
Abstract
Conservation translocations have the potential to strengthen populations of threatened and endangered species, but facilitating integration of translocated individuals with resident populations remains a substantial challenge. Developing functional social relationships like cooperative partnerships or establishing clear dominance hierarchies may be critical to integration of released individuals. Developing such relationships has not received much attention in translocation research, especially for long-lived, socially complex animals for which establishment and navigation of social environments is often a lengthy process that requires sustained monitoring to understand. Here, we present a case study of the social associations of African savannah elephant (Loxodonta africana) calves that have been rehabilitated and released into a fenced wildlife sanctuary in northern Kenya with a resident population of elephants. We use focal follows of interactions pre-release and GPS tracking post-release to quantify social associations of calves with each other and with resident elephants at the release site. We demonstrate how this approach supports translocation monitoring by capturing temporal trends in social patterns within and between release cohorts and among released elephants and wild elephants already resident at the site during a transitional soft release period. Our results show that initial post-release social behavior of rehabilitated calves is related to histories of interaction with familiar individuals and cohort membership and that released calves increased their associations with residents over time. This information provides new behavioral insights for guiding elephant release projects, like the strength of relationships within and among release cohorts, the time to integration with the resident population, and the occurrence and increased incidence of societal fission–fusion. Further, this study provides an example of the utility of animal behavior research to achieve and assess progress towards conservation objectives, and to develop monitoring tools for conservation managers.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wildlife translocation is a valuable applied tool in conservation and is used in a variety of contexts and across a wide range of species (International Union for Conservation of Nature 2013; Seddon et al. 2014). Among conservation translocations, rehabilitation of wildlife with the goal of eventual release is a widespread practice in response to orphaning, injury, and confiscation (Cheyne 2009; Kelly et al. 2010), and can be undertaken to address individual animal welfare concerns and to reinforce wild populations. There is particular interest in such practices where rehabilitation and release to the wild involve species of conservation concern (International Union for Conservation of Nature 2013). However, rehabilitation and release projects are typically resource intensive, requiring considerable investment in each animal and often over sustained periods of time (Cheyne 2009). Additionally, periods under human care may inadvertently alter animal behavior as animals may learn to rely on or have positive associations with humans. Identifying the factors that promote the survival and reproduction of released wildlife and establishing standardized benchmarks of success based on these factors is key to improve the effectiveness of such programs.
Challenges related to behavioral responses of translocated animals are particularly prevalent in conservation translocations, whereby released animals do not behave appropriately for their release environment (Berger-Tal et al. 2020). One behavioral factor that should be considered in translocations is the social dynamics of released animals, including social integration within release cohorts and with the resident population at the release site, if applicable (Pinter-Wollman et al. 2009; Berger-Tal and Saltz 2014; Poirier and Festa-Bianchet 2018; Goldenberg et al. 2019). This may be relevant to released animals regardless of the social structure of the species of interest, as many types of social relationships (e.g., territorial, Shier and Swaisgood 2012; cooperative (Shier 2006) require investment to establish. Promoting integration of released individuals within release cohorts and with resident populations may be particularly important for long-lived species that invest heavily in social relationships (Cheyne 2009) that are known to be negatively affected by social disruption (Engh et al. 2006; Foley et al. 2008; Nunez et al. 2015), or whose knowledge of the new landscape may be facilitated by social relationships (Goldenberg et al. 2021). Such integration may be especially challenging in orphaning, rehabilitation and release contexts in which animals may have had reduced exposure to normal social development and mentorship, and/or have spent time under human care and consequently may be more disconnected from social behavior typical of their species. In such cases, following the social behavior of both released animals and animals in resident populations may provide insight to improve the efficacy of rehabilitation and release programs (Berger-Tal and Saltz 2014). Such an approach may inform expectations for timelines of social integration, illuminate characteristics of resident or rehabilitated individuals that hasten or lengthen integration, or identify key management actions that facilitate integration. There is thus great potential in following individuals over time and identifying challenges to success, which are often behavioral in the wider conservation translocation literature (Berger-Tal et al. 2020).
Elephants are a particularly relevant taxon that may benefit from simultaneous individual-based monitoring of translocated and resident animals at release sites. There are several elephant rehabilitation projects across both Asian (Elephas maximus) and African elephant (Loxodonta spp.) range states that require significant resources like time, personnel, and funding (McKnight 1995; Perera et al. 2016; Thitaram et al. 2015; also see www.gamerangersinternational.org/wildlife-rescue; www.retetielephants.org/). But, while resource intensive, the cascading benefits of elephant translocations can facilitate ecosystem management goals. Elephants are considered keystone species because of their ability to disperse seeds across vast distances and to convert woodland into savannah by browsing, among other ecosystem structuring roles (Campos-Arceiz and Blake 2011); elephant release projects may, therefore, contribute substantially to ecosystem management goals in addition to species-level management goals (Louys et al. 2014; Snijders 2020). Furthermore, release projects hold enormous potential for engaging the public in wider biodiversity and conservation issues surrounding these high-profile and charismatic species that are red listed as endangered (www.iucnredlist.org). For these reasons, elephant rehabilitation and release projects may be an important tool for conservation, and increasing their efficacy is paramount.
Social relationships influence elephant survival in a multitude of ways. For example, elephant family groups that ventured together outside of a protected area during a drought had lower calf mortality, attributed to matriarch knowledge of the landscape (Foley et al. 2008); groups with older matriarchs exhibited greater knowledge of other individuals within the population (McComb et al. 2001); dominance relationships among females structured their landscape use during periods of resource constraint (Wittemyer et al. 2007); and dominance relationships among males influenced their reproductive opportunities (Rasmussen 2005). Given this dependence on long-term bonds, characterizing the formation and nature of social interactions amongst rehabilitated orphans and the social integration process of rehabilitated elephants into wild populations may generate applied strategies that can be used in an iterative way to support future translocations (Kuiper et al. 2018). Investigation into social interactions is thus a worthwhile avenue to increase the conservation value of these projects as the social relationships that released elephants form or maintain may determine their ability to survive, locate resources, and reproduce (Evans et al. 2013). Additionally, because elephants rehabilitated under human care may be less fearful of humans than elephants without this experience, social integration with wild elephants that maintain a fear of humans may be important for released animals in learning appropriate behavior around humans they may encounter once released.
Individual-based monitoring has proven to be a powerful research tool for illuminating diverse aspects of elephant social behavior, e.g., emergent social structure that arises from individual relationships (Douglas-Hamilton 1972; Moss 1988; Wittemyer et al. 2005), social responses to human threats (Goldenberg et al. 2016; Shannon et al. 2013), and we suggest that it can be effectively used to characterize the social integration process of released elephants as well. Indeed, research using individual-based long-term demographic and social behavioral records of wild African elephant orphans has provided insight into the process of social integration for individuals facing family disruption due to poaching and drought (Goldenberg et al. 2016). Despite resilient social structure (determined from the composition of elephant groupings) within a population experiencing heavy mortality and thus many orphaning events (Goldenberg et al. 2016), orphans received greater rates of aggression (Goldenberg and Wittemyer 2018) and interacted less with mature adults when compared with non-orphans (Goldenberg and Wittemyer 2017), likely related to their peripheral positioning in social groups. While orphans in the wild may remain with their natal families, some do not and may eventually join with groups they were not born into (Vidya et al. 2007; Wittemyer et al. 2009; Goldenberg et al. 2016). Particular individuals may facilitate this process like young calves (Thitaram et al. 2015) or peripheral animals including independent bulls (Goldenberg and Wittemyer 2017). Such insights from wild elephants may inform understanding of the social processes of rehabilitated orphaned or abandoned calves, which comprise one of the more common forms of elephant rehabilitation and release endeavors across range countries.
The Reteti Elephant Sanctuary (RES) of the Namunyak Wildlife Conservation Trust (NWCT) in Kenya was established in 2016 as a rehabilitation site for abandoned or orphaned elephant calves (www.retetielephants.org). Once rehabilitated and weaned, calves are soft released in small cohorts into a fenced black rhinoceros sanctuary within the neighboring Sera Conservancy where they become accustomed to life in the wild prior to hard release out of the fenced sanctuary. A small population of wild elephants resides within the fenced sanctuary, including a few families with calves of different ages and several independent bulls. Here, we present results on the social behavior of three cohorts (hereafter C1, C2, and C3) of calves released into the rhino sanctuary in 2019 and 2020. We rely on an individual identification database of released calves and resident elephants built from aerial, ground, and motion-triggered camera surveys. We use focal follows of calf social interactions conducted pre-release to characterize orphan social networks and we use inter-individual distances based on GPS tracking to assess social integration post-release. We focus on the following two primary aspects of post-release orphan social behavior: relationships among orphans and relationships between orphans and resident elephants. Specifically, we test (Q1) whether social interaction rates or temporal overlap at the RES prior to release predict orphan cohesion post-release, and (Q2) whether orphan cohesion with other orphans and with resident elephants changes as a function of time. We discuss our results in the context of this and similar projects that may benefit from long-term post-release monitoring of individual relationships.
Materials and methods
Study site and animals
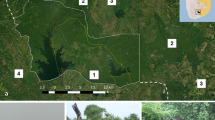
Rescued elephant calves are resident at the Reteti Elephant Sanctuary (1.11° N, 37.46° E) within NWCT for varying lengths of time where they are rehabilitated (Figs. 1, 2). NWCT is situated within a wider network of community conservancies across the north of Kenya. Calves reported to authorities north of the equator that cannot be reunited with their families are brought to RES, with most coming from Laikipia and Samburu counties, in which RES is located. Upon arrival to RES, calves are given veterinary care, bottle-fed formula every three hours, and introduced to the orphan herd at the sanctuary once in healthy condition. They forage in the surrounding area between bottle feedings during daylight hours, where they may encounter wild elephant dung, urine, and other olfactory elephant signs, and on occasion, wild elephants themselves. At night they are brought into a fenced enclosure and are always under the supervision of Reteti keepers. Once a cohort of calves is selected for release, they are gradually weaned from bottle feedings and released into the approximately 107 km2 fenced black rhinoceros sanctuary in the neighboring Sera Conservancy (1.04–1.66° N, 37.75–37.92° E). The fenced release site is itself a transition point for calves (soft release stage), with the planned final stage a release out of the fenced area and into the wider landscape used by the free-ranging Laikipia-Samburu elephant population (hard release stage). The soft release site was chosen following a rapid habitat assessment and carnivore pressure survey.

The Reteti Elephant Sanctuary is located within Namunyak Conservancy, and the fenced rhino sanctuary where orphans are released is within the neighboring Sera Conservancy. The conservancies are separated by the A2 highway

Timeline of orphans released into Sera Conservancy indicating the periods during which they were in the wild prior to rescue, the periods during which they were resident at Reteti following rescue, and the periods during which they were in the wild in the fenced rhino sanctuary. Bars begin at estimated birth dates. Note the varying lengths of time and temporal overlap of orphans resident at Reteti pre-release
The first cohort of three male elephants (C1: “Warges”, “Sosian”, and “Ilngwesi”) was released on May 2, 2019; the second cohort, comprised of two females and one male (C2: “Shaba”, “Mpala”, and “Pokot”, respectively), was released on November 16, 2019; and the third cohort, comprised of two females and two males (C3: “Nadosoit”, “Nchurai”, “Loisaba”, and “Baawa”, respectively), was released on May 28, 2020. All releases were timed with favorable ecological conditions in the sanctuary following the onset of the wet season. These ten calves were between the ages of 3 and 5 years at release and were present at RES prior to release for time periods ranging from ~ 2 to 3.5 years (Fig. 2).
In addition to the resident black rhinoceros for which the fenced sanctuary was established, the rhino sanctuary is home to a representative variety of other fauna native to the northern Kenya landscape, including reticulated giraffe, Grevy’s zebra, hyena, wild elephants (both family groups and independent bulls), and the occasional lion, as well as diverse species guilds of antelope and mesocarnivores. The landscape is characterized as semi-arid savannah and is punctuated by seven permanent water points and eleven seasonal water sources. Pastoralists from the surrounding areas occasionally use the area inside of the fence to graze livestock during seasonal periods when pasture is degraded and dry elsewhere.
Data collection
Focal follows
In order to characterize social relationships among calves prior to release, we conducted focal follows of individual calves at RES. Calves were observed for up to 10 min at a time, during which all interactions with other elephants were recorded (Appendix Fig. A1). Follows were ended earlier than ten minutes if calves were not clearly visible, and variable lengths of focal follows were accounted for in subsequent rate calculations. Interactions recorded included affiliative behaviors like body rubbing, comforting, greeting, playing, putting a trunk to another’s mouth, and trunk touching; and aggressive or dominance related behaviors including chasing, displacing, and pushing, all of which were considered behavioral events that occur over brief periods (Altmann 1974; Goldenberg and Wittemyer 2018). We refer to these as “interactions” throughout the manuscript.
Individual identification
In January 2019 in anticipation of the first calf release, we surveyed the release site on foot, by vehicle, by aircraft (Cessna 206), and later with camera traps in order to construct an individual identification database of the resident elephants within the sanctuary and to determine family group and age structure among them, which was later used to guide GPS collar deployment (Douglas-Hamilton 1972). Aircraft surveys were systematic across the study area, flying set grid lines. Vehicle surveys were limited to roads and conducted to maximize encounter rates with elephants. Foot surveys were used to augment vehicle surveys, where elephants were approached clandestinely to enable identification. Camera traps were placed at water holes and other high traffic locations. We used characteristics like ear tears and tusk shape as identifying features, and assigned ages according to standard visual estimation methods for the species (Moss 1996). Surveys indicated that there were 39 elephants present in the sanctuary. In an effort to reduce elephant density within the fenced area, the Sera Conservancy and Northern Rangelands Trust drove a cohesive group of wild elephants out of the sanctuary by aircraft in September 2019, which reduced the wild population within the fenced sanctuary by eight individuals.
GPS tracking
Most (N = 8) released orphan calves were fitted with GPS satellite collars during bottle feedings prior to release (savannahtracking.com), with at least two individuals collared in each cohort. Two elephants, Mpala and Loisaba, were not collared; analyses that include tracking data, therefore, exclude these two calves. Additionally, Warges’ collar failed to send locations beginning in mid-October 2019; thus he was excluded from tracking analysis following collar failure. In addition to the calves, a selection of five wild elephants based on surveys of the resident population within the fenced area were collared in late May 2019 (Nfemales = 4: “Serteta”, est. age 40–45; “Kaingus”, est. age 30; “Lpupo”, est. age 12; and “Kalama”, est. age 30–35; Nmales = 1: “Chapulo”, est. age 12). Females represented distinct social units. Serteta and Kaingus were part of the eight elephants that left the sanctuary and were thus unable to interact with orphans after September 2019. Both orphan and wild resident elephant collars were programmed to collect a GPS location every 30 min, but the resident collars were switched to collect locations every 1 h at the end of 2019 to extend battery life.
Data analysis
Focal follow datasets included in analyses spanned December 20, 2018 to September 16, 2019, the period during which both the second and third release cohorts were present in the larger orphan herd prior to separation in preparation for release (Nhours = 147.74; Mean hours per calf = 21.11; Ncalves = 7). Data were not available for this analysis for the three calves in the first release cohort because they were already separated from the larger herd in preparation for release when behavioral monitoring began in December 2018.
Elephants reinforce bonds using affiliative interactions and use dominance interactions and aggression to negotiate hierarchies with one another; their familiarity with one another based on these interactions in turn affects how they socialize and use landscapes (Archie et al. 2006; Wittemyer et al. 2007). We, therefore, constructed pre-release social networks for orphans that included both affiliative and aggressive interactions. For every pair of elephants included in the social network (Npairs = 21), the network tie connecting them represents a rate of interaction. A rate of interaction between elephants A and B was calculated as follows: \(\mathrm{Rate}=\frac{{\mathrm{Int}}_{\mathrm{AB}}+{\mathrm{Int}}_{\mathrm{BA}}}{{\mathrm{Time}}_{\mathrm{AB}}+{\mathrm{Time}}_{\mathrm{BA}}}\), where IntAB is the number of interactions between A and B during focal follows of A, IntBA is the number of interactions between A and B during focal follows of B, TimeAB is the total length of time that A was followed while B was present at RES prior to preparation for release, and TimeBA is the total length of time that B was followed while A was present at RES prior to preparation for release. Rates of interaction between pairs were then used in subsequent analysis.
To determine whether social interaction patterns or temporal overlap at the sanctuary prior to release predict orphan cohesion post-release (Q1), or whether cohesion changed with time post-release (Q2), we determined inter-individual distances between tracked orphan pairs and orphan–resident pairs using the Conspecific Proximity tool in the ArcMET 10.6.1 (Movement Ecology Tools for ArcGIS) package and ArcGIS 10.7.1. This tool calculates inter-distances, or spatial proximities, of two different animals that have temporally overlapping datasets, i.e., fixes (Wall 2014). Proximities between orphans were calculated at 30-min resolution. Proximities between orphans and resident elephants were calculated at 1-h resolution given resident elephants had collars on an hourly fix schedule. Daily inter-individual distances were calculated as the 24-h means of these proximities, whether at 30-min intervals (orphan–orphan pairs) or 1-h intervals (orphan–resident pairs). There was negligible GPS drift; therefore, proximity calculations were either on the hour or on the half hour. Trajectories are drawn by connecting successive GPS fixes using straight lines. For each output line segment, standard metrics such as speeds and headings were derived (Wall 2014). We applied temporal and spatial filters to the GPS data to exclude GPS fixes collected before calves were translocated and released into Sera Rhino Sanctuary, fixes prior to collar deployment, fixes after collar failures, and fixes for the two resident elephants (Serteta and Kaingus) after they were driven out of the sanctuary. For each pair of calves that were GPS tracked post-release and focal followed pre-release (Npairs = 10), we calculated the proportion of fixes in their first 100 days of temporal overlap at the release site during which they were within communication distance (1.5 km) of one another (Baotic et al. 2018). Given the relational nature of the data, we then conducted Mantel tests with the “Spearman” method and 1000 permutations out of the distance matrix of these proportions with (1) the distance matrix of the rates of social interaction of pairs pre-release, and (2) the distance matrix of the time that pairs overlapped at the sanctuary pre-release (“distance matrix” refers to distances between variables, not spatial distances). Mantel tests were run using the vegan package in R v.3.5.1 (R Development Core Team 2010; Oksanen et al. 2020).
To investigate whether orphan cohesion with other orphans and with residents changes as a function of time (Q2), we classified each mean daily inter-individual distance as whether (1) or not (0) it was within communication distance (≤ 1.5 km). We then used this binary classification as a response variable in logistic regression models treating pairs of elephants as a random effect. The number of days that the pair overlapped post-release and whether (1) or not (0) they were from the same release cohort (for the orphan models) were included as predictor variables. Because these data are a time series with the potential for temporal autocorrelation, we used AIC (Burnham and Anderson 2002) to compare two models for each analysis: one including a first order autoregressive covariance term and one without. We report the top model from each of the orphan–orphan and orphan–resident sets as the model with the lowest AIC value, which in both cases was the model including the autoregressive term. Models were coded using the glmmTMB package (Magnusson et al. 2017) and checked using the DHARMa package in R (Hartig 2021). Unless otherwise indicated, plots were drawn using the ggplot2 package (Wickham 2016).
Results
Pre-release data and post-release pairwise distance
Pre-release interaction rates between orphans excluding the first cohort (Npairs = 21) ranged across an order of magnitude from 0.005 to 0.064 interactions/min (mean: 0.024, inter-quartile range: 0.014–0.033; Fig. 3). Temporal overlap at RES pre-release (all orphan pairs, Npairs = 45) ranged from 570 to 1167 days (mean: 819.511, inter-quartile range: 693–878). The proportion of fixes within the first 100 days of co-presence at the sanctuary that tracked orphan–orphan pairs spent within 1.5 km of one another (Npairs = 10) ranged from 0.383 to 1 (mean: 0.641, inter-quartile range: 0.416–0.969). Mantel tests on the subset of calves for which pre-release interaction rates and tracking distances were available (Npairs = 10) demonstrated a significantly positive correlation between the matrices representing pre-release interaction rates and the proportion of close fixes (r = 0.288, p < 0.05; Fig. 3), indicating pairs with higher interaction rates tended also to be in closer proximity (higher proportion of fixes within communication distance). We found no significant correlation between the temporal overlap at RES pre-release and the proportion of close fixes matrices (r = 0.129, p = 0.176). These trends should be interpreted with caution given the small sample size and may have been affected by a few high rates of aggression/dominance among three elephants that were closely associated post-release (Appendix Fig. A2).

Interaction network (left) for C2 and C3 based on focal follow data collected at Reteti prior to release. Thicker lines (edge weights) represent higher rates of interaction. Nodes are sized by age, and color distinguishes males (orange) and females (blue). The plot on the right relates interaction rates pre-release to the proportion of fixes in the first 100 days of overlap at the release site that were within 1.5 km of one another. The network figure was drawn in Gephi 0.9.1 (Bastian et al. 2009)
Orphan–orphan inter-individual distances tended to be smaller than orphan–resident inter-individual distances (Fig. 4). The period after the first cohort was released was characterized by strong orphan–orphan cohesion; most orphan–orphan inter-individual distances within the first cohort were within a couple of kilometers. There was greater variation in inter-individual distances after the second cohort was introduced, with greater representation at larger distances. This pattern continued after the third cohort was introduced, though orphan–orphan distances were primarily low during all three periods. Orphan–resident density plots represent an increase in the number of inter-individual distances at the low end between the first two periods, and a slight decrease at the low end between the second and third periods.

Ridge density plots of inter-individual distances for all available pairs of elephants based on tracking data. Plots are divided into three periods ordered from the bottom to the top: between the first release and the second release, between the second and third releases, and after the third release
There was considerable variation within the orphan–orphan and orphan–resident categories. While some orphan pairs have demonstrated very strong within-cohort cohesion (Nadosoit-Nchurai, Shaba-Pokot), others have exhibited fissions over the monitored period (Fig. 5). Strong cohesion within the first cohort decreased toward the end of the monitored period, potentially as a result of a greater number of orphans in the sanctuary (Fig. 6). However, tracked orphans always seem to have been with at least one other orphan, and shuffling between groups typically occurred between groups containing other orphans. In the orphan–orphan model, days of overlap at the release site was not significantly related to the probability of a pair of orphans being within 1.5 km of one another (β = 0.000, SE = 0.006, p = 0.971; Fig. 7), whereas being in the same cohort was significantly positively related to the probability of being within 1.5 km (β = 11.285, SE = 1.408, p < 0.001), indicating that cohort membership predicts proximity and that orphans did not detectably disperse further than communication distance over the time period analyzed.

Orphan–orphan pairs exhibited distinct patterns in inter-individual distances. While some within-cohort pairs have remained consistently close to one another (represented here by Pokot and Shaba from the second cohort; left), others have established patterns early on indicative of fission and fusion (represented here by Baawa and Nadosoit from the third cohort) while coordinating movement with other cohorts (represented by Baawa in the third cohort and Pokot in the second cohort; right). Note differences in y-axis scales

Ilngwesi and Sosian, from the first release cohort, remained strongly cohesive for over a year following release. Sosian and Ilngwesi began to spend time apart within the second year post-release. Vertical dotted lines demarcate the second and third releases into the sanctuary

Predicted probabilities of tracked elephant pairs being within communication distance of one another for (left) orphan–orphan pairs and (right) orphan–resident pairs indicated no predicted change over time for orphan–orphan pairs and a significant increase over time for orphan–resident pairs. Shaded areas represent 95% confidence levels
Among the collared resident elephants, orphans showed the smallest distances with the adult female Kalama (Fig. 8; Appendix Fig. A3). However, there was a strong release cohort effect, with the calves in the first cohort settling into a fission–fusion pattern with Kalama, manifested as multiple days with near zero proximities punctuated by temporary periods of larger distances that has not been apparent among the latter two release cohorts. In the orphan–resident model, days of overlap was significantly positively related to being within 1.5 km among pairs (β = 0.009, SE = 0.002, p < 0.001), indicating closer proximity over time (Fig. 7).

a Orphans from C1 (represented by Ilngwesi) over time settled into a predictable fission–fusion pattern with the resident adult female elephant Kalama. Such a pattern has thus far not been apparent with the b C2 (represented by Shaba) and c C3 (represented by Nadosoit) release cohorts
Discussion
While a critical tool used for the conservation of many threatened and endangered species, translocations involve social upheavals for individual animals that necessitate investment in new relationships. They may also represent a disruption in maternal and familial bonds pre-weaning that are critical to development. Whether translocated animals integrate within resident populations, how long this process takes, and whether there are predictable conditions or cohort structures that facilitate this process are important concepts to understand when translocating species that depend on established social relationships to survive. Such understanding will stem from detailed and long-term monitoring of individuals pre- and post-release (Berger-Tal et al. 2020). As long-lived species that exhibit cohesive extended family structure and lifelong bonds, elephants are dependent on established social relationships; elephant translocations will, therefore, be improved by extracting insights gained through monitoring the social integration process post-release, and iteratively applying findings to establish conditions that support the development of appropriate social relationships (Slotow et al. 2005; Pinter-Wollman et al. 2009; Goldenberg et al. 2019).
Our study of three cohorts of rehabilitated and released African elephant calves into a soft release site in northern Kenya demonstrated that social interactions pre-release are related to proximity during the first 100 days post-release, that release cohort membership predicts proximity post-release, and that orphan–resident proximity increases over time post-release (Table 1). Our preliminary findings contribute to conservation translocation knowledge by highlighting the importance of existing relationships to the transitional period post-release, and by documenting the length of time needed for signs of social integration and fission–fusion patterns to emerge. Analyses into the exploratory movement behavior of these calves indicates that social association with more knowledgeable released calves and with residents may be critical to facilitate knowledge of the landscape (Goldenberg et al. 2021), which in turn may influence calf foraging efficiency and body condition. Detailed understanding of the development of such relationships as conducted here may illuminate the ways in which relationships among elephants may be leveraged to improve post-release outcomes. This research will be deepened by continued monitoring of this population and comparison with similar populations of released calves throughout their range. Further, our results highlight the value of GPS tracking to monitor released animals where minimal interaction with humans post-release is desired: in addition to providing insight into landscape exploration and resource use (Goldenberg et al. 2021), tracking collars on both released calves and resident elephants allowed us to infer social relationships and relationship progression remotely.
Our comparison of social interactions and temporal overlap at the orphanage pre-release to the proportion of fixes within communication distance (1.5 km) of one another over the first 100 days of overlap at the post-release site indicated that social interaction pre-release may be predictive of social cohesion post-release (Dunston et al. 2016). In contrast, temporal overlap at RES pre-release, which may be thought of as a more general measure of familiarity, was not correlated with this proportion of close fixes. These results underscore the value of pairwise relationships and the value in defining them using fine scale social data on animals slated for release (Dunston et al. 2016). The nature of the relationship between calves and their post-release outcomes (e.g., whether pairs with high rates of aggression/dominance between them experience discrepancies in body condition even if they remain in close proximity) is beyond the scope of this study and should be further investigated; our results point to the importance of continuous monitoring of fine scale interactions among calves and may yield greater insights later relating to specific types of interactions.
If close proximity among released pairs post-release is a desired management outcome, e.g., to facilitate predation avoidance or knowledge exchange about the new environment (Poirier and Festa-Bianchet 2018; Goldenberg et al. 2021), information about existing relationships can be leveraged to select release cohorts, or to facilitate interactions among release cohorts. It is possible that Baawa, in the third release cohort, may have facilitated interactions among the second and third release cohorts as a result of his strong existing relationship with Pokot, a similar aged male in the previous release cohort. Facilitating interactions of recently released orphans with orphans that have had time to accrue knowledge of the release site may be an important factor to consider as it may take time for orphans to integrate with wild residents (Figs. 7, 8), and ecological knowledge gained from associations with other elephants is likely essential for avoiding predation and locating resources (Kuiper et al. 2018; Goldenberg et al. 2021). However, we note that these results should be interpreted with caution as the small sample size of individuals whose interactions were documented pre-release did not allow us to control for the effects of age and sex on pairwise interactions. Additionally, to date the released calves all overlapped considerably at Reteti. Temporal overlap at the sanctuary regardless of interaction rate may be a more important factor in predicting association post-release if there is a larger range of temporal overlap within the sample. This will be worth investigating in the future as more calves move through the rescue, rehabilitation, soft release, and hard release process, as will the influence of socially central individuals in facilitating cohesion or bridge-building post-release (Snijders et al. 2017; Goldenberg et al. 2019).
When investigating whether the probability of orphans being within communication distance of one another changed over time post-release, we found no temporal relationship, with tracked orphans usually within 1.5 km of each other regardless of time post-release. The same model indicated a strong positive relationship between being in the same cohort and the probability of being within 1.5 km. Together, these results highlight the importance of orphan–orphan relationships upon release. While there were exceptions, cohesion within cohorts was strong, and calves tended to stay together for extended periods. Released calves remaining cohesive with other released calves—or at least staying nearby—may be an important stepping stone as they navigate their new social and ecological environment (Pinter-Wollman et al. 2009; Kuiper et al. 2018).
In contrast to the orphan–orphan model, our model for orphan–resident pairs found a significant positive relationship between time overlapping at the release site and the probability that orphan–resident pairs were within 1.5 km of one another, such that orphans became closer to residents over time. This result indicates a gradual process of association with the resident population, though distances were smaller among orphan–orphan pairs as compared to orphan–resident pairs (Fig. 4). An increase in association with residents is consistent with results reported from another translocation of a large group of African elephants (Pinter-Wollman et al. 2009) and with the release of a captive-reared female African elephant (Evans et al. 2013). While this trend suggests that orphan calves become more integrated with resident elephants over time, it is important to note that proximity should not necessarily be equated with social integration. Relationships in elephants may take years to establish following an altered social environment (Goldenberg et al. 2016), and orphans may experience more difficult circumstances even if they are cohesive with a family (Goldenberg and Wittemyer 2017, 2018; Parker et al. 2021). A monitored female elephant that was released from captivity in Botswana was observed socializing with wild herds but remained on the periphery of those groups (Evans et al. 2013). It is possible that released elephants stay in close proximity with limited interaction as a way to gradually integrate over extended periods of time, which may still provide important opportunities for learning from wild counterparts, particularly if they are staying within communication distance. In the future, we will combine inter-individual distances derived from GPS tracking with camera trapping imagery and vehicle-based observations to provide finer scale detail into the types of interactions elephants are having with one another, as well as any interactions among elephants that are not GPS tracked.
Here, as in other wildlife translocations, it is critical to iteratively evaluate management protocols in order to improve future conservation outcomes (Berger-Tal et al. 2020). Although the release of orphaned elephant calves into the Sera Conservancy is still in its early stages and represents the soft release stage prior to hard release out of the fenced sanctuary, social integration patterns are emerging that will contribute to the formulation of benchmarks of social behavior that can be used to guide management decisions, assess the post-release performance of individuals, and assess the success of this conservation translocation program and others like it (International Union for Conservation of Nature 2013). First, within-cohort and among-cohort relationships are clearly important in the initial period post-release when translocated animals are learning their new environment. Data collected on behavior pre-release may inform cohort composition and timing of releases between cohorts. For example, dyadic interaction rates could be an indicator for release, and specific pairings or individuals with particular social tendencies may be used to foster cohesion within or among cohorts, or to bridge release cohorts separated over longer periods of time by releasing bonded individuals in separate cohorts (Dunston et al. 2016; Goldenberg et al. 2019).
Second, time periods prior to integration with resident elephants may be long, and may lengthen as the ratio of released calves to residents increases. The first cohort of released calves experienced a period of over 150 days during which there was no discernable pattern in their inter-individual distances with the adult female Kalama (Fig. 8). Following that point, however, they formed an apparent fission–fusion pattern. An expected length of such initial periods before released calves manage to close the gap with residents may be a useful metric for determining whether they have integrated to some degree with the resident population, as has been investigated in other species translocations (Poirier and Festa-Bianchet 2018). However, we have not yet seen any such behavior with calves from the second and third cohorts, which may indicate that there is a threshold number of orphans that may be absorbed into resident families or associate frequently with bulls. This should be kept in mind when release sites are chosen, as the ability of translocated animals to learn from residents may be key to establishment at a release site, though we note it is still possible to learn from conspecifics even if not fully integrated into wild groups. Absorption into families may be harder to achieve if there is a high ratio of translocated to resident animals. Nonetheless, indicators of increased social interactions with residents, even if it is not complete integration within a family, may be useful to gauge readiness to move from soft release within the fenced area to hard release into the larger landscape. Such changes over time may suggest that calves are learning skills to navigate their social environment.
A third metric that may be useful for defining social benchmarks of success in elephant release projects may be the occurrence and increased incidence of societal fission–fusion (Kuiper et al. 2018). Under natural conditions elephants separate from and join with others for varying lengths of time. Separations are driven by seasonal resource competition, and fusions allow elephants to maintain relationships of different types that may be beneficial (Douglas-Hamilton 1972; Moss 1988; Wittemyer et al. 2005). One such social benefit that may be exceptionally important for translocated elephants learning a new environment is information exchange (McComb et al. 2000, 2001). Increased instances of regular fission–fusion patterns, and the occurrence of such instances with an increasing number of individuals, may reflect the growth of a translocated calf’s information network and their settling into a social pattern reflective of fluctuating ecological conditions. Further research into the development of fission and fusion patterns is warranted as monitoring continues.
Post-release monitoring is critical to improving outcomes of wildlife conservation translocations, particularly as such projects tend to be resource intensive and have considerable potential for failure (International Union for Conservation of Nature 2013; Berger-Tal et al. 2020). Functional social behavior may be an underappreciated aspect of the survival of translocated animals (Shier and Swaisgood 2012; Goldenberg et al. 2019; Franks et al. 2020). Thus, documenting social behavior pre- and post-release, as well as the behavior of residents at release sites (McKnight 1995; Pinter-Wollman et al. 2009; Evans et al. 2013; Dunston et al. 2016; Poirier and Festa-Bianchet 2018), may provide managers with insight to hone protocols and expectations in an iterative fashion as new results emerge. The work presented here from the soft release stage for three elephant calf cohorts lays a foundation for understanding the processes these socially complex and individualistic animals undergo as they settle into a novel environment. Continued monitoring, incorporation of other types of data that quantify health and physiology, and comparison with other projects to standardize benchmarks and characterize the full range of behavior exhibited will enrich our understanding further.
References
Altmann J (1974) Observational study of behavior: sampling methods. Behaviour 49:227–267
Archie EA, Morrison TA, Foley CAH, Moss CJ, Alberts SC (2006) Dominance rank relationships among wild female African elephants, Loxodonta africana. Anim Behav 71:117–127
Baotic A, Garcia M, Boeckle M, Stoeger A (2018) Field propagation experiments of male African savanna elephant rumbles: a focus on the transmission of formant frequencies. Animals 8:167
Bastian M, Heymann S, Jacomy M (2009) Gephi: an open source software for exploring and manipulating networks. In: Third international AAAI conference on weblogs and social media
Berger-Tal O, Saltz D (2014) Using the movement patterns of reintroduced animals to improve reintroduction success. Curr Zool 60:515–526
Berger-Tal O, Blumstein DT, Swaisgood RR (2020) Conservation translocations: a review of common difficulties and promising directions. Anim Conserv 23:121–131
Burnham KP, Anderson DR (2002) Model selection and multi-model inference: a practical information-theoretic approach. Springer, Berlin
Campos-Arceiz A, Blake S (2011) Megagardeners of the forest—the role of elephants in seed dispersal. Acta Oecol 37:542–553
Cheyne S (2009) Challenges and opportunities of primate rehabilitation—gibbons as a case study. Endang Species Res 9:159–165
Douglas-Hamilton I (1972) On the ecology and behaviour of the African elephant
Dunston EJ, Abell J, Doyle RE, Kirk J, Hilley VB, Forsyth A, Jenkins E, Freire R (2016) An assessment of African lion Panthera leo sociality via social network analysis: prerelease monitoring for an ex situ reintroduction program. Curr Zool 63:301–311
Engh AL, Beehner JC, Bergman TJ, Whitten PL, Hoffmeier RR, Seyfarth RM, Cheney DL (2006) Behavioural and hormonal responses to predation in female chacma baboons (Papio hamadryas ursinus). Proc R Soc B Biol Sci 273:707–712
Evans K, Moore R, Harris S (2013) The social and ecological integration of captive-raised adolescent male African elephants (Loxodonta africana) into a wild population. PLoS ONE 8:18–21
Foley C, Pettorelli N, Foley L (2008) Severe drought and calf survival in elephants. Biol Lett 4:541–544
Franks VR, Andrews CE, Ewen JG, McCready M, Parker KA, Thorogood R (2020) Changes in social groups across reintroductions and effects on post-release survival. Anim Conserv 23:443–454
Goldenberg SZ, Wittemyer G (2017) Orphaned female elephant social bonds reflect lack of access to mature adults. Sci Rep 7:14408
Goldenberg SZ, Wittemyer G (2018) Orphaning and natal group dispersal are associated with social costs in female elephants. Anim Behav 143:1–8
Goldenberg SZ, Douglas-Hamilton I, Wittemyer G (2016) Vertical transmission of social roles drives resilience to poaching in elephant networks. Curr Biol 26:75–79
Goldenberg SZ, Owen MA, Brown JL, Wittemyer G, Oo ZM, Leimgruber P (2019) Increasing conservation translocation success by building social functionality in released populations. Glob Ecol Conserv 18:e00604
Goldenberg SZ, Hahn N, Stacy-Dawes J, Chege SM, Daballen D, Douglas-Hamilton I, Lendira RR, Lengees MJ, Loidialo LS, Omengo F, Pope F, Thouless C, Wittemyer G, Owen MA (2021) Movement of rehabilitated African elephant calves following soft release into a wildlife sanctuary. Front Conserv Sci 2:720202
Hartig F (2021) DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models
International Union for Conservation of Nature (2013) Guidelines for reintroductions and other conservation translocations
Kelly A, Scrivens R, Grogan A (2010) Post-release survival of orphaned wild-born polecats Mustela putorius reared in captivity at a wildlife rehabilitation centre in England. Endang Species Res 12:107–115
Kuiper TR, Druce DJ, Druce HC (2018) Demography and social dynamics of an African elephant population 35 years after reintroduction as juveniles. J Appl Ecol 55:2898–2907
Louys J, Corlett RT, Price GJ, Hawkins S, Piper PJ (2014) Rewilding the tropics, and other conservation translocations strategies in the tropical Asia-Pacific region. Ecol Evol 4:4380–4398
Magnusson A, Skaug HJ, Nielsen A, Berg CW, Kristensen K, Maechler M, van Bentham KJ, Bolker BM, Brooks ME (2017) glmmTMB: generalized linear mixed models using template model builder. R package version 013
McComb K, Moss C, Sayialel S, Baker L (2000) Unusually extensive networks of vocal recognition in African elephants. Anim Behav 59:1103–1109
McComb K, Moss C, Durant SM, Baker L, Sayialel S (2001) Matriarchs as repositories of social knowledge in African elephants. Science 292:491–494
McKnight BL (1995) Behavioural ecology of “hand-reared” African elephants (Loxodonta africana (Blumenbach)) in Tsavo East National Park, Kenya. Afr J Ecol 33:242–256
Moss CJ (1988) Elephant memories: thirteen years in the life of an elephant family. University of Chicago Press, Chicago
Moss CJ (1996) Getting to know a population. In: Kangwana K (ed) Studying elephants. African Wildlife Foundation, Nairobi, pp 58–74
Nunez CMV, Adelman JS, Rubenstein DI (2015) Sociality increases juvenile survival after a catastrophic event in the feral horse (Equus caballus). Behav Ecol 26:138–147
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner (2020) vegan: community ecology package
Parker JM, Webb CT, Daballen D, Goldenberg SZ, Lepirei J, Letitiya D, Lolchuragi D, Leadismo C, Douglas-Hamilton I, Wittemyer G (2021) Poaching of African elephants indirectly decreases population growth through lowered orphan survival. Curr Biol 31:4156–4162
Perera BV, Jayawardena BADS, Kumudini N (2016) Success of rehabilitation of orphaned elephant (Elephas maximus) calves in Sri Lanka, vol 21, pp 177–178
Pinter-Wollman N, Isbell LA, Hart LA (2009) The relationship between social behaviour and habitat familiarity in African elephants (Loxodonta africana). Proc R Soc B 276:1009–1014
Poirier M-A, Festa-Bianchet M (2018) Social integration and acclimation of translocated bighorn sheep (Ovis canadensis). Biol Cons 218:1–9
R Development Core Team (2010) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. www.R-project.org
Rasmussen HB (2005) Reproductive tactics of male African savannah elephants (Loxodonta africana). Oxford University
Seddon PJ, Griffiths CJ, Soorae PS, Armstrong DP (2014) Reversing defaunation: Restoring species in a changing world. Science 345:406–412
Shannon G, Slotow R, Durant SM, Sayialel KN, Poole J, Moss C, McComb K (2013) Effects of social disruption in elephants persist decades after culling. Front Zool 10:62
Shier DM (2006) Effect of family support on the success of translocated black-tailed prairie dogs. Conserv Biol 20:1780–1790
Shier DM, Swaisgood RR (2012) Fitness costs of neighborhood disruption in translocations of a solitary mammal. Conserv Biol 26:116–123
Slotow R, Garaï ME, Reilly B, Page B, Carr RD (2005) Population dynamics of elephants re-introduced to small fenced reserves in South Africa. S Afr J Wildl Res 35:10
Snijders L (2020) Ecological and evolutionary dynamics of elephant rewilding. Anim Sentience 311:6
Snijders L, Blumstein DT, Stanley CR, Franks DW (2017) Animal social network theory can help wildlife conservation. Trends Ecol Evol 32:567–577
Thitaram C, Dejchaisri S, Somgird C, Angkawanish T, Brown J, Phumphuay R, Chomdech S, Kangwanpong D (2015) Social group formation and genetic relatedness in reintroduced Asian elephants (Elephas maximus) in Thailand. Appl Anim Behav Sci 172:52–57
Vidya TNC, Varma S, Dang NX, Van Thanh T, Sukumar R (2007) Minimum population size, genetic diversity, and social structure of the Asian elephant in Cat Tien National Park and its adjoining areas, Vietnam, based on molecular genetic analyses. Conserv Genet 8:1471–1478
Wall J (2014) Movement ecology tools for ArcGIS® (ArcMET)
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer, New York
Wittemyer G, Douglas-Hamilton I, Getz W (2005) The socioecology of elephants: analysis of the processes creating multitiered social structures. Anim Behav 69:1357–1371
Wittemyer G, Getz WM, Vollrath F, Douglas-Hamilton I (2007) Social dominance, seasonal movements, and spatial segregation in African elephants: a contribution to conservation behavior. Behav Ecol Sociobiol 61:1919–1931
Wittemyer G, Okello JBA, Rasmussen HB, Arctander P, Nyakaana S, Douglas-Hamilton I, Siegismund HR (2009) Where sociality and relatedness diverge: the genetic basis for hierarchical social organization in African elephants. Proc R Soc B Biol Sci 276:3513–3521
Acknowledgements
We thank the Kenyan Office of the President, the Kenya Wildlife Service, and the Sera and Namunyak communities for permission to conduct this research. We are grateful to the staff of the Reteti Elephant Sanctuary, the rangers of Sera Conservancy, and the Save the Elephants monitors in Sera for their conservation efforts that make this work possible. Funding for this work was provided by the Harriet E. Pfleger Foundation, the W T Yoshimoto Foundation Charitable Trust, and other donors to San Diego Zoo Wildlife Alliance and Save the Elephants.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Handling editors: Scott Y.S. Chui and Leszek Karczmarski.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue on “Individual Identification and Photographic Techniques in Mammalian Ecological and Behavioural Research – Part 2: Field Studies and Applications” — Editors: Leszek Karczmarski, Stephen C.Y. Chan, Scott Y.S. Chui and Elissa Z. Cameron.
Supplementary Information
Below is the link to the electronic supplementary material.
Appendix
Appendix

Photo credit: Shifra Goldenberg
Elephant calves frequently interact with one another, as with these calves touching their trunks to each other’s mouths.

Pairwise interaction rates of the six calves that were both focal followed pre-release and GPS tracked post-release. Pairs differed in the extent to which higher rates of affiliative interactions were correlated with higher rates of aggressive/dominance interactions. Because elephants navigate relationships with one another using all types of interactions, these were combined in analyses

Inter-individual distances between the other adult resident female elephant collared within the sanctuary throughout the study, Lpupo, and a member of each release cohort (a C1; b C2; c C3). In contrast to Kalama, no released calves established pattern of association with Lpupo over the study period
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Goldenberg, S.Z., Chege, S.M., Mwangi, N. et al. Social integration of translocated wildlife: a case study of rehabilitated and released elephant calves in northern Kenya. Mamm Biol 102, 1299–1314 (2022). https://doi.org/10.1007/s42991-022-00285-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-022-00285-9