
Abstract
Purpose
Recovery within and between rounds is crucial to combat sports performance. We sought to determine whether sprint interval training (SIT) improves recovery dynamics and aerobic performance.
Methods
Eleven male kickboxing athletes (26 ± 5 years; body mass index 25 ± 3 kg/m2) were recruited. Participants were tested three times for VO2peak/time to exhaustion and critical power; baseline, 3 weeks control, 3 weeks of SIT (8 × 10 s lower body sprints followed by a maximum of 10 min recovery before completing 8 × 10 s upper body sprints). During SIT session 1 and 9 continuous gas analysis was performed.
Results
There was a significant reduction in recovery time between lower and upper body sprints with training (session 1: 441 ± 150 s; session 9: 268 ± 10 s; P < 0.01; d = 2.77) and change in oxygen off-kinetics amplitude (session1: 3.0 ± 0.7 L/min, session 9: 3.6 ± 1.0 L/min; P < 0.05; d = − 1.77), VO2 end (session 1: 0.59 ± 0.19 L/min, session 9: 0.81 ± 0.21 L/min; P < 0.05, d = − 0.90), time constant (session 1: 81 ± 21 s; session 9: 60 ± 11 s; P < 0.05; d = 1.03). Following training there was a significant improvement in critical power (P < 0.05; η2p = 0.72) time to exhaustion (P < 0.05; η2p = 0.30) but not VO2peak (P > 0.05).
Conclusion
SIT improves recovery time associated and aerobic performance associated with improved oxygen off-kinetics. Therefore, training needs to focus on improving oxygen off-kinetics to enhance combat performance.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Kickboxing is characterised as dynamic, intermittent, high intensity activities that require complex skills and tactical awareness for success [28, 33]. The duration of bouts varies depending on the competition but generally it is between 3 and 12 rounds of combat with a 1–2 min recovery period [35]. The work to rest ratio within rounds is similar between the winning and losing athletes [34], however there is a greater number of attacking movements from winning athletes in kickboxing [28]. Across a fight the work to rest ratio has been shown to increase as athletes tire, with an increasing cardiovascular load and perception of effort [34], reflecting the need to for a high capacity for recovery within and between rounds.
Recovery from high intensity intermittent exercise within and between a round is suggested to be biphasic [38] and speed of recovery is linked to a faster rate of phosphocreatine resynthesis, a greater acid/base buffering capacity and a greater mitochondrial enzyme capacity for oxidative metabolism [41]. Therefore, endurance capacity has been seen as a major determinant of recovery kinetics post exercise. When looking at repeated high intensity exercise, endurance trained athletes have been shown to consume a greater volume of oxygen over the course of the exercise for the same power output [38]. This is then suggested to result in a lower reliance on anaerobic metabolism. Following submaximal exercise, the greater oxygen consumption of athletes with a high endurance capacity has been shown to result in a faster excessive post exercise oxygen consumption (EPOC) [38]. In contrast, following a single 30 s Wingate there was no relationship between EPOC and endurance capacity as assessed by determination of maximal volume of oxygen consumption (VO2max) [9]. This suggests the relationship between endurance and recovery may not be as simple as an athletes VO2max. For example, there is a strong positive relationship between faster oxygen kinetics and endurance capacity of athletes [32].
Given the association of endurance with recovery, there is a need to find training modalities that enhance both in combat sports. Typically, kickboxing athletes have been shown to have a VO2max of 53.9 mL/min/kg [35] but can be as low as 48.5 mL/min/kg [35], which is similar to other combat sports [5]. This may suggest that combat training does not target large endurance adaption which may limit the athlete’s ability to recover between rounds. Cycle based sprint interval training (SIT) can be defined as supramaximal all-out efforts for a short time period (< 30 s) interspersed with period of recovery [23]. Typical adaptations to SIT are increased mitochondrial oxidative enzyme content and activity, increased muscle capillarisation and improved lactate metabolism [23], leading to decreased fatiguability of the skeletal muscle [23]. In contrast, improvements in VO2max following 6 weeks of SIT has also been associated with central adaptations with increased cardiac output and stroke volume reported [24]. This improvement may be related to improved vascular distensibility following SIT [30]. In contrast, Raleigh et al. [31] found no change in cardiac output following 4 weeks of training but showed a greater extraction across the muscle. Within combat sports, SIT interventions have been shown to increase both aerobic and anaerobic performance [1, 27] suggesting it is an effective training modality to improve performance. However, to date no studies have looked at the effectiveness of SIT for improving recovery dynamics. Given the changes reported with SIT then it could be that both central and peripheral adaptations will drive recovery.
Given that endurance capacity is strongly associated with recovery and SIT has been shown to elicit both central and peripheral improvements associated with endurance, then the aim of the current study was to determine whether SIT reduces self-selected recovery time between high intensity bouts of exercise. It is hypothesised that improvements in oxygen kinetics and muscular performance following SIT will increase the speed of recovery.
Methods
Participants
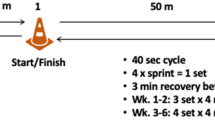
Eleven male kickboxers (26 ± 5 years; height 177 ± 7 cm; body mass 81 ± 11 kg; body mass index 25 ± 3 kg/m2) were recruited for the study. All participants had a minimum of 2 years experience in kickboxing and had no musculoskeletal injuries over the previous 6 months. Participants were informed of the study verbally and in writing before providing written consent. A completed Physical Activity Readiness Questionnaire screened for any health issues. The study was fully approved by the Abertay University Ethics Committee and carried out in line with the Declaration of Helsinki, 2013. Overview of the study design is shown in Fig. 1.

Overview of the study design
Testing
Participants were asked to refrain from alcohol and strenuous exercise for 24 h prior to testing and to consume no food or fluids for 4 h beforehand. All testing was carried out at the same time of day and in the same order across the testing periods.
Incremental Test
Seat height was adjusted on the cycle ergometer (Monark 894E, Monark, Varberg, Sweden) to ensure full extension of the leg on the pedal downstroke. Participants were then connected to the gas analyser (Metalyzer 3B, Cortex, Leipzig, Germany) and asked to cycle at 60 revolutions per minute (r/min) throughout the duration of the test. Testing started with 1 kg of resistance on the ergometer and was increased by 0.5 kg/min. When the participant could no longer maintain 60 rpm the test was stopped, and the time taken and recorded as the participants time to exhaustion. VO2peak was taken as the highest 30 s average across the test.
3-min Critical Power
On a separate day (a minimum of 48 h post incremental test), participants carried out a 3-min critical power test [39]. Seat height was set at the same height as the incremental test, participants were then asked to cycle at 60 r/min (Monark 894E, Monark, Varberg, Sweden) against 1 kg resistance for 4 min. Participants then performed a 3 min ‘all-out’ critical power test against 4.5% body mass. The test began as soon as the participants reached 110 r/min with strong verbal encouragement throughout, but no time indication given. Power output (Watts) was recorded using Monark software (Monark Anaerobic Test Software Version 2.24.2, Monark Exercise AB), with the average final 30 s of the test taken as CP. The power time curve was then analysed using a first order exponential decay;
where YO is the asymptotic value for power; TD is the time constant and A is the difference between Wpeak and Wo. Critical power was taken as the asymptotic value for power (YO).
Control Period
Given that the homeostatic stress to an exercise session is different between people [25], then this would impact on recovery duration selected and result in greater variability in the outcome measure if a separate control group were utilised. Therefore, a self-controlled design was used where participants were asked to continue with their own training for 3 weeks after baseline testing and retested before undertaking the training intervention.
Training Intervention
Training sessions were carried out three times per week over 3 weeks, with 48 h between sessions. Session warm-up consisted of a 4-min lower body cycle at 60 r/min against 1 kg resistance. Following warm-up, participants carried out 8 × 10 s sprints against 8% body mass with 30 s recovery between each sprint. Resistance dropped once participants reached 110 r/min. After lower body sprints, participants had a seated recovery period which could last no longer than 10 min, with the participant indicating when they felt that they were sufficiently recovered. Participants then moved to the upper body arm ergometer (Monark 891EW, Monark, Varberg, Sweden), kneeling in front with buttocks touching heels. Participants then carried out 8 × 10 s sprints against 4.5% body mass with 30 s recovery between each sprint. Resistance was constantly applied.
Session 1 and 9
During the first and last training session, participants had a fingerprick blood sample prior to starting the training session to measure baseline blood lactate concentration (Lactate pro, Arkay Inc., Kyoto, Japan). Blood samples were then collected immediately after lower body sprints and immediately before upper body sprints. During training session 1 and 9 participants were connected to a calibrated breath by breath oxygen analyser (Metalyzer®3B gas analyser, Cortex, Leipzig, Germany) and VO2 and VCO2 recorded throughout. Participants were told they has a maximum of 10 min recovery time and should inform the researcher once they felt that they were sufficiently recovered. Recovery time between lower and upper body sprints was then recorded and VO2 and VCO2 data exported as 1 s averages. Oxygen off-kinetics was fitted with a mono-exponential function [6], using QtiPlot Software (Version 1.1, Bucuresti, Romania).
where, A is the asymptotic value for the exponential term, τ is the time constant for decay, and TD is the time delay and XEE is the end VO2 for recovery. Due to data corruption, VO2 and VCO2 data was only available for 9 participants for both session 1 and 9. The goodness of fit for the exponential function for session 1 was R2 0.92 and session 9 was R2 0.91.
Data Analysis
All data are presented as mean ± standard deviation. All data analysis were completed using Jamovi (version 1.1.9). Shapiro–Wilk test and visual analysis were conducted to ensure normal distribution and a repeated analysis of variance (ANOVA) was used to determine the difference between each testing session, with LSD post hoc analysis. Data for training session 1 and 9 were analysed using a paired samples t-test. Significance was accepted at P < 0.05. Partial eta squared (η2p) were calculated for effect size for ANOVA where 0.02 is a small effect size, 0.13 is a medium effect size and 0.26 is a large effect size [3]. Cohens d were calculated for effect size for t-test where 0.2–0.5 small, 0.6–1.1moderate, and 1.2–1.9 as a large effect [12].
Multivariate linear analysis: Multivariate linear regression was carried out for recovery time, TTE and CP30. Prior to being included in the model covariates were checked for independence using Pearson’s correlation. There was a significant correlation between VO2 and VCO2 area under curve (r = 0.92; P < 0.001), last 60 s VO2 and VCO2 (r = 0.92; P < 0.001), therefore all multivariate regression analysis were only run on VO2 data. Covariates for the recovery time model were the last 60 s VO2, recovery between sets VO2 area under the curve, amplitude, time constant and change in blood lactate after exercise. There was a significant correlation between CP30 and YO (r = 0.97; P < 0.001), therefore model was only run on CP30. Covariates for TTE and CP30 were last 60 s VO2, total work done in the lower body sprints, Amplitude, and time constant of oxygen off kinetics. All models were checked for collinearity and if the variance inflation factor (VIF) value for each covariate was less than 3 with a tolerance greater than 0.2 then they were assumed to be independent factors [17].
Results
Training
There was no significant difference between training sessions for peak or average power for any sprint (data not shown). Following 9 SIT sessions, lower body VO2 area under the curve (AUC) during sprint and recovery was significantly different (sprint AUC: P < 0.001; η2p = 0.46; recovery AUC: P < 0.001; η2p = 0.25; Table 1). For lower body VCO2 AUC there was a significant effect of training session on for sprints but not recovery (sprint AUC: P < 0.001; η2p = 0.24; recovery AUC: P > 0.05; η2p = 0.01; Table 2). Area under the curve for VO2 for overall recovery from lower body sprints was significantly lower in session 9 compared to session 1 (P < 0.05; d = 0.95; Table 1). Likewise, AUC for VCO2 for overall recovery from lower body sprints was significantly lower in session 9 compared to session 1 (P < 0.05; d = 1.34; Table 2). There was a significant effect of training session on upper body VO2 area under the curve (sprint AUC: P < 0.001; η2p = 0.57; recovery AUC: P < 0.001; η2p = 0.20; Table 1) and upper body VCO2 area under the curve for sprints and recovery (sprint AUC: P < 0.001; η2p = 0.31; recovery AUC: P < 0.05; η2p = 0.11; Table 2).
Recovery time between sprint sets (i.e. from lower body to upper body sprints) was significantly lower in session 9 compared to session 1 (session 1: 441 ± 150 s; session 9: 268 ± 10 s; P < 0.01; d = 2.77; Fig. 2C). During the recovery, VO2 and VCO2 plateaued prior to recovery and the average of the last 60 s for VO2 and VCO2 was significantly higher in session 9 compared to session 1 (VO2 session 1: 771 ± 254 mL/min; VO2 session 9: 1007 ± 292 mL/min; P < 0.01; d = − 1.36; VCO2 session 1: 859 ± 316 mL/min; VCO2 session 9: 1190 ± 398 mL/min; P < 0.01; d = − 2.76; Fig. 1A, B). This represents a significantly greater percentage of incremental VO2peak in session 9 compared to session 1 (% peak session 1: 20% ± 5%; session 9: 26% ± 5%; P = 0.002). There was a significant difference in the amplitude of oxygen off kinetics in session 9 compared to session 1 (session 1: 3.0 ± 0.7 L/min, session 9: 3.6 ± 1.0 L/min; P < 0.05; d = − 1.77), VO2 end (session 1: 0.59 ± 0.19 L/min, session 9: 0.81 ± 0.21 L/min; P < 0.05, d = − 0.90) and in time constant (session 1: 81 ± 21 s; session 9: 60 ± 11 s; P < 0.05; d = 1.03).

Change in recovery metrics. A Volume of oxygen consumed, B volume of carbon dioxide produced, C self-selected recovery time. *P < 0.05 session 1 compared to session 9
There was no significant difference in blood lactate between session 1 and 9 either immediately post sprint or after the recovery period (post sprint session 1: 13.3 ± 1.7 mmol/L; post sprint session 9: 14.5 ± 0.7 mmol/L; post recovery session 1: 12.5 ± 1.6 mmol/L; post recovery session 9: 13.6 ± 0.9 mmol/L). The change in blood lactate from rest to immediately post sprint was not significantly different between sessions (session 1: 11.2 ± 1.6 mmol/L; session 9: 12.1 ± 1.8 mmol/L). The change in blood lactate from rest to immediately post sprint was not significantly different between sessions (session 1: − 0.8 ± 0.8 mmol/L; session 9: − 0.6 ± 1.0 mmol/L).
Following multiple regression analysis of the recovery time, using last 60 s VO2, recovery between sets VO2 area under the curve, Amplitude, time constant and change in blood lactate after exercise as covariates, there was an R2 of 0.903 (P < 0.001). There was a significant effect for VO2 area under the curve (P = 0.015), last 60 s VO2 (P = 0.003), and amplitude (P = 0.041) with recovery time but not for time constant (P = 0.627) or change in blood lactate after exercise (P = 0.733). The standard β estimate for VO2 area under the curve was 0.49 (lower 95% CI 0.12, upper 95% CI 0.86); for last 60 s VO2 was − 0.55 (lower 95% CI − 0.85, upper 95% CI − 0.24) and for amplitude was − 0.33 (lower 95% CI − 0.64, upper 95% CI − 0.02).
Performance
Across the study there was no significant time effect for body mass, BMI, VO2peak, amplitude of power decline or time constant for power decline (P > 0.05; Table 3). There was a significant time effect for TTE (P < 0.05; η2p = 0.30; Table 3), CP30 (P < 0.05; η2p = 0.72; Table 3) and Y0 (P < 0.05; η2p = 0.68; Table 3). Following the intervention, TTE, CP30 and Y0 were significantly improved compared to pre 1 and pre 2 (P < 0.05; Table 3).
Following multiple regression analysis of the time to exhaustion or critical power, using last 60 s VO2, total work done in the lower body sprints, Amplitude, and time constant of oxygen off kinetics as covariates, there was an R2 of 0.96 for TTE (P < 0.001) and 0.94 for CP30 (P < 0.01). There was a significant effect for amplitude (P < 0.001), and time constant (P = 0.016) for both TTE and CP30. For TTE, the standard β estimate for amplitude was 0.95 (lower 95% CI 0.77, upper 95% CI 1.13) and for time constant was 0.22 (lower 95% CI 0.05, upper 95% CI 0.38). For CP30, the standard β estimate for amplitude was 0.87 (lower 95% CI 0.64, upper 95% CI 1.10) and for time constant was 0.31 (lower 95% CI 0.02, upper 95% CI 0.61).
Discussion
The major finding from this study is the improvement in self-selected recovery time across a lower and upper body sprint training protocol. Following 9 sessions, recovery time was reduced by 41% ± 10% compared to session 1 (Fig. 2C). This improvement in recovery time was associated with lower between sets recovery area under the curve for VO2 or VCO2 and a higher plateau value for VO2 or VCO2 but not to the metabolic demand of training or blood lactate levels post exercise or at the end of recovery. We also report significant training adaptations in aerobic performance without any improvement in maximal aerobic capacity (Table 3).
Lower Body Sprints
Over the 9 sessions there was no significant change in lower body peak or average power across any sprint. Despite no change in power production there was a significant increase in the extent of oxygen consumption as determined by area under the curve for each sprint (Table 1). This suggests a greater aerobic demand for each sprint with potentially a lower anaerobic demand following training. A progressive 30 s sprint protocol over 8 weeks has been shown to have no effect on anaerobic demand during sessions but result in an increased aerobic load [4]. In contrast to the current study the authors report an increase in average power across the training sessions [4] however training frequency was greater and there was a larger work to rest ratio then used in this study. It has been well documented that larger work to rest ratios promote changes in power production [20]. Sprint duration does not impede aerobic adaptations to SIT [40, 42] suggesting similar mitochondrial adaptations or capillarisation changes regardless. Therefore, the greater VO2 AUC during the lower body sprints, reflects the greater capacity for the muscle to utilise aerobic metabolism.
The overall AUC for the recovery after the lower body sprints was significantly lower from session 1 and session 9 (Table 1). Whilst the AUC is lower, this reflects the shorter recovery time, and the overall oxidative demand of the recovery period is higher (Fig. 2A). This is reflected by the significant increase in VO2 during the last 60 s of recovery. There is also a lower AUC for VCO2, with a greater VCO2 during the last 60 s of recovery (Table 2, Fig. 2B). Together with the greater VO2 load this suggests a greater metabolic load during the recovery phase following 9 sessions of SIT. With greater oxygen consumption over the recovery period there will be greater opportunity for muscle oxygen transport and increased oxidative metabolism leading to a faster rate of phosphocreatine recovery [26].
In session 9 the oxygen off kinetics were different from session 1 with a significant increase in amplitude and a significant reduction for the time constant. The increase in amplitude reflects the increased oxygen demand of the recovery period post sprints as shown by the change in AUC. The reduction in time constant of off kinetics has been demonstrated previously in response to training [6] or with endurance training status [10]. In judo fighters, the time constant for recovery was reduced by 22% over 12 weeks of judo specific high intensity training [7], which is similar to the reduction seen following 9 sessions of cycle based SIT in the current study (23% ± 19%). Within football players, faster time constant of VO2 off kinetics has been strongly associated with improved intermittent exercise performance [15]. Together this improved speed of oxygen recovery post sprints suggests a faster rate of removal of possible fatigue metabolites and a maintenance of intermittent performance.
Time to Recovery
Following 9 sessions of SIT, there was a 173 s reduction in self-selected recovery time between lower body and upper body sprints (Fig. 2C). This much faster recovery from exercise was strongly associated with the amplitude of oxygen off kinetics, last 60 s VO2 during recovery and the AUC for VO2. This suggests that time for recovery is linked to the oxidative demand during recovery and the new plateau seen for both VO2 and VCO2 following training (Figs. 2A and B). Given that the same power output was seen across sprint session then the elevated levels of VO2 and increased amplitude reflect greater oxidative metabolism across the recovery time in the skeletal muscle. Recovery of force post exercise has been shown to be related to increased citrate synthase activity and oxidative potential of the skeletal muscle [19] and PCr recovery has been shown to be limited by oxygen availability post exercise [18]. Group III.IV afferent activity have been shown to limit oxygen delivery to the skeletal muscle [2], therefore the SIT training may induce less afferent feedback during recovery which promotes a faster recovery time. Further research is needed to look at the effect of SIT on fixed recoveries that are typically seen in combat sports.
Upper Body Sprints
The aerobic demand, as assessed by VO2 AUC, of the upper body sprints are significantly different in session 9 compared to session 1 for all sprints except the final sprint (Table 1). Across the first three sprints there is a gradual increase in VO2 AUC and then oxidative contribution to the sprints plateau (Table 1). A similar plateau is seen in oxidative contribution across 4 × 30 s upper body Wingates [16]. However, following upper body SIT utilising longer work to rest ratios there was no change in aerobic demand during an upper body Wingate [22]. For lower body sprint training it has been established that longer work to rest ratios limit aerobic adaptation [20] and it seems reasonable to assume this will be a key component for upper body adaptation as well. Given that aerobic metabolism has been suggested to contribute close to 50% of total energy during an upper body Wingate [29], then this increased aerobic demand may be beneficial to overall upper body performance.
Aerobic Performance Adaptations
Following 9 sessions of SIT, there were significant improvements in incremental time to exhaustion and critical power/Y0 but no change in VO2peak (Table 3). The improvement in both time to exhaustion and critical power are similar to those reposted previously following SIT [40] and were strongly related to the change in amplitude and time constant of the off-oxygen kinetics. Given the symmetry reported between on and off oxygen kinetics [36] then this relationship may reflect improvements in on oxygen kinetics during the exercise. It has been suggested that poor oxygen kinetics leads to the build up of anaerobic by-products which leads to the early onset of fatigue [37]. Improvements in oxygen delivery and utilisation during exercise will increase the intensity where anaerobic demand is delivered and lead to a higher power output through oxidative metabolism [8]. It has been established that improved mitochondrial density and mitochondrial enzyme activity within skeletal muscle occurs following SIT training [23] and computer modelling highlights the importance of mitochondrial function for oxygen on kinetics [21]. Following 2 weeks of high intensity interval training, using 60 s intervals, an improvement in mitochondrial enzyme function has been linked to improvements in oxygen on kinetics during exercise [11]. Given that duration of interval for training has been shown to have no impact on mitochondrial adaptation [23] then it seems reasonable to assume a similar response in the current study. Further research is needed to determine the upper body adaptation to this type of training protocol.
Practical Applications and Conclusion
The ability to rapidly utilise oxidative metabolism is crucial for repeated high intensity events to prevent the early onset of fatigue [37]. Given the intermittent nature of kickboxing improving skeletal muscle oxidative function is crucial for recovery and to maintain a high work rate across competition [13]. The findings from this study suggest that the adaptations from cycle-based SIT would be beneficial for kickboxers, promoting faster recovery and delaying the onset of fatigue, without any change in maximal aerobic capacity. Further the increased aerobic function of the upper body during sprints may allow an athlete to maintain a high upper body work rate meaning the athlete is able to maintain defensive as well as offensive work rate [14]. Given that winners have a higher frequency of attacking [28] then improved aerobic function will enhance ability to maintain the required work rate. There is a strong correlation between endurance components and self-selected recovery, however further work is required to look at recovery in a more sport specific way (such as simulated competition) to fully understand the important physiological changes to enhance performance.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Akgul MS. Effect of Wingate-based high intensity interval training on aerobic and anaerobic performance of kick boxers. Phys Educ Stud. 2019;23(4):167–71. https://doi.org/10.15561/20755279.2019.0401.
Amann M. Significance of group III and IV muscle afferents for the endurance exercising human. Clin Exp Pharmacol Physiol. 2012;39(9):831–5. https://doi.org/10.1111/j.1440-1681.2012.05681.x.
Bakeman R. Recommended effect size statistics for repeated measures designs. Behav Res Methods. 2005;37(3):379–84. https://doi.org/10.3758/bf03192707.
Barnett C, Carey M, Proietto J, Cerin E, Febbraio MA, Jenkins D. Muscle metabolism during sprint exercise in man: influence of sprint training. J Sci Med Sport. 2004;7(3):314–22. https://doi.org/10.1016/s1440-2440(04)80026-4.
Beekley MD, Abe T, Kondo M, Midorikawa T, Yamauchi T. Comparison of normalized maximum aerobic capacity and body composition of sumo wrestlers to athletes in combat and other sports. J Sports Sci Med. 2006;5(CSSI):13.
Billat VL, Mille-Hamard L, Demarle A, Koralsztein JP. Effect of training in humans on off- and on-transient oxygen uptake kinetics after severe exhausting intensity runs. Eur J Appl Physiol. 2002;87(6):496–505. https://doi.org/10.1007/s00421-002-0648-7.
Bonato M, Rampichini S, Ferrara M, Benedini S, Sbriccoli P, Merati G, Franchini E, La Torre A. Aerobic training program for the enhancements of HR and VO2 off-kinetics in elite judo athletes. J Sports Med Phys Fitness. 2015;55(11):1277–84.
Burnley M, Jones AM. Oxygen uptake kinetics as a determinant of sports performance. Eur J Sport Sci. 2007;7(2):63–79. https://doi.org/10.1080/17461390701456148.
Campos EZ, Bastos FN, Papoti M, Freitas Junior IF, Gobatto CA, Balikian Junior P. The effects of physical fitness and body composition on oxygen consumption and heart rate recovery after high-intensity exercise. Int J Sports Med. 2012;33(8):621–6. https://doi.org/10.1055/s-0031-1295442.
Caputo F, Denadai BS. Effects of aerobic endurance training status and specificity on oxygen uptake kinetics during maximal exercise. Eur J Appl Physiol. 2004;93(1–2):87–95. https://doi.org/10.1007/s00421-004-1169-3.
Christensen PM, Jacobs RA, Bonne T, Flück D, Bangsbo J, Lundby C. A short period of high-intensity interval training improves skeletal muscle mitochondrial function and pulmonary oxygen uptake kinetics. J Appl Physiol (Bethesda, Md: 1985). 2016;120(11):1319–27. https://doi.org/10.1152/japplphysiol.00115.2015.
Cohen J. A power primer. Psychol Bull. 1992;112(1):155–9. https://doi.org/10.1037//0033-2909.112.1.155.
Davis P, Leithäuser RM, Beneke R. The energetics of semicontact 3 x 2-min amateur boxing. Int J Sports Physiol Perform. 2014;9(2):233–9. https://doi.org/10.1123/IJSPP.2013-0006.
Dunn EC, Humberstone CE, Iredale KF, Martin DT, Blazevich AJ. Human behaviours associated with dominance in elite amateur boxing bouts: a comparison of winners and losers under the Ten Point Must System. PLoS ONE. 2017;12(12): e0188675. https://doi.org/10.1371/journal.pone.0188675.
Dupont G, McCall A, Prieur F, Millet GP, Berthoin S. Faster oxygen uptake kinetics during recovery is related to better repeated sprinting ability. Eur J Appl Physiol. 2010;110(3):627–34. https://doi.org/10.1007/s00421-010-1494-7.
Franchini E, Takito MY, Dal’Molin Kiss MA. Performance and energy systems contributions during upper-body sprint interval exercise. J Exerc Rehabil. 2016;12(6):535–41. https://doi.org/10.12965/jer.1632786.393.
Hair JF, Black WC, Babin BJ, Anderson RE. Multivariate data analysis. 7th ed. Upper Saddle River: Prentice Hall; 2010.
Haseler LJ, Hogan MC, Richardson RS. Skeletal muscle phosphocreatine recovery in exercise-trained humans is dependent on O2 availability. J Appl Physiol (Bethesda, Md 1985). 1999;86(6):2013–8. https://doi.org/10.1152/jappl.1999.86.6.2013.
Jansson E, Dudley GA, Norman B, Tesch PA. Relationship of recovery from intensive exercise to the oxidative potential of skeletal muscle. Acta Physiol Scand. 1990;139(1):147–52. https://doi.org/10.1111/j.1748-1716.1990.tb08907.x.
Kavaliauskas M, Aspe RR, Babraj J. High-intensity cycling training: the effect of work-to-rest intervals on running performance measures. J Strength Cond Res. 2015;29(8):2229–36. https://doi.org/10.1519/JSC.0000000000000868.
Korzeniewski B, Rossiter HB, Zoladz JA. Mechanisms underlying extremely fast muscle \(\dot {\text{V}}\)O2 on-kinetics in humans. Physiol Rep. 2018;6(16):e13808. https://doi.org/10.1414/phy2.13808.
La Monica MB, Fukuda DH, Starling-Smith TM, Clark NW, Panissa V. Alterations in energy system contribution following upper body sprint interval training. Eur J Appl Physiol. 2020;120(3):643–51. https://doi.org/10.1007/s00421-020-04304-w.
MacInnis MJ, Gibala MJ. Physiological adaptations to interval training and the role of exercise intensity. J Physiol. 2017;595(9):2915–30. https://doi.org/10.1113/JP273196.
Mandić M, Eriksson LMJ, Melin M, Skott V, Sundblad P, Gustafsson T, Rullman E. Increased maximal oxygen uptake after sprint-interval training is mediated by central haemodynamic factors as determined by right heart catheterization. J Physiol. 2023;601:2359–70. https://doi.org/10.1113/JP283807.
Mann TN, Lamberts RP, Lambert MI. High responders and low responders: factors associated with individual variation in response to standardized training. Sports Med. 2014;44(8):1113–24.
McMahon S, Jenkins D. Factors affecting the rate of phosphocreatine resynthesis following intense exercise. Sports Med (Auckland, NZ). 2002;32(12):761–84. https://doi.org/10.2165/00007256-200232120-00002.
Monks L, Seo MW, Kim HB, Jung HC, Song JK. High-intensity interval training and athletic performance in Taekwondo athletes. J Sports Med Phys Fitn. 2017;57(10):1252–60. https://doi.org/10.23736/S0022-4707.17.06853-0.
Ouergui I, Davis P, Houcine N, Marzouki H, Zaouali M, Franchini E, Gmada N, Bouhlel E. Hormonal, physiological, and physical performance during simulated kickboxing combat: differences between winners and losers. Int J Sports Physiol Perform. 2016;11(4):425–31. https://doi.org/10.1123/ijspp.2015-0052.
Price M, Beckford C, Dorricott A, Hill C, Kershaw M, Singh M, Thornton I. Oxygen uptake during upper body and lower body Wingate anaerobic tests. Appl Physiol Nutr Metab Physiol appliquee, nutrition et metabolisme. 2014;39(12):1345–51. https://doi.org/10.1139/apnm-2013-0405.
Rakobowchuk M, Tanguay S, Burgomaster KA, Howarth KR, Gibala MJ, MacDonald MJ. Sprint interval and traditional endurance training induce similar improvements in peripheral arterial stiffness and flow-mediated dilation in healthy humans. Am J Physiol Regul Integr Comp Physiol. 2008;295(1):R236–42.
Raleigh JP, Giles MD, Islam H, Nelms M, Bentley RF, Jones JH, Neder JA, Boonstra K, Quadrilatero J, Simpson CA, Tschakovsky ME. Contribution of central and peripheral adaptations to changes in maximal oxygen uptake following 4 weeks of sprint interval training. Appl Physiol Nutr Metab. 2018;43(10):1059–68. https://doi.org/10.1139/apnm-2017-086.
Reinpõld K, Rannama I. Oxygen uptake and bilaterally measured vastus lateralis muscle oxygen desaturation kinetics in well-trained endurance cyclists. J Funct Morphol Kinesiol. 2023;8(2):64. https://doi.org/10.3390/jfmk8020064.
Silva JJR, del Vecchio FB, Picanso LM, Takito MY, Franchini E. Time motion analysis in Muay Thai and Kickboxing amateur matches. J Hum Sport Exerc. 2011;6(3):490–6.
Slimani M, Chaabene H, Miarka B, Chamari K. The activity profile of elite low-kick kickboxing competition. Int J Sports Physiol Perform. 2015;12(2):182–9. https://doi.org/10.1123/ijspp.2015-0659.
Slimani M, Chaabene H, Miarka B, Franchini E, Chamari K, Cheour F. Kickboxing review: anthropometric, psychophysiological and activity profiles and injury epidemiology. Biol Sport. 2017;34(2):185–96. https://doi.org/10.5114/biolsport.2017.65338.
Sousa A, Rodríguez FA, Machado L, Vilas-Boas JP, Fernandes RJ. Exercise modality effect on oxygen uptake off-transient kinetics at maximal oxygen uptake intensity. Exp Physiol. 2015;100(6):719–29. https://doi.org/10.1113/EP085014.
Temesi J, Mattioni Maturana F, Peyrard A, Piucco T, Murias JM, Millet GY. The relationship between oxygen uptake kinetics and neuromuscular fatigue in high-intensity cycling exercise. Eur J Appl Physiol. 2017;117(5):969–78. https://doi.org/10.1007/s00421-017-3585-1.
Tomlin DL, Wenger HA. The relationship between aerobic fitness and recovery from high intensity intermittent exercise. Sports Med (Auckland NZ). 2001;31(1):1–11. https://doi.org/10.2165/00007256-200131010-00001.
Vanhatalo A, Doust JH, Burnley M. Determination of critical power using a 3-min all-out cycling test. Med Sci Sports Exerc. 2007;39(3):548–55. https://doi.org/10.1249/mss.0b013e31802dd3e6.
Yamagishi T, Babraj J. Effects of reduced-volume of sprint interval training and the time course of physiological and performance adaptations. Scand J Med Sci Sports. 2017;27(12):1662–72. https://doi.org/10.1111/sms.12831.
Zafeiridis A, Dalamitros A, Dipla K, Manou V, Galanis N, Kellis S. Recovery during high-intensity intermittent anaerobic exercise in boys, teens, and men. Med Sci Sports Exerc. 2005;37(3):505–12. https://doi.org/10.1249/01.mss.0000155394.76722.01.
Zelt JG, Hankinson PB, Foster WS, Williams CB, Reynolds J, Garneys E, Tschakovsky ME, Gurd BJ. Reducing the volume of sprint interval training does not diminish maximal and submaximal performance gains in healthy men. Eur J Appl Physiol. 2014;114(11):2427–36. https://doi.org/10.1007/s00421-014-2960-4.
Acknowledgements
The authors would like to thank the participants for their time when taking part in this study and the support of the University for carrying out the study.
Funding
No external funding was received for this study.
Author information
Authors and Affiliations
Contributions
Conceptualization, AH and JB; methodology AH and JB.; formal analysis, AH, AW and JB; investigation, AH; writing—original draft preparation, JB; writing—review and editing, AH, AW and JB; visualization, AH, AW and J.B.; supervision, AW and JB. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
No conflicts of interest.
Ethical approval
Ethical approval was obtained from the ethics committee of Abertay University. The procedures used in this study adhere to the tenets of the Declaration of Helsinki.
Consent to participate
All participants gave informed consent prior to taking part.
Consent to publish
All participants gave consent for publication of data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hall, A., Williams, A.K. & Babraj, J. Training for Recovery: Impact of Sprint Interval Training on Recovery Dynamics and Aerobic Performance in Kickboxing Athletes. J. of SCI. IN SPORT AND EXERCISE (2024). https://doi.org/10.1007/s42978-023-00273-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42978-023-00273-6