer areas with relatively dense understories (Sunarto et al. 2012). Most likely, in less dense forests, more sunlight can reach the forest ground and promote the growth of grasses and new seedlings, thereby increasing the availability of food types that attract more ungulates, the prey base of tigers. With regards to acacia plantations, tigers do not like areas containing young acacia plants and intensive human activities and prefer to inhabit old plantations with relatively thick subcanopies and more leaf litter (Sunarto et al. 2012).
From the habitat suitability map obtained herein, we predicted that the amount of available tiger habitat in the study area spanned 60% of the entire landscape or 3870 km2; one-fourth of this area constituted marginal habitats. Therefore, habitat quality improvements should be a focus in this area, if successful tiger translocation are a future objective. These various available habitat regions are still connected; thus, tigers can roam freely inside the habitats. The remaining 40% of the study area is not viable due to natural environmental constraints such as peat domes or heavy modification by humans, thus causing these habitats to be entirely unsuitable for tigers.
However, the area of available habitat is not sufficient to support a viable tiger population. The tiger density varies from location to location. In mineral soil habitats, such as in the Bukit Barisan Selatan National Park, the estimated tiger density is approximately 1.6 tigers/100 km2 (O’Brien et al. 2003); in Leuser National Park, the estimated density is 0.3 tigers/100 km2 (Wibisono and Pusporini 2010); and in Kerinci Seblat National Park, the estimated density is 1.5–3.3 tigers/100 km2 (Linkie et al. 2006). In low-density habitats, such as peatland ecosystems, the size of the home range of a tiger is approximately 289 km2 (Sunarto et al. 2012). Based on this estimate, we can assume that approximately 13 tigers can live on the Kampar Peninsula.
Peat swamp forests, especially peatland domes, cannot support an abundance of terrestrial wildlife as a consequence of their limited primary production due to the low soil nutrient levels (MacKinnon et al. 1996). Our research showed that a large block of undisturbed peatland dome was an unsuitable habitat for the Sumatran tiger. This area is a poor habitat that is out of the reach of floodwaters and receives nutrient inputs only from rain and the local plants themselves (Wahyunto et al. 2013). The infertility of the soil in this region is reflected by vegetation features such as the decreasing canopy height, decreasing total biomass, decreasing tree diameters, and increasing leaf thicknesses (MacKinnon et al. 1996). Thick leaves are likely a strategy with which plants defend themselves against herbivory. In addition, medium and large herbivores with pointed feet face difficulties travelling in terrain with soft soils and porous ground surfaces (MacKinnon et al. 1996). Therefore, the very low density of herbivores in this area results in a low probability of Sumatran tiger occurrences in the peatland dome. More productive peatlands are located near rivers and in areas where the peat layer is thin (Wahyunto et al. 2013); these regions are more suitable as wildlife habitats.
The other important environmental limitation of peat swamp habitats for terrestrial mammals is the presence of excessive water; in peatlands, some areas are always saturated by freshwater. Areas near rivers are usually flooded during rainy seasons. Wide and small rivers, canals, and flooded areas determine and restrict the movement patterns of animals. The Kampar Peninsula is bordered by two large rivers and a strait, namely, the Siak River in the west, the Kampar River in the east, and the Panjang Strait in the north. The width of these rivers varies from one hundred metres in the upper reaches to more than one kilometre downstream. These rivers act as barriers for tiger movement at some locations, as tigers will usually not cross a river more than 400 m in width. Inside the studied habitats, many small rivers and water canals present radial drainage patterns in which streams flow in different directions from a central peak. These streams might not impose barriers for tigers. In mangrove forests in Sundarbans, tigers crossed canals with a mean width of 54 m at an average rate of 5 times per day (Naha et al. 2016).
For an isolated population, the probability of extinction is a function of the population size. Small populations with high variations face higher extinction risks than large populations (Legendre et al. 2008). A large population of top carnivores requires a large and connected habitat to support viability (Huck et al. 2010), and the Kampar Peninsula is suitable for tiger reintroduction but is not sufficiently large to support large-scale tiger conservation. Minimizing poaching and improving the habitat quality in the study region is necessary but not sufficient to secure a viable tiger population. The maintenance of connective habitats between the Kampar and Kerumutan landscapes is crucial for saving Sumatran tigers in the future. With a total area larger than 10,000 km2, the landscape is sufficient for maintaining the minimum viable Sumatran tiger population. (Luskin et al. 2017) estimated that the number of adult tigers on the Kampar Peninsula and in Kerumutan decreased from 42 individuals in 2000 to 26 individuals in 2012. Thus, the reintroduction of tigers in the Kampar Peninsula is feasible for increasing the tiger population size.
Conclusion
Although the peatland ecosystems studied herein occupy a large area and serve as an important landscape for wild animals in Indonesia, including tigers, peatlands are commonly considered poor habitats with limited carrying capacities for supporting apex predator populations. However, the role of peatlands in economic development has been continuously increasing. Natural peatlands have been converted into many modified land-use types, such as plantation forests, agricultural areas, settlements, and other infrastructures. Investigating the suitability of these new landscapes as tiger habitats is essential for obtaining data that can be used to improve upon existing conservation practices, such as site-specific wildlife reintroductions and landscape management. This research revealed that 60% of the total Kampar landscape can be considered tiger habitats with varying degrees of suitability. Tigers occupy a narrow niche within the whole habitat, and the difference between the habitat requirements of tigers and the average habitat condition in the study area is quite large. The restoration of marginal habitats, effective control of poaching, and maintenance of landscape connectivity are the keys to successful tiger reintroduction and the conservation of a long-term viable tiger population.
Data availability
The data that support the findings of this study are available on request from the corresponding author (SP). The data of tiger locations are not publicy available in order to protect this endangered species.
References
Basille M, Calenge C, Marboutin É, Andersen R, Gaillard J-M (2008) Assessing habitat selection using multivariate statistics: some refinements of the ecological-niche factor analysis. Ecol Modell 211(1–2):233–240. https://doi.org/10.1016/j.ecolmodel.2007.09.006
Boast LK, Good K, Klein R (2016) Translocation of problem predators: is it an effective way to mitigate conflict between farmers and cheetahs Acinonyx jubatus in Botswana? Oryx 50(3):537–544. https://doi.org/10.1017/S0030605315000241
Brun C, Cook AR, Lee JSH, Wich SA, Koh LP, Carrasco LR (2015) Analysis of deforestation and protected area effectiveness in Indonesia: a comparison of bayesian spatial models. Glob Environ Change 3:285–295. https://doi.org/10.1016/j.gloenvcha.2015.02.004
Cheyne SM (2006) Wildlife reintroduction: considerations of habitat quality at the release site. BMC Ecol 6(5):1–8. https://doi.org/10.1186/1472-6785-6-5
Cianfrani C, Lay GL, Hirzel AH, Loy A (2010) Do habitat suitability models reliably predict the recovery areas of threatened species? J Appl Ecol 47(2):421–430. https://doi.org/10.1111/j.1365-2664.2010.01781.x
Cook CN, Morgan DG, Marshall DJ (2010) Reevaluating suitable habitat for reintroductions: lessons learnt from the eastern barred bandicoot recovery program. Anim Conserv 13(2):184–195. https://doi.org/10.1111/j.1469-1795.2009.00320.x
European Space Agency ESA (2015) Sentinel-2, User Handbook.
Fielding AH, Bell JF (1997) A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ Conserv 24(1):38–49. https://doi.org/10.1017/S0376892997000088
Goenarto P, Gunaryadi D (2018) Biodiversity of Kampar Peninsula. In: Shimatani Y, Natori Y, Nakamura R (Eds.), 8th Asian Wetland Symposium-Wetlands for Sustainable Life Saga: AWS2017. pp. 307–308
Goodrich JM, Miquelle DG (2005) Translocation of problem Amur tigers Panthera tigris altaica to alleviate tiger-human conflicts. Oryx 39(4):454–457. https://doi.org/10.1017/S0030605305001146
Hirzel AH, Arlettaz R (2003) Modeling habitat suitability for complex species distributions by environmental-distance geometric mean. Environ Manage 32(5):614–623. https://doi.org/10.1007/s00267-003-0040-3
Hirzel AH, Gwenaelle LL (2008) Habitat suitability modelling and niche theory. J Appl Ecol 45:1372–2138. https://doi.org/10.1111/j.1365-2664.2008.01524.x
Hirzel AH, Hausser J, Perrin N (2002) Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data? Ecology 83(2):2027–2036. https://doi.org/10.2307/3071784
Hirzel AH, Posse B, Oggier PA, Crettenand Y, Glenz C, Arlettaz R (2004) Ecological requirements of reintroduced species and the implications for release policy: the case of the bearded vulture. J Appl Ecol 41:1103–1116. https://doi.org/10.1111/j.0021-8901.2004.00980.x
Hirzel AH, Lay GL, Helfer V, Randin C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Modell 9:142–152. https://doi.org/10.1016/j.ecolmodel.2006.05.017
Huck M, Jedrzejewski W, Borowik T, Miłosz-Cielma M, Schmidt K, Jedrzejewska B, Nowak S, Mysłajek RW (2010) Habitat suitability, corridors and dispersal barriers for large carnivores in Poland. Acta Theriol 55(2):177–192. https://doi.org/10.4098/j.at.0001-7051.114.2009
Kanagaraj R, Wiegand T, Kramer-Schadt S, Anwar M, Wiegand TR, Goyal SP (2011) Assessing habitat suitability for tiger in the fragmented Terai Arc landscape of India and Nepal. Ecography 34(6):970–981. https://doi.org/10.1111/j.1600-0587.2010.06482.x
Karanth KU, Chellam R (2009) Carnivore conservation at the crossroads. Oryx 43(01):1–2. https://doi.org/10.1017/S003060530843106X
Kywe TZ (2012) Habitat suitability modeling for tiger (Panthera tigris) in the Hukaung Valley Tiger Reserve, Northern Myanmar. Dissertation, George August University
Legendre S, Schoener TW, Clobert J, Spiller DA (2008) How is extinction risk related to population-size variability over time? A family of models for species with repeated extinction and immigration. Am Nat 172(2):282–298. https://doi.org/10.1086/589454
Linkie M, Chapron G, Martyr DJ, Holden J, Leader-Williams N (2006) Assessing the viability of tiger subpopulations in a fragmented landscape. J Appl Ecol 43(3):76–586. https://doi.org/10.1111/j.1365-2664.2006.01153.x
Linkie M, Haidir IA, Nugroho A, Dinata Y (2008) Conserving tigers Panthera tigris in selectively logged Sumatran forests. Biol Conserv 141(9):2410–2415. https://doi.org/10.1016/j.biocon.2008.07.002
Linkie M, Martyr D, Harihar A, Mardiah S, Hodgetts T, Risdianto D, Subchaan M, Macdonald D (2018) Asia’s economic growth and its impact on Indonesia’s tigers. Biol Conserv 219:105–109. https://doi.org/10.1016/j.biocon.2018.01.011
Luskin MS, Albert WR, Tobler MW (2017) Sumatran tiger survival threatened by deforestation despite increasing densities in parks. Nat Commun 1783:1–9. https://doi.org/10.1038/s41467-017-01656-4
MacKinnon K, Hatta G, Halim H, Mangalik A (1996) The ecology of Kalimantan (Indonesian Borneo) 3. Periplus Editions (HK) Limited, Canada
Margono BA, Potapov PV, Turubanova S, Stolle F, Hansen MC (2014) Primary forest cover loss in Indonesia over 2000–2012. Nat Clim Change 4:1–6. https://doi.org/10.1038/NCLIMATE2277
Marsh CW, Wilson WL (1981) Primate research and conservation in Malaysia. CAB Rev 14(4):1–10. https://doi.org/10.1079/PAVSNNR201914004
Meng Y, Liu X, Wu L, Liu M, Zhang B, Zhao S (2019) Spatio-temporal variation indicators for landscape structure dynamics monitoring using dense normalized difference vegetation index time series. Ecol Indic 107:1–11. https://doi.org/10.1016/j.ecolind.2019.105607
Miettinen J, Liew SC (2010) Status of peatland degradation and development in Sumatra and Kalimantan. Ambio 39(5):394–401. https://doi.org/10.1007/s13280-010-0051-2
Miettinen J, Shi C, Liew SC (2016) Land cover distribution in the peatlands of Peninsular Malaysia, Sumatra and Borneo in 2015 with changes since 1990. Glob Ecol Conserv 6:67–78. https://doi.org/10.1016/j.gecco.2016.02.004
Morehouse AT, Hughes C, Manners N, Bectell J, Bruder T (2020) Carnivores and communities: a case study of human-carnivore conflict mitigation in Southwestern Alberta. Front Ecol Evol 8:1–15. https://doi.org/10.3389/fevo.2020.00002
Naha D, Jhala YV, Qureshi Q, Roy M (2016) Ranging, activity and habitat use by tigers in the mangrove forests of the Sundarban. PLoS One 6:1–16. https://doi.org/10.1371/journal.pone.0152119
Nyhus PJ, Tilson R (2004) Characterizing human-tiger conflict in Sumatra, Indonesia: implications for conservation. Oryx 38(1):68–74. https://doi.org/10.1017/S0030605304000110
O’Brien TG, Kinnaird MF, Wibisono HT (2003) Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim Conserv 6(2):131–139. https://doi.org/10.1017/S1367943003003172
Olaya V (2004) A gentle introduction to SAGA GIS. The SAGA User Group eV, Gottingen, Germany. https://doi.org/10.1017/CBO9781107415324.004
Pakpien S, Simcharoen A, Duangchantrasiri S, Chimchome V, Pongpattannurak N, Smith JLD (2017) Ecological covariates at kill sites influence tiger (Panthera tigris) hunting success in Huai Kha Khaeng wildlife sanctuary Thailand. Trop Conserv Sci 10:1–7. https://doi.org/10.1177/1940082917719000
Priatna D, Santosa Y, Prasetyo LB, Kartono AP (2012) Home range and movements of male translocated problem tigers in Sumatra. Asian J Conserv Biol 1(1):20–30
Rayan DM, Mohamad SW (2009) The importance of selectively logged forests for tiger Panthera tigris conservation: a population density estimate in Peninsular Malaysia. Oryx 43(1):48–51. https://doi.org/10.1017/S0030605308001890
RER-FFI (2016) Biodiversity of the Kampar Peninsula: summary report. RER Publication No. 1, Jakarta
RER-FFI (2020) Current status of hutan rawa gambut Siak-Kampar important bird area. RER Publication Series No.4, Jakarta
Resende PS, Viana-Junior AB, Young RJ, de Azevedo CS (2020) A global review of animal translocation programs. Anim Biodivers Conserv 43(2):221–232. https://doi.org/10.32800/abc.2020.43.0221
Sanderson E, Forrest J, Loucks C, Ginsberg J, Dinerstein E, Seidensticker J, Wikramanayake E (2010) Setting priorities for the conservation and recovery of wild tigers: 2005–2015. In: Tilson R, Nyhus P (eds) Tigers in the world, 2nd edn. Academic Press, Cambrigde
Santos X, Brito JC, Sillero N, Pleguezuelos JM, Llorente GA, Fahd S, Parellada X (2006) Inferring habitat-suitability areas with ecological modelling techniques and GIS : a contribution to assess the conservation status of Vipera latastei. Biol Conserv 130:416–425. https://doi.org/10.1016/j.biocon.2006.01.003
Smith O, Wang J, Carbone C (2018) Evaluating the effect of forest loss and agricultural expansion on Sumatran tigers from scat surveys. Biol Conserv 221:270–278. https://doi.org/10.1016/j.biocon.2018.03.014
Sunarto S, Kelly MJ, Parakkasi K, Klenzendorf S, Septayuda E, Kurniawan H (2012) Tigers need cover: multi-scale occupancy study of the big cat in Sumatran forest and plantation landscapes. PLoS One 7:e3085. https://doi.org/10.1371/journal.pone.0030859
Sunarto S, Kelly MJ, Klenzendorf S, Vaughan MR, Zulfahmi HMB, Parakkasi K (2013) Threatened predator on the equator: multi-point abundance estimates of the tiger Panthera tigris in Central Sumatra. Oryx 47(2):211–220. https://doi.org/10.1017/S0030605311001530
Sunarto S, Kelly MJ, Parakkasi K, Hutajulu MB (2015) Cat coexistence in central Sumatra: ecological characteristics, spatial and temporal overlap, and implications for management. J Zool 296(2):104–115. https://doi.org/10.1111/jzo.12218
The World Bank and BPS (2019) Pilot ecosystem account for Indonesian peatlands (Sumatera—Kalimantan). Report
Wahyunto W, Supriatna W, Agus F (2013) Land use change and recommendation for sustainable development of peatland for agriculture: case study at Kubu Raya and Pontianak Districts, West Kalimantan. Indones J Agric Sci 11(1):32–40. https://doi.org/10.21082/ijas.v11n1.2010.p32-40
Wantzen KM, Yule CM, Tockner K, Junk WJ (2008) 7-Riparian wetlands of tropical streams. In: Dudgeon D (ed) Tropical stream ecology. Elsevier, pp 199–217. https://doi.org/10.1016/B978-012088449-0.50009-1
Wibisono HT, Pusparini W (2010) Sumatran tiger (Panthera tigris sumatrae): a review of conservation status. Integr Zool 5(4):313–323. https://doi.org/10.1111/j.1749-4877.2010.00219.x
Wibisono HT, Linkie M, Guillera-Arroita G, Smith JA, Sunarto et al (2011) Population status of a cryptic top predator: an island-wide assessment of tigers in Sumatran rainforests. PLoS One 6(11):e25931. https://doi.org/10.1371/journal.pone.0025931
Xue J, Su B (2017) Significant remote sensing vegetation indices: a review of developments and applications. J Sens 2017:1353691. https://doi.org/10.1155/2017/1353691
Xuezhi W, Weihua X, Zhiyun O, Jianguo L, Yi X, Youping C (2008) Application of ecological-niche factor analysis in habitat assessment of giant pandas. Acta Ecol Sin 28(2):821–828. https://doi.org/10.1016/S1872-2032(08)60030-X
Zhenxin L, Zimmermann F, Hebblewhite M, Purekhovsky A, Morschel F, Chunquan Z, Miquelle D (2010) Technical report on the identification of potential tiger habitat in the Changbaishan ecosystem. WWF, WCS, Northeast Normal University, KORA, the University of Montana, Northeast China
Acknowledgements
This research is conducted as an institutional collaboration among Gadjah Mada University, the Ministry of Environment and Forestry of the Republic of Indonesia, and the APRIL Group. The authors wish to thank these institutions for their financial and logistical support during the collection of field data.
Author information
Authors and Affiliations
Contributions
Manuscript title: habitat suitability of a peatland landscape for tiger translocation on Kampar Peninsula, Sumatra, Indonesia. The authors confirm contribution to the paper as follows: study conception and design: SP, AB; data collection: SP, AB, AHS; analysis and interpretation of results: SP; AB; AHS; original draft preparation: SP; AB; AHS.; writing—review and editing: SP; AB; AHS. All authors reviewed the results and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
All authors have no conflicts of interest.
Additional information
Handling editor: Michael Somers.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pudyatmoko, S., Budiman, A. & Siregar, A.H. Habitat suitability of a peatland landscape for tiger translocation on Kampar Peninsula, Sumatra, Indonesia. Mamm Biol 103, 375–388 (2023). https://doi.org/10.1007/s42991-023-00361-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-023-00361-8