
Abstract
Little Bahama Bank in the northern Bahamas supports several populations of bottlenose dolphins (Tursiops truncatus). We provide the first estimates of birth rate and age-class-specific apparent survival rates for the local South Abaco population using data from a long-term (1997–2014) photo-identification (photo-ID) study and use the estimated life history parameters in a population viability analysis (PVA) to predict future population trends. Hurricane events are predicted to become more intense due to climate change but knowledge of how hurricanes may impact cetacean populations is limited. Little Bahama Bank is subject to hurricane activity, so we also investigate the potential impact of hurricanes on calf, juvenile and adult survival. Photo-ID data confirmed the existence of a core adult population with relatively high site fidelity in South Abaco, but also evidence of transient animals. Estimated annual birth rate was 0.278 (95% CI: 0.241–0.337). We found strong support for a decline in apparent survival for all age-classes. Estimated survival declined by 9% in adults (0.941 in 1998, to 0.855 in 2013), 5% in juveniles (0.820 in 2000, to 0.767 in 2013) and 36% in calves (0.970 in 1997, to 0.606 in 2013). Evidence that survival was influenced by repeated hurricane activity leading to increased mortality and/or emigration was stronger for calves and juveniles than for adults. PVA simulations of an assumed isolated South Abaco population showed that declines would lead to extinction within decades, even under the most optimistic scenario. Future work should focus on establishing if South Abaco is part of natural source–sink metapopulation dynamics on Little Bahama Bank by assessing trends in abundance in local populations and establishing how they interact; this will be important for assessing their conservation status in a potentially increasingly changing environment.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Knowledge of birth and survival rates of populations is needed to describe a species’ life history but can also be important in the context of conservation. If species monitoring indicates a decline in abundance, unless the cause is already clear (e.g. Taylor et al. 2017), estimates of birth and survival rates can help identify possible reasons for the decline (e.g. Currey et al. 2011). Investigation of how these rates have varied over time may also provide insight into the influence of environmental changes (e.g. Agrelo et al. 2021; Jourdain et al. 2021). Incorporating population-specific birth and survival rates in population dynamics models, such as a population viability analysis, allows population growth rates to be estimated and inferences made about future conservation action that may be needed to halt a decline in abundance (e.g. Bezamat et al. 2021).
Little Bahama Bank (surrounding Great Abaco and Grand Bahama Islands) in the northern Bahamas supports several populations of bottlenose dolphins (Tursiops truncatus), totalling an estimated 1100 individuals (Durban 2002). Due to the abrupt drop-off in water depth immediately offshore of Little Bahama Bank, the overall population is believed to be closed, with no immigration or emigration occurring (Parsons et al. 2006). Within this overall population, three genetically distinct local populations have been described, although some gene flow (via both females and males) is evident (Parsons et al. 2006). These are the East Abaco population (east of Great Abaco; Fearnbach et al. 2012), the Northern community (north of Grand Bahama and encompassing the White Sand Ridge region; Rossbach and Herzing 1999; Rogers et al. 2004) and the South Abaco population, which forms the focus of the current study. A fourth population (the Southern community) has also been identified off the north-west coast of Grand Bahama (Rossbach and Herzing 1999; Rogers et al. 2004). The distinctness of the local populations combined with some interchange among them indicates that bottlenose dolphins on Little Bahama Bank may form a metapopulation.
In this paper, we analyse data from a long-term photo-identification study to estimate the birth rate and age-class-specific apparent survival rates of bottlenose dolphins in South Abaco. We use this new information on life history parameters to explore the future prospects of this local population using a population viability analysis (Brook et al. 2000; Thompson et al. 2000; Manlik et al. 2016). Results from these analyses may help inform the extent to which it is reasonable to consider the bottlenose dolphins on Little Bahama Bank as a metapopulation.
Climate change appears to be contributing to the observed increase in intensity and frequency of at least some forms of extreme weather events (e.g. National Academy of Sciences 2020). Evidence suggests that the intensity of hurricanes, one form of extreme weather, is increasing and that this is likely to be due to global warming (e.g. Knutson et al. 2021). Hurricanes have been linked to negative environmental consequences in a variety of ecosystems (e.g. Tanner et al. 1991; Gardner et al. 2005) and also to impacts on species, including permanent or temporary emigration (e.g. Preen and Marsh 1995), increases in habitat range (e.g. White et al. 2005), and abundance declines due to potential reductions in particular food resources (e.g. Wauer and Wunderle 1992). In marine mammals, hurricane activity may impact survival (Preen and Marsh 1995; Langtimm and Beck 2003), reproduction (Miller et al. 2010), foraging (Smith et al. 2013), social structure (Elliser and Herzing 2011) and stranding events (Mignucci-Giannoni et al. 2000).
A future increase in the proportion of high intensity storms (Category 4 and 5 on the Saffir-Simpson Hurricane Wind Scale; NOAA 2021a) is predicted to be more likely than an increase in the frequency of hurricane events (NOAA GFDL 2021). However, in the northern Bahamas, there has been a marked increase in hurricane events in the South Abaco area following the passage of Hurricane Andrew in 1992 (NOAA 2021b), including, recently, Hurricane Dorian, a Category 5 hurricane which directly hit Great Abaco and Grand Bahama in 2019. Before Hurricane Andrew in 1992, the last hurricane to impact South Abaco was Hurricane Betsy in 1965. Several studies have documented potential effects of hurricane activity on bottlenose dolphins in The Bahamas (Elliser and Herzing 2011; Fearnbach et al. 2011, 2012), but knowledge of how hurricanes may affect cetacean survival is limited. Thus, we also explore whether hurricanes may have influenced the apparent survival rates of calves, juveniles or adults in the South Abaco bottlenose dolphin population, and which features of hurricanes (frequency, intensity, proximity) may best explain any effect.
Methods
Data collection and photographic identification
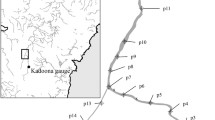
Boat-based photo-identification surveys were conducted between 1997 and 2014 off the west coast of South Abaco, within the waters of Little Bahama Bank (northern Bahamas), in an estimated 50 km2 study site (Fig. 1; Fig. A1). This site was located between the north of Gorda Cay (approx. 26°20′ N, 77°62′ W) and the south of Cross Harbour (approx. 25°90′ N, 77°25′ W). Photo-identification survey equipment and protocols were similar to those documented in Fearnbach et al. (2012), with surveys conducted between January and October every year, with increased effort during the summer months (June to August). Photographs of individual dorsal fins were graded for photographic quality following the method outlined by Durban et al. (2000) based on image size, focus, angle and percentage of the fin visible in the frame. Only good-quality photographs were used in analysis. All photographed individuals were identified based on the unique markings on their dorsal fin and matched against a catalogue of known bottlenose dolphins from this population.

Calving data and birth rate analysis
A dataset containing calving histories of individual females for the years 1996–2013 was constructed. The year 1996 was included because some calves were identified in 1997 (when photo-identification effort started) as 1-year-olds based on their relative size and appearance. A confidence level of 1 (certain) or 2 (uncertain) was assigned to both year of birth and mother-ID in the dataset. Certainty of mother-ID was based on the criteria that mother and calf were seen together, with calf in echelon position, on more than two unique days (e.g. Arso Civil et al. 2017). Certainty of year of birth was based on a combination of ageing of the calf based on body size, skin coloration, presence of foetal folds, field observations and expert opinion by DC based on the long-term study of individuals in this population.
Only those mother–calf pairs that had a confidence of 1 (certain) for mother-ID were included in the birth rate analysis. However, calves with either a confidence 1 (certain) or 2 (uncertain) for year of birth were included because, for this analysis, a 1-year error in birth date (the maximum possible) is less problematic than a false lengthening or discontinuation of an inter-birth interval. Three females considered part of the East Abaco population (BMMRO unpublished data) but seen occasionally with calves in South Abaco were excluded from the analysis.
Mean inter-birth interval (IBI; time in years between births) and annual birth rate (probability of a female having a calf in a year) were estimated for this population following the methods described in Arso Civil et al. (2017). In summary, generalised linear mixed models (GLMM) were used to model the probability of a female giving birth as a function of the number of years since the previous birth and/or the quadratic form of this covariate included as fixed effects, and mother-ID and/or year included as random effects. As in Arso Civil et al. (2017), it was assumed that females do not give birth in consecutive years. This assumption was based on information collected over the long-term photo-identification study in South Abaco in which there is no evidence for females giving birth in consecutive years (BMMRO unpublished data). All models assumed binomial error in the response and used a logit link. Analysis was undertaken in R Version 4.1.1 (R Core Team 2021), with package lme4 (Bates et al. 2015). Support from the data for each model was based on Akaike’s Information Criterion (AIC).
Estimating apparent survival
Open-population capture–recapture Cormack–Jolly–Seber (CJS) models (Lebreton et al. 1992) were used to estimate apparent survival probabilities of calves, juveniles and adults.
Calf and juvenile survival
To estimate calf and juvenile survival, annual capture histories of all individuals with known year of birth (with confidence 1, see above) were generated. Calves and juveniles in the South Abaco population can be tracked with confidence as they age using either temporary or permanent marks, as well as identified via mother–calf association. Age-class models were fitted to estimate the probability of survival of calves (defined here as 0–2 years) and juveniles (defined here as 3–8 years). The age of sexual maturity, i.e. exiting the juvenile age-class, was set at 9 years based on mid-value estimates for female bottlenose dolphins in Wells and Scott (2018). Two age-class structures were investigated, in which calf survival was modelled as either (1) non-age-specific, i.e. a single parameter for all ages, or (2) age-specific, i.e. separate parameters for first-, second- and third-year survival. Juvenile survival was modelled as non-age-specific in both age-class structures.
Goodness of fit tests of CJS model assumptions about apparent survival probabilities and recapture probabilities (Lebreton et al. 1992), were run using package R2ucare (Gimenez et al. 2018) in R Version 4.1.1 (R Core Team 2021), following the schematic in Figure 1 in Gimenez et al. (2018). The overall test was not significant (χ235 = 30.80, p = 0.67) indicating no lack of fit, including no overdispersion in the data.
Survival (Phi) was modelled as age-class-specific (calves and juveniles, as described above), and was also allowed to vary annually (time) or with a linear time trend (Time). The modelled relationships between age-class and time or Time were allowed to be additive (a common effect of time or Time on both age-classes) or interactive (separate effects of time or Time on each age-class). Recapture probability (p) was modelled as constant (.) or varying by year (time). We refer to these models of calf/juvenile survival as base models.
Adult survival
To estimate adult survival, annual capture histories of all individuals that were permanently marked, i.e. animals with dorsal fin nicks, and that were believed to be an adult on first capture were generated. Goodness of fit tests were run as described above. The overall test was highly significant (\({\chi }_{48}^{2}\) = 106.6, p < 0.01) indicating lack of fit of the CJS model. Tests 3.SR and 2.CT were both significant (Test 3.SR: \({\chi }_{12}^{2}\)= 30.01, p < 0.01; Test 2.CT: \({\chi }_{15}^{2}\) = 60.22, p < 0.01), indicating lack of fit likely resulting from a transience effect and a trap dependence effect, respectively. All models of adult survival thus incorporated two features. First, survival probability was modelled by transient-class, in which one class represented survival in the year following first capture and a second class represented survival in all subsequent years. The first class includes so-called transient individuals (captured only once) for which survival will be negatively biased (Pradel et al. 1997), thus only estimates of survival for the second class (non-transients) are subsequently considered. Second, annual recapture probability was modelled to vary between individuals captured or not captured in the previous year, modelled as an individual time-varying covariate (see, e.g. Schleimer et al. 2019). Component tests 3.SR and 2.CT were removed from the overall goodness of fit test (Pradel et al. 2005), which then showed no lack of fit (\({\chi }_{21}^{2}\)= 16.3, p = 0.75), including no evidence of overdispersion in the data.
Survival (Phi) was modelled as transient-class-specific (trans) and was also allowed to vary annually (time) or with a linear time trend (Time). The modelled relationships between trans and time or Time were only allowed to be interactive because in these models there are separate effects on transients and non-transients; we do not want the modelled effect on non-transients to be influenced by transients, as would occur in an additive model. Recapture probability (p) was modelled as trap dependent (td) and was also allowed to vary annually as an additive or interactive relationship with time. We refer to these models of adult survival as base models.
All models assumed binomial error in the response and used a logit link function. Models were run using the R package RMark (Version 2.2.7; Laake 2013), implementing software MARK (Version 9.0; White and Burnham 1999). Support from the data for each model was based on AICc (AIC adjusted for small sample size; Burnham and Anderson 2002).
Modelling the effect of hurricanes on survival
Data on hurricanes affecting the South Abaco study site during this study were gathered from the National Oceanic and Atmospheric Administration (NOAA) National Hurricane Center (NOAA 2021b). These were additionally confirmed in a local context (Table 1). A hurricane was defined as a direct hit if it passed within 50 miles (approx. 80 km) of the South Abaco study site, and non-direct if it passed within 100 miles (approx. 160 km), based on the 100 mile average diameter of hurricane force winds (e.g. Willoughby et al. 2007). Major hurricanes were defined as those that achieved Category 3 (or greater) wind speeds on the Saffir-Simpson Hurricane Wind Scale (NOAA 2021a) over the South Abaco study site; Category 1 or 2 hurricanes were defined as Minor.
To model the effect of hurricanes on apparent survival, a set of covariates with annual values for the period 1997–2014 was compiled. Three types of covariate were created to represent different possible effects on survival (Table 2). Binary covariates categorised years as 1 or 0 to represent the presence or absence of a hurricane. ‘Time since’ covariates categorised years as the time since the last hurricane to investigate the potential lagged effect of hurricanes. Cumulative count covariates categorised years by the number of years with hurricanes since 1992. To investigate whether major and/or direct hurricanes may have had a greater influence on survival, hurricane covariates were created that included (a) only years with hurricanes that were major and/or direct and (b) years with all hurricanes (Table 2). In 2 years, 1999 and 2004, there were two hurricanes, one of which was major and direct (Table 1).
The effect of hurricanes on apparent survival of calves/juveniles and of adults was modelled by incorporating each of the hurricane covariates in Table 2 in the base models described above. For calves/juveniles, hurricane covariates were modelled as additive or interactive relationships with age-class and with time (varying by year) or Time (linear time trend). For adults, hurricane covariates were modelled as additive or interactive relationships with trans and with time or Time. We refer to these models as hurricane effect models.
Population viability analysis (PVA)
Population viability analysis was conducted in software Vortex Version 10 (Lacy and Pollak 2020). The South Abaco population was treated as isolated and independent for the purposes of the PVA. All simulations were run as individual-based models using birth rate and age-class-specific apparent survival rates estimated during this study. Mortality rates for calves, juveniles and adults required as inputs to Vortex were calculated as 1 − survival rates estimated by the most-supported base models. Three scenarios were modelled to explore the impact of different survival rates estimated by these models, which all predicted a declining trend in survival (see “Results”). The Baseline (2005) scenario included survival estimates for calves, juveniles and adults from 2005, a year midway within the study period that thus represents mid-range (approximately average) estimates of survival (Table 3). The High (2000) scenario included survival estimates from 2000, the first year when survival estimates were available for all age-classes, and thus representing high estimated survival (Table 3). The Low (2010) scenario included survival estimates from 2010, avoiding the estimates for the final three years, in which terminal bias could be present (e.g. Schleimer et al. 2019), and thus representing low estimated survival (Table 3). The estimated birth rate was the same in all three scenarios (Table 3; see “Results”).
Values for maximum lifespan, maximum age of female/male reproduction and female/male age at first offspring were based on values from Wells and Scott (2018). Inbreeding depression was omitted; however, the default inbreeding depression value of 6.29 (O’Grady et al. 2006) was also tested (see below). A polygynous mating system, non-density-dependent reproduction and an equal sex ratio at birth were selected (c.f. Thompson et al. 2000) and it was assumed that all mature males were part of the breeding pool. Initial population size was set at 50, based on the average estimated abundance of the South Abaco population from 2001 to 2013 (Webber 2018). Carrying capacity was set at 70, based on the highest upper 95% confidence limit from 2001 to 2013 (Webber 2018). For each scenario simulated, a single population was stipulated, with 5000 iterations (see Currey et al. 2009a) over 150 years, with extinction defined as only one sex remaining. Reproduction and survival EV (environmental variation) correlation was set at the Vortex default of 0.5. Maximum number of broods per year and maximum number of progeny per brood were both set as 1.
Sensitivity testing
The Baseline (2005) survival and birth rates were varied by + 1% and + 1SDev to test the relative influence of fixed proportion and observed variation, respectively, in these parameters on the population projection (c.f. Manlik et al. 2016). The default inbreeding depression value of 6.29 was also tested. Testing involved calculating the difference between the predicted mean (stochastic) instantaneous population growth rate (stoch-r in Vortex, where λ = er; hereafter referred to as mean growth rate) of Baseline (2005) and the mean growth rate when a single parameter was altered, by + 1% or + 1SDev, whilst not altering any other values. To compare survival and birth rate parameter influence on population dynamics, a percentage change in the mean growth rate was calculated by dividing the difference in mean growth rate by the Baseline (2005) estimate of mean growth rate.
Population stabilisation
Given the results (see below), PVA simulations were run to establish the increases in survival (from Baseline (2005) values) needed to stabilise the South Abaco population. Keeping birth rate at the value in the Baseline (2005) scenario, survival rates were gradually increased by 1% across all age-classes simultaneously until stability was reached. This simplistic approach was chosen in the absence of information to warrant consideration of potentially more realistic scenarios in which targeted conservation measures might differentially affect the survival of different age-classes. The SDev values for mortality in the PVA were proportionally scaled to the respective survival estimates as survival rates were artificially increased. These same simulations were also run with birth rate increased to 33.3% (representing a likely minimum 3-year inter-birth interval). The SDev value for birth rate was also proportionally scaled when birth rate was artificially increased.
Results
Estimated birth rate
Only three models (within 2 AIC units of each other) received any support from the data; all other models had a delta-AIC of greater than 21.8 from the third most-supported model. These three models included years since previous birth and years since previous birth in quadratic form as fixed effects, and mother-ID and/or year as random effects. To account for model uncertainty, the estimated coefficients from these three models were model-averaged using AIC weights using the multi-model inference package MuMIn (Version 1.43.17; Bartoń 2020) implemented in R Version 4.1.1 (R Core Team 2021), to estimate the mean inter-birth interval and hence birth rate. Model-averaged inter-birth interval was 3.60 years (95% CI: 2.97–4.15) and annual birth rate was 0.278 (95% CI: 0.241–0.337).
Base estimates of survival
Calves and juveniles
In total, 56 calves with known year of birth were identified between 1996 and 2013. Preliminary analysis indicated that modelling calf survival as a single parameter for all calf ages received more support from the data than modelling age-specific calf survival, i.e. separate parameters for first-, second-, and third-year survival. Thus, calf survival was estimated only as a single parameter (non-age-specific). Within these models with non-age-specific calf survival, a model with linear time trend on survival interacting with age-class and constant recapture probability had much more support than any other model (Table 4). The results of this model indicated a decreasing trend in calf and juvenile survival (Fig. 2a, b). Estimated calf survival decreased from 0.970 (95% CI: 0.875–0.993) in 1997, to 0.876 (95% CI: 0.795–0.927) in 2005 (equivalent to approximately half-way through the time series) and 0.606 (95% CI: 0.368–0.802) in 2013, representing an approximate 36% decrease in apparent survival over the time series. Estimated juvenile survival declined less steeply from 0.820 (95% CI: 0.598–0.933) in 2000, to 0.800 (95% CI: 0.695–0.876) in 2005 and 0.767 (95% CI: 0.548–0.899) in 2013, representing an approximate 5% decrease in apparent survival over the time series. Recapture probability was estimated at 0.85 (95% CI: 0.79–0.90).

Estimates of apparent survival for (a) calves, (b) juveniles and (c) adults. For calves and juveniles, model-averaged estimates from the hurricane effect models are shown as well as estimates from the base model (see text and Tables 4 and 5). The hurricane covariates included in these models, either ‘time since last direct and/or major hurricane year’ or ‘cumulative count of direct and/or major hurricane years since 1992’, are shown on the top axis in panels (a) and (b)
Adults
The adult dataset included 84 individuals. Two models with linear time trend on survival interacting with transient-class had much more support than any other model and differed only in whether trap dependency was modelled as interacting with time or not (Table 5). The estimated coefficients from these two models were model-averaged using AICc weights to obtain base estimates of adult survival. Results indicated a declining trend in survival (Fig. 2c). Adult survival declined from 0.941 (95% CI: 0.876–0.973) in 1998, to 0.910 (95% CI: 0.878–0.935) in 2005 and 0.855 (95% CI: 0.718–0.932) in 2013, representing an approximate 9% decrease in apparent survival over the time series. Estimated recapture probability varied between 0.46 and 0.77.
Effect of hurricanes on survival
For calves and juveniles, six models had most support from the data within a delta-AICc of 3; other models had a delta-AICc of greater than 5 and thus little support (Table 4). Of these six models, two had more support than the others (delta-AICc > 2.5) and also slightly more support than the base survival model (delta-AICc = 0.8; Table 4). In both these models, the hurricane covariate was included as well as the linear time trend in survival in the base model, indicating that these hurricane covariates helped to explain additional variation in the data. The hurricane covariates in these two models both related to direct and/or major (DM) hurricane years, modelled in two different ways: ‘time since last direct and/or major hurricane year’ or ‘cumulative count of direct and/or major hurricane years since 1992’. Coefficients from these two most-supported DM hurricane effect models were model-averaged to produce a set of hurricane-impacted survival rates for calves and juveniles to explore how these estimates differed from the base survival estimates. Estimated calf survival dropped by approximately 3% in 1999 (Hurricanes Dennis and Floyd), 8% in 2004 (Hurricanes Frances and Jeanne), 19% in 2011 (Hurricane Irene) and 12% in 2012 (Hurricane Sandy) (Fig. 2a). Between these direct and/or major hurricane years, survival was estimated to continue to decline slowly, i.e. no recovery in estimated calf survival rates was predicted. Estimated juvenile survival dropped by approximately 11% in 2004, 13% in 2011 and 7% in 2012; however, in contrast to calf survival, juvenile survival was predicted to recover between direct and/or major hurricane years (Fig. 2b).
For adults, many hurricane effect models had some support from the data within a delta-AICc of 5 but only one had more support (delta-AICc = 0.3) than the base model (Table 5). In this model, the hurricane covariate, cumulative count of all hurricane years since 1992, replaced the linear time trend in survival in the most-supported base models (Table 5). That is, the hurricane covariate did not explain variation in the data additional to the trend in the base models. Of the most-supported hurricane effect models (delta-AICc < 2), only one included the hurricane covariate as well as the linear time trend. Survival estimates from these most-supported hurricane effect models were very similar to base estimates and are not shown in Fig. 2c.
Population viability analysis (PVA)
Model scenarios
The Baseline (2005) scenario estimated a mean growth rate of − 0.0662 for the South Abaco population, with a probability of extinction of 1 (i.e. all 5000 iterations were predicted to become extinct within 150 years), and a mean time to extinction of 41.9 years (Fig. 3). The High (2000) and Low (2010) scenarios estimated mean growth rates of − 0.0361 and − 0.1109, with a probability of extinction of 0.99 and 1, and a mean time to extinction of 73.0 years and 25.7 years, respectively (Fig. 3). Thus, even the most optimistic scenario predicted a strong population decline.

Population viability analysis for the South Abaco population using estimated survival values for calves, juveniles and adults from the base models (see text). Stoch-r mean (stochastic) instantaneous population growth rate, SD(r) standard deviation of r, PE probability of extinction, mean TE mean time to extinction (years)
Sensitivity testing
Mean growth rate was most sensitive to a + 1% increase in adult survival (Table 6), causing a + 11.3% increase in mean growth rate and resulting in the largest change in mean time to extinction. A + 1% increase in juvenile survival caused the mean growth rate to increase by + 3.3%. Changes in mean growth rate were smaller for + 1% increases in calf survival (+ 2.1%) and estimated birth rate (+ 1.8%). The inbreeding depression default value of 6.29 (O’Grady et al. 2006) had a small negative effect on mean growth rate (Table 6).
Observed variation sensitivity testing showed that a + 1SDev increase in juvenile survival had the largest proportional influence on mean growth rate, causing a + 20.4% increase (Table 6). A + 1SDev increase in adult survival had the second largest influence on mean growth rate, causing a proportional change of + 14.8%. Changes of + 1SDev to both calf survival and estimated birth rate had smaller effects on the mean growth rate of + 6.0% and + 3.6%, respectively (Table 6).
Population stabilisation
PVA simulations showed that survival estimates for calves, juveniles and adults would have to increase by + 6% from the Baseline (2005) values to stabilise the South Abaco population (Fig. 4). This would equate to survival rates of 0.936 for calves, 0.860 for juveniles and 0.970 for adults. When the estimated birth rate was increased to 33.3% (an inter-birth interval of 3 years), a + 5% increase in survival across all age-classes was not sufficient to stabilise population size, but a + 6% increase in survival would allow the population to grow (Fig. 4).

Results of population stabilisation simulations for the South Abaco population. Stoch-r mean (stochastic) instantaneous population growth rate, SD(r) standard deviation of r, yr year, IBI inter-birth interval. Increases in survival are from the Baseline (2005) values and were applied to calves, juveniles and adults simultaneously
Discussion
In this study, we used individual sighting histories to produce the first estimates of birth rate and age-class-specific apparent survival rates for the South Abaco population of bottlenose dolphins (Tursiops truncatus) on Little Bahama Bank (northern Bahamas), used these estimated life history parameters in a population viability analysis (PVA) to predict future population trends, and investigated the potential effect of hurricanes on calf, juvenile and adult survival. We found that there is a core adult population (with relatively high site fidelity) in South Abaco, based on regularly sighted marked adult individuals in the photographic-identification data and relatively high recapture probabilities estimated from the mark-recapture models. However, we also found evidence of transient adults, indicating that there are mature individuals that are infrequently seen in South Abaco. These findings are comparable with those reported for the East Abaco population (Fearnbach et al. 2012) and the Northern community (Rossbach and Herzing 1999; Rogers et al. 2004) on Little Bahama Bank. Population viability analysis simulations showed that the detected decline in survival of all age-classes would drive the South Abaco population (if isolated) to extinction in a relatively short time frame, even under the most optimistic scenario. Evidence that hurricanes had an effect on apparent survival was stronger for calves and juveniles than for adults.
Birth and survival rates
The estimated birth rate for the South Abaco population of 0.278 (95% CI: 0.241–0.337) is slightly higher than that reported for the east coast of Scotland (0.222, 95% CI = 0.218–0.253; Arso Civil et al. 2017) and considerably higher than that reported, using different methodology, for Sarasota, Florida (0.144, SD = 0.0244; Wells and Scott 1990). This suggests that birth rate is not currently a cause for concern for the South Abaco population. Adult apparent survival at the beginning of the time series (0.941; 95% CI: 0.876–0.973) is within the range reported for other bottlenose dolphin populations worldwide (e.g. Silva et al. 2009; Daura-Jorge et al. 2013; Arso Civil et al. 2019). It is also similar to the average survival rate reported for the East Abaco population between 1993 and 2009 (0.94; Fearnbach et al. 2012). Juvenile apparent survival in South Abaco was found to be markedly lower than for adults. Lower juvenile survival (where the juvenile definition is similar to the current study) has also been reported for bottlenose dolphin populations off the Azores (Silva et al. 2009) and in the Sado Estuary, Portugal (Gaspar 2003).
A decline in apparent survival was detected in calves, juveniles and adults, with a particularly strong effect in calves (a decline of approximately 36% over the study period), leading to low calf survival in later years in the time series. A strong decline in first-year calf survival in the population of bottlenose dolphins in Doubtful Sound, New Zealand occurred in a short period of time (Currey et al. 2009b). Estimates of survival may be subject to negative so-called terminal bias at the end of a time series, particularly in the final 3 years (e.g. Schleimer et al. 2019). In our study, the decline in estimated survival in the base models occurs across the whole study period, not only in the final three years, as found by Schleimer et al. (2019). The declines in the three final years estimated from the hurricane models for calves and juveniles were a result of including the hurricane covariates. Therefore, there is no evidence that the estimated declining trends in apparent survival are a result of terminal bias. We cannot distinguish between mortality and permanent (or long-term temporary) emigration in apparent survival and thus have no information to indicate whether the estimated declines are caused by one or the other or both (e.g. Baker et al. 2010). Several mother–calf pairs have been sighted within multiple local populations on Little Bahama Bank (Rogers et al. 2004; Parsons et al. 2006; the current study). It is, therefore, possible that declining calf survival rates could reflect mother–calf emigration from South Abaco instead of, or as well as, a decline in true survival. The estimated decline in juvenile survival was much less than for calves but the low estimates of juvenile survival overall may also be a result of emigration instead of, or as well as, true mortality.
Effect of hurricanes on survival rates
For adults, although inclusion of hurricane covariates did not improve our base survival models, results did show that replacing a simple time trend with a covariate representing cumulative hurricane activity had equivalent support from the data. Thus, it is possible that repeated hurricane activity may have caused or contributed to the estimated decline in survival over time. However, any such inference must be made very cautiously. Estimates of survival from the hurricane effect models were very similar to those from the base models providing little evidence that hurricanes had any effect on adult survival rates. It is also possible that other unmeasured factors that have increased over time could have caused or contributed to the estimated decline. Bassos-Hull et al. (2013) reported that bottlenose dolphins on the west coast of Florida were relatively unaffected at a population level by an individual major, direct hurricane event (Charley) in 2004. In contrast, a 3% decline in survival was reported in the East Abaco population in 1999 that was speculated to be due to the impact of Hurricanes Dennis and Floyd (Fearnbach et al. 2012). Repeated hurricane activity has also previously been speculated to be a potential contributing stressor to the long-term gradual decline in abundance in the East Abaco population between 1996 and 2009 (the end of the study time series; Fearnbach et al. 2012).
In contrast, there was stronger evidence from our analysis that direct and/or major (DM) hurricane events affected survival of calves and juveniles. Hurricane covariates describing time since the last DM hurricane year or the cumulative number of years with DM hurricanes were included in well-supported models as well as the time trend in survival, indicating that these hurricane covariates explained variation in the data additional to that trend. Calves have previously been reported to be more affected by extreme weather in other mammalian species, such as reindeer (Rangifer tarandus; Hansen et al. 2019). Miller et al. (2010) suggested that a potential increase in reproduction in a bottlenose dolphin population in the Mississippi Sound following a major hurricane (Katrina) in 2005 may have been partially influenced by an increase in calf mortality as a result of the hurricane. However, Bassos-Hull et al. (2013) argued that this could have been due to natural variation in birth rate. In 3 of the 4 years with the strongest decreases in estimated calf survival rate in our study (1999, 2004 and 2011), all five hurricanes occurred in the months of August and September, whereas in all other hurricane years, these events only occurred in the months of October and November (Table 1; NOAA 2021b). In East Abaco, neonates are mostly documented in the winter months (Fearnbach et al. 2011). Assuming the same calving seasonality in South Abaco, one possible contributing factor to the stronger decreases in calf survival in 1999, 2004 and 2011 could be that the first-year calves were relatively younger in these years and were thus more vulnerable when these hurricanes hit the South Abaco area.
A decrease in apparent survival rates may indicate an increase in emigration instead of, or as well as, an increase in mortality. The decline and recovery pattern in estimated juvenile survival (Fig. 2b) could indicate that disruptive DM hurricane events result in movement out of the South Abaco area, rather than, or as well as, true mortality. A large change in the Northern community social structure was seen following Hurricanes Frances and Jeanne in 2004, when 30% of the resident dolphins were replaced by immigrants (Elliser and Herzing 2011). However, the extent to which juvenile animals were involved is unclear, as the study mostly did not differentiate between juveniles and adults.
Predicted population decline
Assuming the bottlenose dolphins in South Abaco form an isolated population and that survival rates do not improve, our PVA simulations predict a strong decline towards extinction within decades. To stabilise the population, survival rates of calves, juveniles and adults would need to increase by 6% from our suggested Baseline (2005), which represents the approximate average over the study period. These survival rates that would allow a viable population fall within the upper end of the range reported for other populations of bottlenose dolphins (e.g. Gaspar 2003; Silva et al. 2009; Arso Civil et al. 2019). Currey et al. (2011) identified reduced survival in calves (animals less than 3 years old) as the likely cause of the decline in the Doubtful Sound bottlenose dolphin population. In our study in South Abaco, the decline in calf survival is greater than the declines in juvenile and adult survival, but even our most optimistic PVA scenario projected a steep population decline when calf survival was high at approximately 95%. Given that estimated birth rate in South Abaco is similar to or higher than in other bottlenose dolphin populations, it is likely that low adult and/or low juvenile survival rates are driving the predicted population decline. This tentative conclusion is also supported by the results of the sensitivity analysis, which show that population growth rate is most influenced by variation in adult and juvenile survival. However, evidence of movements of dolphins between populations on Little Bahama Bank (Rogers et al. 2004; Parsons et al. 2006; the current study) indicates that the South Abaco population is unlikely to be isolated. The declining survival rates estimated here may thus reflect a combination of both true mortality and permanent (or long-term temporary) emigration, the relative contribution of which cannot be estimated without additional data and analysis.
In some situations, there may be an identifiable cause of a decline in a local small cetacean population, which may be able to be directly targeted for management action (e.g. Currey et al. 2009b; Slooten 2013). However, where populations may be impacted by multiple pressures, it is more difficult to identify specific conservation measures to arrest a decline (e.g. Tezanos-Pinto et al. 2015). Despite indications that hurricane activity may have affected bottlenose dolphin survival in South Abaco, especially in calves, the extent to which the estimated long-term declines in survival in all age-classes are caused by repeated hurricane activity in this area is unknown. There are other pressures that might be affecting this population. Suspected underlying impacts of overfishing (e.g. Bearzi et al. 2008) and disturbance from continued presence of small boats (e.g. Bejder et al. 2006; Pirotta et al. 2015) could potentially be influencing survival rates (BMMRO unpublished data). Shark predation risk to younger dolphins (e.g. Mann and Barnett 1999) may also be a contributing factor, which could increase following hurricane events, as seen in the East Abaco population (Fearnbach et al. 2011). If hurricanes are a contributing cause of a decline in the South Abaco population, and a continued high frequency and/or potential increase in intensity of hurricane events occurs due to climate change (e.g. NOAA GFDL 2021), this would be impossible to mitigate against.
Little Bahama Bank bottlenose dolphins as a metapopulation
Our results provide some support for the hypothesis that South Abaco bottlenose dolphins could form part of a larger metapopulation on Little Bahama Bank. The generally high site fidelity but also evidence of infrequently sighted individuals would be expected if the local South Abaco population was part of a metapopulation on the Bank. A similar pattern of relatively high site fidelity, but with some movement of individuals (and limited gene flow) has also been observed in other populations (East Abaco, Northern community) on Little Bahama Bank (Rossbach and Herzing 1999; Rogers et al. 2004; Parsons et al. 2006; Fearnbach et al. 2012).
If the small local population in South Abaco is part of a metapopulation, its decline as predicted by the population viability analysis, whether a result of mortality and/or emigration, would form a natural part of the source–sink dynamics (Dias 1996). In that scenario, a small South Abaco sink may have little impact on a larger metapopulation on Little Bahama Bank. However, the loss of a local population of bottlenose dolphins could result in reduced genetic and behavioural variation (e.g. Reeves 2018) and the regional ecosystem in South Abaco might also be disturbed (Heithaus et al. 2008), so this would not be without impact. If, instead, Little Bahama Bank does not act as a metapopulation, or if there were other local sink populations within it (as potentially indicated by the abundance decline in the East Abaco population; Fearnbach et al. 2012), this would represent a greater conservation concern.
A focus of future work on the South Abaco population should be on assessing trends in local abundance to determine if the estimated declines in apparent survival rates and the predicted decline in population size are reflected in an observed decline in abundance. In a wider context, future work should aim to update the status of all local populations on Little Bahama Bank and to co-ordinate research efforts to establish how the local populations interact as well as to study their dynamics jointly. This could help to provide wider context to potential emigration (permanent or temporary) from South Abaco. Continued long-term photographic-identification studies will be essential in elucidating the existence of a metapopulation of bottlenose dolphins on Little Bahama Bank, as well as assessing their conservation status with potentially increasing environmental changes and anthropogenic pressures.
References
Agrelo M, Daura-Jorge FG, Rowntree VJ, Sironi M, Hammond PS, Ingram SN, Marón CF, Vilches FO, Seger J, Payne R, Simões-Lopes PC (2021) Ocean warming threatens southern right whale population recovery. Sci Adv 7:2823. https://doi.org/10.1126/sciadv.abh2823
Arso Civil M, Cheney B, Quick NJ, Thompson PM, Hammond PS (2017) A new approach to estimate fecundity rate from inter-birth intervals. Ecosphere 8(4):e01796. https://doi.org/10.1002/ecs2.1796
Arso Civil M, Cheney B, Quick NJ, Islas-Villanueva V, Graves JA, Janik VM, Thompson PM, Hammond PS (2019) Variations in age- and sex-specific survival rates help explain population trend in a discrete marine mammal population. Ecol Evol 9(1):533–544. https://doi.org/10.1002/ece3.4772
Baker JD, Westgate A, Eguchi T (2010) Vital rates and population dynamics. In: Boyd IL, Bowen WD, Iverson SJ (eds) (2010) Marine mammal ecology and conservation: a handbook of techniques. Oxford University Press, pp 119–143
Bartoń K (2020) MuMIn: multi-model inference. R package, version 1.43.17. https://CRAN.R-project.org/package=MuMIn. Accessed 12 Nov 2021
Bassos-Hull K, Perrtree RM, Shepard CC, Schilling S, Barleycorn AA, Allen JB, Balmer BC, Pine WE, Wells RS (2013) Long-term site fidelity and seasonal abundance estimates of common bottlenose dolphins (Tursiops truncatus) along the southwest coast of Florida and responses to natural perturbations. J Cetac Res Manage 13(1):19–30
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67(1):1–48. https://doi.org/10.18637/jss.v067.i01
Bearzi G, Agazzi S, Gonzalvo J, Costa M, Bonizzoni S, Politi E, Piroddi C, Reeves RR (2008) Overfishing and the disappearance of short-beaked common dolphins from western Greece. Endanger Species Res 5:1–12. https://doi.org/10.3354/esr00103
Bejder L, Samuels A, Whitehead H, Gales N, Mann J, Connor R, Heithaus M, Watson-Capps J, Flaherty C, Krützen M (2006) Decline in relative abundance of bottlenose dolphins exposed to long-term disturbance. Conserv Biol 20(6):1791–1798. https://doi.org/10.1111/j.1523-1739.2006.00540.x
Bezamat C, Hammond PS, Castilho PV, Simões-Lopes PC, Daura-Jorge FG (2021) Dolphin population specialized in foraging with artisanal fishers requires zero-bycatch management to persist. Aquat Conserv Mar Freshwat Ecosyst 31(11):3133–3145. https://doi.org/10.1002/aqc.3694
Brook BW, O’Grady JJ, Chapman AP, Burgman MA, Akçakaya HR, Frankham R (2000) Predictive accuracy of population viability analysis in conservation biology. Nature 404:385–387. https://doi.org/10.1038/35006050
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Currey RJC, Dawson SM, Slooten E (2009a) An approach for regional threat assessment under IUCN Red List criteria that is robust to uncertainty: the Fiordland bottlenose dolphins are critically endangered. Biol Cons 142(8):1570–1579. https://doi.org/10.1016/j.biocon.2009.02.036
Currey RJC, Dawson SM, Slooten E, Schneider K, Lusseau D, Boisseau OJ, Haase P, Williams JA (2009b) Survival rates for a declining population of bottlenose dolphins in Doubtful Sound, New Zealand: an information theoretic approach to assessing the role of human impacts. Aquat Conserv Mar Freshwat Ecosyst 19(6):658–670. https://doi.org/10.1002/aqc.1015
Currey RJC, Dawson SM, Schneider K, Lusseau D, Boisseau OJ, Haase PA, Slooten E (2011) Inferring causal factors for a declining population of bottlenose dolphins via temporal symmetry capture-recapture modeling. Mar Mamm Sci 27(3):554–566. https://doi.org/10.1111/j.1748-7692.2010.00417.x
Daura-Jorge FG, Ingram SN, Simões-Lopes PC (2013) Seasonal abundance and adult survival of bottlenose dolphins (Tursiops truncatus) in a community that cooperatively forages with fishermen in southern Brazil. Mar Mamm Sci 29(2):293–311. https://doi.org/10.1111/j.1748-7692.2012.00571.x
Dias PC (1996) Sources and sinks in population biology. Trends Ecol Evol 11(8):326–330. https://doi.org/10.1016/0169-5347(96)10037-9
Durban JW (2002) Bayesian methods for marine mammal population assessment. PhD Thesis, University of Aberdeen, UK
Durban JW, Parsons KM, Claridge DE, Balcomb KC (2000) Quantifying dolphin occupancy patterns. Mar Mamm Sci 16(4):825–828. https://doi.org/10.1111/j.1748-7692.2000.tb00975.x
Elliser CR, Herzing DL (2011) Replacement dolphins? Social restructuring of a resident pod of Atlantic bottlenose dolphins, Tursiops truncatus, after two major hurricanes. Mar Mamm Sci 27(1):39–59. https://doi.org/10.1111/j.1748-7692.2010.00403.x
Fearnbach H, Durban J, Parsons K, Claridge D (2011) Seasonality of calving and predation risk in bottlenose dolphins on Little Bahama Bank. Mar Mamm Sci 28(2):402–411. https://doi.org/10.1111/j.1748-7692.2011.00481.x
Fearnbach H, Durban J, Parsons K, Claridge D (2012) Photographic mark-recapture analysis of local dynamics within an open population of dolphins. Ecol Appl 22(5):1689–1700. https://doi.org/10.1890/12-0021.1
Gardner TA, Côté IM, Gill JA, Grant A, Watkinson AR (2005) Hurricanes and Caribbean coral reefs: impacts, recovery patterns, and role in long-term decline. Ecology 86(1):174–184. https://doi.org/10.1890/04-0141
Gaspar R (2003) Status of the resident bottlenose dolphin population in the Sado Estuary: past, present and future. PhD Thesis, University of St Andrews, UK
GEBCO Compilation Group (2020) GEBCO 2020 Grid. https://www.gebco.net/data_and_products/gridded_bathymetry_data/gebco_2020/
Gimenez O, Lebreton J-D, Choquet R, Pradel R (2018) R2ucare: an R package to perform goodness-of-fit tests for capture-recapture models. Methods Ecol Evol 9(7):1749–1754. https://doi.org/10.1111/2041-210X.13014
Hansen BB, Gamelon M, Albon SD, Lee AM, Stien A, Irvine RJ, Sæther B-E, Loe LE, Ropstad E, Veiberg V, Grøtan V (2019) More frequent extreme climate events stabilize reindeer population dynamics. Nat Commun 10:1616. https://doi.org/10.1038/s41467-019-09332-5
Heithaus MR, Frid A, Wirsing AJ, Worm B (2008) Predicting ecological consequences of marine top predator declines. Trends Ecol Evol 23(4):202–210. https://doi.org/10.1016/j.tree.2008.01.003
Jourdain E, Goh T, Kuningas S, Similä T, Vongraven D, Karoliussen R, Bisther A, Hammond PS (2021) Dynamics of a killer whale (Orcinus orca) population in response to a period of rapid ecosystem change in the eastern North Atlantic. Ecol Evol 11(23):17289–17306. https://doi.org/10.1002/ece3.8364
Knutson TR, Chung MV, Vecchi G, Sun J, Hsieh T-L, Smith AJP (2021) Climate change is probably increasing the intensity of tropical cyclones. Zenodo. https://doi.org/10.5281/zenodo.4570334
Laake JL (2013) RMark: an R interface for analysis of capture-recapture data with MARK. AFSC processed Rep. 2013–01. Alaska Fisheries Science Center, NOAA, National Marine Fisheries Service, Seattle
Lacy RC, Pollak JP (2020) Vortex: a stochastic simulation of the extinction process. Version 10.3.8. Chicago Zoological Society, Brookfield, Illinois, USA. https://scti.tools/vortex/. Accessed 12 Nov 2021
Langtimm CA, Beck CA (2003) Lower survival probabilities for adult Florida manatees in years with intense coastal storms. Ecol Appl 13(1):257–268. https://doi.org/10.1890/1051-0761(2003)013[0257:LSPFAF]2.0.CO;2
Lebreton J-D, Burnham KP, Clobert J, Anderson DR (1992) Modeling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecol Monogr 62(1):67–118. https://doi.org/10.2307/2937171
Manlik O, McDonald JA, Mann J, Raudino HC, Bejder L, Krützen M, Connor RC, Heithaus MR, Lacy RC, Sherwin WB (2016) The relative importance of reproduction and survival for the conservation of two dolphin populations. Ecol Evol 6(11):3496–3512. https://doi.org/10.1002/ece3.2130
Mann J, Barnett H (1999) Lethal tiger shark (Galeocerdo cuvier) attack on bottlenose dolphin (Tursiops sp.) calf: defense and reactions by the mother. Mar Mamm Sci 15(2):568–575. https://doi.org/10.1111/j.1748-7692.1999.tb00823.x
Mignucci-Giannoni AA, Toyos-González GM, Pérez-Padilla J, Rodríguez-López MA, Overing J (2000) Mass stranding of pygmy killer whales (Feresa attenuata) in the British Virgin Islands. J Mar Biol Assoc UK 80(2):383–384. https://doi.org/10.1017/S0025315499002076
Miller LJ, Mackey AD, Hoffland T, Solangi M, Kuczaj SA II (2010) Potential effects of a major hurricane on Atlantic bottlenose dolphin (Tursiops truncatus) reproduction in the Mississippi Sound. Mar Mamm Sci 26(3):707–715. https://doi.org/10.1111/j.1748-7692.2010.00371.x
National Academy of Sciences (2020) Climate change: evidence and causes: update 2020. The National Academies Press, Washington. https://doi.org/10.17226/25733
National Oceanic and Atmospheric Administration (2021a) Saffir-Simpson Hurricane Wind Scale. National Hurricane Center. https://www.nhc.noaa.gov/aboutsshws.php. Accessed 12 Nov 2021
National Oceanic and Atmospheric Administration (2021b) Past track seasonal maps (Atlantic Basin)/Tropical cyclone reports: Atlantic, Caribbean, and the Gulf of Mexico. National Hurricane Center. https://www.nhc.noaa.gov/data/. Accessed 12 Nov 2021
NOAA Geophysical Fluid Dynamics Laboratory (GFDL) (2021) Global warming and hurricanes: an overview of current research results. NOAA GFDL. https://www.gfdl.noaa.gov/global-warming-and-hurricanes/. Accessed 12 Nov 2021
O’Grady JJ, Brook BW, Reed DH, Ballou JD, Tonkyn DW, Frankham R (2006) Realistic levels of inbreeding depression strongly affect extinction risk in wild populations. Biol Cons 133(1):42–51. https://doi.org/10.1016/j.biocon.2006.05.016
Parsons KM, Durban JW, Claridge DE, Herzing DL, Balcomb KC, Noble LR (2006) Population genetic structure of coastal bottlenose dolphins (Tursiops truncatus) in the northern Bahamas. Mar Mamm Sci 22(2):276–298. https://doi.org/10.1111/j.1748-7692.2006.00019.x
Pirotta E, Merchant ND, Thompson PM, Barton TR, Lusseau D (2015) Quantifying the effect of boat disturbance on bottlenose dolphin foraging activity. Biol Cons 181:82–89. https://doi.org/10.1016/j.biocon.2014.11.003
Pradel R, Hines JE, Lebreton J-D, Nichols JD (1997) Capture-recapture survival models taking account of transients. Biometrics 53(1):60–72. https://doi.org/10.2307/2533097
Pradel R, Gimenez O, Lebreton J-D (2005) Principles and interest of GOF tests for multistate capture–recapture models. Anim Biodivers Conserv 28(2):189–204
Preen A, Marsh H (1995) Response of dugongs to large-scale loss of seagrass from Hervey Bay, Queensland, Australia. Wildl Res 22(4):507–519. https://doi.org/10.1071/WR9950507
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Reeves RR (2018) Conservation. In: Würsig B, Thewissen JGM, Kovacs KM (eds) Encyclopedia of marine mammals, 3rd edn. Academic Press, pp 215–229
Rogers CA, Brunnick BJ, Herzing DL, Baldwin JD (2004) The social structure of bottlenose dolphins, Tursiops truncatus, in the Bahamas. Mar Mamm Sci 20(4):688–708. https://doi.org/10.1111/j.1748-7692.2004.tb01188.x
Rossbach KA, Herzing DL (1999) Inshore and offshore bottlenose dolphin (Tursiops truncatus) communities distinguished by association patterns near Grand Bahama Island, Bahamas. Can J Zool 77(4):581–592. https://doi.org/10.1139/z99-018
Schleimer A, Ramp C, Delarue J, Carpentier A, Bérubé M, Palsbøll PJ, Sears R, Hammond PS (2019) Decline in abundance and apparent survival rates of fin whales (Balaenoptera physalus) in the northern Gulf of St. Lawrence. Ecol Evol 9(7):4231–4244. https://doi.org/10.1002/ece3.5055
Silva MA, Magalhães S, Prieto R, Serrão Santos R, Hammond PS (2009) Estimating survival and abundance in a bottlenose dolphin population taking into account transience and temporary emigration. Mar Ecol Prog Ser 392:263–276. https://doi.org/10.3354/meps08233
Slooten E (2013) Effectiveness of area-based management in reducing bycatch of the New Zealand dolphin. Endanger Species Res 20:121–130. https://doi.org/10.3354/esr00483
Smith CE, Hurley BJ, Toms CN, Mackey AD, Solangi M, Kuczaj SA II (2013) Hurricane impacts on the foraging patterns of bottlenose dolphins Tursiops truncatus in Mississippi Sound. Mar Ecol Prog Ser 487:231–244. https://doi.org/10.3354/meps10372
Tanner EVJ, Kapos V, Healey JR (1991) Hurricane effects on forest ecosystems in the Caribbean. Biotropica 23(4):513–521. https://doi.org/10.2307/2388274
Taylor BL, Rojas-Bracho L, Moore J, Jaramillo-Legorreta A, Ver Hoef JM, Cardenas-Hinojosa G, Nieto-Garcia E, Barlow J, Gerrodette T, Tregenza N, Thomas L, Hammond PS (2017) Extinction is imminent for Mexico’s endemic porpoise unless fishery bycatch is eliminated. Conserv Lett 10(5):588–595. https://doi.org/10.1111/conl.12331
Tezanos-Pinto G, Constantine R, Mourão F, Berghan J, Baker CS (2015) High calf mortality in bottlenose dolphins in the Bay of Islands, New Zealand—a local unit in decline. Mar Mamm Sci 31(2):540–559. https://doi.org/10.1111/mms.12174
Thompson PM, Wilson B, Grellier K, Hammond PS (2000) Combining power analysis and population viability analysis to compare traditional and precautionary approaches to conservation of coastal cetaceans. Conserv Biol 14(5):1253–1263. https://doi.org/10.1046/j.1523-1739.2000.00099-410.x
Wauer RH, Wunderle JM Jr (1992) The effect of hurricane Hugo on bird populations on St. Croix, U.S. Virgin Islands. Wilson Bull 104(4):656–673
Webber T (2018) Abundance and survival estimates of common bottlenose dolphins (Tursiops truncatus) in South Abaco, The Bahamas. Unpublished BSc Thesis. University of Plymouth, UK.
Wells RS, Scott MD (1990) Estimating bottlenose dolphin population parameters from individual identification and capture-release techniques. Rep Int Whal Commn (Special Issue 12):407–415
Wells RS, Scott MD (2018) Bottlenose dolphin, Tursiops truncatus, common bottlenose dolphin. In: Würsig B, Thewissen JGM, Kovacs KM (eds) Encyclopedia of marine mammals, 3rd edn. Academic Press, pp 118–125
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46:S120–S139. https://doi.org/10.1080/00063659909477239
White TH Jr, Collazo JA, Vilella FJ, Guerrero SA (2005) Effects of Hurricane Georges on habitat use by captive-reared Hispaniolan parrots (Amazona ventralis) released in the Dominican Republic. Ornitologia Neotropical 16(3):405–417
Willoughby HE, Rappaport EN, Marks FD (2007) Hurricane forecasting: the state of the art. Nat Hazard Rev 8(3):45–49. https://doi.org/10.1061/(ASCE)1527-6988(2007)8:3(45)
Acknowledgements
This study was made possible with financial support from Earthwatch Institute, Atlantis Blue Project Foundation, with permission to conduct research granted by the Bahamas Department of Marine Resources. Many Earthwatch volunteers, volunteer staff members, and student interns assisted in data collection and processing. The authors would like to specifically thank Ken Balcomb, John Durban, Kim Parsons, Leigh Hickmott and Felice Major for their contribution to this work.
Author information
Authors and Affiliations
Contributions
JC and PSH developed the principal ideas and designed the study methodology; DC and CD sourced funding for photo-identification surveys, collected and processed photo-identification data; MAC provided the birth rate estimation methodology and code; JC analysed the data; JC, MAC and PSH led the writing of the manuscript; all the authors critically reviewed manuscript drafts and approved the final manuscript for publication.
Corresponding authors
Ethics declarations
Conflict of interest
On behalf of all the authors, the corresponding authors state that there is no conflict of interest.
Additional information
Handling Editors: Stephen C.Y. Chan and Leszek Karczmarski.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue on “Individual Identification and Photographic Techniques in Mammalian Ecological and Behavioural Research – Part 2: Field Studies and Applications” — Editors: Leszek Karczmarski, Stephen C.Y. Chan, Scott Y.S. Chui and Elissa Z. Cameron.
Supplementary Information
Below is the link to the electronic supplementary material.
Appendix
Appendix
See Fig. A1.

Bottlenose dolphin (Tursiops truncatus) mother-calf pair from the local South Abaco population on Little Bahama Bank (northern Bahamas). Photo credit: Bahamas Marine Mammal Research Organisation
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Coxon, J., Arso Civil, M., Claridge, D. et al. Investigating local population dynamics of bottlenose dolphins in the northern Bahamas and the impact of hurricanes on survival. Mamm Biol 102, 1133–1148 (2022). https://doi.org/10.1007/s42991-021-00208-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-021-00208-0