
Abstract
Cryptogams of ten urban flatroofs, contrasting in their age and size, were studied between 2016 and 2018. Siliceous (bituminous felt, gravel, brick) and calcareous (concrete) substrata occurred at each site. Microclimate (T, RH) at two sites of contrasting shading was monitored from September 2016 to January 2017. Biomass of two differently aged, exposed flatroofs was sampled in October 2018. Taxa of Cladonia and Xanthoparmelia have been identified by spot tests and HPTLC. A total of 61 taxa (25 bryophytes, 36 lichens), mostly widespread synanthropic species, have been detected with an explicit difference of species composition between shaded and exposed sites. Floristically interesting species included acidophilous bryophytes (Hedwigia ciliata, Racomitrium canescens) and lichens (Xanthoparmelia conspersa, Stereocaulon tomentosum) of montane character. The most widespread lichen is Cladonia rei which accounted for a significant part of the biomass at selected sites. Species-area curves for bryophytes at exposed sites have become saturated at 100–150 m2. In contrast, saturation of lichen diversity has not been reached even at the largest sites. Flatroofs with traditional roofing techniques can harbour relatively diverse microhabitats and species-rich synanthropic vegetation. It is urgent to study these sites before renovation with modern roofing techniques eliminates them. Diversification of urban surroundings is possible in the future via application of various substrats in renovated and newly constructed roofs.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Cryptogams are known as effective first colonizers in natural (Gilbert 1990) as well as in urban habitats. Construction activity in cities continuously creates new habitats on buildings, but compared to their large extension very little information has been gathered on their cryptogamic flora. Species composition of certain microhabitats has been somewhat better explored for bryophytes than for lichens (Fudali 2006; Sabovljević and Grdović 2009), but overall diversity of the bryophyte flora within settlements is generally neglected. The bryophytes are likely to form a significant part of urban floras, but the available data on them are limited (Skudnik et al. 2013). In Hungary, apart from a few bryophyte records from some towns, only two studies are known, which examined the bryophyte diversity of villages (Zsólyom and Szűcs 2018). Recently the bryophytes are increasingly used in’green roofs’ (Glime 2017; Studlar and Peck 2011), but the number of publications dealing with spontaneous appearance of mosses on the roofs remains very low. These mainly focused on the patterns of bryophyte species richness and community structure of thatched roofs, but their microenvironmental variables are not described directly (Hedderson et al. 2003).
Although the urban lichen floras, as indicators of air pollution, have been subjects to intensive studies, the surveys are mostly confined to epiphytes (Skye 1968; Farkas et al. 2001). Much less recent information is available on epilithic and epigeic, especially building-dwelling lichens (Lisická 2008; Matwiejuk 2009). Some historical buildings (e.g. churches) represent important exceptions (Piervittori et al. 1994; Bungartz 1999). In contrast, data on establishment of lichens on modern buildings, including the recently widespread flatroofs, are very scarce. Spontaneous colonization of lichens has been observed on some roofs (Thuring and Dunnett 2019; Köehler and Kaiser 2021).
A similar data shortage prevails for Hungary as we mostly have historical records of synanthropic lichen vegetation, from river embankments and bridges (Gallé 1965). Only sporadic synanthropic records are scattered in floristic studies (e.g. Lőkös and Farkas 1998, 2000; Farkas and Lőkös 2003; Szűcs and Lóth 2008). No modern survey is available concentrating on floristic effect of roofing techniques (cf. Favero-Longo et al. 2005; Gazzano et al. 2009), although techniques and materials applied during construction have greatly changed meanwhile. Modern techniques were applied on lichens colonizing monuments and while bioremediation potential of lichens were studied on various artificial and natural substrates (Cilia et al. 2015; Morando et al. 2019).
Cryptogams, due to their dispersal by spores and contrary to dispersal-limited seed plants, are known to map microclimatic heterogeneities and make excellent indicator organisms of microhabitats (Birks et al. 1998). Microclimate is a crucial determinant of ecological patterns and is a driver of process in both vascular (Kershaw and Larson 1974; Behera et al. 2012) and non-vascular plant communities. Most studies concerning microclimate and surface temperature have focused on vascular plants, with only a few studies on bryophytes (Van Tooren et al. 1985) and even fewer on lichens (Aartsma et al. 2021) despite them being an important component in some communities. Temperature and water availability are the primary factors of the growth rate and the distribution of cryptogams (Nash 2008; Glime 2017). Temperature typically defines the boundaries of their distribution while water availability limits their growth and distribution within those boundaries (Glime 2017). As for taxonomic groups, humidity has been identified as a decisive factor for the bryophytes. Bryophytes generally prefer more shaded conditions and higher air humidity (Humphrey et al. 2002; Ranius et al. 2008). In contrast, in lichens sensitive to light availability proved more pronounced (Humphrey et al. 2002; Ranius et al. 2008; Aptroop 2010; Nascimbene et al. 2013; Ódor et al. 2013).
In contrary to natural pioneer habitats (volcanoes, glacier forelands), their dispersal limitation has hardly been studied in urban surroundings where they might colonize apparently inhospitable habitats (bare rocks, walls, roofs). Goffinet and Shaw (2009) suspect that these associations often have a chemical basis as far as the substratum is concerned, although in other cases some other ecophysiological explanations may exist. They can fairly be considered as habitat forming organisms which enabled the subsequent succession of the vascular plants.
Due to their reduced extension, survey of cryptogamic assemblages, differs greatly from those in vascular communities. Berg et al. (2016) demonstrated a large overall variation of applied plot sizes ranging from 0.0002 to 30 m2 for lichen communities. These figures spread over five orders of magnitude, which is even more extreme than those reported for relevés in vascular plant communities (Chytrý and Otýpková 2003). The most frequently used cryptogam plot sizes were 0.04, 0.01 and 0.03 m2, respectively. Güler et al. (2016) recommend that, wherever possible, plots of cryptogamic micro-communities should be square shaped (20 cm × 20 cm or 10 cm × 10 cm).
The basic aim of our study was to take an inventory of cryptogams (lichens and bryophytes) dwelling on a representative number of closely located urban flatroofs. We wanted to characterize microclimatic conditions of exposed and shaded flatroofs, respectively, as the most obvious abiotic background factor of their contrasting species compositions under the same macroclimate. We intended to compare total cryptogamic biomass as well as its division between lichen and bryophyte fractions on flatroofs of contrasting ages. This way we wanted to contribute to the knowledge of phasing and timing of primary succession acting on urban flatroofs. By comparing lichen and bryophyte species richness of different-sized flatroof we intended to establish a species-area relationship for urban flatroof cryptogams. This way we could also test whether is there a correlation between lichen and bryophyte species richness, respectively, in these widespread urban habitats.
Material and methods
Site selection and survey
Suitable sampling sites have been selected using public databases of colour aerial photographs (GoogleEarth, http://www.viamichelin.com). Pictures taken 0–10 years prior to surveys have been screened to select old, non-renovated flatroofs. These show an apparent patchy pattern of cryptogamic vegetation, in contrast to the uniform surface of new or renovated ones. Access to roofs was permitted by management of the selected institutions upon request. Information on the age of buildings was gained from facility managers. Roof dimensions were measured on public aerial photos. Participants of field sampling have accomplished a course of labour safety tuition by the Autonomous Division of Labour Safety, Debrecen University prior to working on roofs.
Floristical surveys (photodocumentation, species inventory, collection of specimens) were carried out from March to June, 2016. The whole surface of ten urban roofs has been surveyed, and the presence-absence data of cryptogams have been recorded for the whole roof area. Specimens have been collected when necessary, and finally, species lists have been compiled. Many cryptogam species have been identified on the spot, while others have been transported to the laboratory and then have further been examined. Survey time varied proportional to roof size and also to structural diversity of roof surface (e.g. amount of walls, chimneys present). Duration of surveys lasted from 0.5 to 2 h with two to three surveyors, respectively, in a way that longer times and more surveyors have been applied in larger and more diverse roofs. Usually one survey per roof has been applied but in case of the largest roofs some additional survey has also been conducted when the microclimate measuring apparatus has been checked.
Site characteristics
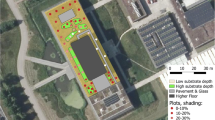
All selected sites are non-exploited flatroofs (i.e. except for repairs no access of people is allowed), therefore human disturbance is very scarce. All sites are almost level with a fall as little as 1:80 to 1:40 to ensure the rainwater runoff. At most studied sites insulation regime was ‘normal’ (also referred to as ‘conventional’) meaning that thermal insulation is placed under a protective membrane. In this case this membrane was bituminous felt covered by a weatherproofing layer of gravel coat (consisting mostly of siliceous pea gravel of 10–25 mm diameter). At a single site (1, LSB) ‘inverted roof’ regime (also referred to as ‘upside down’ roof) was applied where the waterproofing layer is beneath the thermal insulation. In this case, the uppermost cover layer was gravel ballast (typically siliceous gravel of 25–60 mm diameter). Roofing also contained vertical structures (chimneys, ventilation structures, small bordering walls) made of non-calcareous (brick) as well as calcareous materials (concrete walls and wall cappings) at all sites. Location, size, age and other site characteristics are summarized in Table 1.
Identification
Lichen identification has been completed by manuals of Verseghy (1994), Smith et al. (2009) and Wirth et al. (2013). Nomenclature of lichens follows the public database of IndexFungorum (http://www.indexfungorum.org/, accessed on 9th March, 2023). Not-collected thalli of crustose lichens have been observed by hand lens and documented by macro photos in the field while collected specimens have been identified using stereo and compound microscopes (Olympus SZX9 and Olympus BX50) as well as by spot tests (Orange et al. 2010). For 40 specimens from the critical lichen genera Cladonia, Stereocaulon and Xanthoparmelia standard high-performance thin-layer chromatography (HPTLC) in solvent system C (toluene: acetic acid = 20: 3, V/V) has been applied (Arup et al. 1993). This has been applied in order to determine the pattern of secondary metabolites, an inevitable trait for identification. Approximately 5 mm × 5 mm air-dried thallus fragments were soaked in 0.2 ml acetone for 30 min in order to extract the lichen substances. Pretreated (50 °C for 5 min, CAMAG TLC Plate Heater III, then cooled to room temperature), 10 cm × 10 cm thin-layer chromatographic plates (Merck, Kieselgel 60 F254) were used. Eight µl acetone extracts (1 µl at each time) were applied to each position (5 mm apart) on the plate using a microapplicator (CAMAG Micro Applicator I). Pleurosticta acetabulum (Neck.) Elix & Lumbsch (norstictic acid) and Heterodermia leucomelos (L.) Poelt (atranorin, zeorin) were used as controls. After chromatographic development, the plates were examined under UV light (254 and 366 nm), then sprayed (CAMAG TLC Sprayer) with water, dried (to investigate fatty acids), then subsequently sprayed with a 10% sulphuric acid solution and heated at 110 °C for 5–10 min. Finally, the plates were cooled to room temperature and studied under UV light (366 nm).
Bryophytes have been identified by consulting the relevant handbooks and papers as Erzberger and Schröder (2013), Erzberger et al. (2016), while the nomenclature for bryophytes follows Hodgetts et al. (2020). A Wild dissecting microscope and a Leica light microscope were used.
Collected bryophyte specimens are deposited in the Botanical Garden and Botanical Museum Berlin-Dahlem (B) and in the Cryptogamic Herbarium of the Department of Botany and Plant Physiology at the Eszterházy Károly University, Eger (EGR). Lichen specimens have been deposited in the Herbarium of the Hungarian Natural History Museum, Budapest (BP), and the Herbarium of the Debrecen University, Debrecen (DE).
For all cryptogamic specimens identifiers of the collecting sites by quadrates ([8495.2] and [8495.4]) according to the Central European Flora Mapping System (Niklfeld 1971; Király et al. 2003) were indicated.
Microclimate meaurement
Microclimate measurements have been conducted on two roofs of different exposition. The selected buildings, the 4th Nurse Hostel (NH, exposed roof) and the Life Sciences Building (LSB, highly shaded terrace) are located close to each other (< 450 m). Temperature (T) has been recorded at 5 cm and 30 cm height, resprectively, while relative humidity (RH) was measured at 30 cm height. Measurements have been stored in 60 min intervals and analyzed between 1st September 2016 and 31st January 2017 using HOBO Temperature/Relative humidity dataloggers (Onset Computer Corporation, USA, series H08–003–02 and U12–011).
Due to low batteries, data have not been gathered at one datalogger between 16 and 20th of December, 2016. Data of other loggers from this period have therefore been excluded from analyses and a total duration of 148 days has been analysed.
Duration of direct sunlight has been calculated using HemiView hemispherical canopy analyser (Delta-T Devices Ltd, UK) at the Life Sciences Building. Based upon measurements and in coincidence with personal observations, we have shown that only small parts of this site receive any direct sunlight during the year in the morning and evening, respectively. Direct sunlight enters here only between 8th May and 4th August. The maximum length of direct insolation at these parts is only 75 min on 22nd June.
Temperature measurements have additionally been compared with standard records (at 2 m height) of the nearby meteorological station (< 400 m from LSB and < 700 m from NH) operated at the university campus by the Department of Meteorology and the Renewable Energy Park (N 47° 33.355′, E 21° 36.925′). Comparison of our microclimate measurements and the reference readings was, however, possible only to a limited extent due to a repeated malfunctioning of measurement tools at campus station. Thus we have selected three days in each month (7th, 12th and 21st, respectively, representing ca. 10% of measuring days) when standard measurement have been compared with ours.
Biomass sampling
Cryptogamic biomass was sampled on the flatroofs of contrasting age of the Lehel street kindergarten (37 years old) and the big garage of the Nursery Home (49 years old) on 16 October 2018. These are located about 780 m apart and were similarly exposed and isolated with identical technologies. In each flatroof, 10 samples of 10 cm × 10 cm (0.01 m2) were collected, then samples were dried, hand sorted into bryophytes and lichens and these fractions were measured with an accuracy of 0.01 g.
Data processing
Microclimate
Pairwise and multiple comparisons have been performed by Sigmaplot 12.0 programme package. For comparison of daily average, maximum, minimum as well as range of temperature and of relative humidity records for the full period (148 days) ANOVAs on ranks (Kruskal–Wallis test) have been applied whereas pairwise significancies among differently positioned dataloggers (shaded vs. exposed, 5 cm vs. 30 cm) has been determined by the Dunn’s test with p < 0.05 significance values (Zar 2010). Paired comparisons of records from the 5 cm readings of shaded and the exposed roof, respectively, has been performed by Wilcoxon signed rank tests (non-parametric version of paired t-test).
Similarly, ANOVAs on ranks (Kruskal–Wallis test) have been applied to compare daily average, maximum, minimum as well as range of temperature and of relative humidity records in each sampled months. In case of significant ANOVAs pairwise comparisons have been completed by Dunn’s tests with p < 0.05 significance values (Zar 2010). Paired comparisons of records from the 5 cm readings of shaded and the exposed roof, respectively, has been performed by Wilcoxon signed rank tests.
On hourly temperature readings of selected days (three days per months) we used Wilcoxon signed rank tests for paired comparisons of microclimate dataloggers with standard reference dataset (five comparisons).
Biomass
Pairwise comparison of biomass samples (lichens, bryophytes and all cryptogams, respectively) collected on roofs of contrasting age has been completed by Mann–Whitney rank sum test (Mann and Whitney 1947).
Floristic composition
Correlation of bryophyte and lichen species numbers per site has been tested by Spearman rank order correlation (Spearman 1904). Species composition of sites has been analysed using DCA in the programme package CANOCO and visualized in CANODRAW (Braak and Smilauer 2002). Rare species, occurring at a single site and the species-poor site 10 have been excluded from multivariate analysis. Presence-absence records for a total of 29 taxa (20 lichens, 9 bryophytes, thus 47.5% of all recorded taxa, representing 79.4% of all records) have been analysed.
Species richness
Species-area analyses have been applied for lichen, bryophyte and total species richness of sites, except for the LSB roof with deep shade and a species composition contrasting all the rest. We used a single component, two parameter version of exponential rise to maximum equations, a widely accepted one used in describing saturation curves from accidental sources. The used equation was f = a*(1-exp(− b*x)) where x is site area in m2 while f is the species number of bryophytes and lichens at given site, respectively, whereas a and b are parametres to be calculated by an iterative approach with automatic initial parameter estimate functions. We used the software package option dynamic fitting (with 200 fits while the maximal number of iterations was also 200) so as to get the best possible estimates for parametres and to achieve the best possible fit.
Results
Microclimate
For the entire sampling period (148 days), average of temperature (T) fell between 3.97 and 7.02 °C among the four dataloggers (Suppl. Table 1). These differences, however, did not prove to be significant by the Kruskal–Wallis test due to their large variation through the sampled period (Suppl. Table 2). The same holds for absolute minimum scores (between −16.02 and −17.17 °C). In contrast, absolute maximum values (between 25.95 and 47.43 °C) and ranges of temperature (between 41.97 and 64.11 °C) were already highly significantly different (p < 0.001) among the stations (Suppl. Table 1; Suppl. Table 2). All comparisons of relative humidity (average, maximum, minimum, range) for the whole sampled period resulted in highly significant values according to the Kruskal–Wallis tests (Suppl. Table 2). Paired comparisons of the surface (5 cm) measurements at shaded and exposed roofs proved to be highly significant for average, maxima and range in temperature, but non-significant for the minima (Wilcoxon signed rank tests, N = 148). For relative humidity, differences between the shaded and exposed roofs proved highly significant in all parametres but the maxima (Suppl. Table 2, grey columns). The exposed roof had much larger range of variation both for temperature (due to extreme high values) and relative humidity (due to extreme low values) data in both heights (5 cm and 30 cm, respectively) than did loggers of the shaded one. Absolute temperature maximum of exposed roof was about 10 °C higher at 30 cm and over 20 °C higher at 5 cm height (Suppl. Table 1).
Monthly mean (daily mean, daily maximum, daily minimum, daily range) at the exposed and shaded sites at 5 cm height for temperature are summarized in Table 2 and for relative humidity in Suppl. Table 3. Similar records are summarized at 30 cm height for temperature and relative humidity in Suppl. Table 4. Monthly comparisons have shown a similar patterns. No significant difference occured for minimum temperature records, only a few for averages whereas all montly comparisons of maximum temperatures as well as of temperature ranges proved to be significant by ANOVAs on ranks (Suppl. Table 5). Frequencies of significant pairwise comparisons of temperature by Dunn’s test were also similar (in the same order as the previous one, counting 0, 4, 18 and 16 significant differences, respectively). Paired comparisons of temperature maxima and range of surface measurements (5 cm) of the exposed and shaded roofs by Wilcoxon signed rank tests differed with high significance for all months (Table 3). Contrary to these, comparisons for average and minimum values often proved to be less or non-significant.
Maxima and ranges of relative humidity (RH) scores proved to be highly significant in all cases, but monthly comparisons of their minima and average resulted in some less or non-significant ANOVAs on ranks. Frequencies of significant pairwise comparisons of RH by Dunn’s test, however, differed somewhat less frequently than those of temperature. Paired comparisons of ranges of relative humidity in surface measurements (5 cm) always differed with high significance by Wilcoxon signed rank tests. Somewhat less highly significant differences have been revealed for maximum, average and minimum values (Table 3).
Differences among differently positioned datasets have shown a clear seasonality. Most of the highly significant ANOVAs on ranks (p < 0.001) occured in September but non-significant results peaked in December and January. Frequencies of significant pairwise differences (by the Dunn’s tests) had their maxima in September both for temperature and relative humidity data. Contrary to the previous results, their minima fell in December (T) and January (RH), respectively (Suppl. Table 5). Similarly, all paired comparisons between exposed and shaded surfaces (5 cm) proved to be highly significant in September, but only four to five of them from November to January. We have found that data from loggers of the exposed roof differed half as often from the reference than did those from the shaded roof (Table 3).
On selected days, pairwise comparisons of microclimate dataloggers with standard reference dataset at hourly temperature readings have shown that the highly significant differences (by Wilcoxon signed rank tests) have been about twice as frequent at shaded roof records than at the exposed ones (Suppl. Table 6, 7).
Daily regimes of temperature and relative humidity in September 2016 measured at 5 cm are shown in Fig. 1. (Temperature and RH data for 30 cm height can be found at Suppl. Figure 1). At a selected typical early autumn day (12 September 2016) both at 5 cm and 30 cm height, much lower and delayed daily maxima (between 2 and 3 p.m.) have been recorded on the shaded site than at the exposed one where maxima fell between 12 a.m. and 1 p.m. (Fig. 2). A similar delay of daily RH minima of RH has been found in autumn months (Suppl. Figure 2).

Daily regimes of microclimatic parameters in September 2016 measured at 5 cm height on exposed (NH) and shaded (LSB) flatroofs, respectively. Hourly readings have been used in constructing diagrams. a Temperature records are shown on linear scale whereas b relative humidity records on logarithmic scale

Daily temperature regimes of the standard measurement (2 m) and of microclimate dataloggers at 5 cm and 30 cm height in exposed and shaded flatroofs, respectively, on 12 September 2016
Seasonal pattern of differences coincided with those found in monthly dataset comparisons. Highly significant differences have been more frequent at warmer periods (9 and 10 ones in September and October, respectively) than in November or December (as low as 4 ones out of the maximally possible 15). Results for January have shown an intermediate position.
Biomass
Total cryptogamic biomass at the two selected exposed sites proved to be as high as 370 g/m2 (37 yrs) and 640 g/m2 (49 yrs), respectively. It comprised of 3.1 times (T = 57, p < 0.001) and 7.5 times (T = 65, p = 0.003) more lichens than bryophytes at the younger and older sites, respectively (Table 4). Cladonia rei and Ceratodon purpureus have been identified as community dominants. The community had a typically patchy appearance as lichens colonized on the surface of the firstly established bryophyte ‘islands’ scattered on barren areas of gravel (Fig. 3).

1 Patchy pattern of Cladonietum rei community, dominated by Ceratodon purpureus and C. rei. Diameter of the beer cap is 26 mm, 2 detail of a 37 years old site (Lehel street kindergarten), 3 detail of 49 years old site (big garage of Nursery Home). All photos had been taken on 16 October 2018
On the older roof, significantly more total cryptogamic biomass (about 1.7 times, T = 75.5, p = 0.028) as well as more bryophyte biomass (over 3.5 times, T = 64.5, p = 0.002) been sampled while about 1.5 times more lichen biomass has been recorded (ns) than on the younger one (Table 4).
Floristic composition
A total of 61 species (36 lichens and 25 bryophytes), mostly common and widespread throughout Hungary, was identified (Suppl. Table 8). Based on the morphological characters and the lichen substances detected by HPTLC (atranorin, barbatic acid, fumarprotocetraric acid, homosekikaic acid, norstictic acid, rangiformic acid, stictic acid, usnic acid) the presence of seven Cladonia taxa (C. coniocrea, C. conista, C. fimbriata, C. furcata, C. macilenta/floerkeana, C. rangiformis, C. rei), Stereocaulon tomentosum and Xanthoparmelia conspersa was confirmed.
Multivariate analysis has indicated a different composition of the more or less shaded roofs (1,2) from those of exposed ones (Fig. 4).

Species composition of the studied sites, according to the detrended correspondence analysis (DCA). Scientific names were abbreviated in 8 letters where the first four letters stand for the genus name while the last four ones for the species name, respectively. Bryophyte names are underlined. List of species used in DCA with their abbreviations and frequencies: lichens: Acarospora sp. (Acar sp.) 3, Caloplaca decipiens (Calo deci) 4, Caloplaca sp. (Calo sp.) 6, Caloplaca cf. citrina (Calo citr) 4, Caloplaca cf. pusilla (Calo pusi) 2, Candelariella aurella (Cand aure) 6, Cladonia coniocraea (Clad coni) 2, Cladonia fimbriata (Clad fimb) 4, Cladonia rei (Clad rei) 8, Lecanora dispersa (Leca disp) 6, Lecanora muralis (Leca mura) 7, Phaeophyscia nigricans (Phae nigr) 4, Phaeophyscia orbicularis (Phae orbi) 4, Physcia adscendens (Phys adsc) 2, Physcia caesia (Phys caes) 3, Sarcogyne regularis (Sarc regu) 3, Verrucaria nigrescens (Verr nigr) 6, Xanthoparmelia conspersa (Xant cons) 3, Xanthoria elegans (Xant eleg) 4, Xanthoria parietina (Xant pari) 2. bryophytes: Brachythecium rutabulum (Brac ruta) 3, Brachythecium salebrosum c.spg. (Brac sale) 2, Bryum argenteum (Bryu arge) 6, Ceratodon purpureus (Cera purp) 9, Grimmia pulvinata (Grim pulv) 5, Hedwigia ciliata (Hedw cili) 4, Hypnum cupressiforme (Hypn cupr) 2, Racomitrium canescens (Raco cane) 4, Syntrichia ruralis (Synt rura) 5
Abbreviations are resolved in detail in the Supplementary files. Letter size is proportional to species frequencies (small letters: present at 2 to 3 sites, medium letters: 4 to 6 sites, large letters: from 7 to 9 sites). Species recorded on a single site as well as a species-poor site have been excluded from the analysis.
Bryophytes characteristic for shaded roofs include Brachythecium rutabulum, B. salebrosum, Hypnum cupressiforme and Racomitrium canescens. Contrary, typical bryophytes of exposed roofs are Syntrichia ruralis, Bryum argenteum and Grimmia pulvinata. These are accompanied by a number of lichen species among which the most widespread is Lecanora muralis. Except for Stereocaulon tomentosum, we have not detected further lichen species confined to shaded sites. Frequent taxa present both on exposed and shaded roofs, included the bryophytes Ceratodon purpureus and Hedwigia ciliata as well as the lichens Cladonia fimbriata and C. rei.
Species richness
No significant correlation of bryophyte and lichen species richness was found between sites. Some sites proved to be more bryophyte-rich, some sites more lichen-rich while other ones were rich in both.
No correlation of total cryptogamic species richness (i.e. bryophytes plus lichens) and site area was revealed. However, when separated, saturation of species richness in relation to site area has been found both for lichens and bryophytes. For bryophytes species richness has already approached its maximum at a small area as low as 100–150 m2. In contrast, lichen species richness continued to increase over 500 m2 suggesting that saturation level is well above the area of studied sites (Fig. 5). Equation parameters for bryophytes are as a = 4.6306, b = 0.0394, pregression = 0.0538, r2 = 0.4336 whereas for lichens as a = 16.577, b = 0.0039, pregression = 0.0053, r2 = 0.6934.

Saturation curves of species richness of bryophytes and lichens, respectively, on different sized flatroofs, constructed by the equation f = a*(1-exp(− b*x)). Filled symbols and solid line: bryophytes, empty symbols and dashed line: lichens
Dicussion
First data on anthropogenic vegetation in Hungay date back several decades (Gallé 1965) but little information has been gathered since then and hardly any records stemmed from city build-ups. As it turned out in our study, vegetation of flatroofs, a widespread urban habitat recently, mostly consists of cryptogams. Vascular species only have scattered individuals of a handful species among bryophytes and lichens at most sites.
Most of the recorded roof-dwelling cryptogams are widespread and common in Hungary (Orbán and Vajda 1983; Verseghy 1994) but some of them are floristically interesting. These include the lichen, Stereocaulon tomentosum Fr. which was found as the second record for Hungary (Verseghy 1994; Matus et al. 2017, 2018). Most Stereocaulon species, originally from subalpine to alpine siliceous cliffs, heaths, are endangered and already regionally extinct in Central Europe (Wirth et al. 2013). Scattered occurrences of some Stereocaulon species have already been reported from Central and Western European anthropogenic habitats, including flatroofs (Wirth et al. 2013, https://www.verspreidingsatlas.nl; http://www.lichenology.info). Characteristic of montane siliceous rocks, Xanthoparmelia conspersa (Verseghy 1994), has scattered lowland records from Hungary (e.g. on andesite stones of an old bridge; Gallé 1965). We have detected it on three exposed flat roofs suggesting that in urban habitats it can be more frequent than believed.
Floristically interesting bryophytes include Ctenidium molluscum, Dicranum scoparium and Ptychostomum elegans characteristic of rock outcrops and of acidophilous forest soils, with scattered records from the Great Hungarian Plain. These three taxa have been confined to the only deep shade site. Two further bryophytes, Racomitrium canescens and Hedwigia ciliata are characteristic of siliceous rocky outcrops and acidic dry sandy grasslands and only have sparse records in the Hungarian Great Plain (Boros 1968; Erzberger et al. 2016).
Major identification challenges came from lichen genus Cladonia, of which seven taxa were found. The morphologically similar pair, Cladonia subulata and C. rei are chemically distinct species (Dolnik et al. 2010; Wirth et al. 2013). While C. rei produces homosekikaic acid (with ± sekikaic acid) and usually with fumarprotocetraric acid, C. subulata contains only fumarprotocetraric acid (Wirth et al. 2013). Only C. rei was found in the present study.
As our investigation revealed, not only the flora of the flat roofs, but also its vegetation is dominated by cryptogams, especially in terms of biomass. Planning the biomass sampling setup we only had a single Hungarian predecessor, studies from sandy grassland by Verseghy and Kovács–Láng (1971), Verseghy (1976, 1977). They sampled cryptogams at hectare sized areas in ten replicates of 20 cm × 20 cm sized plots (4000 cm2 in total). Although our biomass sampling setup applied only a fourth of the abovementioned area (1000 cm2 per site) we still it considered representative because of the identical sample size (n = 10) in a significantly smaller area (290 and 400 m2, respectively).
The largest portion of lichen biomass as well as of total biomass is coming again from the the morphologically diverse, species-rich genus Cladonia. Cladonia species are well known as important constituents of pioneer vegetation, both in natural and anthropogenic habitats (Rola and Osyczka 2014). They are considered as first colonizers in early stages of primary succession also at anthropogenic sites (Hajdúk and Lisická 1999; Osyczka and Rola 2013). Dominant of the studied roofs Cladonia rei proved to be a widespread species in various cryptogamic associations (Khodosovtsev et al. 2011; Rola et al. 2014). Vegetation of our exposed sites can be classified as the common assamblage Cladonietum rei (Günzl 2005; Rola et al. 2014) which could be recognized as characteristic of anthropogenic and disturbed habitats throughout Europe (Syrek and Kukwa 2008; Rola et al. 2014).
Contrastingly steep saturation curves suggest different minimal areas for bryophytes and lichens, respectively. Explaining this finding quantitatively would require specific data on rates of propagule rain and establishment. In absence of these, we can only outline certain assumptions. Firstly, dispersal of both groups rely on microscopic airborne propagules (spores and gemmae in bryophytes (Van Zanten and Pócs 1981), spores, picnidia, soredia and isidia in lichens (Wirth et al. 2013).
Spore dispersal has been studied with greatly contrasting spatial and temporal scales from local to transcontinental and from a vegetation season to geological timescale. The former, such as dispersal of pathogenic fungi spores on arables, show an exponentially or polynomially decreasing rate from source distance (Stockmarr et al. 2007). In the latter several specific factors (e.g. drought tolerance or colouration of spores) have been identified transcontinental bryophyte dispersal (van Zanten 1984; van Zanten and Pócs 1981).
Scale of our study is positioned in between the above two. Many of the lichen species found have their closest propagule sources at as much as 80–120 km from the studied sites (on hard rocks of Zemplén Mts, Bükk Mts, Mátra Mts and Bihar Mts, respectively). In contrast, all bryophyte species are known to have populations within a 10–30 km range (Erzberger et al. 2023, unpublished). This could explain why many lichens are likely more dispersal limited at the studied sites than bryophytes. While most bryophytes can reach even the smaller roofs, still new lichen species can be detected as roof sizes increase.
Secondly, establishment of lichens, as stable symbiotic organisms or tiny ecosystems (Farrar 1976), is more complex than just arrival of spores to a location. Being comprised of more than one organism (at least a mycobiont, a photobiont and some further microorganisms) (Hawksworth and Grube 2020), can reduce they establishment rate compared to that of the non-symbiotic bryophytes.
The reason for cryptogamic dominance on most flatroofs has become clear from microclimate measurements as only poikilohydric organisms can endure large fluctuations of climate elements. Microclimate has already been shown to have a profound influence on composition of various cryptogamic dominated communities (Kersaw and Larson 1974; Van Tooren et al. 1985; Aartsma et al. 2021) and the same applies to the studied flatroof assamblages. The most important biotic effects of microclimate are manifested through contrasting water availability. Marked differences of the exposed and shaded roofa are explained by reduced fluctuation and higher averages of RH as well as lower maxima and limited ranges of T of the latter (Suppl. Tables 1, 2, Fig. 1). This is accompanied by a reduced amount of light (not tested). At the deep shades site water stays available for prolonged periods compared to an exposed one. This effect has also been shown to change with the seasons (Tables 2, 3, Suppl. Table 5): it iss more significant in periods with longer daylight but fading away from late autumn. The consequence is a shift to a bryophyte dominated assamblage at the deep shade site, first in terms of species richness then possibly by biomass. This also stands for partially shaded sites (2,6,10, Suppl. Tables 3, 4, 6) to a smaller extent. A similar separation of bryophyte and lichen species due to differences in water and light availability has been demostrated in temperate forest habitats (Király and Ódor 2010; Király et al. 2013; Ódor et al. 2013).
Contrasting water availability may even lead to different successional pathways. In spite of its young age, the only deep shade roof harbours about four times as many bryophyte species as the average of the other nine roofs in an area about a fifth of the average of the others. This shaded pathway is already not exclusively characterized by cryptogams but also numerous fern species (Takács and Löki 2015) and some flowering vascular plants can also establish. The above microclimatic differences can explain the apparent separation of the deep shade site (1) from the rest detected by DCA and an intermediate position of partially shaded roofs (2, 6). Key species in this separation are the bryophytes Hypnum cupressiforme, Brachythecium rutabulum, B. salebrosum and Racomitrium canescens. The group of exposed roofs is, in contrast, distinguished by the bryophytes Bryum argenteum, Grimmia pulvinata and Syntrichia ruralis as well as numerous lichens taxa such as Caloplaca spp., Candelaria aurella, Lecanora dispersa, L. muralis, Phaeophyscia spp., Verrucaria spp., Xanthoria elegans.
There is a consensus that biomass accumulation during primary cryptogamic succession can be very slow lasting for decades (Farrar 1976; Verseghy 1977; Löbel et al. 2006). Although a full reconstruction of roof cryptogam successional pathways by the space-for-time approach (Picket 1989) still seems difficult. The reason for this is that we don’t have enough data from younger roofs (our records refer mostly to the 35 to 50 yrs age class) moreover the exact age of many roofs is unknown. We have, however, gained some insight through the biomass accumulation data of the similarly exposed 37 and 49 yrs old roofs, respectively. Our records have confirmed that there is still a statistically significant increase of bryophyte (and of total) biomass between these dates. Shifting proportions of biomass in dominants, Ceratodon purpureus and Cladonia rei, respectively, is remarkable. At the 37 yrs old site C. purpureus comprised only 8.5% of total biomass but its proportion has already been over 24% at the 49 yrs old one (Table 4). Both species are known as pioneers or first colonizers ( for C. rei see Hajdúk and Lisická 1999; Osyczka and Rola 2013; Wirth et al. 2013, https://www.verspreidingsatlas.nl/4188; for C. purpureus see Orbán 1984; https://www.verspreidingsatlas.nl/2642; https://www.britishbryologicalsociety.org.uk/) but it is unknown if there is any difference between their establishment dates. So this proportion shift may either suggest contrasting time of their colonization or differences of their growth rates.
Thus, already established thalli of C. rei may facilitate C. purpureus growth by draining fog and rain to their bases providing an extra water supply and some shelter for the bryophyte. As C. rei is also known to establish on bryophytes (Syrek and Kukwa 2008), it is also possible that those mats of C. purpureus which had C. rei thalli established atop could have performed an increased growth. Patchy appearance of Cladonietum rei (Fig. 3) also seem to support this assumption but further experiments are necessary to quantify this successional mechanism. Successional facilitation (Connell and Slatyer 1977) between two cryptogamic organisms has rarely been tested so far (Colesie et al. 2012).
A further increase of bryophyte biomass and withdrawal of C. rei during succession seems likely. This assuption is supported by a bryophyte dominated site (not sampled for biomass). Although this site (Waterworks Building, Botanical Garden) is not older but experiences some shade from surrounding trees. A somewhat longer availability of water may have accelerated the succession here resulting in an already non-patchy, uninterrupted bryophyte cover.
Conclusion for future biology
Flatroofs are ideal sites for studying diversity and undisturbed primary succession in cryptogamic communities due to their restricted accessibility. Old, non-renovated flatroofs are still widespread in all major cities in Hungary, but only a few of them have been surveyed. Consequently, most of their cryptogamic biodiversity remains unexplored. Roofing techniques, however, are being modernized and old roofs constantly renovated, mostly in framework of EU funded projects. The old-fashioned roof covers of bituminous felt with gravel coat, providing a vulnerable insulation (Varghese 2007), but also suitable habitats for cryptogamic communities, are going to be replaced. By 2020 four out of ten study sites, surveyed in 2016 have been, at least partly, renovated and one site demolished. Surveying on remaining old roofs should therefore have a priority in the next few years.
It is not realistic to declare any roof as a protected area as safety and cost effective insulation are the main objectives for urban buildings. Choosing among feasible techniques in construction of new buildings and in renovation of old ones can, however, promote flatroof biodiversity greatly. While modified bituminous membrane roofing (often reinforced with glass fibres) provide a rather uniform substrate with no microclimatic variability, the inverted roofing with gravel ballast can support relatively diverse microhabitats, especially if siliceous and calcareous gravels are applied alternately. This can provide a more strong and lasting insulation therefore stable cryptogam habitats (https://www.building.co.uk).
Using these techniques and to boost cryptogamic diversity also in roof gardens can add to the ornamental purpose. In addition to flowering plants (succulents such as Sedum, Sempervivum, Aeonium arboreum), larger bryophytes and macrolichens on green roofs can draw attention to these organisms, often neglected by the public. Up to now, only a few studies have included the use of cryptogams for roof greening (Drake et al. 2018). There are already attempts in modern urban architecture to couple cryptogams (Cladonia, Polytrichum) with flowering plants (Dvorak and Volder 2010) to establish stable, drought resistant, as well as aesthetic assemblages (Heim and Lundholm 2014; Lee et al. 2014). A practical outcome for these experiments is an improved hydrological function of roofs turned green (Bengtsson 2005).
References
Aartsma P, Asplund J, Odland A, Reinhardt S, Renssen H (2021) Microclimatic comparison of lichen heaths and shrubs: shrubification generates atmospheric heating but subsurface cooling during the growing season. Biogeosciences 18(5):1577–1599. https://doi.org/10.5194/bg-18-1577-2021
Aptroot A (2010) Changes in the epiphytic flora on four tilia trees in belgium over 59 years. Herzogia 25(1):39–45. https://doi.org/10.13158/heia.25.1.2010.39
Arup U, Ekman S, Lindblom L, Mattson J-E (1993) High performance thin layer chromatography (HPTLC), an improved technique for screening lichen substances. Lichenologist 25:61–71. https://doi.org/10.1006/lich.1993.1018
Behera SK, Mishra AK, Sahu N et al (2012) The study of microclimate in response to different plant community association in tropical moist deciduous forest from northern India. Biodivers Conserv 21:1159–1176. https://doi.org/10.1007/s10531-012-0230-5
Bengtsson L (2005) Peak flows from thin sedum-moss roof. Hydrol Res 36:269–280. https://doi.org/10.2166/nh.2005.0020
Berg C, Schwager P, Pöltl M, Dengler J (2016) Plot sizes used for phytosociological sampling of bryophyte and lichen micro-communities. Herzogia 29(2):654–667. https://doi.org/10.13158/heia.29.2.2016.654
Birks HJB, Heegaard E, Birks HH, Jonsgard B (1998) Quantifying bryophyte environment relationships. In: Bates JW, Ashton NW, Duckett JG (eds.) Bryology for the twenty-first century, maney and british bryological society. pp 305–319
Boros Á (1968) Bryogeographie und Bryoflora Ungarns. Akadémiai Kiadó, Budapest 466 pp
Braak CJF ter, Smilauer P (2002) CANOCO reference manual and CanoDraw for windows user's guide: software for canonical community ordination (version 4.5).
Bungartz F (1999) Die Flechten des Kölner Doms—Wiederbesiedelung oder Reliktvorkommen? Rheinische Heimatpflege 36:14–24
Chytrý M, Otýpková Z (2003) Plot sizes used for phytosociological sampling of European vegetation. J Veg Sci 14:563–570. https://doi.org/10.1111/j.1654-1103.2003.tb02183.x
Cilia C, Panigada C, Rossini M, Candiani G, Pepe M, Colombo R (2015) Mapping of asbestos cement roofs and their weathering status using hyperspectral aerial images. J Geo Inf 4(2):928–941. https://doi.org/10.3390/ijgi4020928
Colesie C, Scheu S, Green TGA, Weber B, Wirth R, Büdel B (2012) The advantage of growing on moss: facilitative effects on photosynthetic performance and growth in the cyanobacterial lichen Peltigera rufescens. Oecologia 169:599–607. https://doi.org/10.1007/s00442-011-2224-5
Connell JH, Slatyer RO (1977) Mechanisms of succession in natural communities and their role in community stability and organization. Am Nat 111(982):1119–1144
Dolnik C, Beck A, Zarabaska D (2010) Distinction of Cladonia rei and C. subulata based on molecular, chemical and morphological characteristics. Lichenologist 42:373–386. https://doi.org/10.1017/S0024282910000071
Drake P, Grimshaw-Surette H, Heim A, Lundholm J (2018) Mosses inhibit germination of vascular plants on an extensive green roof. Ecol Eng 117:111–114. https://doi.org/10.1016/j.ecoleng.2018.04.002
Dvorak B, Volder A (2010) Green roof vegetation for North American ecoregions: a literature review. Landsc Urban Plan 96:197–213. https://doi.org/10.1016/j.landurbplan.2010.04.009
Erzberger P, Schröder W (2013) The genus Bryum (Bryaceae, Musci) in Hungary. Stud Bot Hung 44:5–192
Erzberger P, Bednarek-Ochyra H, Ochyra R (2016) Grimmiaceae subfam. Racomitrioideae (Bryophyta) in Hungary. Pol Bot J 61(1):23–51
Erzberger P, Csiky J, Baráth K, Beránek Á, Deme J, Fintha G, Jakab G, Matus G, Mesterházy A, Nagy J, Nagy Z, Németh Cs, Pócs T, Schmidt D, Schmotzer A, Szűcs P, Wolf M, Papp B (2023) Preliminary data on frequency and distribution of Hungarian bryophytes based on ‘recent’ (1974-2022) records. J Bryol (in press)
Farkas E, Lőkös L (2003) Pyrenolichens of the Hungarian lichen flora II. Sarcopyrenia gibba (Nyl.) new to Hungary. Acta Bot Hung 45(3–4):273–278
Farkas E, Lőkös L, Molnár K (2001) Lichen mapping in Komárom. NW Hung Acta Bot Hung 43(1–2):147–162
Farrar JF (1976) The lichen as an ecosystem: observation and experiment. In: Brown DH, Hawksworth DL, Bailey RH (eds) Progress and problems. Academic Press, London, pp 385–406
Favero-Longo SE, Turci F, Tomatis M, Castelli D, Bonfante P, Hochella MF, Piervittori R, Fubini B (2005) Chrysotile asbestos is progressively converted into a non-fibrous amorphous material by the chelating action of lichen metabolites. J Environ Monit 7:764–766
Fudali E (2006) Influence of city on the floristical and ecological diversity of bryophytes in parks and cemeteries. Biodivers Res Conserv 1–2:131–137
Gallé L (1965) A Tisza menti kövesgátak zuzmócönózisai (Gesellschaften von auf Steinen wohnenden Flechten auf Überschwemmungsgebieten der Theiss). Móra Ferenc Múzeum Évkönyve 1964–65(1):265–286
Gazzano C, Favero-Longo SE, Matteucci E, Piervittori R (2009) Image analysis for measuring lichen colonization on and within stonework. Lichenologist 41(3):299–313. https://doi.org/10.1017/S0024282909008366
Gilbert O (1990) The lichen flora of Urban Wasteland. Lichenologist 22:87–101
Glime JM (2017) Construction. Chapt. 5. In: Glime, J. M. Bryophyte Ecology. Volume 2. Uses. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. Last updated 22 March 2023 and available at https://digitalcommons.mtu.edu/bryophyte-ecology5/
Goffinet B, Shaw AJ (eds) (2009). Cambridge University Press, Cambrige, p 565
Güler B, Jentsch A, Bartha S, Bloor JMG, Campetella G, Canullo R, Házi J, Kreyling J, Pottier J, Szabó G, Terziyska T, Uğurlu E, Wellstein C, Zimmermann Z, Dengler J (2016) How plot shape and dispersion affect plant species richness counts: implications for sampling design and rarefaction analyses. J Veg Sci 27:692–703. https://doi.org/10.1111/jvs.12411
Günzl B (2005) Erdflechtengesellschaften der Klasse Ceratodonto-Polytrichetea piliferi in Nordhessen—aktuelle Erfassung und Gliederung. Tuexenia 25:317–333
Hajdúk J, Lisická E (1999) Cladonia rei (lichenized Ascomycotina) at sites contaminated by emissions from Kovohút Krompachy (NE Slovakia). Bull Slovenskej Botanickej Spoločnosti, Bratislava 21:49–51
Hawksworth DL, Grube M (2020) Lichens redefined as complex ecosystems. New Phytol 227:1281–1283. https://doi.org/10.1111/nph.16630
Hedderson TAJ, Letts JB, Payne K (2003) Bryophyte diversity and community structure on thatched roofs of the Holnicote Estate, Somerset, U.K., Journal of Bryology 25(1):49–60. doi:https://doi.org/10.1179/037366803125002653
Heim A, Lundholm J (2014) Species interactions in green roof vegetation suggest complementary planting mixtures. Landsc Urban Plan 130:125–133. https://doi.org/10.1016/j.landurbplan.2014.07.007
Hodgetts NG, Söderström L, Blockeel TL, Caspari S, Ignatov MS, Konstantinova NA, Lockhart N, Papp B, Schröck C, Sim-Sim M, Bell D, Bell NE, Blom HH, Bruggeman-Nannenga MA, Brugués M, Enroth J, Flatberg KI, Garilleti R, Hedenäs L, Holyoak DT, Hugonnot V, Kariyawasam I, Köckinger H, Kučera J, Lara F, Porley RD (2020) An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J Bryol 42(1):1–116. https://doi.org/10.1080/03736687.2019.1694329
Humphrey JW, Davey S, Peace AJ, Ferris R, Harding K (2002) Lichens and bryophyte communities of planted and semi–natural forests in Britain: the influence of site type, stand structure and deadwood. Biol Conserv 107:165–180. https://doi.org/10.1016/S0006-3207(02)00057-5
Kershaw KA, Larson DW (1974) Studies on lichen-dominated systems. IX. Topographic influences on microclimate and species distribution. Can J Bot 52:1935–1945. https://doi.org/10.1139/b74-248
Khodosovtsev OY, Boiko MF, Nadyeina OV, Khodosovtseva YA (2011) Lichen and bryophyte associations on the lower Dnieper sand dunes: syntaxonomy and weathering indication. Chornomors’k bot 7:44–66. https://doi.org/10.14255/2308-9628/11.71/5
Király I, Ódor P (2010) The effect of stand structure and tree species composition on epiphytic bryophytes in mixed deciduous-coniferous forests of Western Hungary. Biol Conservation 143(9):2063–2069. https://doi.org/10.1016/j.biocon.2010.05.014
Király G, Balogh L, Barina Z, Bartha D, Bauer N, Bodonczi L, Dancza I, Farkas S, Galambos I, Gulyás G, Molnár VA, Nagy J, Pifkó D, Schmotzer A, Somlyai L, Szmorad F, Vidéki R, Vojtkó A, Sz Z (2003) A magyarországi flóratérképezés módszertani alapjai (Methodological fundaments of Hungarian floristic mapping). Flora Pannonica 1(1):3–20
Király I, Nascimbene J, Tinya F, Ódor P (2013) Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers Conserv 22(1):209–223. https://doi.org/10.1007/s10531-012-0415-y
Köehler M, Kaiser D (2021) Green roof enhancement on buildings of the University of Applied Sciences in Neubrandenburg (Germany) in times of climate change. Atmosphere 12(3):382. https://doi.org/10.3390/atmos12030382
Lee KE, Williams KJH, Sargent LD, Farrell C, Williams NS (2014) Living roof preference is influenced by plant characteristics and diversity. Landsc Urban Plan 122:152–159. https://doi.org/10.1016/j.landurbplan.2013.09.011
Lisická E (2008) Lichens on an acrylic-coated aluminium roof. Graphis Scripta 20:9–12
Löbel S, Dengler J, Hobohm C (2006) Species richness of vascular plants, bryophytes and lichens in dry grasslands: The effects of environment, landscape structure and competition. Folia Geobot 41:377–393. https://doi.org/10.1007/BF02806555
Lőkös L, Farkas E (1998) Az Őrség zuzmóflórájának alapvetése. Savaria A Vas Megyei Múzeumok Értesítője 24(2):127–160
Lőkös L, Farkas E (2000) Contributions to the knowledge of lichens of the forests along the Fekete-Körös, SE Hungary. Studia Botanica Hungarica 30–31:69–78
Mann HB (1947) Whitney DR (1947) On a test of whether one of two random variables is stochastically larger than the other. Ann Math Statist 18(1):50–60
Matus G, Freytag Cs, Adorján B, Oláh V, Mészáros I, Lőkös L (2017) Occurrence of Stereocaulon Hoffm. (Stereocaulaceae, Ascomycota) in Hungary (A Stereocaulon Hoffm. genus (Stereocaulaceae, Ascomycota) előfordulása Magyarországon). In: Activity of the Botanical Section of the Hungarian Biological Society, Journal of the Botanical Section of the Hungarian Biological Society 104(2):251–258.
Matus G, Freytag CS, Varga Z, Mészáros I, Adorján B, Oláh V, Szűcs P, Erzberger P, Balogh R, Lőkös L, Farkas E (2018) Caution, the botanist are working on the roof! (Vigyázat, a botanikusok a tetőn dolgoznak!) In: Activity of the Botanical Section of the Hungarian Biological Society. Journal of the Botanical Section of the Hungarian Biological Society 105(2):292.
Matwiejuk A (2009) Lichens of the cemeteries in Białystok (north-eastern Poland). Bot Stec 13:49–56
Morando M, Matteucci E, Nascimbene J, Borghi A, Piervittori R, Favero-Longo SE (2019) Effectiveness of aerobiological dispersal and microenvironmental requirements together influence spatial colonization patterns of lichen species on the stone cultural heritage. Sci Total Environ 685:1066–1074. https://doi.org/10.1016/j.scitotenv.2019.06.238
Nascimbene J, Dainese M, Sitzia T (2013) Contrasting responses of epiphytic and dead wood-dwelling lichen diversity to forest management abandonment in silver fir mature woodlands. For Ecol Manag 289:325–332. https://doi.org/10.1016/j.foreco.2012.10.052
Nash TH III (2008) Lichen Biology, 2nd edn. University Press, Cambridge, UK, pp 1–486
Niklfeld H (1971) Bericht über die Kartierung der Flora Mitteleuropa. Taxon 20(4):545–571. https://doi.org/10.2307/1218258
Ódor P, Király I, Tinya F (2013) Patterns and drivers of species composition of epiphytic bryophytes and lichens in managed temperate forests. For Ecol Manag 306:256–265. https://doi.org/10.1016/j.foreco.2013.07.001
Orange A, James PW, White FJ (2010) Microchemical methods for the identification of lichens. British Lichen Society, London, p 101
Orbán S (1984) A magyarországi mohák stratégiái és T. W. R. értékei (Strategy types and T, W, R indicator values of bryophytes in Hungary). Az Egri Ho Si Minh Tanárképző Főiskola Füzetei 17:755–765
Orbán S, Vajda L (1983) Magyarország mohaflórájának kézikönyve. Akadémiai Kiadó, Budapest, 518 pp. + XII pp.
Osyczka P, Rola K (2013) Cladonia lichens as the most effective and essential pioneers in strongly contaminated slag dumps. Cent Eur J Biol 8:876–887. https://doi.org/10.2478/s11535-013-0210-0
Pickett STA (1989) Space-for-time substitution as an alternative to long-term studies. In: Likens GE (eds) Long-term studies in ecology. Springer, New York, NY. https://doi.org/10.1007/978-1-4615-7358-6_5
Piervittori R, Salvadori O, Laccisaglia A (1994) Literature on lichens and biodeterioration of stonework. I Lichenologist 26(2):171–192. https://doi.org/10.1006/lich.1994.1014
Ranius T, Johansson P, Berg N, Niklasson M (2008) The influence of tree age and microhabitat quality on the occurrence of crustose lichens associated with old oaks. J Veg Sci 19:653–662. https://doi.org/10.3170/2008-8-18433
Rola K, Osyczka P (2014) Cryptogamic community structure as a bioindicator of soil condition along a pollution gradient. Environ Monit Assess 186(9):5897–5910. https://doi.org/10.1007/s10661-014-3827-1
Rola K, Osyczka P, Nobis M (2014) Cryptogamic communities dominated by the lichen Cladonia rei—a case study of polish post-smelting dumps in a worldwide context. Herzogia 27:121–135. https://doi.org/10.13158/heia.27.1.2014.121
Sabovljević M, Grdović S (2009) Bryophyte diversity within Urban Areas: case study of the City of Belgrade (Serbia). Int J Bot 5:85–92. https://doi.org/10.3923/ijb.2009.85.92
Skudnik M, Sabovljevic A, Batic F, Sabovljevic M (2013) The bryophyte diversity of Ljubljana (Slovenia). Pol Bot J 58(1):319–324. https://doi.org/10.2478/pbj-2013-0031
Skye E (1968) Lichens and air pollution: a study of cryptogamic epiphytes and environment in the Stockholm region. Acta Phytogeographica Suecica 52:123
Smith CW, Aptroot A, Coppins BJ, Fletcher A, Gilbert OL, James PW, Wolseley PA (eds) (2009). British Lichen Society, London, p 1046
Spearman C (1904) The proof and measurement of association between two things. Am J Psychol 15(1):72–101. https://doi.org/10.2307/1412159
Stockmarr A, Andreasen V, Østergård H (2007) Dispersal distances for airborne spores based on deposition rates and stochastic modeling. Phytopathology 97:1325–1330. https://doi.org/10.1094/PHYTO-97-10-1325
Studlar SM, Peck JE (2011) Extensive green roofs and mosses: reflections from a pilot study in terra alta. West Virginia Evansia 26(2):52–63. https://doi.org/10.1639/0747-9859-26.2.52
Syrek M, Kukwa M (2008) Taxonomy of the lichen Cladonia rei and its status in Poland. Biologia 63:493–497. https://doi.org/10.2478/s11756-008-0092-1
Szűcs P, Lóth H (2008) Almásfüzitő antropogén élőhelyeinek mohaflórája. Komárom-Esztergom Megyei Múzeumok Közleményei 13–14:399–424
Takács A, Löki V (2015) Néhány Adat Debrecen Urbán-Flórájához. Kitaibelia 20(1):168–170
Thuring C, Dunnett N (2019) Persistence, loss and gain: characterising mature green roof vegetation by functional composition. Landsc Urban Plan 185:228–236. https://doi.org/10.1016/j.landurbplan.2018.10.026
Van Tooren BF, During HJ, Lensink MJG (1985) The influence of the bryophyte layer on the microclimate in chalk grasslands. Abstr Bot 9:219–230
van Zanten BO (1984) Some consideration on the feasibility of long-distance transport in bryophytes. Acta Boot Neerl 33:231–232
van Zanten BO, Pócs T (1981) Distribution and dispersal of bryophytes. Adv Bryol 1:479–562
Varghese PC (2007) Building construction. Prentice-Hall of India 427. pp
Verseghy K (1976) Quantitative investigation of xerothermophilous lichens of sandy soil. Ann Hist Nat Mus Natn Hung 68:59–63
Verseghy K (1977) Investigation on lichen production of grassland communities of sandysoil. II. Acta Biol Acad Sci Hung 28(1):25–32
Verseghy K, Kovács-Láng E (1971) Investigations on production of grassland communities of sandy soil in the IBP area near Csévharaszt (Hungary). I. Production of lichens. Acta Biol Acad Sci Hung 22(4):393–411
Verseghy K (1994) Magyarország zuzmóflórájának kézikönyve. Magyar Természettudományi Múzeum, Budapest, p 414
Wirth V, Hauck M, Schultz M (2013) Die flechten deutschlands. Ulmer, Stuttgart, p 1244
Zar JH (2010) Biostatistical analysis. 5th Edition, Prentice-Hall/Pearson, Upper Saddle River, xiii, p 944
Zsólyom D, Szűcs P (2018) Balaton település (Heves megye) mohaflórája. (The bryophyte flora of Balaton village (Heves county, Hungary)). Botanikai Közlemények 105(2):231–242. https://doi.org/10.17716/BotKozlem.2018.105.2.231
Acknowledgements
We are thankful for. G. Bene (Division of Labour Safety, Debrecen University) for contacting us with facility managers of the buildings of the University. Facility managers as well as heads of the institutions have all been supporting our research, we are thankful for the cooperation of I. H. Bordás (4th Nurse Hostel, Debrecen University), L. Papp (Botanic Garden, Debrecen University), M. Sz. Erdelics (Lehel street kindergarten), J. K. Kucskár (Home for Visually Impaired), as well as B. Kmetty, L. Nagy, K. K. Krajnik (Nursery home, city of Debrecen). Help of L. Laczkó, N. Nagy, Z. Varga, students of MSc course Lichenology, in sampling is thankfully acknowledged. Records of standard meteorological observations have been provided by the Observatory of Department of Meteorology, University of Debrecen and the Renewable Energy Park.
Funding
Open access funding provided by University of Debrecen. Financial support has been provided by NKFIH under K_17/124341 (2017–2023). The publication is supported by the EFOP-3.6.1-16-2016-00022 project. The project is co-financed by the European Union and the European Social Fund. Supported by the ÚNKP-19-3-I-DE-392 New National Excellence Program of the Ministry for Innovation and Technology to R. Aszalósné Balogh.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Aszalósné Balogh, R., Matus, G., Lőkös, L. et al. Cryptogamic communities on flatroofs in the city of Debrecen (East Hungary). BIOLOGIA FUTURA 74, 183–197 (2023). https://doi.org/10.1007/s42977-023-00166-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42977-023-00166-3