
Abstract
Tuta absoluta, an invasive pinworm characterized by high reproductive potential and adaptation to different agroecological conditions, cause serious damage to tomato crops. Chemical control with synthetic insecticides is widely used to control this pest, although pesticides exhibit side effects on non-target organisms and negatively impact the environment, with the occurrence of resistance to some active substances in the target pest. The use of essential oils (EOs) from aromatic or officinal plants could represent an environmentally safe control method, alternative to synthetic insecticide application. In this work we investigated the effect of EOs from Spanish oregano, laurel, basil, garlic, peppermint, cypress, and eucalyptus EOs on T. absoluta oviposition deterrence and larval repellency. Results showed a remarkable oviposition deterrence using laurel, Spanish oregano, basil, and garlic EOs in both laboratory and greenhouse experiments. In no-choice repellence tests carried 1, 2, 6, and 22 h after treatment application, peppermint and Spanish oregano EOs showed the most lasting effects while cypress and eucalyptus EOs were not effective after 6 and 22 h, respectively. In choice tests, at either 5 or 22 h after treatment application, low preference for the test over control by larvae was observed in all treatments, except for basil EO. The most promising effects were observed in Spanish oregano and laurel treatments where no larvae were detected. Repellent effectiveness of EOs does not seem to be influenced when larvae did not get a chance to do a choice between treated and untreated leaves. This study highlights the effectiveness of EOs of aromatic or officinal plants as an alternative to synthetic insecticides for the control of T. absoluta.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The South American tomato pinworm, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) is one of the most damaging pests of tomato (Solanum lycopersicum L.) crops, both in open fields and under greenhouses. This invasive pest, originating from South America, cause economic and ecological problems due to its rapid dispersion challenging pest control programs. Currently it can be found throughout Africa, Europe, Middle East and Asia (Campos et al. 2017; Biondi et al. 2018; Mansour et al. 2018; Han et al. 2019). In Italy, T. absoluta was reported for the first time in 2008 and rapidly spread to the tomato growing areas in all regions of the country (Speranza and Sannino 2012). In new colonization areas, the tomato pinworm has displayed great ability to adapt to different agro-ecological conditions, exhibiting high reproductive potential and significant economic damage (Desneux et al. 2010, 2011; Urbaneja et al. 2013). Under Mediterranean conditions, adults of T. absoluta can be detected throughout the whole year. Females generally mate only once in a day and up to six times during their lifespan. They oviposit preferentially on both sides of the leaves located in the plant apex (73%) and to a lesser extent on leaf veins and stem margin (21%), sepals (5%) or green fruits (1%) (Estay 2000). At 25–27 °C, the pre-oviposition period lasts for 2–3 days. A single female can lay up to 260 eggs throughout its life (Uchoa-Fernandes et al. 1995). After hatching, larvae explore the substrate for 5–10 min to about an hour, before digging a mine in the leaf parenchyma. Larvae feed and develop on leaves, stems, buds, flowers and fruits, damaging crop with yield losses that can reach 100% (Desneux et al. 2010, 2011; Biondi et al. 2018). The control of T. absoluta in tomato largely relies on intensive use of synthetic insecticides. However, chemical control of T. absoluta is very difficult, because the moth has a remarkable tendency to rapidly acquire resistance against insecticides. Furthermore, synthetic insecticides exhibit side effects on non-target organisms and negatively impact the environment (Han et al. 2018; Guedes et al. 2019). Therefore, researchers have focused on control approaches of T. absoluta that exhibit limited negative effects on the environment, which are based on the use of aromatic or officinal plant compounds such essential oils (EOs) (Soares et al. 2019). Indeed, plant-based bioinsecticides are cheap, biodegradable, more effective and species-specific than synthetic insecticides (Andrade-Ochoa et al. 2018). A number of different EOs shows a broad spectrum of activity against pests of many crops and stored products, exhibiting insecticidal, antifeedant, repellent, oviposition deterrent, growth regulatory, and antivector effects (Isman 2006; Said-Al Ahl et al. 2017; Bett et al. 2017; Rajkumar et al. 2019; Abdelkader et al. 2020; Chintalchere et al. 2021). In particular, the repellent and oviposition deterrence activities of EOs extracted from aromatic or officinal plants are well known. However, most of these studies were conducted on pests belonging to Coleoptera and Diptera species (Nerio et al. 2010; Mossa 2016; Marsin et al. 2020). Given the efficacy of some botanicals against T. absoluta, it is possible to use them in combination with soft pesticides in order to develop effective integrated pest management approaches that target this pest (Tarusikirwa et al. 2020). Currently, many studies report ovicidal, larvicidal and adulticidal activities of EOs against this pinworm (Magalhães et al. 2001; Umpiérrez et al. 2012; Alam et al. 2019; Esther et al. 2019; Lo Pinto et al. 2019, 2020; Tarusikirwa et al. 2020; Mansour and Biondi 2021); however oviposition deterrence and repellence effects are less investigated (Aynalem 2018; Marsin et al. 2020). In this perspective, the aim of the present study was to assess the effectiveness of EOs from selected plants that are understudies in terms of oviposition deterrence and repellency activity against T. absoluta. Specifically, the effectiveness of EOs of Spanish oregano (Thymus capitatus L., synonym Thymbra capitata, subgenus Coridothymus capitatus L. Reichemb. F.), laurel (Laurus nobilis L.), basil (Ocimum basilicum L.), garlic (Allium sativum L.), peppermint (Mentha piperita L.), cypress (Cupressus sempervirens L.), and eucalyptus (Eucalyptus globulus L.) was investigated. Our objective was to assess: 1) effects on the oviposition deterrence in choice tests using (i) Spanish oregano and laurel EOs in laboratory and (ii) Spanish oregano, laurel, basil, and garlic EOs in greenhouse; 2) effects on repellency against third instar larvae in laboratory, using (iii) Spanish oregano, peppermint, cypress, and eucalyptus EOs in no-choice tests and (iv) Spanish oregano, peppermint, cypress, eucalyptus, laurel, and basil EOs in choice tests.
Materials and methods
Plant essential oils
Natural EOs (100% pure industrial products obtained by steam distillation method, standardized and marketed as food supplements, ERBAMEA) of basil (leaves), Spanish oregano (flowering aerial parts), laurel (leaves), peppermint (leaves), cypress (cones), and eucalyptus (leaves), and macerate oil of garlic (bulbs), were used. Garlic macerate oil was prepared using the method described by Patil and Ravindra (2008).
To perform laboratory experiments, an emulsion of 200 μl of EOs with 2% Tween® 20 (SigmaAldrich) in distilled water (50 ml) per each treatment was used as test, and 2% Tween® 20 in distilled water (50 ml) was used as control. For the greenhouse experiment, an emulsion of 400 μl of EOs with 2% Tween® 20 (SigmaAldrich) in distilled water (100 ml) per each treatment was used as test and 2% Tween® 20 in distilled water (100 ml) was used as control. For garlic, two emulsions of 300 μl and 600 μl of EO were obtained as described above.
Tomato plants and insects
The biological material used in the bioassays was obtained from tomato seedlings kept in two greenhouses (greenhouse-1 and greenhouse-2). Tomato plants, Solanum lycopersicum L var. Pizzutello, were grown under greenhouse-1 (25 ± 5 °C, 50–70% relative humidity [RH], 16:08 L:D photoperiod) in plastic pots (7 × 7 × 8 cm) and were watered every 2 days. Seedlings for experiments were used when they reached about 30–40 cm in height. Tuta absoluta third instar larvae used in the bioassays were obtained from rearing of the insect on tomato plants under greenhouse-2 (in the aforementioned controlled climatic conditions). To obtain mated adults of T. absoluta, leaves with larvae were collected from plants of greenhouse-2 and placed in a plastic tray (40 × 30 × 6 cm) over a layer of sand (0.5 cm) arranged on a sheet of filter paper. Furthermore, leaves were covered with a sheet of filter paper. The tray was kept in a thermo-conditioned room in the aforementioned controlled climatic conditions. Daily, the sand was sieved and the underlying sheet of paper was observed until insect reached the pupal stage. Pupae were transferred to a Plexiglas cage (50 × 30 × 30 cm) and observed periodically until adult emergence. After mating, pairs of adults were taken from the cage to be used in tests. To obtain larvae for experiments, leaves from infested plants of greenhouse-2 were collected and brought to the laboratory. After observation under stereomicroscope, third instar larvae were carefully removed from their mines by using a fine brush and were used in the bioassays.
Experiment 1: oviposition deterrent effects in laboratory
EOs of laurel and Spanish oregano were used. Tomato leaves were taken from uninfested seedlings kept in greenhouse. In laboratory, each leaf, made up of three apical leaflets distributed along the leaf rachis, was wet with the emulsion with EOs (as test), or with water and Tween only (as control). Then leaves were singly introduced inside a glass tubes (10 × 1 cm) containing water to maintain the leaf turgor. Single pairs of compound leaves, one for the test and one for the control, were placed vertically inside Plexiglas boxes (40 × 25 × 25 cm) at a distance from each other of 30 cm. These boxes featured two side walls in tulle and a removable front window (15 × 15 cm) to facilitate transfer of leaves and insects inside the box. Subsequently, two mated pairs of T. absoluta were released into the box and left for 36 h. Insects were fed with a drop of honey placed centrally on the top of the box. Each treatment (laurel and Spanish oregano EOs) was replicated 40 times. Boxes were kept in a thermo-conditioned room under the aforementioned controlled climatic conditions. Observations were made after 36 h from the beginning of the experiment by counting the number of eggs laid on the leaves for each box.
Experiment 2: oviposition deterrent effects in greenhouse
EOs of Spanish oregano, basil, laurel and garlic were used, and for each treatment the following procedure was used: 40 uninfested plants were taken from greenhouse-1 and numbered from 1 to 40. At the beginning of the experiment, 20 plants were sprayed with the emulsion with EO (= test) and 20 plants with water and Tween (= control). Plants were transferred into a greenhouse-2 under a tunnel covered with tulle (200 × 150 cm and 90 cm roof height) without openings and provided of 4 removable windows (40 × 40 cm) along a side (at a distance of 40 cm from each other) to facilitate transfer of plants and insects inside the tunnel. Plants were randomly placed in 5 rows of 8 plants each, at a distance of 10 cm on the row and 20 cm between rows. Soon after, twenty mated pairs of T. absoluta were released into the tunnel and left for 4 days, feeding whit drops of honey placed on the top of the tunnel. After 4 days, plants were transferred to the laboratory and observed under a stereomicroscope to record the number of eggs found on leaves.
Experiment 3: repellency effects against larvae in no-choice experiments
EOs of Spanish oregano, peppermint, cypress, and eucalyptus were used. To perform experiments, uninfested compound leaves were collected from plants of the greenhouse-1 and brought to the laboratory where leaflets of similar development stage, shape and size (4 cm in length and 3 cm in width) were selected and removed from the plant. Leaflets were treated by dipping them in the solution of EOs (= test) or only in water and Tween (= control) and then left to dry for 5 min at room temperature on a non-absorbent surface. After evaporation of the access water, the leaflets were placed singly with the abaxial side into a Petri dish (5 cm diameter) containing a moist filter paper disk. For each treatment, 20 Petri dishes as tests (EOs) and 20 as controls (distilled water) were prepared. Then, one T. absoluta larva of the 3rd instar was put into each Petri dish. Three replicates were run for each tested oil. Petri dishes were kept in a thermo-conditioned room under the aforementioned controlled climatic conditions. To detect any larval activity on the leaf, the presence or absence of larvae producing mines were recorded after 1, 2, 6, and 22 h from the beginning of the test.
Experiment 4: repellency effects against larvae in choice experiments
EOs of laurel, Spanish oregano, peppermint, cypress, eucalyptus, and basil were tested. Treatments were prepared as described in the previous section (experiment-3). Bioassays were performed as follows: single pairs of leaflets, one for the test and one for the control, were randomly placed at a distance of 2 cm from each other in a Petri dish (9 cm diameter). For each treatment, five Petri dishes were prepared and one T. absoluta third instar larva was put into each Petri dish in the central area between leaflets. Five replicates were run for each tested oil under the same climatic conditions described previously. The presence or absence of larvae producing mines on the leaflets of each Petri dish was recorded after half an hour, and 5, 20, and 24 h from the beginning of the experiment.
Statistical analysis
To assess the effects of EOs on the oviposition deterrence, data were analysed using paired t-test (for dual-choice tests between test and control) in experiment-1 (laboratory). A one-way ANOVA followed by Tukey's post-hoc test (P < 0.05) was used for experiment-2 (greenhouse). To evaluate repellency effects, two-way ANOVA followed by Tukey's post-hoc test (P < 0.05) was used for experiment-3 (no-choice test), in addition to paired t-test for experiment-4 (dual-choice test). Before analysis, a logarithmic transformation of the average values and arcsine transformation of the percentage values were carried out. Percentage repellency (PR) was calculated according to McDonald formula (McDonald et al. 1970) as follows:
\(PR = [(Nc-Nt) / (Nc + Nt)] 100\)
Where: Nc: Number of larvae in the control.
Nt: number of larvae in the treated leaves.
Statistical analyses were performed using Statistica 7.0 for Windows Package (Stat Soft Inc. 2001).
Results
Experiment 1: oviposition deterrent effects in laboratory
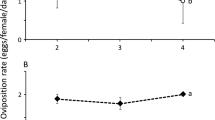
The oviposition deterrent effect after 36 h from the treatment was remarkable for both Spanish oregano (4 μl/ml) and laurel (4 μl/ml) EOs (Fig. 1). The average number of eggs laid on leaves treated with Spanish oregano and laurel EOs and control is shown in Table 1. Statistical analysis (paired t-test) showed that differences between test and control were significant for both Spanish oregano (t = -7.75, df = 78, P < 0.001) and laurel (t = -4.29, df = 78, P < 0.001) EOs, but not significant between these two EOs (t = -0.71, df = 78, P = 0.48).

Percentage of eggs laid by mated females of T. absoluta subjected to dual-choice test on tomato leaves treated with laurel and Spanish oregano EOs (test) or distilled water (control) after 36 h from the beginning of the experiment in laboratory
Experiment 2: oviposition deterrent effects in greenhouse
The observations after 4 days from the beginning of the experiment showed oviposition deterrent effects of Spanish oregano (4 μl/ml), basil (4 μl/ml), and garlic at the dose 6 μl/ml, but not of laurel (4 μl/ml), and garlic at the dose 3 μl/ml (Fig. 2). The average number of eggs per plants for each EO is shown in Table 2. Statistical analysis (paired t-test) indicated significant differences in mean number of eggs between test and control in trials with EOs of Spanish oregano (t = -7.72, df = 38, P < 0.001), basil (t = -4.52, df = 38, P < 0.001), and garlic at the dose 6 μl/ml (t = -2.44, df = 38, P = 0.019). On the contrary, non-significant differences were found in trials with EOs of laurel (t = -0.36, df = 38, P = 0.71) and garlic at the dose 3 μl/ml (t = -1.15, df = 38, P = 0.25). Significant differences were found among EOs (one-way ANOVA: F = 11.30, df = 4, P < 0.001), due to EO of garlic at the dose 3 μl/ml, that differed from the other EOs, and EO of garlic (6 μl/ml) that differed from laurel EO only (Tukey post hoc, P = 0.005).

Percentage of eggs laid by mated females of T. absoluta subjected to dual-choice test on tomato plants treated with Spanish oregano, basil, laurel, and garlic EOs (test) or distilled water (control) detected after 4 days from the beginning of each EO experiment in greenhouse
Experiment 3: repellency effects against larvae in no-choice test
The percentage of larvae detected on the leaflets treated with EOs (4 μl/ml) and control at 1, 2, 6 and 22 h after releasing the larvae are shown in Fig. 3. In the first observation, larvae were only detected on cypress EO and control, at 2 h on eucalyptus, at 6 h on Spanish oregano, and at 22 h on peppermint. In the last observation, a higher percentage of larvae was observed in the control, followed by cypress, eucalyptus, oregano, and peppermint. The percentage of repellency (PR) of EOs detected after releasing the larvae on the treated leaflets is shown in Table 3. All EOs elicited a strong repellence at 2 h from the treatment. PR of peppermint was always high, while decreased that of the other EOs, with a small increasing at 22 h. Statistical analysis (two-way ANOVA) indicated that there was a significant difference in percentage of larvae detected among EOs (F = 6.89, df = 12, P < 0.001). In all experiments, tests with EOs were statistically different from control, except for cypress EO at 6 and 22 h, and eucalyptus EO at 22 h. In addition, statistical differences were found at 22 h, specifically among peppermint EO and other EOs except Spanish oregano EO, and between Spanish oregano EO and eucalyptus EO. Considering time effects of each EO, significant differences were observed after 6 h for cypress and eucalyptus EOs, while no differences were detected for all EOs, except for peppermint EO, where larvae were detected on the test at 22 h (Tukey post hoc, P = 0.005).

Percentage of T. absoluta larvae on tomato leaf associated with EOs and control detected after 1, 2, 6 and 22 h from the beginning of the experiment in no-choice test (bars with the same letter are not significantly different (Tukey post hoc P < 0.05)
Experiment 4: repellency effects against larvae in choice test
The response of larvae in dual-choice treatments with EOs (4 μl/ml) is shown in Fig. 4. After 5 h, all larvae in bioassays with Spanish oregano, cypress and eucalyptus EOs, made a choice. In bioassays with basil, peppermint, and laurel EOs, larvae made a choice in 60%, 75%, and 80% of the cases respectively, while the remaining larvae exhibited a no choice. The low preference of the larvae for the test over control was observed for all EOs except basil. No larval damage was ever detected in tests with Spanish oregano and laurel EOs. After 20 h, all larvae eventually made a choice. In particular, in the treatment with basil, larvae moved towards the test in 50% of the replicates, in the treatments with peppermint and laurel EOs all larvae preferred the control condition (100%). PR of EOs detected after releasing the larvae on the treated leaflets is shown in Table 4. At 5 h from the treatment, oregano and laurel EOs showed a total repellence effect, while the others EOs showed values not exceeding 30%. PR did not change at 20 h, except for peppermint where an increase up to 50% was found. Statistical analysis (paired t-test) highlighted significant differences between test and control in Spanish oregano (t = -4.0, df = 48, P < 0.001) and laurel (t = -6, df = 48, P < 0.001) EOs at either 5 or 20 h after treatment, and in peppermint (t = -4.52, df = 48, P < 0.001) at 20 h. No significant differences for cypress, basil and eucalyptus EOs were found (P < 0.05). In addition, two-ways ANOVA indicated that there were significant differences among EOs at either 5 or 20 h after treatment (F = 9.07, df = 5, P < 0.001), due to peppermint EO and Spanish oregano EOs (Tukey post hoc, P < 0.05). No significant differences between observation were found over time (F = 1.74, df = 1, P = 0.19).

Percentage of T. absoluta larvae on tomato leaves associated with EOs and control detected after 5 (a) and 20 h (b) from the beginning of the experiment in choice test (asterisks indicate a preference which is significantly different within a choice test, P < 0.001, n = 25, df = 48, paired t-test analysis)
Discussion
Natural products of plants, especially extracts or essential oils, are traditionally used against economically important pests, and some of them have provided potential alternatives to currently used synthetic insecticides (Isman 2006). These natural products are less harmful to the environment and human health and their utilization has increased in recent years. Despite the large number of essential oils used against pest insects, only few have been tested on the tomato borer (Soares et al. 2019). Our study investigated the effectiveness in terms of oviposition deterrence and repellency of EOs of O. basilicum (basil), T. capitatus, subgenus C. capitatus (Spanish oregano), L. nobilis (laurel), M. piperita (peppermint), C. sempervirens (cypress), E. globulus (eucalyptus), and A. sativum (garlic) against T. absoluta. In relation to the oviposition deterrence, the control of T. absoluta by preventing egg deposition on plant tissues is a potentially very attractive approach in order to avoid increasing pest population on tomato fields. However, there is still little knowledge on the activity of natural products about this aspect. A literature survey revealed that some works on the oviposition deterrence of T. absoluta have been conducted using aqueous extracts of Nerium oleander L., Olea europea L., Laurus nobilis L. (Natour and Karrom 2017), EOs of Azadirachta indica A. Juss (Tomé et al. 2013), Tanacetum vulgare L., T. parthenium L., Aleo vera (L), Melaleuca alternifolia (Maiden and Betce) and Juglans regia L. (Erdogan 2019) in laboratory, EOs of Ocimum gratissimum L. and O. basilicum L. (Yarou et al. 2017) in flight tunnels, and aqueous extracts of Allium sativum L. and Cymbopogon citratus (DC.), EOs of Eucalyptus spp., Ruta graveolens L., O. basilicum and Pimpinella anisum L. (Hussein et al. 2015) in field conditions. This paper is the first record on the oviposition deterrence effects by EOs of Spanish oregano and laurel against T. absoluta, which highlighted a strong effectiveness of this EOs in laboratory. In greenhouse conditions, a similar deterrent activity was obtained with Spanish oregano followed by basil and garlic at dose of 6 μl/ml, but no significant effects were observed with laurel and garlic at dose of 3 μl/ml. Taken together, these results indicate that deterrence caused by garlic increased with dosage. Results about basil are in agreement with other works reporting that basil oil reduces the population of T. absoluta per plant up to 63% in field (Hussein et al. 2015) and caused significant ovipositional deterrent effects in a flight tunnel (Yarou et al. 2017). Moreover, our results confirm those of previous studies on other pests. For instance, basil reduced the oviposition behaviour of Spodoptera littoralis (Boisduval) (Abd El Aziz and El Hawary 1997), Phthorimaea operculella Zell. (Sharaby et al. 2009), Callosobruchus chinensis L. (Kiradoo and Srivastava 2010) and Aedes aegypti L. (Warikoo et al. 2011).
In relation to repellent effects by EOs on T. absoluta, some recent studies have been conducted under laboratory conditions using EOs of O. gratissimum and O. kilimandscharicum Guerke against adults (Essoung et al. 2020), and Cinnamomum zeylanicum Blume (Adil et al. 2015), A. sativum, Syzygium aromaticum, Eucalyptus spp., M. piperita, Simmondsia chinensis, Artemisia absinthium (Salama and Shehata 2017) and Lantana camara L. (Liambila et al. 2021) against larvae. In our study EOs of Spanish oregano, laurel, basil, cypress, and eucalyptus have been tested as repellents against T. absoluta for the first time. The results obtained provide a scientific contribution for the use of EOs to control of this pest. Peppermint, Spanish oregano and laurel EOs showed a strong efficacy while cypress, eucalyptus, and basil EOs were moderately repellent. In no-choice tests, repellent effects of EOs were affected by the time after treatments, showing variable responses from test larvae. In particular, the major repellency was obtained in experiments with Spanish oregano and peppermint EOs, where larvae moved on test after 6 and 22 h after treatment, respectively, and differences from the control were significant in all observations. Cypress and eucalyptus EOs were less repellent than other EOs, considering larval presence occurred on the test leaves 1 and 2 h after treatment application, respectively. The repellent activity of these two EOs decreased when time after treatment application increased, because no significant differences from the control were recorded from 6 and 22 h, respectively. The major percentage of repellency (PR) was detected with peppermint EO in all observations, pointing out its potential effectiveness in the control of T. absoluta. These results are in agreement with a previous study in which peppermint EO was highly repellent against first instar larvae of T. absoluta in laboratory (Salama and Shehata 2017). Many studies on this essential oil demonstrate deleterious effects on a wide range of insects. Indeed, peppermint EO was reported having repellent effects of ants, flies, lice, beetles and moths (Ansari et al. 2000; Hori 2003; Hussein et al. 2015; Saeidi and Mirfakhraie 2017). Our experiments on repellency under choice conditions showed variable responses from larvae in dependence on the EO tested, regardless of the time elapsed after treatments, with the exception of peppermint EO. Indeed, after 20 h, this EO caused some degree of repellency, because significant differences from the control were recorded, with PR up to 50%. The most promising effects were observed in Spanish oregano and laurel EOs treatments, where larvae were never detected in the test and differences from the control were always significant. EOs of basil, cypress and eucalyptus showed a lower efficacy in terms of larval response which never differed significantly from the control and PR levels were around 30% in both time observations. It is revealed by comparing the types of experiments (no-choice and choice test), that the repellent effectiveness of EOs does not seem to be influenced when larvae did not get a chance to make a choice between treated and untreated leaves.
The positive effects of repellency by Spanish oregano and laurel EOs against tomato pinworm observed in our work are in accordance with studies that tested these plant extracts against stored product pests (Papachristos and Stamopoulos 2002; Cosimi et al. 2009; Jemâa et al. 2011; Ncibi et al. 2019). EOs of basil, cypress and eucalyptus were weak repellent against T. absoluta and the lower efficacy detected in our study could due to the concentration used. Indeed, in contrast with our results, these EOs are reported as good repellents against many harmful pests such as stored product insects, moths, mosquitoes, flies and aphids (Landolt et al. 1999; Harrewijn et al. 2001; Papachristos and Stamopoulos 2002; Yang and Ma 2005; Prajapati et al. 2005; Tapondjou et al. 2005; Erler et al. 2006; Hori 2003; Mishra et al. 2012; Hasaballah et al. 2018; Madreseh-Ghahfarokhi et al. 2018; Chandel et al. 2019; Ncibi et al. 2019; Abdelkader et al. 2020). However, the low repellent activity of eucalyptus was found also against moths and mosquitoes (Landolt et al. 1999; Trongtokit et al. 2005). Overall, the results of this research indicate that EOs from plants such as Spanish oregano, basil, laurel, peppermint and garlic could be potentially implemented as biopesticides against T. absoluta. Further study using cypress and eucalyptus EOs should be carried out in order to evaluate their potential use against this pest.
Conclusions
EOs used in this study gave promising results against T. absoluta. The oviposition deterrence effect of laurel, oregano, basil and garlic EOs was remarkable both under laboratory and greenhouse conditions. Repellence tests against larvae highlighted that peppermint, Spanish oregano and laurel EOs had a strong efficacy, while cypress, eucalyptus and basil EOs displayed moderately repellent effects. To conclude, the results obtained in this work could be useful to develop pest control programs against T. absoluta but further studies under field conditions are needed to confirm oviposition deterrence and repellence effects.
References
Abd El-Aziz SE, El-Hawary FMA (1997) Inhibitory effects of some essential oils on the development of the cotton leafworm, Spodoptera littoralis (Boisd.). J Egypt German Soc Zool 22:117–130
Abdelkader H, Abdelkader B, Yahia B (2020) Toxicity and repellency of Eucalyptus globulus L. essential oil against Aphis fabae Scopoli, 1973 (Homoptera: Aphididae). J Ent Res 44(1):147–152. https://doi.org/10.5958/0974-4576.2020.00027.4
Adil B, Tarik A, Abderrahim K, Khadija O (2015) Evaluation of the insecticidal effect of the essential oil of Cinnamomum zeylanicum against Tuta absoluta (Meyrick). Inter J Innov Res Sci Eng Tech 4(9):8033–8037. https://doi.org/10.15680/IJIRSET.2015.0409005
Alam M J, Ahmed KS, Rony MNH, Islam NET, Bilkis SE (2019) Bio-efficacy of bio-pesticides against tomato leaf miner, Tuta absoluta, a threatening pest of tomato. Biosci Agric Res 22(02):1852–1862 https://doi.org/10.18801/jbar.220219.229
Andrade-Ochoa S, Sánchez-Aldana D, Chacón-Vargas KF, Rivera-Chavira BE, Sánchez-Torres LE, Camacho AD, Nogueda-Torres B, Nevárez-Moorillón GV (2018) Oviposition deterrent and larvicidal and pupicidal activity of seven essential oils and their major components against Culex quinquefasciatus Say (Diptera: Culicidae): synergism–antagonism effects. Insects 9:25. https://doi.org/10.3390/insects9010025
Ansari MA, Vasudevan P, Tandon M, Razdan RK (2000) Larvicidal and mosquito repellent action of peppermint (Mentha piperita) oil. Biores Technol 71:267–271. https://doi.org/10.1016/S0960-8524(99)00079-6
Aynalem B (2018) Tomato leafminer [(Tuta absoluta Meyrick) (Lepidoptera: Gelechiidae)] and its current eco-friendly management strategies: A review. J Agric Biotechnol Sustain Dev 10(2):11–24. https://doi.org/10.5897/JABSD2018.0306
Bett PK, Deng AL, Ogendo JO, Kariukia ST et al (2017) Residual contact toxicity and repellence of Cupressus lusitanica Miller and Eucalyptus saligna Smith essential oils against major stored product insect pests. Ind Crop Prod 110:65–74. https://doi.org/10.1016/j.indcrop.2017.09.046
Biondi A, Guedes RNC, Wan F-H, Desneux N (2018) Ecology, worldwide spread, and management of the invasive South American tomato pinworm, Tuta absoluta: past, present, and future. Ann Rev Ent 63:239–258. https://doi.org/10.1146/annurev-ento-031616-034933
Campos MR, Biondi A, Adiga A, Guedes RNC, Desneux N (2017) From the Western Palaearctic region to beyond: Tuta absoluta 10 years after invading Europe. J Pest Sci 90:787–796. https://doi.org/10.1007/s10340-017-0867-7
Chandel RK, Nebapure SM, Sharma M, Subramanian S et al (2019) Insecticidal and repellent activities of Eucalyptus oil against lesser grain borer Rhyzopertha Dominica (Fabricius). J Microbiol Biotech Food Sci 9(3):525–529. https://doi.org/10.15414/jmbfs.2019/20.9.3.525-529
Chintalchere JM, Dar MA, Raut KD, Pandit RS (2021) Bioefficacy of lemongrass and tea tree essential oils against house fly, Musca domestica. Proc Natl Acad Sci, India, Sect B Biol Sci 91(2):307–318. https://doi.org/10.1007/s40011-020-01220
Cosimi S, Rossi E, Cioni PL, Canale A (2009) Bioactivity and qualitative analysis of some essential oils from Mediterranean plants against stored-product pests: Evaluation of repellency against Sitophilus zeamais Motschulsky, Cryptolestes ferrugineus (Stephens) and Tenebrio molitor (L). J Stored Prod Res 45:125–132. https://doi.org/10.1016/j.jspr.2008.10.002
Desneux N, Luna MG, Guillemaud T, Urbaneja A (2011) The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: the new threat to tomato world production. J Pest Sci 84:403–408. https://doi.org/10.1007/s10340-011-0398-6
Desneux N, Wajnberg E, Wyckhuys KAG, Burgio G et al (2010) Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. J Pest Sci 83:197–215. https://doi.org/10.1007/s10340-010-0321-6
Erdogan P (2019) Oviposition deterrent activities of some plant extracts against tomato leaf miner, Tuta Absoluta Meyrick (Lepidoptera: Gelechiidae). J Bacter Myc Open Access 7(6):139‒142. https://doi.org/10.15406/jbmoa.2019.07.00259
Erler F, Ulug I, Yalcinkaya B (2006) Repellent activity of five essential oils against Culex pipiens. Fitoterapia 77:491–494
Essoung FRE, Tadjong AT, Chhbra SC, Mohamed SA, Hassanali A (2020) Repellence and fumigant toxicity of essential oils of Ocimum gratissimum and Ocimum kilimandscharicum on Tuta absoluta (Lepidoptera: Gelechiidae). Environ Sci Poll Res 27:37963–37976
Estay P (2000) Polilla del Tomate Tuta absoluta (Meyrick). Impresos CGS Ltda. http://www.inia.cl/medios/Descargas/CRI/Platina/Informativos/Informativo9.pdf
Esther MR, Emmanuel NH, George BG, Bintu NN et al (2019) Controlling Tuta absoluta (Meyrick) by selected crude plant extracts in the laboratory and in the screen house. J Agric Sci Tech A 9:227–239. https://doi.org/10.17265/2161-6256/2019.04.003
Guedes RNC, Roditakis E, Campos MR, Haddi K, Bielza P, Siqueira HAA, Tsagkarakou A, Vontas J, Nauen R (2019) Insecticide resistance in the tomato pinworm Tuta absoluta: patterns, spread, mechanisms, management and outlook. J Pest Sci 92:1329–1342
Han P, Zhang Y, Lu Z, Wang S, Ma D, Biondi A, Desneux N (2018) Are we ready for the invasion of Tuta absoluta? Unanswered key questions for elaborating an Integrated Pest Management package in Xinjiang, China. Entomol Gen 38:113–125
Han P, Bayram Y, Shaltiel-Harpaz L, Sohrabi F, Saji A et al (2019) Tuta absoluta continues to disperse in Asia: Damage, ongoing management and future challenges. J Pest Sci 92:1317–1327
Harrewijn P, van Oosten AM, Piron PG (2001) Natural terpenoids as messengers: a multidisciplinary study of their production, biological functions, and practical applications. Kluwer Academic Publishers, The Netherlands
Hasaballah AI, Shehata AZ, Fouda MA, Hassan MI, Gad ME (2018) The biological activity of Cupressus sempervirens extracts against Musca domestica. Asian J Biol 5(1):1–12. https://doi.org/10.9734/AJOB/2018/38023
Hori M (2003) Repellency of essential oils against the cigarette beetle, Lasioderma serricorne (Fabricius) (Coleoptera: Anobiidae). Appl Entomol Zool 38:467–473. https://doi.org/10.1303/aez.2003.467
Hussein NM, Hussein MI, Gadel Hak SH, Shaalan HS, Hammad MA (2015) Effect of two plant extracts and four aromatic oils on Tuta absoluta population and productivity of tomato cultivar gold stone. Nat Sci 12:108–118. https://doi.org/10.21608/jppp.2015.74529
Isman MB (2006) Botanical insecticides, deterrents and repellents in modern agriculture and an increasingly regulated world. Annu Rev Entomol 51:45–66. https://doi.org/10.1146/annurev.ento.51.110104.151146
Jemâa JMB, Tersim N, Khouja ML (2011) Composition and repellent efficacy of essential oil from Laurus nobilis against adults of the Cigarette beetle Lasioderma serricorne (Coleoptera: Anobiidae). Tunisian J Plant Prot 6(1):29–42
Kiradoo MM, Srivastava MA (2010) Comparative study on the efficacy of two lamiaceae plants on egg - laying performance by the pulse beetle Callosobruchus chinensis Linn. (Coleoptera : Bruchidae). J Biopest 3(3):590–595
Landolt PJ, Hofstetter RW, Biddick LL (1999) Plant essential oils as arrestants and repellents for neonate larvae of the codling moth (Lepidoptera: Tortricidae). Environ Entomol 28(6):954–960. https://doi.org/10.1093/ee/28.6.954
Liambila RN, Wesonga JM, Ngamau CN, Wallyambillah W (2021) Chemical composition and bioactivity of Lantana camara L. essential oils from diverse climatic zones of Kenya against leaf miner (Tuta absoluta Meyrick). Afr J Agric Res 17(9):1198–1208. https://doi.org/10.5897/AJAR2020.15243
Lo Pinto M, Vella L, Agrò A (2019) Investigations on Tuta absoluta (Lepidoptera: Gelechiidae): larval infestation on the tomato cultivated in open field and evaluation of five essential oils against larvae in laboratory. Int J Ent Res 4(4):7–14 www.entomologyjournals.com
Lo Pinto M, Vella L, Agrò A (2020) Adulticidal activity of essential oils of Mentha piperita L., Cupressus sempervirens L., and Eucalyptus globulus Labill. against the tomato leafminer Tuta absoluta Meyrick (Lepidoptera: Gelechiidae). J Ent Zool St 8(6):1721–1728. https://www.entomologyjournals.com
Madreseh-Ghahfarokhi S, Pirali Y, Dehghani-Samani A, Dehghani-Samani A (2018) The insecticidal and repellent activity of ginger (Zingiber officinale) and eucalyptus (Eucalyptus globulus) essential oils against Culex theileri Theobald, 1903 (Diptera: Culicidae). Ann Parasitol 64(4):351–360. https://doi.org/10.17420/ap6404.171
Magalhães STV, Jham GN, Picanço MC, Magalhães G (2001) Mortality of second-instar larvae of Tuta absoluta produced by the hexane extract of Lycopersicon hirsutum f. glabratum (PI 134417) leaves. Agr for Ent 3:297–303. https://doi.org/10.1046/j.1461-9555.2001.00116.x
Mansour R, Brevault T, Chailleux A, Cherif A, Grissa-Lebdi K, Haddi K, Mohamed SA, Nofemela RS, Oke A, Sylla S, Tonnang HEZ, Zappalà L, Kenis M, Desneux N, Biondi A (2018) Occurrence, biology, natural enemies and management of Tuta absoluta in Africa. Entomol Gen 38:83–112
Mansour R, Biondi A (2021) Releasing natural enemies and applying microbial and botanical pesticides for managing Tuta absoluta in the MENA region. Phytoparasitica 49:179–194
Marsin AM, Muhamad II, Anis SNS, Lazim NAM, Ching LW, Dolhaji NH (2020) Essential oils as insect repellent agents in food packaging: a review. Eur Food Res Technol 246:1519–1532. https://doi.org/10.1007/s00217-020-03511-1
McDonald LL, Guy RH, Speirs RD (1970) Preliminary evaluation of new candidate materials as toxicants, repellents, and attractants against storedproduct insects-1. Preliminary evaluation of new candidate materials as toxicants, repellents, and attractants against stored-product insects. Marketing Res Rep Agric Res Serv USDA 1(882):35
Mishra BB, Tripathi SP, Tripathi CPM (2012) Repellent effect of leaves essential oils from Eucalyptus globulus (Mirtaceae) and Ocimum basilicum (Lamiaceae) against two major stored grain insect pests of Coleopterons. Nat Sci10(2):50–54. http://www.sciencepub.net
Mossa ATH (2016) Green pesticides: essential oils as biopesticides in insect-pest management. J Environ Sci Technol 9:354–378. https://doi.org/10.3923/jest.2016.354.378
Natour M, Karrom M (2017) The effect study of oviposition deterrent of some plant aqueous extracts of moth tomato females Tuta absoluta (Meyrick), (Lepidoptera: Gelechiidae). J King Abdulaziz Univ - Sci 29(2):77–85
Ncibi S, Naima B, Soumaya HH, Ncibi MA et al (2019) Insecticidal activity of several Tunisian essential oils against two major pests of stored grain Rhyzopertha dominica (Fabricius) and Tribolium castaneum (Herbest). J New Sci Agr Biotechnol 66(5):4182–4194. https://www.jnsciences.org
Nerio LS, Olivero-Verbel J, Stashenko E (2010) Repellent activity of essential oils: A review. Bioresour Technol 101:372–378. https://doi.org/10.1016/j.biortech.2009.07.048
Papachristos DP, Stamopoulos DC (2002) Repellent, toxic and reproduction inhibitory effects of essential oil vapours on Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J Stored Prod Res 38:117–128. https://doi.org/10.1016/S0022-474X(01)00007-8
Patil R, Ravindra R (2008) Formulation development and evaluation from garlic oil macerate. The Internet J Nutr Welln 8(1). https://doi.org/10.5580/25c0
Prajapati V, Tripathi AK, Aggarwal KK, Khanuja SPS (2005) Insecticidal, repellent and oviposition-deterrent activity of selected essential oils against Anopheles stephensi, Aedes aegypti and Culex quinquefasciatus. Bioresour Technol 96:1749–1757. https://doi.org/10.1016/j.biortech.2005.01.007
Rajkumar V, Gunasekaran C, Christy IK, Dharmaraja J et al (2019) Toxicity, antifeedant and biochemical efficacy of Mentha piperita L. essential oil and their major constituents against stored grain pest. Pest Biochem Physiol 156:138–144. https://doi.org/10.1016/j.pestbp.2019.02.016
Saeidi K, Mirfakhraie S (2017) Chemical composition and insecticidal activity Mentha piperita L. essential oil against the cowpea seed beetle Callosobruchus maculatus F. (Coleoptera: Bruchidae). J Entomol Acarol Res 49:127–134. https://doi.org/10.4081/jear.2017.6769
Said-Al Ahl H, Hikal WM, Tkachenko KG (2017) Essential oils with potential as insecticidal agents: a review. J Environ Plann Man 3:23–33
Salama HS, Shehata IE (2017) Bioactivity and repellency of some plant extracts against the Tomato leaf miner Tuta absoluta (Meyrick 1917) (Lepidoptera: Gelechiidae). Res J Pharm Bio Chem Sci 8(4):1021–1036
Sharaby A, Abdel-Rahman H, Moawad S (2009) Biological effects of some natural and chemical compounds on the potato tuber moth, Phthorimaea operculella Zell. (Lepidoptera:Gelechiidae). Saudi J Biol Sci 16:1–9. https://doi.org/10.1016/j.sjbs.2009.07.001
Soares MA, Campos MR, Passos LC, Carvalho GA, Haro MM, Lavoir AV, Biondi A, Zappalà L, Desneux N (2019) Botanical insecticide and natural enemies: A potential combination for pest management against Tuta absoluta. J Pest Sc 92:1433–1443
Speranza S, Sannino L (2012) The current status of Tuta absoluta in Italy. Bulletin OEPP/EPPO Bulletin 42(2):328–332. https://doi.org/10.1111/epp.2579
Statsoft Inc Statistica (Data Analysis Software System) (2001) Version 6 StatSoft Italia S.r.l, Vigonza (PD)
Tapondjou AL, Adler C, Fontem DA, Bouda H, Reichmuth C (2005) Bioactivities of cymol and essential oils of Cupressus sempervirens and Eucalyptus saligna against Sitophilus zeamais Motschulsky and Tribolium confusum du Val. J Stored Prod Res 41(1):91–102. https://doi.org/10.1016/j.jspr.2004.01.004
Tarusikirwa VL, Machekano H, Mutamiswa R, Chidawanyika F, Nyamukondiwa C (2020) Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) on the “Offensive” in Africa: prospects for integrated management initiatives. Insects 11:764. https://doi.org/10.3390/insects11110764
Tomé HVV, Martins JC, Corrêa AS, Galdino TVS et al (2013) Azadirachtin avoidance by larvae and adult females of the tomato leafminer Tuta absoluta. Crop Prot 46:63–69. https://doi.org/10.1016/j.cropro.2012.12.021
Trongtokit Y, Rongsriyam Y, Komalamisra N, Apiwathnasorn C (2005) Comparative repellency of 38 essential oils against mosquito bites. Phytother Res 19:303–309. https://doi.org/10.1002/ptr.1637
Uchôa-Fernandes MA, Della Lucia TMC, Vilela EF (1995) Mating, oviposition and pupation of Scrobipalpuloides absoluta (Meyr.) (Lepidoptera: Gelechiidae), An Soc Entomol Brasil 24(1):159–164
Umpiérrez ML, Lagreca ME, Cabrera R, Grille G, Rossini C (2012) Essential oils from Asteraceae as potential biocontrol tools for tomato pests and diseases. Phytochem Rev 11:339–350. https://doi.org/10.1007/s11101-012-9253-5
Urbaneja A, Desneux N, Gabarra R, Arno J et al (2013) Biology, ecology and management of the tomato borer, Tuta absoluta. In: Peña JE (ed) Potential Invasive Pests of Agricultural Crops. Wallingford: CABI pp 98–132. https://doi.org/10.1079/9781845938291.0098
Warikoo R, Wahab N, Kumar S (2011) Oviposition-altering and ovicidal potentials of five essential oils against female adults of the dengue vector, Aedes aegypti L. Parasitol Res 109:1125–1131. https://doi.org/10.1007/s00436-011-2355-y
Yang P, Ma Y (2005) Repellent effect of plant essential oils against Aedes albopictus. J Vector Ecol 30:231–234
Yarou BB, Bawin T, Boullis A, Heukin S et al (2017) Oviposition deterrent activity of basil plants and their essentials oils against Tuta absoluta (Lepidoptera: Gelechiidae). Environ Sci Pollut Res 25:29880–29888. https://doi.org/10.1007/s11356-017-9795-6
Funding
Open access funding provided by Università degli Studi di Palermo within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest:
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lo Pinto, M., Vella, L. & Agrò, A. Oviposition deterrence and repellent activities of selected essential oils against Tuta absoluta Meyrick (Lepidoptera: Gelechiidae): laboratory and greenhouse investigations. Int J Trop Insect Sci 42, 3455–3464 (2022). https://doi.org/10.1007/s42690-022-00867-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42690-022-00867-7