
Abstract
Understanding how active and healthy ageing can be achieved is one of the most relevant global problems. In this review, I use the “Four questions” framework of Tinbergen to investigate how ageing works, how it might contribute to the survival of species, how it develops during the lifetime of (human) individuals and how it evolved. The focus of ageing research is usually on losses, although trajectories in later life show heterogeneity and many individuals experience healthy ageing. In humans, mild changes in cognition might be a typical part of ageing, but deficits are a sign of pathology. The ageing of the world's populations, and relatedly, the growing number of pathologically ageing people, is one of the major global problems. Animal models can help to understand the intrinsic and extrinsic factors contributing to ageing.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
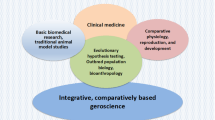
Biological ageing is an age-related decline in physiological function, leading to decreased survival and reproductive rate. Biological ageing affects almost all organisms, although in significantly different forms. Phylogenetically, it is an ancient process (Bonduriansky et al. 2008), already present in the single-cell bacteria Escherichia coli (Nyström, 2007). In humans, population ageing is the most important medical and social demographic problem worldwide, according to the World Health Organization (2020). This problem is relatively new. In 70 years, especially in the most developed countries, the proportion of people over 65 has risen from 11 to 18%, and by 2050, it could be as high as 38%. Scientific research is a crucial element to maximise the number of people who achieve healthy trajectories of ageing. The “four questions” framework (Tinbergen 1963) helps to obtain a complete understanding of the costs, the potential benefits and constraints that might have shaped ageing throughout the evolution of species. Below I summarise some hypotheses and results on the mechanism of ageing, how it contributes to the survival of species, how it develops during the lifetime of (human) individuals and how it evolved (Fig. 1).

Applying Tinbergen’s four questions (1963) to understand ageing
Causation: What is the cause of ageing?
Hallmarks of ageing form three main groups: (1) damage to cellular functions: genomic instability, telomere attrition, epigenetic alterations, and loss of proteostasis; (2) antagonistic responses: deregulated nutrient sensing, altered mitochondrial function, and cellular senescence; (3) integrative hallmarks: stem cell exhaustion and altered intercellular communication (López-Otín et al. 2013; Aunan et al. 2016; Sándor and Kubinyi 2019). Stem cells avoid apoptosis, i.e. they are immortal (Dunham et al. 2000). Less abundant stem cells mean that an organism’s renewal capacity is limited. Differentiated cells of more complex animals age and die, eventually leading to the animal’s death. Cancer cells that express a telomere-lengthening enzyme are also immortal (Dunham et al. 2000). In a way, cancer is related to embryogenesis, just like stem cells. The main mammalian tumour-suppressor mechanisms evolved from ancient mechanisms that regulate embryogenesis/developmental maintenance but now contribute to ageing (i.e. inducing cell death (Campisi 2003).
The conserved role of signalling pathways is well-established. For example, activation of the nutrient-sensing TOR (Target of Rapamycin) drives growth, and when growth is completed, TOR enhances the ageing process (Blagosklonny 2010). This is probably the reason, on a cellular level, why women outlive men. Men are larger and stronger. Hyperactive mTOR contributes to the physical robustness of young males at the cost of accelerated ageing (Blagosklonny 2010).
During ageing, cells cease to divide, and the number of senescent cells in tissues rises, impairing renewal and homeostasis as well as decreasing organ function. Old cells are usually characterised by bigger size, more diverse morphotypes, increased lysosomal beta-galactosidase activity, more chromosomes (i.e. three or more in humans instead of two), shortened telomeres (non-coding regions at the tips of chromosomes), and changes in several genes’ expression levels (Rodier and Campisi 2011). Biological mechanisms ultimately contribute to the clinical effects of ageing, as seen in organ decline and therefore reduced function. In humans, the mechanisms behind the decline of brain functions include a decrease in grey matter volume (after the age of 20 years), especially in the frontal cortex. White matter volume also decreases, and its function declines (Harada et al. 2013). The rate of such physiological changes can often be accounted for by certain genetic variants. For example, the beta-2-adrenergic receptor (ADRB2) gene was shown to affect white matter integrity and cognitive ability in old age (Penke et al. 2010).
Survival value: Is ageing an adaptive process?
“If organisms can function well in youth, why can they not continue to do so in old age?” – asks Partridge and Barton in their influential review (Partridge and Barton 1993). Several theories propose answers (Milewski 2010). The key conceptual insight of all theories is that the old count less than the young: with age, even immortal organisms have impaired fertility and die from random injuries and disease. By the time genes supporting later life would take effect, most of the carriers are already dead or infertile and harmful mutations already have been passed on to the offspring of the individuals bearing them. Therefore, natural selection becomes less efficient in old age, i.e. there could be only a weak selection against ageing.
According to the antagonistic pleiotropy hypothesis (Williams 1957; Williams and Day 2003), a gene with a benefit in early life but a detrimental effect later has a net positive impact and will be favoured by natural selection. Many findings support antagonistic pleiotropy. For example, a moderate level of radical oxidative species (ROS) is beneficial at a young age, but a high level of ROS is detrimental at old age (Dai et al. 2017). In humans, cardiovascular diseases and diabetes are examples of antagonistic pleiotropy, meaning that specific gene variants contribute to female reproductive advantages but later increase the risk of diseases (Byars & Voskarides 2020).
Therefore, the ageing process can be a genetically programmed, adaptive trait because it prevents overcrowding, accelerates generations’ turnover, and may even favour closely related individuals (Longo et al. 2005). For example, the popular grandmother-hypothesis claims that menopause evolved in social species because old females with a long post-fertility lifespan (PFSL) increase their inclusive fitness by investing in their grand offspring. Indeed, in addition to humans, in killer whales (Orcinus orca), the presence of post-fertile grandmothers increases the reproductive success of daughters (Brent et al. 2015). Note, however, that young, fertile Asian elephant grandmothers also increase the survival of grand calves and decrease their daughters’ inter-birth intervals in contrast to the hypothesis prediction (Lahdenperä et al. 2016). The positive effect of grandmothers is irrespective of their reproductive status. Therefore, menopause, when the oocyte number falls below the threshold required for ovarian function, is most probably an age-related decline and is not beneficial.
Another influential hypothesis, the disposable soma theory, proposes that organisms adjust their investments into either maintenance or reproduction to maximise fitness (Kirkwood 1977; Abrams and Ludwig 1995). Ageing emerges when an organism allocates resources from anti-ageing repairs to other needs. Thus, ageing is due to an energy-saving strategy, i.e. an evolutionary trade-off between gains during early life (growth, reproduction) and maintenance. The rate of ageing depends on the allocation of resources.
The competing mutation accumulation theory rejects that ageing could be adaptive. Instead, it assumes that ageing is purely maladaptive due to the build-up of random deleterious mutations that are only expressed beyond a certain age. Natural selection cannot eliminate these deleterious genes because organisms usually die due to unavoidable environmental risks before reaching the age these genes would be expressed (see above). Therefore, ageing-related genes can accumulate over successive generations, even in potentially immortal populations, because selection does not oppose the spread of deleterious mutations in the relative lack of old individuals.
The hyperfunction theory of ageing assumes that ageing is a continuation of development driven by, for example, the inappropriate activation of growth-promoting pathways (Blagosklonny 2013). Specifically, signalling pathways that are essential in early life (such as the insulin and TOR pathways), also drive cellular senescence. However, this obvious link does not mean that ageing is programmed. It is just the aimless continuation of the programme. From an evolutionary viewpoint, the death of an individual with the “best fitting” genome in a certain environment cannot be considered adaptive. If adaptation is regarded as a phenotypic trait with a functional role in each individual, we find that the ageing process does not support the survival of an individual, on the contrary. Thus, programmed theories are based on circular reasoning: they claim that ageing eliminates the less valuable old animals for the benefit of the species and groups. However, old animals are less valuable precisely because of ageing. The most plausible explanation and the only way out of the circle is that ageing has no function; it is simply the by-product of development (Kowald and Kirkwood 2016).
Ontogeny: The process of becoming older
The focus of ageing research is usually on losses. However, not every old individual shows a deficit in functioning. Rowe and Kahn were the first to popularise the heterogeneity of health trajectories in later life (Rowe and Kahn 1987). They differentiated between “usual” or typical ageing, i.e. functioning well but with a high risk for disease and disability, and “successful” or healthy ageing, i.e. demonstrating a high level of functioning across several domains. Since then, most definitions have included physiological constructs (e.g. physical functioning), engagement constructs (e.g. participation in volunteer work), and well-being constructs (e.g. life satisfaction). Based on the definitions, the prevalence of healthy agers highly varies from less than 1% up to 90% in different publications (Cosco et al. 2014). For example, in a longitudinal study in Taiwan, based on multidimensional indicators on a representative sample, 29.1% of the population belonged to the successful ageing group, 36.3% to the usual ageing group, 23.2% to the health declining group and 11.4% demanded care (Hsu and Jones 2012). In the USA, community-living persons aged at least 70 years were followed for 9 years. 31.7% showed no decline in the Mini-Mental State Examination scores, 43.5% minimal decline, 14.9% moderate decline, 7.4% progressive decline, and 2.5% rapid decline (Han et al. 2016).
Cognitive health is an inherent part of healthy ageing in humans, characterised by social activity and independent life until death. In healthy, educated humans, the age-related cognitive decline begins in the mid-twenties (Salthouse 2009). Human cognitive abilities can be divided into several domains, and they may change differently with age. For example, vocabulary and general knowledge (‘crystallised abilities’) remain stable or gradually improve up to 60–70 years of age. In contrast, attentional abilities, memory functions, performance on verbal tasks, problem-solving, processing and learning new information, and attending to the environment (‘fluid abilities’) peak in the third decade of life and then decline steadily. Mild changes in cognition might be a typical part of ageing, but deficits are a sign of pathology, which impairs daily functioning and results in an inability to lead an independent life. In addition, a reduced schedule of brain activity due to behavioural change and/or loss due to ageing of brain function can create a self-reinforcing downward spiral in impaired brain function in older adults.
Evolution: How does ageing change across species?
The rate and onset of ageing show large variation between species (Nussey et al. 2013). Mortality may increase, is constant or decrease with age, linked to energy allocation characteristic of a species, but little is known about what constraints favour a life trajectory (Baudisch and Vaupel 2012).
In some species, the mortality rate from ageing is stable, i.e. older individuals have no higher chance to die than young ones; in other words, the species is “immortal”. A jellyfish species (Turritopsis dohrnii) and a planarian flatworm (Schmidtea mediterranea), are well-known examples. Throughout animal evolution, there is a gradual decline in the abundance of cardinal stem cells for regeneration. Higher bilaterians, including humans, “opted for” greater complexity but less abundant stem cells and consequently lost immortality (Petralia et al. 2014).
Among species with a pattern of ageing with increasing mortality and decreasing fertility, lifespan is generally inversely correlated with metabolic rates. If ageing is due to a gradual accumulation of damage from metabolic by-products, species with slower metabolisms and higher weight live longer. However, there are several exceptions. Birds outlive mammals of comparable size, which clearly shows that organisms vary in the extent to which they combat the proximate causes of ageing. Mice and rats have a 2–3 years lifespan, but the naked mole rat (Heterocephalus glaber) lives up to 30 years due to several factors such as enhanced antioxidant defence, lower insulin levels, and fewer aberrant proteins (Kim et al. 2011). The evolutionary theory of ageing predicts delayed ageing in species with reduced vulnerability to environmental hazards. For example, flying may have a protective effect against predators, explaining why birds and bats live considerably longer than expected based on their metabolic rate (Austad and Fischer 1991).
Model organisms of ageing
Human lifespan is expected to increase by 4.4 years by 2040, exceeding 85 years for both sexes in some countries (Foreman et al. 2018) due to better nutrition, health care, sanitation, and economic well-being. The growing number of older people with age-related diseases poses an increased burden on health care and pension systems. Therefore, ageing research focuses on increasing the health span, i.e. the length of time the person is healthy. Animal models can help to understand the intrinsic and extrinsic factors contributing to ageing.
By examining animal species with extreme longevity or immortality, such as the Hydra genus (Martínez and Bridge 2012), the bivalve (Arctica islandica, > 500 years, (Ungvari et al. 2013), and the bowhead whale (Balaena mysticetus, > 200 years, (Keane et al. 2015)), researchers might uncover the genetic elements behind a long (and healthy) life. However, these animals are usually challenging to study in the laboratory. Short-lived organisms are more efficient experimental models. Nematode worms (Caenorhabditis elegans), fruit fly (Drosophila melanogaster), mice (Mus musculus), turquoise killifish (Nothobranchius furzeri), and the unicellular yeast (Saccharomyces cerevisiae), with a lifespan from a few days to a few years, have shed light on many regulatory mechanisms behind ageing (e.g. Fabrizio et al. 2001; Juhász et al. 2003; Vellai et al. 2003; Kovács-Valasek et al. 2017; Hu and Brunet 2018). Based on these studies, it was found that genes related to ageing are components of essential metabolic and signalling pathways, such as autophagic activity and cellular metabolism (Sándor and Kubinyi 2019). Although these laboratory model animals help to uncover evolutionarily conserved mechanisms, they do not necessarily reflect the variance found in natural populations, nor the interaction between the cellular mechanisms of ageing and complex extrinsic factors. Therefore, even valid findings in model laboratory animals might not directly correspond to genetic polymorphisms linked to health- and lifespan in humans living in a very different environment compared to laboratories. Moreover, some genes linked to the central nervous system development are unique to the primate lineage or can be found only in humans (Bitar and Barry 2017).
Where many animal models fail: cognitive ageing
Cognitive ageing refers to an age-related decline in cognitive functioning experienced by almost all older people in several cognitive abilities such as memory, processing speed, and conceptual reasoning. In humans, together with age-related neurodegenerative diseases, non-pathological variation in cognitive ageing has also been well documented in the scientific literature (Cosco et al. 2014). Although normal cognitive ageing can influence the day-to-day life of the elderly, the most common health issues posing a great burden on the healthcare systems of developed countries are related to pathological cognitive ageing. Over 85 years of age, 25–45% of individuals have dementia, a severe cognitive decline (Bird 2008). Identifying the genetic and environmental factors that influence the development of impaired cognition among the elderly has been a major quest for gerontology research. As it seems, the genetic background of age-related cognitive decline, even that of specific neurodegenerative states, is very complex, and further research is needed to reveal the interactions between genetic variants, with each having subtle effects.
However, the most traditional animal models, rodents and non-mammalian animals do not develop age-related neurodegenerative disorders by nature. Although this limitation of worms, flies, mice and other organisms has been overcome by different techniques used to induce neurodegenerative processes in the central nervous system, the findings of such studies may not be easily implemented in humans. The causes of neurodegeneration can be many and are influenced by cognitive and environmental factors that cannot be assessed in laboratory animals because of their limited cognitive and social capacities. Dementia can result from multiple small strokes in the brain, certain diseases (AIDS, Huntington’s disease), and Alzheimer’s disease (AD). AD is characterised by amyloid beta and tau protein pathology inside the brain (Khan and Bloom 2016). Although transgenic mouse models have amyloid beta plaque formation in the brain, they naturally have resistance to amyloid beta pathology and therefore do not show an extensive neuronal loss. Notably, the brain of rodents is lissencephalic, i.e. lacking surface convolutions. Unsurprisingly, the findings from AD people were not consistent with those in transgenic AD mouse models (Ambrosini et al. 2019). This may be one of the reasons why the 40-year search for effective therapeutics to treat AD has resulted in a clinical failure rate of 100% (Mullane and Williams 2019). Dogs can provide a solution to this problem because individuals with more severe symptoms of cognitive dysfunction have higher levels of amyloid beta in their brains (Urfer et al. 2020).
As mentioned above, humans are also unique with their highly extended post-fertility lifespan. Some researchers argue that the high prevalence of Alzheimer’s disease (AD) is linked to PFLS (Gunn-Moore et al. 2018). The mean PFLS is 45, while the maximum lifespan is 110 years. Thus, PFLS as a percentage of maximum lifespan is 40.9%. Organisms with a similarly long post-fertility lifespan are at a high risk of AD because of links with longevity and the malfunction of the insulin signalling pathway. The killer whale has a similarly long PFLS (48.7%) to humans. Although orcas with dementia have not been reported, studies found evidence of both amyloid deposits and tau pathology in related cetaceans and three species of dolphins (Gunn-Moore et al. 2018). The PFLS of chimpanzees (Pan troglodytes) is 18.8, the domestic dog (beagle breed, Canis familiaris) 23.3, and the domestic cat (Felis silvestris catus) is 38.0. Thus, humans, sea mammals, cats and maybe some dog breeds with especially long PFLS might have AD-like neurodegeneration (Gunn-Moore et al. 2018). However, there are serious ethical reasons against keeping these animals in large numbers for research purposes and especially against invasive experimental interventions. Besides, characterising longevity phenotypes would be almost as challenging in long-living primates and orcas as in humans. The limitation that traditional laboratory animals do not develop dementia has been overcome by transgenic and gene-edited animals. However, the long history of failed AD trials challenges the validity of these models (Götz et al. 2018). Consequently, there are still many unanswered questions about the biology of cognitive ageing which cannot be addressed adequately in current laboratory animal models.
According to several authors (e.g. Ambrosini et al. 2019; Chapagain et al. 2018; Creevy et al. 2022; Zakošek Pipan et al. 2021), dogs provide a good model for human cognitive ageing with unprecedented advantages in terms of general validity. Companion or pet dogs share our environment more than any other species. The same environmental factors affect them as people (chemicals, air pollution, noise pollution, lack of exercise, etc.), which are suspected risk factors of cognitive decline in humans but cannot be appropriately modelled in the laboratory. Individuals, like humans, vary in life expectancy and have multiple diseases treated in an advanced healthcare system. They allow for longitudinal and interventional studies because they age 4–11 times faster than humans. Pet dogs also excel in displaying socio-cognitive skills in interaction with humans compared to other animals (Miklósi and Kubinyi 2016) and consequently develop homologues of age-related human diseases, such as dementia-like canine cognitive dysfunction (Madari et al. 2015) and functional declines (Szabó et al. 2018). Canine research can help us to identify suitable biomarkers, for example, allowing early detection and predictions of AD in humans; study the interplay of genetic and environmental effects and the unique ageing trajectories, which can be the basis of appropriate interventions. Recent research in our group at ELTE supports these assumptions. Since the establishment of the Canine Brain and Tissue Bank (CBTB) in 2017 (Sándor et al. 2021a), we obtained brains and other tissue samples from 171 donated pet dogs representing various breeds. Based on these samples, it was found that a conserved ageing biomarker, CDKN2A, had similar tissue-specific age-related gene expression patterns to that of humans (Sándor et al. 2021b); the levels of amyloid beta peptide 42 (Abeta42) in three brain regions positively correlated with age and cognitive decline (Urfer et al. 2021); and the list of 3,436 differentially expressed genes detected by poly-A RNA sequencing from the frontal cortex of young and old dogs corresponded with human data to a greater extent than with rodent data (Sándor et al. 2022). Moreover, in a pilot study, a negative association was reported between the memory performance of old dogs and the levels of Actinobacteria in the gut, mimicking observations in people with AD (Kubinyi et al. 2020). These results support that the dog model of human cognitive (dys)function reflects the phenotypes of human age-related neurodegenerations such as dementia.
Conclusion for future biology
Both the number and proportion of older people are expected to increase at an accelerating rate in the future. Therefore, the study of ageing and longevity is of great importance. It enables us to better understand the ageing process, which allows the promotion of a healthy lifestyle among the general population and provides humankind to successfully cope with the long-term socioeconomic consequences of an ageing population. Most research on human ageing looks at older adults, many of whom have chronic illnesses, and we know little about the ageing of healthy young people. However, to prevent age-related diseases and declines, interventions to slow ageing and prolong healthspan should be applied to people while they are young and healthy.
Ageing is a conservative phenomenon. Therefore, findings on simple model organisms are generally translatable to humans. However, features related to the unique human cognition have no counterparts in these organisms. Animals prone to developing human-like neurodegenerative disorders and age-related cognitive abnormalities, such as dogs, have grown to be a precious model species for researchers who aim to understand the biological background of ageing. Research should focus on the mechanisms and phenotype of ageing in these animals. Data from companion dogs representing a broader spectrum of genetic and environmental variability would be most beneficial for modelling human populations. However, obtaining biological samples, such as muscle or brain tissue, is a more significant challenge for molecular and histological research than for laboratory animals; therefore, large-scale medical databases and biobanking services must be established and maintained.
References
Abrams PA, Ludwig D (1995) Optimality theory, Gompertz’ law, and the disposable soma theory of senescence. Evolution (n y) 49:1055–1066. https://doi.org/10.1111/j.1558-5646.1995.tb04433.x
Ambrosini YM, Borcherding D, Kanthasamy A, Kim HJ, Willette AA, Jergens A et al (2019) The gut-brain axis in neurodegenerative diseases and relevance of the canine model: a review. Front Aging Neurosci 11:130. https://doi.org/10.3389/fnagi.2019.00130
Aunan JR, Watson MM, Hagland HR, Søreide K (2016) Molecular and biological hallmarks of ageing. Br J Surg 103:e29–e46. https://doi.org/10.1002/bjs.10053
Austad SN, Fischer KE (1991) Mammalian aging, metabolism, and ecology: evidence from the bats and marsupials. J Gerontol 46:B47–B53. https://doi.org/10.1093/geronj/46.2.B47
Baudisch A, Vaupel JW (2012) Evolution. Getting to the root of aging. Science 338:618–9. https://doi.org/10.1126/science.1226467
Bird TD (2008) Genetic aspects of Alzheimer disease. Genet Med 10:231–239. https://doi.org/10.1097/GIM.0b013e31816b64dc
Bitar MA, Barry G (2017) Multiple innovations in genetic and epigenetic mechanisms cooperate to underpin human brain evolution. Mol Biol Evol 35:263–268. https://doi.org/10.1093/molbev/msx303
Blagosklonny MV (2010) Why the disposable soma theory cannot explain why women live longer and why we age. Aging 2:884–887. https://doi.org/10.18632/aging.100253
Blagosklonny MV (2013) Aging is not programmed: Genetic pseudo-program is a shadow of developmental growth. Cell Cycle 12:3736. https://doi.org/10.4161/CC.27188
Bonduriansky R, Maklakov A, Zajitschek F, Brooks R (2008) Sexual selection, sexual conflict and the evolution of ageing and life span. Funct Ecol 22:443–453. https://doi.org/10.1111/j.1365-2435.2008.01417.x
Brent LJN, Franks DW, Foster EA, Balcomb KC, Cant MA, Croft DP (2015) Ecological knowledge, leadership, and the evolution of menopause in killer whales. Curr Biol 25:746–750. https://doi.org/10.1016/J.CUB.2015.01.037
Byars SG, Voskarides K (2020) Antagonistic pleiotropy in human disease. J Mol Evol 88:12–25. https://doi.org/10.1007/S00239-019-09923-2/TABLES/2
Campisi J (2003) Cancer and ageing: rival demons? Nat Rev Cancer 3:339–349. https://doi.org/10.1038/nrc1073
Chapagain D, Range F, Huber L, Virányi Z (2018) Cognitive Aging in Dogs. Gerontology 64:165–171. https://doi.org/10.1159/000481621
Cosco TD, Prina AM, Perales J, Stephan BCM, Brayne C (2014) Operational definitions of successful aging: a systematic review. Int Psychogeriatr 26:373–381. https://doi.org/10.1017/S1041610213002287
Creevy KE, Akey JM, Kaeberlein M, Promislow DEL, Barnett BG, Benton B et al (2022) An open science study of ageing in companion dogs. Nature 602:51–57. https://doi.org/10.1038/s41586-021-04282-9
Dai DF, Chiao YA, Martin GM, Marcinek DJ, Basisty N, Quarles EK et al (2017) Mitochondrial-targeted catalase: extended longevity and the roles in various disease models. Prog Mol Biol Transl Sci 146:203–241. https://doi.org/10.1016/BS.PMBTS.2016.12.015
Dunham MA, Neumann AA, Fasching CL, Reddel RR (2000) Telomere maintenance by recombination in human cells. Nat Genet 26:447–450. https://doi.org/10.1038/82586
Fabrizio P, Pozza F, Pletcher SD, Gendron CM, Longo VD (2001) Regulation of longevity and stress resistance by Sch9 in yeast. Science 1979(292):288–290. https://doi.org/10.1126/science.282.5390.943
Foreman KJ, Marquez N, Dolgert A, Fukutaki K, Fullman N, McGaughey M et al (2018) Forecasting life expectancy, years of life lost, and all-cause and cause-specific mortality for 250 causes of death: reference and alternative scenarios for 2016–40 for 195 countries and territories. Lancet 392:2052–2090. https://doi.org/10.1016/S0140-6736(18)31694-5
Götz J, Bodea L-G, Goedert M (2018) Rodent models for Alzheimer disease. Nat Rev Neurosci 19:583–598. https://doi.org/10.1038/s41583-018-0054-8
Gunn-Moore D, Kaidanovich-Beilin O, Gallego Iradi MC, Gunn-Moore F, Lovestone S (2018) Alzheimer’s disease in humans and other animals: a consequence of postreproductive life span and longevity rather than aging. Alzheimer’s Dementia 14:195–204. https://doi.org/10.1016/J.JALZ.2017.08.014
Han L, Gill TM, Jones BL, Allore HG (2016) Cognitive aging trajectories and burdens of disability, hospitalization and nursing home admission among community-living older persons. J Gerontol Series A 71:766–771. https://doi.org/10.1093/GERONA/GLV159
Harada CN, Natelson Love MC, Triebel KL (2013) Normal cognitive aging. Clin Geriatr Med 29:737–752. https://doi.org/10.1016/j.cger.2013.07.002
Hsu HC, Jones BL (2012) Multiple trajectories of successful aging of older and younger cohorts. Gerontologist 52:843–856. https://doi.org/10.1093/GERONT/GNS005
Hu C-K, Brunet A (2018) The African turquoise killifish: A research organism to study vertebrate aging and diapause. Aging Cell. https://doi.org/10.1111/acel.12757
Juhász G, Csikós G, Sinka R, Erdélyi M, Sass M (2003) The Drosophila homolog of Aut1 is essential for autophagy and development. FEBS Lett 543:154–158. https://doi.org/10.1016/S0014-5793(03)00431-9
Keane M, Semeiks J, Webb AE, Li YI, Quesada V, Craig T et al (2015) Insights into the evolution of longevity from the bowhead whale genome. Cell Rep 10:112–122. https://doi.org/10.1016/J.CELREP.2014.12.008
Khan SS, Bloom GS (2016) Tau: the center of a signaling nexus in alzheimer’s disease. Front Neurosci 10:31. https://doi.org/10.3389/fnins.2016.00031
Kim EB, Fang X, Fushan AA, Huang Z, Lobanov AV, Han L et al (2011) Genome sequencing reveals insights into physiology and longevity of the naked mole rat. Nature 479:223–227. https://doi.org/10.1038/nature10533
Kirkwood TBL (1977) Evolution of ageing. Nature 270:301–304. https://doi.org/10.1038/270301a0
Kovács-Valasek A, Szabadfi K, Dénes V, Szalontai B, Tamás A, Kiss P et al (2017) Accelerated retinal aging in PACAP knock-out mice. Neuroscience 348:1–10. https://doi.org/10.1016/J.NEUROSCIENCE.2017.02.003
Kowald A, Kirkwood TBL (2016) Can aging be programmed? A critical literature review. Aging Cell 15:986–998. https://doi.org/10.1111/acel.12510
Kubinyi E, Bel Rhali S, Sándor S, Szabó A, Felföldi T (2020) Gut Microbiome composition is associated with age and memory performance in pet dogs. Animals 10:1488. https://doi.org/10.3390/ani10091488
Lahdenperä M, Mar KU, Lummaa V (2016) Nearby grandmother enhances calf survival and reproduction in Asian elephants. Sci Rep 6:27213. https://doi.org/10.1038/srep27213
Longo VD, Mitteldorf J, Skulachev VP (2005) Programmed and altruistic ageing. Nat Rev Genet 6:866–872. https://doi.org/10.1038/nrg1706
López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G (2013) The hallmarks of aging. Cell 153:1194–1217. https://doi.org/10.1016/J.CELL.2013.05.039
Madari A, Farbakova J, Katina S, Smolek T, Novak P, Weissova T et al (2015) Assessment of severity and progression of canine cognitive dysfunction syndrome using the CAnine DEmentia Scale (CADES). Appl Anim Behav Sci 171:138–145. https://doi.org/10.1016/j.applanim.2015.08.034
Martínez DE, Bridge D (2012) Hydra, the everlasting embryo, confronts aging. Int J Dev Biol 56:479–487. https://doi.org/10.1387/ijdb.113461dm
Miklósi Á, Kubinyi E (2016) Current trends in canine problem-solving and cognition. Curr Dir Psychol Sci 25:300–306. https://doi.org/10.1177/0963721416666061
Milewski LAK (2010) The evolution of ageing. Biosci Horizons 3:77–84. https://doi.org/10.1093/biohorizons/hzq001
Mullane K, Williams M (2019) Preclinical models of alzheimer’s disease: relevance and translational validity. Curr Protoc Pharmacol 84:e57. https://doi.org/10.1002/CPPH.57
Nussey DH, Froy H, Lemaitre J-F, Gaillard J-M, Austad SN (2013) Senescence in natural populations of animals: Widespread evidence and its implications for bio-gerontology. Ageing Res Rev 12:214–225. https://doi.org/10.1016/J.ARR.2012.07.004
Nyström T (2007) A Bacterial Kind of Aging. PLoS Genet 3:e224. https://doi.org/10.1371/journal.pgen.0030224
Partridge L, Barton NH (1993) Optimally, mutation and the evolution of ageing. Nature 362:305–311. https://doi.org/10.1038/362305a0
Penke L, Muñoz Maniega S, Houlihan LM, Murray C, Gow AJ, Clayden JD et al (2010) White matter integrity in the splenium of the corpus callosum is related to successful cognitive aging and partly mediates the protective effect of an ancestral polymorphism in ADRB2. Behav Genet 40:146–156. https://doi.org/10.1007/s10519-009-9318-4
Petralia RS, Mattson MP, Yao PJ (2014) Aging and longevity in the simplest animals and the quest for immortality. Ageing Res Rev 16:66–82. https://doi.org/10.1016/J.ARR.2014.05.003
Rodier F, Campisi J (2011) Four faces of cellular senescence. J Cell Biol 192:547–556. https://doi.org/10.1083/JCB.201009094
Rowe J, Kahn R (1987) Human aging: usual and successful. Science 237:143–149. https://doi.org/10.1126/science.3299702
Rudnicka E, Napierała P, Podfigurna A, Męczekalski B, Smolarczyk R, Grymowicz M (2020) The World Health Organization (WHO) approach to healthy ageing. Maturitas 139:6–11. https://doi.org/10.1016/J.MATURITAS.2020.05.018
Salthouse TA (2009) When does age-related cognitive decline begin? Neurobiol Aging 30:507–514. https://doi.org/10.1016/J.NEUROBIOLAGING.2008.09.023
Sándor S, Czeibert K, Salamon A, Kubinyi E (2021a) Man’s best friend in life and death: scientific perspectives and challenges of dog brain banking. Geroscience 43:1653–1668. https://doi.org/10.1007/S11357-021-00373-7/FIGURES/1
Sándor S, Jónás D, Tátrai K, Czeibert K, Kubinyi E (2022) Poly(A) RNA sequencing reveals age-related differences in the prefrontal cortex of dogs. Geroscience. https://doi.org/10.1007/S11357-022-00533-3/FIGURES/8
Sándor S, Kubinyi E (2019) Genetic pathways of aging and their relevance in the dog as a natural model of human aging. Front Genet. https://doi.org/10.3389/fgene.2019.00948
Sándor S, Tátrai K, Czeibert K, Egyed B, Kubinyi E (2021b) CDKN2A gene expression as a potential aging biomarker in dogs. Front Veter Sci 8:348. https://doi.org/10.3389/FVETS.2021.660435
Szabó D, Miklósi Á, Kubinyi E (2018) Owner reported sensory impairments affect behavioural signs associated with cognitive decline in dogs. Behav Proc 157:354–360. https://doi.org/10.1016/j.beproc.2018.07.013
Tinbergen N (1963) On aims and methods of Ethology. Z Tierpsychol 20:410–433. https://doi.org/10.1111/j.1439-0310.1963.tb01161.x
Ungvari Z, Sosnowska D, Mason JB, Gruber H, Lee SW, Schwartz TS et al (2013) Resistance to genotoxic stresses in arctica islandica, the longest living noncolonial animal: Is extreme longevity associated with a multistress resistance phenotype? J Gerontol A Biol Sci Med Sci 68:521–529. https://doi.org/10.1093/gerona/gls193
Urfer S, Darvas M, Keene D, Czeibert K, Kubinyi E, Sándor S et al (2020) Amyloid beta-42 levels in companion dog brains correlate with age and cognitive function. Innov Aging 4:887–887. https://doi.org/10.1093/GERONI/IGAA057.3273
Urfer SR, Darvas M, Czeibert K, Sándor S, Promislow DEL, Creevy KE et al (2021) Canine Cognitive Dysfunction (CCD) scores correlate with amyloid beta 42 levels in dog brain tissue. GeroScience 2021:1–8. https://doi.org/10.1007/S11357-021-00422-1
Vellai T, Takacs-Vellai K, Zhang Y, Kovacs AL, Orosz L, Müller F et al (2003) Influence of TOR kinase on lifespan in C elegans. Nature 426:620–621
Williams GC (1957) Pleiotropy, natural selection, and the evolution of senescence. Evolution 11:398–411. https://doi.org/10.2307/2406060
Williams PD, Day T (2003) Antagonistic pleiotropy, mortality source interactions, and the evolutionary theory of senescence. Evolution 57:1478–1488. https://doi.org/10.1111/J.0014-3820.2003.TB00356.X
Zakošek Pipan M, Prpar Mihevc S, Štrbenc M, Košak U, German Ilić I, Trontelj J et al (2021) Treatment of canine cognitive dysfunction with novel butyrylcholinesterase inhibitor. Sci Rep 2021(11):1–11. https://doi.org/10.1038/s41598-021-97404-2
Acknowledgements
This project has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (Grant Agreement No. 680040) and Hungarian Academy of Sciences via a grant to the MTA-ELTE ‘Lendület/Momentum’ Companion Animal Research Group (grant no. PH1404/21).
Funding
Open access funding provided by Eötvös Loránd University.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kubinyi, E. Biologia Futura: four questions about ageing and the future of relevant animal models. BIOLOGIA FUTURA 73, 385–391 (2022). https://doi.org/10.1007/s42977-022-00135-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42977-022-00135-2