
Abstract
Mountainous grassland landscapes are severely threatened by the proliferation of shrub encroachment. So far, it remains unclear though how slope aspect coupled with land disturbances linked to the proximity of homesteads to communal grazing sites affects the distribution, density and structure of the encroaching shrubs in these grassland ecosystems. In this study, we investigated the role that slope aspect plays in determining the density and structure of an encroaching shrub species Euryops floribundus N.E. Br in three communal grazing sites located at varying distances from rural homesteads in a semi-arid mountainous grassland in the Eastern Cape Province of South Africa. Three sites were strategically chosen in Cala communal grazing lands in an encroached mountainous grassland landscape that depicted north and south-facing slope aspects. The selected sites were Tsengiwe; a site located in close proximity to homesteads at a distance of less than 100 m, Upper Mnxe situated at an intermediate distance ranging from 200 to 800 m away from homesteads and Manzimdaka, which was located furthest from homesteads at a distance greater than 1600 m. In each site and corresponding slope aspect, the density and structure of the shrub E. floribundus were evaluated on 36 randomly distributed plots, yielding a total of 108 plots across all sites. We found that shrub density was significantly higher (50%) on the north-facing slope compared to the south-facing slope in Tsengiwe, the site located near homesteads. Shrub height was significantly higher (53 and 17%) on the north-facing slope compared to the south-facing slope at Upper Mnxe and Manzimdaka, which were located at intermediate and furthest distances from the homesteads. Notably, shrub height was significantly lower (37%) in the north-facing slope compared to the south-facing slope in Tsengiwe. Following a similar pattern to shrub height, total stem number was significantly higher (20 and 85%) in the north-facing slope compared to the south-facing slope at Upper Mnxe and Tsengiwe, respectively. Shrub crown area was higher (33 and 11%) in the north-facing slope compared to the south-facing slope at Upper Mnxe and Manzimdaka, respectively. A strong positive relationship was established between the height of shrubs and their longest crown diameter, shortest crown diameter and shrub crown area across all sites, suggesting that the investigated shrub species E. floribundus employs its structural characteristics to survive, and thrive and this was more evident in the north-facing slope. The results highlight the importance of considering the proximity of homesteads into account in ecological studies, and puts emphasis on improved understanding of the vegetation patterns shaped by shrub encroachment in mountainous grasslands, which is crucial in the development of effective land management strategies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Grasslands are distributed throughout varying topographies globally and makeup about 40% of the surface of the world (Bardgett et al., 2021; Lieffering et al., 2019). This proportion of grasslands worldwide also include grasslands found in mountainous environments. Mountains are distinct and conspicuous parts of the landscape, characterized by extreme topography and complex slopes with steep gradients (Hirmas and Graham, 2011). Specifically, mountainous grasslands are characterized by high elevations and undulating topography where the mixture of grasses and other herbaceous plants predominate (O’Connor, 2005; Sala et al., 2013; Ralph Clark et al., 2021). These grasslands provide a range of material and non-material benefits to human livelihoods (Bardgett et al., 2021). The benefits brought by grasslands include a wide range of ecosystem services including food production, timber, water supply and regulation, protection from natural hazards, storage of carbon, climate change mitigation, recreation and tourism (Bardgett et al., 2021; Gibson, 2009). Mountainous grasslands also provide abundant forage for more than 360 million cattle and 600 million sheep and goats due to their high protein and mineral content (Huntsinger and Hopkinson, 1996; Squires et al., 2018). In addition, these grasslands provide economic activities and livelihood opportunities for many rural people who depend on livestock sales and products (e.g., meat, milk and skins) for their livelihoods (Palmer and Ainslie, 2005; Papanastasis et al., 2017; Squires et al., 2018). Despite the importance of mountainous grasslands, mounting evidence indicates that they are under severe threat from ongoing shrub encroachment (Masibonge et al., 2019; Montané et al., 2007; Steger et al., 2022).
Mountainous grasslands are seriously threatened by the increasing density of native trees and shrubs, a phenomenon referred to as shrub encroachment (Van Auken, 2009). This land cover transformation, which is common in many arid and semi-arid grasslands, leads to a reduction in biodiversity, changes in plant species composition and contributes to overall decrease in productivity of grasslands (Mogashoa et al., 2021; Moleele et al., 2002; Roques et al., 2001; Van Auken, 2000). These changes associated with shrub encroachment negatively impact livestock production and the livelihoods of those living in rural areas (Archer et al., 2017; Scholes and Archer, 1997). The impact of shrub encroachment on grassland ecosystems is not only governed by the density of shrubs, but is also controlled by the structural characteristics of the encroaching plants (Eldridge et al., 2011, Osborne et al., 2018; Zhou et al., 2019; Mogashoa et al., 2021). The structural characteristics of the encroaching plant including the height, stem number and canopy density are critical in governing the accessibility of crucial resources like sunlight, water, nutrients and space (Zizka et al., 2014; Götmark et al., 2016; Gaillard et al., 2018). These structural characteristics significantly influence the ability of various grass species to dominate and thrive within the ecosystem. For instance, taller shrubs with denser canopies may create more shade, reducing light availability for grass species and increasing competition for water and nutrients (Osborne et al., 2018; Mogashoa et al., 2021; Ding and Eldridge, 2022). Despite the negative impact of shrub encroachment on some of the functions and services provided by mountainous grasslands, it has been documented that slope aspect promotes shrub encroachment (Kakembo et al., 2007; Gartzia et al., 2014; Archer et al., 2017; Wu et al., 2018). This land characteristic is a major driver of incoming solar radiation, and creates a range of microclimates that affect the growth and survival of woody and shrubby vegetation (Auslander et al., 2003; Bennie et al., 2008; Carletti et al., 2009; Gallardo-Cruz et al., 2009; Grab, 2013; Longman et al., 2014). Generally, north-facing slopes in the Southern Hemisphere experience warmer temperatures than their south-facing counterparts due to their exposure to increased solar radiation, resulting in drier and warmer conditions (Holland and Steyn, 1975; Schulze and McGee, 1978). This is in stark contrast to the conditions in the Northern Hemisphere, where south-facing slopes receive more solar radiation, making them warmer compared to the cooler north-facing slopes (Kutiel and Lavee, 1999; Jasińska et al., 2019; Li et al., 2022). In this article, we will use the terms “warmer aspect” and “cooler aspect” where appropriate to ensure clarity and account for the different slope aspect terminology used in the Southern and Northern hemispheres. Despite the impact of slope aspect in determining microclimatic conditions, its effect on the distribution of encroaching shrubs is still not well understood.
There is no explicit agreement on how slope aspect influences the structure of encroaching woody plants, as evidenced by the contradictory results in the existing literature. Some studies have shown that cooler slope aspects are characterized by greater density and cover of encroaching woody plants than warmer slope aspects (Johnson and Miller, 2006; Kakembo et al., 2007; Weisberg et al., 2007; Wu et al., 2018; Soubry et al., 2022), while other studies have reported greater density and cover of shrubs on warmer slope aspects (Hottman and O’Connor, 1999; Johnson and Miller, 2006; Gartzia et al., 2014; Caviezel et al., 2017). There are also studies that found no discernible effect of slope aspect on the encroaching woody plants (Coop and Givnish, 2007; Endress et al., 2007; Haile et al., 2021). The inconsistent results observed in these studies likely result from (1) the interactive effects of slope aspect with other factors such as human-induced disturbances, particularly in mountainous grazing sites situated in close proximity to homesteads; and (2) the type of encroaching woody plant species such as trees, shrubs, or a mixture of both. Additionally, there is a lack of research investigating the influence of slope aspect on the structural characteristics of encroaching shrub species in mountainous grasslands.
In this study, we investigated the influence of slope aspect on the density and structural characteristics of Euryops floribundus N.E. Br shrub in communal grazing sites located at varying distances from homesteads in a semi-arid mountainous grassland. Euryops floribundus N.E. Br is one of the shrub species that is aggressively encroaching the mountainous grasslands in the Eastern Cape Province of South Africa. The mountainous grasslands run north–south producing predominantly north-aspect and south-aspect slopes. This difference in aspect affects the growth patterns of the Euryops floribundus species. The species is an evergreen multi-stemmed perennial shrub that develops to reach a height of 3 m. This shrub species is endemic to the Eastern Cape Province of South Africa, and produces abundant quantities of tiny seeds that are easily dispersed by wind, animals and water (Masibonge et al., 2019). Euryops floribundus shrub is widely found in communal grazing lands, but it is not palatable to livestock such as goats, sheep and cattle. Local communities utilize the shrub for its medicinal purposes and as a source of fuel for cooking and building material (Vetter et al., 2006; Shackleton and Gambiza, 2008; Gxasheka, 2013). The extent of utilization of the benefits derived from the woody plants and the impact of livestock grazing in communal grasslands vary depending on the proximity of homesteads to grazing lands (Shackleton, 1993; Inman et al., 2020). This difference in the distance from homesteads to grazing lands has been associated to the extent of shrub encroachment, as grazing land near homesteads tend to have higher shrub encroachment compared to those that are located at far distances from homesteads (Fisher et al., 2011; Bennett et al., 2012). The sites located near homesteads often experience more land disturbance, such as overgrazing and wood harvesting for building and firewood, compared to sites situated further away (Fisher et al., 2011; Gatica et al., 2020). Consequently, nearby sites are characterized by bare soil, which can provide favorable conditions for the establishment and growth of shrubs. Even so, there is a lack of studies examining the underlying ecological and topographical drivers behind the encroachment of Euryops floribundus. We hypothesized that Euryops floribundus density would be higher on the north-facing slope than on the south-facing slope due to warmer and drier conditions favoring shrub establishment, but lower on communal grazing sites closest to homesteads than those farthest away due to increased land disturbance. Additionally, we hypothesized that the structural characteristics of Euryops floribundus including height, total stem number and cover would be higher on the north-facing slope than on the south-facing slope due to warmer and drier conditions favoring shrub development, but lower on communal grazing sites closest to homesteads than those farthest away due to greater land disturbance. Improved understanding of how slope aspect influences the density and structure of encroaching shrubs across different communal grazing sites will help in formulating efficient rangeland management strategies to curb shrub encroachment, in turn enhancing the quality and quantity of forage and promoting sustainable livestock production in communal grazing land.
Material and methods
Description of study sites
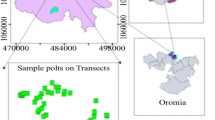
The study was conducted across three communal grazing sites near Cala in the Sakhisizwe Local Municipality in the Eastern Cape Province of South Africa (Fig. 1). Cala is a town located near the Tsomo River, approximately 28 km southwest of Elliott town. The name “Cala” means “adjacent to,” referring to its location near the west of the Drakensberg (Raper et al., 2014). The Sakhisizwe Local Municipality has a population of approximately 64,500 people, of which 14,500 live in Cala (Aliber and Xabadiya, 2020; Sakhisiwe Local Municipality, 2017). To select our communal grazing sites, we used a multi-faceted approach that involved discussions with local extension officers and community leaders as well as extensive field scouting. Our selection was based on the distance of neighboring homesteads to the communal grazing site. The distance to homesteads was estimated using Google Earth (Haile et al., 2021). The selected sites were Tsengiwe (situated near at less than 100 m from homesteads), Upper Mnxe (located at an intermediate distance from homesteads within 200–800 m) and Manzimdaka (located far from homesteads at more than 1600 m) (Table 1). These sites are approximately 10 km apart within the communal grazing land of Cala. The climate at the study site is semi-arid, with an average annual precipitation of 500 mm and average temperatures of 22 °C in January and 12 °C in July. Most of the precipitation in the area occurs during the summer season (November to March) and only small amounts of precipitation fall during the winter months (June to August). For all the selected sites, the woody layer is mainly dominated by Euryops floribundus while the grass layer is dominated by Aristida congesta, Elionurus muticus, Eragrostis chloromelas, Eragrostis plana, Heteropogon contortus, Sporobolus africanus and Themeda triandra (Mucina and Rutherford 2006; Table 1). Mudstones of the Karroo supergroup dominates the geology of the sites, giving rise to shallow and stony soils (Mucina and Rutherford 2006). The communal grazing sites investigated in this study had no fencing or imposed management practices, such as prescribed burning. Over the last three decades, the sites have been continuously used as grazing land for a variety of livestock including cattle, sheep and goats, with no limits or constraints imposed on land management (Gxasheka, 2013). There is a lack of quantified data on stocking rates in the study sites, but the average number of cattle per household ranges from 5 (Upper Mnxe), 6 (Manzimdaka) and 7 (Tsengiwe), while the average number of sheep per household is 10 (Manzimdaka), 13 (Upper Mnxe) and 20 (Tsengiwe) (Gxasheka et al., 2017). As shown in Fig. 2, these sites are subjected to various land disturbances such as overgrazing, unplanned burning of grazing areas, stem cutting and the harvesting and collection of Euryops floribundus for firewood. Among these disturbances, overgrazing and firewood collection of Euryops floribundus are the most significant, with Tsengiwe experiencing higher levels of firewood extraction compared to Upper Mnxe and Manzimdaka (Gxasheka, 2013; Gxasheka et al., 2017).

Location of the three communal grazing sites, a Tsengiwe, b Upper Mnxe and c Manzimdaka) within Cala in Sakhisizwe Local Municipality of the Eastern Cape Province of South Africa. Photo credits: Masibonge Gxasheka

Various observed land disturbance including a Overgrazing, b stem cutting, resulted in no surviving stems, c stem cutting with only one stem remaining, d stem cutting, with only one stem removed, e livestock grazing, f burned shrub with no stems remaining, g burned shrub with some stems remaining, h firewood collection in the three communal grazing sites of Cala, Eastern Cape Province of South Africa. Photo credits: Masibonge Gxasheka
Experimental design and vegetation sampling
In February 2022, a mountainous grassland landscape encroached by Euryops floribundus shrub species, with north-facing and south-facing slope aspects was selected at each communal grazing site after an intensive field vegetation survey. (Table 2, Fig. 2). For each mountainous grassland site, slope aspect was determined using a compass (Majeed et al., 2022). From the bottom of the mountainous grassland to the top, 18 plots (10 m × 10 m) were randomly established approximately 10 m apart at each slope aspect. Overall, this approach resulted in 36 plots per study site and 108 plots across the three sites investigated. To determine density and total stem number, we measured all the shrub live individuals of Euryops floribundus within the demarcated 100 m2 plot (Dlamini et al., 2019; Mogashoa et al., 2021). We measured the height of every shrub using a calibrated 2 m rod from the base of the shrub Euryops floribundus to the top of the green foliage within the 100 m2 plot. We classified the shrub heights into three categories following the work of Masibonge et al. (2019): 0–1 m for seedlings, > 1–2 m for young shrubs and > 2 m for mature shrubs. To estimate the shrub crown area, we determined the longest crown diameter (LCD) and shortest crown diameter (SCD) of each shrub Euryops floribundus using tape measure in 100 m2 plot. Then, we calculated the shrub crown area (SCA) using the equation shown below:
where d1 is the major axis (LCD) and d2 is the minor axis (SCD) of the crown (Hesselbarth et al., 2018). Furthermore, we used a Garmin® GPSMAP 64 × series handheld global positioning system ( Garmin International Incorporated Company in Kansas, USA) to record longitude, latitude and elevation of each plot per sampling site.
Data analysis
The data on density, height class distribution and structural characteristics of Euryops floribundus were analyzed using the R statistical software version 4.2.1 (R Core Team, 2022). Prior to analysis, the data were checked for normal distribution using the Shapiro–Wilk test, and homogeneity of variance using Levene’s test, which indicated non-normal distribution. Shrub density data were normally distributed after they were log 10 transformed. A two-way ANOVA, employing the general linear model (GLM) procedure (Pekár and Brabec, 2016) was used to determine how slope aspect and site influence shrub density and structural characteristics of Euryops floribundus (i.e., shrub height, total stem number, longest crown diameter, shortest crown diameter and shrub crown area). The results were considered significant at p ≤ 0.05. Subsequently, a post hoc analysis was conducted using the Tukey test. To get an overview on how the structural characteristics are related, we conducted a Pearson’s correlation analysis using the R package “corrplot” (Wei et al., 2017). The correlation coefficients were classified as follows: 0.00–0.10, negligible correlation, 0.10–0.39: weak, 0.40–0.69: moderate, 0.70–0.89: strong correlation, 0.90–1.00: very strong (Schober et al., 2018). A p value ≤ 0.05 was considered statistically significant. For data visualization and manipulation, we used the R packages “ggplot2”, “dplyr “and “ggpubr” (Wickham and Grolemund, 2016; Wickham and Wickham, 2020).
Results
Effect of slope aspect on shrub density of Euryops floribundus
Slope aspect and communal grazing site as individual factors did not influence shrub density, but the combined effect of slope aspect and site significantly influenced the density of Euryops floribundus (p < 0.05; Table 3). Specifically, shrub density was significantly higher (50%) on the north-facing slope (2528 shrubs ha−1) compared to the south-facing slope (1689 shrubs ha−1) (p < 0.05; Fig. 3a) in Tsengiwe. In contrast, no significant differences were observed for shrub density in the north-facing and south-facing slopes at Upper Mnxe (1800 and 2156 shrubs ha−1) and Manzimdaka (1572 and 2017 shrubs ha−1) (p > 0.05; Fig. 3a). In Upper Mnxe and Manzimdaka, seedling density (0–1 m) was lower on the north-facing slope relative to the south-facing slope (Fig. 5a). On the contrary, seedling density (0–1 m) was higher on the north-facing slope compared to the south-facing slope in Tsengiwe. The density of young shrubs (> 1–2 m) did not differ between the north-facing slope and the south-facing slope at Upper Mnxe and Manzimdaka (5b). In contrast, the density of young shrubs (> 1–2 m) was lower on the north-facing slope compared to south-facing slope in Tsengiwe (5b). Compared to the south-facing slope, the density of mature shrubs (> 2 m) was higher on the north-facing slope at Upper Mnxe (5c). Similarly, mature shrub density was higher on the north-facing slope than the south-facing slope (5c) in Manzimdaka. Conversely, mature shrubs were not found on the north-facing slope, but only observed on the south-facing slope in Tsengiwe (5c).

Boxplots values of a shrub density, b shrub height and c total number of stems per shrub between north-facing and south-facing slope aspect in Tsengiwe, Upper Mnxe and Manzimdaka. Different letters denote significant differences at p < 0.05 between the slope aspects. Box plot elements: box = values of 25th and 75th percentiles; horizontal line = median; symbol + = mean, whiskers = values of 5th and 95th percentiles. Circle and triangle shapes outside the whiskers represent the outliers
Effect of slope aspect on shrub height and total stem number
Shrub height and total stem number of Euryops floribundus showed significant differences at the different communal grazing sites and varying slope aspects (p < 0.05; Table 3). Notably, shrub height was significantly higher (53%) in the north-facing slope (1.48 m) compared to the south-facing slope (0.97 m) at Upper Mnxe (p < 0.05; Fig. 3b). Similarly, the shrub height on the north-facing slope (1.6 m) was significantly higher (17%) compared to south-facing slope (1.3 m) at Manzimdaka (Fig. 3b). Shrub height at Tsengiwe followed an opposite trend, as it was 37% lower in the north-facing slope (0.88 m) compared to the south-facing slope (1.4 m) (Fig. 3b). Total stem number was significantly higher (20%) on the north-facing slope (4 stems/shrub) compared to the south-facing slope (3 stems/shrub) in Upper Mnxe (Fig. 3c). Similarly, total stem number was significantly higher (85%) on the north-facing slope (7 stems/shrub) compared to the south-facing slope (4 stems/shrub) at Tsengiwe (Fig. 3c). In contrast, the total stem number of the north-facing slope (5 stems/shrub) and south-facing slope (4 stems/shrub) did not differ statistically at Manzimdaka (Fig. 3c).
Effect of slope aspect on crown characteristics of Euryops floribundus shrub
Communal grazing site significantly influenced the short crown diameter, longest crown diameter and shrub crown area, while slope aspect had no direct influence on the crown diameters and crown cover of Euryops floribundus. The combination of slope aspect and communal grazing site, however, significantly influenced the short crown diameter, longest crown diameter and shrub crown area of Euryops floribundus. Specifically, the longest crown diameter was significantly higher (20%) on the north-facing slope (1.14 m) than on the south-facing slope (0.95 m) at Upper Mnxe (p < 0.05; Fig. 4a). However, the longest crown diameter did not show a statistically significant difference between the north-facing slope (1.46 m) and the south-facing slope (1.39 m) in Manzimdaka. In contrast, the longest crown diameter was significantly lower (22%) on the north-facing slope (1.14 m) compared to south-facing slope (1.46 m) (Fig. 4a) in Tsengiwe. In Upper Mnxe, shortest crown diameter was significantly higher (12%) on the north-facing slope (0.80 m) than on the south-facing slope (0.71 m) (Fig. 4b). In Manzimdaka, the shortest crown diameter did not differ significantly between the north-facing slope (1.06 m) and the south-facing slope (1.05 m). Conversely, shortest crown diameter was significantly lower (17%) on the north-facing slope (0.87 m) compared to south-facing slope (1.05 m) at Tsengiwe (Fig. 4b). Shrub crown area was significantly higher (33%) on the north-facing slope (0.86 m2) than the south-facing slope (0.64 m2) in Upper Mnxe (p < 0.05; Fig. 4c). Even though there was no statistical difference between the slope aspects, the shrub crown area showed a marginal increase (11%) on the north-facing slope (1.44 m2) compared to the south-facing slope (1.30 m2) at Manzimdaka. Conversely, shrub crown area was significantly lower (38%) on the north-facing slope (0.85 m2) compared to south-facing slope (1.36 m2) in Tsengiwe (Fig. 4c).

Box plots showing the distribution of a longest crown diameter, b shortest crown diameter and c shrub crown area in north-facing and south-facing slope aspect in Tsengiwe, Upper Mnxe and Manzimdaka. Different letters denote significant differences at p < 0.05 between the slope aspects. Box plot elements: box = values of 25th and 75th percentiles; horizontal line = median; symbol + = mean, whiskers = values of 5th and 95th percentiles. Circle and triangle shapes outside the whiskers represent the outliers
Relationship among structural characteristics of Euryops floribundus in north-facing and south-facing slopes
In Fig. 6, a heat map diagram shows the Pearson correlation coefficients between shrub structural characteristics of Euryops floribundus on the north-facing and south-facing slopes for the three communal grazing sites investigated in this study. In the north-facing slope in Tsengiwe, we established a moderate positive correlation between shrub height and longest crown diameter (r = 0.55) and shrub crown area (r = 0.49) (Fig. 6a), indicating that taller shrubs tend to have larger and wider crowns. Total stem number was moderately positively correlated with shrub crown area (r = 0.53), shortest crown diameter (r = 0.51) and longest crown diameter (r = 0.51), implying that shrubs with more stems tend to have larger crowns. Shrub crown area was very strongly positively correlated with shortest crown diameter (r = 0.96) and longest crown diameter (r = 0.93). At Tsengiwe, shrub height was moderately positively correlated with shrub crown area (r = 0.69), longest crown diameter (r = 0.67) and shortest crown diameter (r = 0.64) in the south-facing slope (Fig. 6a). Total stem number was moderately positively correlated with shrub crown area (r = 0.59), shortest crown diameter (r = 0.58) and longest crown diameter (r = 0.53). Shrub crown area was very strongly positively correlated with shortest crown diameter (r = 0.94; p < 0.001) and longest crown diameter (r = 0.94). These relations suggest that taller shrubs with more stems tend to have larger and dense crowns on the south-facing slope.
In the north-facing slope at Upper Mnxe, shrub height was moderately positively correlated with longest crown diameter (r = 0.44) (Fig. 6b), suggesting that taller shrubs tend to have larger crowns. Shrub crown area (r = 0.58), shortest crown diameter (r = 0.56) and longest crown diameter (r = 0.51) were all moderately positively correlated with total stem number. This infers that shrubs with more stems tend to have bigger crowns with larger surface areas and crown diameters. Shrub crown area was very strongly positively correlated with shortest crown diameter (r = 0.96) and longest crown diameter (r = 0.95), indicating that shrubs with larger crown areas also tend to have larger crown diameters. In the south-facing slope, shrub height was strongly positively correlated with longest crown diameter (r = 0.72) and moderately positively correlated with shrub crown area (r = 0.68) and short crown diameter (r = 0.60) (Fig. 6b). Total stem number was moderately positively correlated with shrub crown area (r = 0.58), shrub crown area (r = 0.56) and longest crown diameter (r = 0.55). Furthermore, shrub crown area was very strongly positively correlated with the longest crown diameter (r = 0.96) and shortest crown diameter (r = 0.96). This correlation suggests that taller shrubs with more stems tend to have larger and wider crowns on the south-facing slope.
As shown in Fig. 6c, the longest crown diameter (r = 0.66), shrub crown area (r = 0.62) and shortest crown diameter (r = 0.55) were all moderately positively correlated to shrub height on the north-facing slope at Manzimdaka. This suggests that as the height of the shrub increases, the length of the longest and shortest branches as well as the size of the crown area also increase. Total stem number was strongly positively correlated with shrub crown area (r = 0.74), shortest crown diameter (r = 0.73) and moderately positively correlated with longest crown diameter (r = 0.69). This indicates that shrubs with more stems tend to have bigger crowns with larger surface areas and crown diameters. Shrub crown area was very strongly positively correlated with shortest crown diameter (r = 0.97) and longest crown diameter (r = 0.96). Shrub height was moderately positively correlated with longest crown diameter (r = 0.54), shrub crown area (r = 0.51) and shortest crown diameter (r = 0.46) in the south-facing slope of Manzimdaka (Fig. 6c). Total stem number was moderately positively correlated with shortest crown diameter (r = 0.55), shrub crown area (r = 0.54) and longest crown diameter (r = 0.48). Shrub crown area was very strongly positively correlated with shortest crown diameter (r = 0.96) and longest crown diameter (r = 0.94). This suggests that as the height and number of stems of the shrub increases, the length of the longest and shortest branches and the size of the crown area also increase.
Discussion
Effect of slope aspect on shrub density of Euryops floribundus
Slope aspect is an important topographical attribute that affects the length and intensity of solar radiation, and this alters the microclimate of the faces of the north-facing and south-facing slopes (Higgins et al., 1999; Schulze and McGee, 1978). Hillslope aspect, therefore, has important effects on vegetation development because of larger differences in soil radiation, soil temperature, evaporation and soil moisture on various hillslope aspects (Pan et al., 2023). Contrasting aspects of the landscape may lead to differences in plant species, density and vegetative productivity (Archer et al., 2017; Gartzia et al., 2014; Gxasheka et al., 2023; Sternberg and Shoshany, 2001). At the communal grazing sites investigated, we found that slope aspect influenced the vegetation characteristics of the Euryops floribundus species in the sampled mountainous grassland. Specifically, we found that the density of E. floribundus shrubs at Tsengiwe, which is geographically located near communal homesteads was 50% higher on the north-facing slope than on the south-facing slope. Notably, slope aspect had no effect on shrub density at Upper Mnxe and Manzimdaka, which were located at intermediate and furthest distances from the homesteads. One possible explanation for the increased shrub density on the north-facing slope at Tsengiwe is that the surface of the north-facing slope in the Southern Hemisphere, where our sites are located faces the sun and receives more solar radiation over a longer period of time (Holland and Steyn 1975; Schulze and McGee, 1978). Elevated intensity of radiation increases soil temperature, resulting in higher seed germination rates, especially for shrubby seedlings (Fig. 4a). In fact, work by Boehm et al. (2021) in a montane rangeland in the United States of America (USA) showed that warmer slope aspects experience higher seed germination, especially in winter than cooler slope aspects. This observation was also captured in a laboratory-based study on seed germination, which found that mountain shrub seeds particularly those of Asteraceae species, germinate more readily in warmer temperatures (25–35 °C), leading to an increase in shrub abundance and dominance in mountainous grasslands (Venn et al., 2021). A study by Sternberg and Shoshany (2001), who investigated the effects of aspect on woody vegetation in a semi-arid shrub land in Israel found that the density of woody plants was 31% higher on warmer slopes than on cooler slopes. The results from these studies and what was found in our work reinforces that slope aspect is indeed an important landscape attribute that influences shrub distribution across a range of environments. Another possible explanation for the higher density of shrubs on the north-facing slope in our study could be linked to the close proximity of the Tsengiwe communal grazing site to homesteads. Communal grazing sites that are located close to homesteads experience more land disturbances caused by overgrazing and harvesting of woody plants by inhabitants compared to sites that located far away from homesteads (Fisher et al., 2011; Bennett et al., 2012). These land disturbances reduce the cover and diversity of grass species leading to creation of bare grounds, which encourage the establishment of undesirable grass species and promote shrub encroachment (Fisher et al., 2011; Bennett et al., 2012; Jamil et al., 2022). In fact, earlier work by Gxasheka et al. (2017) found that the Tsengiwe site was subjected to higher levels of anthropogenic activity, particularly the harvesting and collection of Euryops floribundus shrubs, which are used as building materials and for firewood. The findings obtained in our study are in congruent to the results by Bennett et al. (2012), who found greater density of shrubs on the north-facing slopes of sites close to settlements in communal rangelands in the Eastern Cape, South Africa.
Effect of slope aspect on height, total stem number and crown cover of Euryops floribundus
Height is a key plant characteristic that distinguishes potential fitness variation across individual plants in an ecosystem (Moles et al., 2009; Nagashima and Hikosaka, 2011). It is a crucial attribute of the ecological strategies used by woody plants to ensure their long-term survival in challenging environments (Díaz et al., 2016; Falster and Westoby, 2003). In this study, the Euryops floribundus shrub showed more investment in plant height growth on the north-facing slopes than on south-facing slopes at Upper Mnxe and Manzimdaka, respectively. The taller shrubs on the north-facing slope are probably due to the larger number of mature shrubs observed at these sites (Fig. 5c). The proliferation of mature shrubs in the north-facing slope is associated with increased sunlight and warmth that provides a more favorable environment for the growth of Euryops floribundus shrubs. Mature tall shrubs tend to have a developed root system, which increases water uptake (Fan et al., 2019; Letts et al., 2010; Luo et al., 2020) leading to higher photosynthetic rate and in turn increased plant height (Helluy et al., 2021; Schenk and Jackson, 2002). Additionally, the correlation analysis revealed a strong positive relationship between shrub height of Euryops floribundus and its longest crown diameters implying that an increase in shrub height promotes longest crown diameters. The development of such a shrub architecture allows for the shadowing effect of the longest crown diameters, and in so doing enhances soil moisture availability by reducing evapotranspiration (Dai et al., 2009; Lozano-Parra et al., 2018). Despite the increased shrub height on north-facing slopes in Upper Mnxe and Manzimdaka, our research work also found that the north-facing slope in Tsengiwe had lower shrub height and shrub cover. These results were linked to the abundance of shrub seedlings on the north-facing slope in Tsengiwe (Fig. 5a). The proliferation of shrub seedlings at the site near homesteads may indicate more recent establishment, potentially as a result of human-induced disturbances linked to collection of building materials and firewood from the shrub Euryops floribundus (Gxasheka et al., 2017; Shackleton and Gambiza, 2008).

Boxplots showing Euryops floribundus height class distribution of seedlings (0–1 m) (a), young shrubs (> 1–2 m) (b) and Mature shrubs (> 2 m) (c) between north-facing and south-facing slope aspects in Tsengiwe, Upper Mnxe and Manzimdaka. Box plot elements: box = values of 25th and 75th percentiles; horizontal line = median; whiskers = values of 5th and 95th percentiles. Black dots, circle and triangle shapes outside the whiskers represent the outliers. Photo credits: Masibonge Gxasheka
In Upper Mnxe and Manzimdaka, the shrub crown cover was higher (33 and 11%) on the north-facing slope compared to the south-facing slope. Similar to the results obtained in our study, Hottman and O’Connor (1999) found that woody cover was 56% higher on north-facing slopes than south-facing slopes in semi-arid mountainous rangelands in South Africa. The authors ascribed their findings to the ability of particular woody plants to encroach (such as Acacia tortilis and Maytenus heterophylla), thus reducing grass cover, altering species richness and soil fertility. In southwest Yukon, Canada, Kambo and Danby (2018) found that shrub cover was 47% higher on warmer slopes compared to cooler slopes. The increase in shrub cover of Euryops floribundus on north-facing slopes could be explained by the positive correlation between shrub height, number of stems and crown area. These results suggest that the shrub Euryops floribundus grows taller with more stems on the north-facing slopes, thus providing a large and dense crown cover. Shrubs with large dense crowns on the north-facing slope influence the local microclimate by improving abiotic conditions and enhancing plant growth. Previous studies have shown that shrub architecture in dry environments can reduce seasonal temperatures (i.e., cooler summers and warmer winters), resulting in enhanced soil moisture in summer (D’Odorico et al., 2010, 2013; He et al., 2010; Thomas et al., 2018). Specifically, tall shrubs with large crowns have a significant impact on surface cooling as they cast more extensive shadows, which reduce the amount of solar energy that reaches the ground. Ultimately, this reduces evaporation from the soil surface and enhances plant growth (Thomas et al., 2018). Large crowns also increase shrub fog precipitation during fogging times, which enhances water availability (Brantley and Young, 2010; Kennedy and Sousa, 2006). Furthermore, tall shrubs with large crowns reduce wind permeability, air movement and convection, and this decreases evapotranspiration rates in the surrounding atmosphere (Adams, 2009; Thomas et al., 2018).
In Tsengiwe and Upper Mnxe, north-facing slopes had a higher total stem number than south-facing slopes. The increase in total stem number on the north-facing slope was likely due to higher crown cover of the Euryops floribundus shrubs. This is supported by the positive relationship between total stem number, longest crown diameter, shortest crown diameter and crown area depicted in Fig. 6a-b. This relationship can be interpreted as a self-adaptation strategy used by Euryops floribundus, whereby the large crown cover interacts with the local climate and establishes a more favorable microclimate by providing shade, subsequently reducing the amount of direct sunlight reaching the ground. The shading effect results in more favorable soil temperatures and moisture levels. Consequently, the favorable microclimate encourages the growth of additional stems, which in turn enhances the shrub crown cover. Furthermore, additional stems improve the shrub’s capacity to accumulate resources (i.e., water and nutrients), enhancing the ability to survive and persist in unfavorable environments (Götmark et al., 2016; Matsushita et al., 2010; Ye et al., 2014). Previous research has also suggested that shrub species have more stems in stressful environments (Wilson, 1995; Nzunda and Lawes, 2011; Scheffer et al., 2014; Götmark et al., 2016). For instance, Ye et al. (2014) explored local-scale factors (i.e., topography) that influence the growth of multi-stemmed trees and found that stressful environment (e.g., low soil moisture and nutrient availability) encourage shrubs to develop more stems. Lastly, multi-stemmed trees exchange their carbohydrate reserves among their stems which improves survival, growth and resistance to environmental stressors (Del Tredici, 2001). Therefore, the shrub Euryops floribundus may also benefit from a higher total stem number by storing more carbohydrates among the stems, acting as a buffer against environmental stress, thus improving its survival in the mountainous grassland (del Tredici, 2001; Kobe, 1997; Tanentzap et al., 2012).

Pearson’s correlation’s correlation heatmap among structural characteristics of Euryops loribundus in (a) Tsengiwe, b Upper Mnxe and c Manzimdaka. SH = shrub height, TNS = total number of stem, LCD = longest crown diameter; SCD = shortest crown diameter, SCA = shrub crown area. The blue color gradient shows highest significantly positive correlation while the red color gradient shows highly negative correlation
Limitation of the study
Although our research provides useful insights, it is crucial to acknowledge a significant limitation concerning the lack of replications for the distance to homestead factor. This limitation requires replication of the study sites, which was hindered by logistical and financial constraints, resulting in pseudoreplication (Hurlbert, 1984). While, this limitation is common in field ecological research, it should not limit our ability to draw insights from the results obtained in our study (Davies and Gray, 2015). These findings should be interpreted with caution. Despite this limitation, the study provides valuable insights on the effects of slope aspect and human disturbance on Euryops floribundus encroachment patterns in mountainous grasslands. Additionally, these findings provide vital guidance for stakeholders involved in the management and conservation of mountainous grasslands.
Conclusion
The aim of this study was to investigate the influence of slope aspect on the density and structural characteristics of Euryops floribundus N.E. Br shrub in communal grazing sites located at different distances from homesteads across a semi-arid mountainous grassland. Our research revealed that the north-facing slope at Tsengiwe (site near homesteads) had a higher density of shrubs than the south-facing slope. Additionally, our results revealed that the north-facing slope at Upper Mnxe (intermediate site to homestead) and Manzimdaka (furthest site to homesteads) had higher shrub height, total stem number and crown cover compared to the south-facing slope. These findings suggest that the warmer north-facing slope provides more favorable conditions for shrub growth and survival as the distance from homesteads increases. Also, our research results showed a significant positive relationship between the height of shrubs and their longest crown diameter, shortest crown diameter and shrub crown area. Such a relation suggests that the E. floribundus shrub species employs its structural characteristics to survive and thrive more in the north-facing slope of the communal grazing land. Our research results on the landscape distribution of the density and structural characteristics of E. floribundus shrub can be used as a baseline for future monitoring of the extent and intensity of this shrub species in the studied semi-arid mountainous grassland. This information is also critical in improving our understanding of shrub encroachment patterns, and this will enable the development of a suitable management strategy for restoring the diversity and ecological integrity of semi-arid mountainous grasslands. E. floribundus shrub is a major encroacher in the mountainous grasslands in the Chris Hani District of the Eastern Cape Province, so it must be given top priority by land managers. To unravel the impact of topographic factors like slope aspect on the structure of encroaching shrubs, future research should consider quantifying the microclimatic related data including soil temperature, relative humidity, soil moisture and light intensity on different slope aspects and under the shrub canopy of E. floribundus. This information will provide insight into the thermal, water and light conditions affecting the growth and survival of the shrubs and how they may change with slope aspect and in response to future climate change.
Data availability
The data for the current study will be made available upon reasonable request.
References
Adams, J. (2009). Vegetation-climate interaction: how plants make the global environment. Springer Science & Business Media.
Aliber, M., & Xabadiya, A., (2020). GTAC/CBPEP/EU project on employment-intensive rural land reform in South Africa: policies, programmes and capacities.
Archer, S. R., Andersen, E. M., Predick, K. I., Schwinning, S., Steidl, R. J., & Woods, S. R. (2017). Woody plant encroachment: Causes and consequences. In D. D. Briske (Ed.), Rangeland systems: Processes, management and challenges (pp. 25–48). Springer.
Van Auken, O. W. (2000). Shrub invasions of North American semiarid grasslands. The Annual Review of Ecology, Evolution, and Systematics, pp 197–215.
Auslander, M., Nevo, E., & Inbar, M. (2003). The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores. Journal of Arid Environments, 55(3), 405–416.
Bardgett, R. D., Bullock, J. M., Lavorel, S., Manning, P., Schaffner, U., Ostle, N., Chomel, M., Durigan, G., Fry, E. L., Johnson, D., Lavallee, J. M., Le Provost, G., Luo, S., Png, K., Sankaran, M., Hou, X., Zhou, H., Ma, L., Ren, W., … Shi, H. (2021). Combatting global grassland degradation. Nature Reviews Earth & Environment, 2(10), 720–735.
Bennett, J. E., Palmer, A. R., & Blackett, M. A. (2012). Range degradation and land tenure change: insights from a ‘released’communal area of Eastern Cape Province, South Africa. Land Degrad Dev, 23(6), 557–568.
Bennie, J., Huntley, B., Wiltshire, A., Hill, M. O., & Baxter, R. (2008). Slope, aspect and climate: Spatially explicit and implicit models of topographic microclimate in chalk grassland. Ecological Modelling, 216(1), 47–59.
Boehm, A. R., Hardegree, S. P., Glenn, N. F., Reeves, P. A., Moffet, C. A., & Flerchinger, G. N. (2021). Slope and aspect effects on seedbed microclimate and germination timing of fall-planted seeds. Rangeland Ecology & Management, 75, 58–67.
Brantley, S. T., & Young, D. R. (2010). Linking light attenuation, sunflecks, and canopy architecture in mesic shrub thickets. Plant Ecology, 206(2), 225–236.
Carletti, P., Vendramin, E., Pizzeghello, D., Concheri, G., Zanella, A., Nardi, S., & Squartini, A. (2009). Soil humic compounds and microbial communities in six spruce forests as function of parent material, slope aspect and stand age. Plant and Soil, 315(1), 47–65.
Caviezel, C., Hunziker, M., & Kuhn, N. J. (2017). Green alder encroachment in the European Alps: The need for analyzing the spread of a native-invasive species across spatial data. CATENA, 159, 149–158.
Coop, J. D., & Givnish, T. J. (2007). Spatial and temporal patterns of recent forest encroachment in montane grasslands of the Valles Caldera, New Mexico, USA. Journal of Biogeography, 34(5), 914–927.
D’Odorico, P., Fuentes, J. D., Pockman, W. T., Collins, S. L., He, Y., Medeiros, J. S., De Wekkar, S., & Litvak, M. E. (2010). Positive feedback between microclimate and shrub encroachment in the northern Chihuahuan desert. Ecosphere, 1(6), 1–11
Dai, X., Jia, X., Zhang, W., Bai, Y., Zhang, J., Wang, Y., & Wang, G. (2009). Plant height–crown radius and canopy coverage–density relationships determine above-ground biomass–density relationship in stressful environments. Biology Letters, 5(4), 571–573.
Davies, G. M., & Gray, A. (2015). Don’t let spurious accusations of pseudoreplication limit our ability to learn from natural experiments (and other messy kinds of ecological monitoring). Ecology and Evolution, 5(22), 5295–5304.
Del Tredici, P. (2001). Sprouting in temperate trees: A morphological and ecological review. Botanical Review, 67(2), 121–140.
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S., Reu, S., Kleyer, M., Wirth, C., Prentice, C., Garnier, E., Bönisch, G., Westoby, M., Poorter, H., Reich, P. B., Moles, A. T., Dickie, J., Gillison, A. N., Zanne, A. E., Chave, J., Joseph Wright, S., Sheremet'ev, S. N., Jactel, H., Baraloto, C., Cerabolini, B., Pierce, S., Shipley, B., Kirkup, D., Casanoves, F., Joswig, J. S., Günther, A., Falczuk, V., Rüger, N., Mahecha, M. D., & Gorné, L. D. (2016). The global spectrum of plant form and function. Nature, 529(7585), 167–171
Ding, J., & Eldridge, D. (2022). The success of woody plant removal depends on encroachment stage and plant traits. Nature Plants, 1–10.
Dlamini, P., Mbanjwa, V., Gxasheka, M., Tyasi, L., & Sekhohola-Dlamini, L. (2019). Chemical stabilisation of carbon stocks by polyvalent cations in plinthic soil of a shrub-encroached savanna grassland, South Africa. Catena, 181, 104088.
D’odorico, P., He, Y., Collins, S., De Wekker, S. F., Engel, V., & Fuentes, J. D. (2013). Vegetation–microclimate feedbacks in woodland–grassland ecotones. Global Ecology and Biogeography, 22(4), 364–379.
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Roger, E., Reynolds, J. F., & Whitford, W. G. (2011). Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecology Letter, 14(7), 709–722
Endress, B. A., Naylor, B. J., Parks, C. G., & Radosevich, S. R. (2007). Landscape factors influencing the abundance and dominance of the invasive plant Potentilla recta. Rangeland Ecology & Management, 60(3), 218–224.
Falster, D. S., & Westoby, M. (2003). Plant height and evolutionary games. Trends in Ecology & Evolution, 18(7), 337–343.
Fan, Y., Li, X. Y., Huang, H., Wu, X. C., Yu, K. L., Wei, J. Q., Zhang, C. C., Wang, P., Hu, X., & D’Odorico P. (2019). Does phenology play a role in the feedbacks underlying shrub encroachment? Science of the Total Environment, 657, 1064–1073.
Fisher, J. T., Witkowski, E. T. F., Erasmus, B. F. N., Van Aardt, J., Asner, G. P., Wessels, K. J., & Mathieu, R. (2012). Human-modified landscapes: patterns of fine-scale woody vegetation structure in communal savannah rangelands. Environ Conserv 39(1), 72–82.
Gaillard, C., Langan, L., Pfeiffer, M., Kumar, D., Martens, C., Higgins, S. I., & Scheiter, S. (2018). African shrub distribution emerges via a trade‐off between height and sapwood conductivity. J Biogeogr, 45(12), 2815–2826
Gallardo-Cruz, J. A., Pérez-García, E. A., Meave, J. A. (2009). β-Diversity and vegetation structure as influenced by slope aspect and altitude in a seasonally dry tropical landscape. Landsc Ecol, 24, 473-482.
Grab, S. W. (2013). Fine-scale variations of near-surface-temperature lapse rates in the high Drakensberg Escarpment, South Africa: environmental implications. Arct Antarct Alp Res, 45(4), 500–514.
Gartzia, M., Alados, C. L., & Perez-Cabello, F. (2014). Assessment of the effects of biophysical and anthropogenic factors on woody plant encroachment in dense and sparse mountain grasslands based on remote sensing data. Progress in Physical Geography, 38(2), 201–217.
Gatica, G., Escudero, A., & Pucheta, E. (2020). Livestock settlement affects shrub abundance via plant functional diversity but not species richness in arid environments. Plant Ecology, 221, 1253–1264.
Gibson, D. J. (2009). Grasses and Grassland Ecology. Oxford University Press.
Götmark, F., Götmark, E., & Jensen, A. M. (2016). Why be a shrub? A basic model and hypotheses for the adaptive values of a common growth form. Frontiers in Plant Science, 1095
Gxasheka, M., Beyene, S. T., Mlisa, N. L., & Lesoli, M. (2017). Farmers’ perceptions of vegetation change, rangeland condition and degradation in three communal grasslands of South Africa. Tropical Ecology, 58, 217–228.
Gxasheka, M., Gajana, C. S., & Dlamini, P. (2023). The role of topographic and soil factors on woody plant encroachment in mountainous rangelands: A mini literature review. Heliyon, 9, e20615. https://doi.org/10.1016/j.heliyon.2023.e20615
Gxasheka, M. (2013). Euryops Floribuntus Encroachment in Eastern Cape Communal Rangelands: Indigenous and Scientific Understanding of Effects on Range Ecology, and Food Security. MSc dissertation, University of Fort Hare.
Haile, M., Birhane, E., Rannestad, M. M., & Adaramola, M. S. (2021). Expansive shrubs: Expansion factors and ecological impacts in Northern Ethiopia. Journal for Nature Conservation, 61, 125996.
He, Y., D'Odorico, P., De Wekker, S. F., Fuentes, J. D., & Litvak, M. (2010). On the impact of shrub encroachment on microclimate conditions in the northern Chihuahuan desert. J Geophys Res Atmos 115, (D21).
Helluy, M., Gavinet, J., Prévosto, B., & Fernandez, C. (2021). Influence of light, water stress and shrub cover on sapling survival and height growth: the case of A. unedo, F. ornus and S. domestica under Mediterranean climate. European Journal of Forest Research, 140(3), 635–647
Hesselbarth, M. H. K., Wiegand, K., Dreber, N., Kellner, K., Esser, D., & Tsvuura, Z. (2018). Density-dependent spatial patterning of woody plants differs between a semi-arid and a mesic savanna in South Africa. Journal of Arid Environments, 157, 103–112.
Higgins, S. I., Richardson, D. M., Cowling, R. M., & Trinder-Smith, T. H. (1999). Predicting the landscape-scale distribution of alien plants and their threat to plant diversity. Biological Conservation, 13(2), 303–313.
Hirmas, D. R., & Graham, R. C. (2011). Pedogenesis and soil-geomorphic relationships in an arid mountain range, Mojave Desert, California. Soil Sci Soc Am J, 75(1), 192–206.
Holland, P.G., & Steyn, D. G. (1975) Vegetational responses to latitudinal variations in slope angle and aspect. J Biogeogr, 2, 179-183.
Hottman, M. T., & O’Connor, T. G. (1999). Vegetation change over 40 years in the Weenen/Muden area, KwaZulu-Natal: Evidence from photo-panoramas. The African Journal of Range & Forage Science, 16(2–3), 71–88.
Huntsinger, L., & Hopkinson, P. (1996). Sustaining rangeland landscapes: A social and ecological process. Journal of Range Management, 49(2), 167–173.
Hurlbert, S. H. (1984). Pseudoreplication and the design of ecological field experiments. Ecological Monographs, 54(2), 187–211.
Inman, E. N., Hobbs, R. J., Tsvuura, Z., Valentine, L. (2020). Current vegetation structure and composition of woody species in community‐derived categories of land degradation in a semiarid rangeland in Kunene region, Namibia. Land Degrad Dev, 31(18), 2996–3013.
Jamil, A., Zubair, M., Manzoor, S. A., Wali Muhammad, M., Yasin, G., Ur Rahman, S., Alzain, M. N., Alqarawi, A. A., & Abd_Allah , E. F. (2022). Impact of human settlements on diversity of range vegetation. Sustainability, 14(1), 519.
Jasińska, J., Sewerniak, P., Markiewicz, M. (2019). Links between slope aspect and rate of litter decomposition on inland dunes. Catena, 172, 501–508.
Johnson, D. D., & Miller, R. F. (2006). Structure and development of expanding western juniper woodlands as influenced by two topographic variables. Forest Ecology and Management, 229(1–3), 7–15.
Kakembo, V., Rowntree, K., & Palmer, A. R. (2007). Topographic controls on the invasion of Pteronia incana (Blue bush) onto hillslopes in Ngqushwa (formerly Peddie) district, Eastern Cape, South Africa. Catena, 70(2), 185–199.
Kambo, D., & Danby, R. K. (2018). Factors influencing the establishment and growth of tree seedlings at Subarctic alpine treelines. Ecosphere, 9(4), e02176.
Kennedy, P. G., & Sousa, W. P. (2006). Forest encroachment into a Californian grassland: Examining the simultaneous effects of facilitation and competition on tree seedling recruitment. Oecologia, 148(3), 464–474.
Kobe, R. K. (1997). Carbohydrate allocation to storage as a basis of interspecific variation in sapling survivorship and growth. Oikos, pp 226–233.
Kutiel, P., & Lavee, H. (1999). Effect of slope aspect on soil and vegetation properties along an aridity transect. Isr J Plant Sci, 47(3), 169–178.
Letts, M. G., Johnson, D. R., & Coburn, C. A. (2010). Drought stress ecophysiology of shrub and grass functional groups on opposing slope aspects of a temperate grassland valley. Botany, 88(9), 850–866.
Lieffering, M., Newton, P. C., Brock, S. C., & Theobald, P. W. (2019). Some effects of topographic aspect on grassland responses to elevated CO2. Plant Prod Sci, 22(3), 345–351.
Longman, R. J., Giambelluca, T. W., Alliss, R. J., Barnes, M. L. (2014). Temporal solar radiation changes at high elevations in Hawai ‘i. J Geophys Res Atmos, 119(10), 6022–6033.
Mucina, L., & Rutherford, M. C. (eds) (2006). The vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. Pretoria, South Africa: South Africa National Biodiversity Institute.
Lozano-Parra, J., Pulido, M., Lozano-Fondón, C., & Schnabel, S. (2018). How do soil moisture and vegetation covers influence soil temperature in drylands of Mediterranean regions? Water, 10(12), 1747.
Luo, H., Xu, H., Chu, C., He, F., & Fang, S. (2020). High temperature can change root system architecture and intensify root interactions of plant seedlings. Frontiers in Plant Science, 11, 160.
Masibonge, G., Tefera, S., & Mota, L. (2019). Invasion of Euryops floribundus in degraded South African communal grassland: Unpacking the invasion relationship with elevation, soil properties, the quality and quantity of the herbaceous layer. African Journal of Ecology, 58(1), 69–79.
Matsushita, M., Tomaru, N., Hoshino, D., Nishimura, N., & Yamamoto, S. I. (2010). Factors affecting the production, growth, and survival of sprouting stems in the multi-stemmed understory shrub Lindera triloba. Botany, 88(2), 174–184.
Majeed, M., Khan, A. M., Habib, T., Anwar, M. M., Sahito, H. A., Khan, N., & Ali, K. (2022). Vegetation analysis and environmental indicators of an arid tropical forest ecosystem of Pakistan. Ecol Indic 142, 109291.
Mogashoa, R., Dlamini, P., & Gxasheka, M. (2021). Grass species richness decreases along a woody plant encroachment gradient in a semi-arid savanna grassland, South Africa. Landscape Ecology 36(2), 617–636.
Moleele, N. M., Ringrose, S., Matheson, W., & Vanderpost, C. (2002). More woody plants? The status of bush encroachment in Botswana’s grazing areas. Journal of Environmental Management, 64(1), 3–11.
Moles, A. T., Warton, D. I., Warman, L., Swenson, N. G., Laffan, S. W., Zanne, A. E., Pitman, A., Hemmings, F. A., & Leishman, M. R. (2009). Global patterns in plant height. Journal of Ecology, 97(5), 923–932.
Montané, F., Rovira, P., & Casals, P. (2007). Shrub encroachment into mesic mountain grasslands in the Iberian peninsula: effects of plant quality and temperature on soil C and N stocks. Global Biogeochemical Cycles, 21(4).
Nagashima, H., & Hikosaka, K. (2011). Plants in a crowded stand regulate their height growth so as to maintain similar heights to neighbours even when they have potential advantages in height growth. Annals of Botany, 108(1), 207–214.
Nzunda, E. F., & Lawes, M. J. (2011). Costs of resprouting are traded off against reproduction in subtropical coastal dune forest trees. Plant Ecology, 212(12), 1991–2001.
O’connor, T. G. (2005). Influence of land use on plant community composition and diversity in Highland Sourveld grassland in the southern Drakensberg, South Africa. Journal of Applied Ecology, 42(5), 975–988.
Osborne, C. P., Charles-Dominique, T., Stevens, N., Bond, W. J., Midgley, G., & Lehmann, C. E. (2018). Human impacts in African savannas are mediated by plant functional traits. New Phytologist, 220(1), 10–24.
Palmer, A. R., & Ainslie, A. M. (2005). Grasslands of South Africa. Grasslands of the World, 34, 77.
Pan, Y., Ren, L., Xiang, X., Huo, J., Meng, D., Wang, Y., Yu, C., & Huang, Y. (2023). Effect of slope aspect on plant above‐and belowground functional traits of Alpine Meadow on the Qinghai‐Tibet Plateau, China. Journal of Geophysical Research: Biogeosciences, 128(2), e2022JG007268. https://doi.org/10.1029/2022JG007268
Papanastasis, V. P., Bautista, S., Chouvardas, D., Mantzanas, K., Papadimitriou, M., Mayor, A. G., Koukioumi, P., Papaioannou, A., & Vallejo, R. V. (2017). Comparative assessment of goods and services provided by grazing regulation and reforestation in degraded Mediterranean rangelands. Land Degradation and Development, 28(4), 1178–1187.
Pekár, S., & Brabec, M. (2016). Modern analysis of biological data: Generalized linear models in R. Masaryk University Press.
R Core Team (2022). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available from: https://www.r-project.org/
Ralph Clark, V., Kunkel, K., O’Connor, T., Jewitt, D., Sekhele, N., & Kirkman, K. (2021). Montane rangelands in a changing world. The African Journal of Range & Forage Science, 38(1), iii–vi.
Raper, P. E., Moller, L. A., & du Plessis, T. L. (2014). Dictionary of Southern African place names. Jonathan Ball Publishers.
Roques, K. G., O’connor, T. G., & Watkinson, A. R. (2001). Dynamics of shrub encroachment in an African savanna: Relative influences of fire, herbivory, rainfall and density dependence. Journal of Applied Ecology, 38(2), 268–280.
Sakhisiwe Local Municipality. (2017). Socio-economic review and outlook 2017. Vincent: Eastern Cape Socio-Economic Consultative Council.
Sala, O., Vivanco, L., & Flombaum, P. (2013). Grassland ecosystems. In Encyclopedia of Biodiversity: Second Edition (pp. 1–7). Elsevier Inc. https://doi.org/10.1016/B978-0-12-384719-5.00259-8
Scheffer, M., Vergnon, R., Cornelissen, J. H. C., Hantson, S., Holmgren, M., van Nes, E. H., & Xu, C. (2014). Why trees and shrubs but rarely trubs? Trends in Ecology & Evolution, 29(8), 433–434.
Schenk, H. J., & Jackson, R. B. (2002). Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. Journal of Ecology, pp 480–494.
Schober, P., Boer, C., & Schwarte, L. A. (2018). Correlation coefficients: Appropriate use and interpretation. Anesthesia and Analgesia, 126(5), 1763–1768.
Scholes, R. J., & Archer, S. R. (1997). Tree-grass interactions in savannas. The Annual Review of Ecology, Evolution, and Systematics, pp 517–544.
Schulze, R. E., & McGee, O. S. (1978). Climatic indices and classifications in relation to the biogeography of southern Africa. In Biogeography and ecology of southern Africa (pp. 19–52). Springer.
Shackleton, C. M. (1993). Fuelwood harvesting and sustainable utilisation in a communal grazing land and protected area of the eastern Transvaal lowveld. Biol Conserv, 63(3), 247–254.
Shackleton, C. M., & Gambiza, J. (2008). Social and ecological tradeoffs in combating land degradation: The case of invasion by a woody shrub (Euryops floribundus) at Macubeni, South Africa. Land Degradation and Development, 19(4), 454–464.
Soubry, I., Robinov, L., Chu, T., & Guo, X. (2022). Mapping shrub cover in grasslands with an object-based approach and investigating the connection to topo-edaphic factors. Geocarto Int, 37(27), 16926–16950.
Squires, V. R., Dengler, J., Hua, L., & Feng, H. (Eds.) (2018) Grasslands of the world: diversity, management and conservation. CRC Press.
Steger, C., Boone, R. B., Dullo, B. W., Evangelista, P., Alemu, S., Gebrehiwot, K., & Klein, J. A. (2022). Collaborative agent-based modeling for managing shrub encroachment in an Afroalpine grassland. Journal of Environmental Management, 316, 115040.
Sternberg, M., & Shoshany, M. (2001). Influence of slope aspect on Mediterranean woody formations: Comparison of a semiarid and an arid site in Israel. Ecological Research, 16(2), 335–345.
Tanentzap, A. J., Mountford, E. P., Cooke, A. S., & Coomes, D. A. (2012). The more stems the merrier: Advantages of multi-stemmed architecture for the demography of understorey trees in a temperate broadleaf woodland. Journal of Ecology, 100(1), 171–183.
Thomas, A. D., Elliott, D. R., Dougill, A. J., Stringer, L. C., Hoon, S. R., & Sen, R. (2018). The influence of trees, shrubs, and grasses on microclimate, soil carbon, nitrogen, and CO2 efflux: Potential implications of shrub encroachment for Kalahari rangelands. Land Degradation and Development, 29(5), 1306–1316.
Van Auken, O. W. (2009). Causes and consequences of woody plant encroachment into western North American grasslands. Journal of Environmental Management, 90(10), 2931–2942.
Venn, S. E., Gallagher, R. V., & Nicotra, A. B. (2021). Germination at extreme temperatures: Implications for alpine shrub encroachment. Plants, 10(2), 327.
Vetter, S., Goqwana, W. M., Bond, W. J., & Trollope, W. W. (2006). Effects of land tenure, geology and topography on vegetation and soils of two grassland types in South Africa. The African Journal of Range & Forage Science, 23(1), 13–27.
Wei, T., Simko, V., Levy, M., Xie, Y., Jin, Y., & Zemla, J. (2017). Package ‘corrplot.’ Statistician, 56(316), e24.
Weisberg, P. J., Lingua, E., & Pillai, R. B. (2007). Spatial patterns of pinyon–juniper woodland expansion in central Nevada. Rangeland Ecology & Management, 60(2), 115–124.
Wickham, H., & Grolemund, G, (2016), R for data science: import, tidy, transform, visualize, and model data. " O'Reilly Media, Inc.".
Wickham, H., & Wickham, M. H. (2020). Package ‘plyr’. Obtenido Httpscran Rproject Orgwebpackagesdplyrdplyr Pdf.
Wilson, B. F. (1995). Shrub stems: form and function. In Plant stems (pp. 91–102). Academic Press.
Wu, M., He, H. S., Zong, S., Tan, X., Du, H., Zhao, D., Liu, K., & Liang, Y. (2018). Topographic controls on vegetation changes in alpine tundra of the Changbai Mountains. Forests, 9(12), 756
Ye, J., Hao, Z., Wang, X., Bai, X., Xing, D., & Yuan, Z. (2014). Local-scale drivers of multi-stemmed tree formation in Acer, in a temperate forest of Northeast China. Chinese Science Bulletin, 59(3), 320–325.
Zizka, A., Govender, N., & Higgins, S. I. (2014.) How to tell a shrub from a tree: A life‐history perspective from a South African savanna. Austral Ecol, 39(7), 767–778.
Acknowledgements
Funding for this study was provided by the National Research Foundation (NRF-BAAP) project Ref: BAAP200224506520 and Grant Number: 129426. We wish to extend our heartfelt appreciation to the community members in the study sites for their indispensable contribution and unwavering support throughout the data collection phase of this research. Our thanks also go to the student assistants from the University of Fort Hare and Fort Cox Agriculture and Forestry Training Institute for their support and assistance on the fieldwork. Finally, we thank the anonymous reviewers and editor for their great suggestions and valuable input.
Funding
Open access funding provided by University of Limpopo.
Author information
Authors and Affiliations
Contributions
The experiment was conceived and organized by MG and PD. MG did the field sampling. MG entered the data into MS Excel and conducted statistical analysis. MG wrote the first draft of the paper; PD offered feedback on the earlier drafts. The final manuscript has been read by and approved by all authors.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they do not have any conflicts of interest, either financial or otherwise, that would have affected this manuscript.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gxasheka, M., Dlamini, P. Slope aspect of a mountainous grassland landscape shapes the structure of an encroaching shrub (Euryops floribundus N.E. Br): insights from communal grazing sites located at varying distances from rural homesteads. COMMUNITY ECOLOGY 25, 127–143 (2024). https://doi.org/10.1007/s42974-024-00182-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42974-024-00182-w