
Abstract
The use of plants as sources for novel antimicrobial as well as antioxidant agents offers advantages. Plants are readily accessible and inexpensive, extracts or compounds from plant sources often demonstrate high level of biological activities. Previous studies have reported antibacterial and antifungal activities within the Fabaceae family that included Acacia species. This study aims to determine presence of antibacterial activity, antioxidant activity, and the secondary metabolites of sequential solvent extracts (acetone, methanol, and acetic acid) of Acacia berlandieri and Acacia rigidula leaves. The antibacterial activity was investigated using a disc diffusion assay. The ferric thiocyanate method was used to assess the ability of all extracts to prevent oxidation. Qualitative phytochemical tests, NMR, IR, and UV–Vis spectroscopy were done to identify potential secondary metabolites. P. alcalifaciens (p < 0.001), E. faecalis (p < 0.01), S. aureus (p < 0.001), and Y. enterocolitica (p < 0.001) were significantly inhibited by A. rigidula extracts when compared to A. berlandieri extracts. A. rigidula’s acetone extract exhibited the significantly (p < 0.001) highest inhibition of peroxidation, 42%. Qualitative phytochemical tests showed positive results for presence of phenols, flavonoids, saponins, terpenes and tannins. NMR, IR, and UV–Vis spectroscopy revealed chemical structures found in flavonoids, saponins, terpenes and tannins, supporting the results of qualitative phytochemical tests. A. berlandieri and A. rigidula leaf extracts have revealed presence of medicinally valued bioactive components. The results of this study provide a basis for further investigations of the A. rigidula leaf extracts. A. rigidula leaf extracts have the potential to serve as a source of novel antimicrobial and antioxidant agents.
Graphic abstract

Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
People have used various plants and their derivatives for medical purposes, including the treatment of infectious diseases. The use of plants as sources for novel antimicrobial as well as antioxidant agents offers advantages. Plants are readily accessible and inexpensive, extracts or compounds from plant sources often demonstrate high-level activity against bacteria, and they rarely have severe side effects [1]. Most pharmacological activities of medicinal plants are traced to their secondary metabolites (SMs). SMs are smaller molecules when compared to the constituents of primary metabolites (PMs) like proteins, carbohydrates, and lipids [2]. SMs, unlike PMs are not directly involved in plants’ normal growth or reproduction. SMs on the other hand, provide the plants with defense mechanisms and/or survival mechanisms. SMs are also known to be useful to humans. For example, they are a major source of antibiotics, food additives i.e., antioxidants, and pesticides [3]. Biosynthesized compounds derived from SMs that include alkaloids, phenols, essential oils, steroids, lignins, and tannins have been successful source of potential drug leads [4].
Currently, there are ~ 20,000 species of known medicinally valuable plants; which can serve as a valuable reservoir of SMs for the development of novel drugs. Likewise, only an estimated 10% of the world’s biodiversity has been evaluated for this purpose [5]. Fabaceae is the second largest family currently used for medicinal plant purposes with over 490 plant species [6]. A larger diversity of species suggests a greater supply of raw materials for drug discovery and biotechnology [7]. Studies have highlighted biological activities within the Fabaceae family [3, 8,9,10,11]. Acacia species belong to this family. Several Acacia species were reported to have antibacterial and antifungal, antioxidant, anticancer, antiparasitic, cytotoxicity and immunomodulatory activities [12]. Specific examples include Acacia nilotica and Acacia aroma; both are used in traditional medicine. A. nilotica has been reported to have antimicrobial, antiplasmodial and antioxidant activities. It is also used for the treatment of human immunodeficiency virus, hepatitis C virus and cancer [13, 14]; while, A. aroma is used for wound healing, as antiseptic, and the treatment of gastrointestinal disorders [15].
Plants containing phenolic compounds are possible sources of natural antioxidants that stabilize free readicals by hydrogenation or complexing with oxidizing species [16]. Members of the Fabaceae family, A. nilotica and A. hydaspica, are rich in phenolic compounds and are reported to have inhibitory effects on bacterial pathogens [16, 17]. A. nilotica leaf extracts exhibit reducing capabilities (2.57 ± 0.03) similar to ascorbic acid (2.62 ± 0.07) at 1,000 ug/mL confering to the reducing potential of the species [17]. Acetone extracts of A. senegal and A. dealbata were reported to have high concentrations of phenols and flavonoids and exhibited antioxidant activities [18, 19]. Antioxidants are known to eliminate reactive oxygen species (ROS) involved in human diseases. ROS have harmful effects on DNA, lipid peroxidation, and oxidative deactiviation of proteins associated in pathogenesis of many human diseases, such as cancer, Alzheimer’s disease and atherosclerosis [20].
The species under study, A. rigidula and A. berlandieri are predominantly found in the southern part of Texas and in the northern states of Mexico [21]. To our knowledge, there is very limited studies on the biological activities of the two species understudy. A. rigidula extracts have been extensively used in popular weight-loss supplements [21]. In addition, Garza et al., reported antifungal properties of A. rigidula methanol extracts [22]. More recently, A. rigidula’s root and stem extracts have been used as reducing and capping agent to produce silver nanoparticles that eradicate pathogenic resistant bacteria in vivo [23].
To date there is no report of A. rigidula nor A. berlandieri being used in traditional medicine. Likewise, although the Acacia genus is quite large, little is known about the chemical composition of most species in this genus. From the several species that have been studied, flavonoids, terpenoids and phenolic acids are the most predominant compounds isolated from the genus Acacia. The most frequently reported flavonoids are catechin and quercetin. Triterpenoids are the major class of terpenoids and many gallic acid and coumaric acid derivatives were reported to have been isolated from the genus Acacia [12]. It was also reported that the most evident secondary metabolites present in the Acacia species are polysaccharides (gums) and complex phenolic substances (condensed tannins) [24]. To our knowledge, our study is the first to report on the potential secondary metabolites in A. rigidula and A. berlandieri.
Considering the biological properties reported from other Acacia species, it is of interest and relevant to further study the biological activities and secondary metabolites found in A. rigidula and A. berlandieri. Results of such research study could provide a novel source for the development of effective treatments. The medicinal value and the pattern of global ethnomedical knowledge will broaden as more species within this family are further evaluated [25,26,27,28,29]. Therefore, the objectives of this study are (1) to determine presence of antibacterial, and antioxidant activities in A. rigidula. and A. berlandieri leaf extracts and (2) to identify the potential secondary metabolites in the extract that exhibited the most significant activity.
2 Results and discussion
2.1 2.1 Antibacterial activity
Three leaf extracts (acetone, methanol, and acetic acid) per species (A. rigidula and A. berlandieri) were subjected to antibacterial assays against nine different bacteria where the mean zones of inhibition (mZOI) were measured (Table 1).
Both Acacia species exhibited differences in antibacterial activities based on mZOIs. A. berlandieri displayed minimal inhibitory effects (mZOI, 6.00–8.99 mm) against three (P. alcalifaciens, P. aeruginosa, and Y. enterocolitica) of the nine bacterial species (Table 1). On the other hand, six of the nine bacterial species were susceptible to the A. rigidula extracts (Table 1). Among the six bacterial species, four are gram-negative (P. alcalifaciens, P. aeruginosa, Y. enterocolitica and E. coli) and two are gram-positive (S. aureus and E. faecalis). The mZOIs ranged from 8.70 to 17.56 mm for acetone extract, 7.83–14.43 mm for methanol extract, and 6.00–12.33 mm for acetic acid extract (Table 1). Gram-positive bacteria generally lack the outer membrane, exposing them to antibiotics or in this case, secondary metabolites. Without the outer membrane, secondary metabolites are able to penetrate the peptidoglycan envelope and reach the cell membrane of gram-positive bacteria easier [30]. Conversely, gram-negative bacterial cell wall outer membrane (a lipopolysaccharide) is thought to act as a barrier to many substances including antibiotics [27]. This was also observed in our study, more gram-negative bacteria conferred resistance to the extracts.
An analysis of variance (table not shown) was done to test any significant difference between plant species. For all six bacteria, A. rigidula extracts showed significant differences in mZOI when compared to A. berlandieri extracts (Fig. 1). Of the six susceptible bacteria, P. alcalifaciens (p < 0.001), E. faecalis (p < 0.01), S. aureus (p < 0.001), and Y. enterocolitica (p < 0.001) were significantly inhibited by A. rigidula when compared to A. berlandieri.

Comparison of the antibacterial activities of A. berlandieri and A. rigidula. Non-overlapping CIs for side-by-side bars indicate significant difference in mean ZOI
Studies suggest that Acacia species possess secondary metabolites with antibacterial properties that can be used to inhibit the growth of bacteria [25, 26]. For example, in A. arabica bark extracts, phytochemical screening revealed the presence of terpenoids, phenols, tannins, flavonoids, and cardiac glycosides in both the acetone and the methanol plant extracts [28]. Thus, the antibacterial activities in our study could also be attributed to secondary metabolites. While, the significant difference in antibacterial activities between A. rigidula and A. berlandieri could be attributed to (1) a difference in the specific secondary metabolites present in the respective extracts and/or (2) the concentration of extracted secondary metabolites in A. rigidula is higher than in A. berlandieri, thus contributing to the higher antibacterial activities observed in A. rigidula. Results of phytochemical analysis should reveal any difference in the secondary metabolites (e.g., flavonoids, tannins, saponins of the two extracts) between the two species.
Of the three solvents used for extraction, acetone extracts showed a higher mZOI than those of methanol and acetic acid extracts. However, there was no significant difference among these three solvents. As previously mentioned, P. alcalifaciens (p < 0.001), E. faecalis (p < 0.01), S. aureus (p < 0.001), and Y. enterocolitica (p < 0.001) were significantly inhibited by A. rigidula when compared to A. berlandieri. On the other hand, S. marcescens, E. aerogenes, and K. pneumonia, were not inhibited by any of the extracts, which is attributed to the resistance commonly seen in gram-negative bacteria. Our results were similar from two other studies of acetone and methanol extracts of two species belonging to the Fabaceae family, A. arabica and A. aulacocarpa [27, 28].
Previous study by Lawrence et al. using extracts of A. arabica, and using different solvents, also showed varying degrees of antibacterial activities against gram-positive and gram-negative organisms [26]. More specifically, E. coli, S. aureus, and P. aeruginosa were among three of the six susceptible bacteria [26]. Acetone extracts of A. arabica displayed promising results, specifically against S. aureus with a mZOI ranging between 17.67 and 22.67 mm [26]. On the other hand, A. aulacocarpa methanol extracts inhibited both gram-positive and gram-negative bacteria [25]. These included Y. enterocolitica, E. faecalis and S. aureus. These bacteria were also susceptible to the A. rigidula’s methanol extract in our study. Similar to A. rigidula and A. berlandieri extracts, A. aulacocarpa extract from a previous study was also not inhibitory to the three gram-negative bacteria, K. pneumonia, E. aerogenes and S. marcescens [25]. As previously discussed, gram-negative confers more resistance against antibiotics than gram-positive bacteria
2.2 Minimum inhibitory concentration
Results of our antibacterial assays revealed potentially effective microbial suppressors; therefore, the minimal inhibitory concentration (MIC) of the extracts against the four most susceptible bacteria were determined. Acacia extracts’ MIC values, whereby no detectable bacterial growth was observed, were determined by broth microdilutions (Table 2). MIC against P. alcalifaciens, S. aureus, Y. enterocolitica, and E. faecalis of A. rigidula and A. berlandieri ranged from 37.5 to 75 mg/mL and 37.5 to 150 mg/mL, respectively. A. rigidula displayed lower MIC than A. berlandieri which means greater effectivity against the tested bacteria. In addition, acetone extracts displayed a lower MIC when compared to using other solvents between species.
In comparison with previously reported MIC values of A. nilotica [31], the MIC values of Acacia leaf extracts in our study were higher. MIC values of A. nilotica were in the range of 1.56–3.12 mg/mL. The great variation of MICs when comparing all the three Acacia species could be attributed to the method of extraction, the bacterial strains used, and the volatile nature of the chemical constituents found within each plant [32]. By further evaluating the mode of action by which extracts inhibit the growth of selected pathogens, plants found belonging to the Acacia family can provide an alternative approach to antibiotics that are no longer effective.
2.3 Qualitative phytochemical analysis
Using standard procedures for phytochemical analysis, all extracts were analyzed for the presence of secondary metabolites. Of the six extracts, most contained flavonoids, saponins, phenols, tannins and terpenoids (Table 3). These secondary metabolites may be responsible for the antimicrobial activity through different mechanisms [33,34,35,36]. All extracts of A. rigidula and A. berlandieri were negative for alkaloids. Both A. rigidula and A. berlandieri showed presence of the same set of secondary metabolites tested in our study (Table 3). However, it was observed that A. rigidula extracts when compared to A. berlandieri extracts have potentially higher concentration of diterpenes, and tannins as shown by more intense color formations. Diterpenes and tannins could potentially account for the significantly higher antibacterial activity observed in A. rigidula extracts. The quantitative determination of diterpenes and tannins, and the other secondary metabolites should be done in order to address if there is a significant difference in the concentration of secondary metabolites between A. rigidula and A. berlandieri.
Further investigation of tannins e.g., the quantification and comparison of total tannins between A. rigidula’s and A. berlandieri’s extracts and the isolation of tannins is of interest. Tannin compounds have been noted to possess antibacterial, antiviral, and inhibitory effects on enzymes [37]. Studies have suggested that tannins possess antibacterial capabilities, more specifically and importantly on S. aureus. When exposed to several tannins, S. aureus is unable to coagulate plasma [38]. Results of our qualitative phytochemical analysis revealed that A. rigidula extracts exhibited potentially higher amounts of tannins in comparison to A. berlandieri. A. berlandieri provided little to no evidence when testing for tannins in methanol and acetic acid extract. This can attribute to the inhibitory effects of all three extracts from A. rigidula, all of which showed anti-S. aureus activity.
2.4 Chemical analysis
2.4.1 Nuclear magnetic resonance
Nuclear Magnetic Resonance (NMR) is a sensitive and powerful method of detecting presence of different protons depending on their chemical environment. NMR can be used to monitor reaction progress and to detect the presence of different compounds. A combination of non-destructive spectroscopic methods such as IR and NMR to detect the small amounts of relatively large molecular weight natural products are commonly preferred [39].
Due to the high polarity of the extracts, deuterated water (D2O) was used as NMR solvent. The residual peak for the D2O solvent showed up at 4.8 ppm. The extracts were found to be insoluble in common NMR solvents such as chloroform-d (CDCl3) and benzene-d6 (C6D6). Since phytochemical analysis yielded clues for the structure of the compounds; the chemical shifts (δ, in ppm) observed were used in order to access different types of protons that are indicative of the presence of the class of metabolites present in the extract. Representative 1H NMR spectra of extraction solvents of A. rigidula are reported in Fig. 2. The 1H NMR spectra for the acetone and methanol extracts display peaks around δ 3.3–4.0 ppm due to the presence of –OH group and methoxy group (indicative of the presence of saponins). These peaks conform with the 1H NMR spectral data of isolated saponin reported by Sharma and Paliwal [25]. The broad peaks associated with OH groups seen in the 1H NMR of the acetone and methanol extract may be due to the exchangeable nature of such protons. Because a protic deuterated solvent was used (D2O), the OH protons exchanged with the deuterium and the peaks would appear too broad or disappear entirely, as in the case for the acetic acid extract where broad peaks did not show up in the 1H NMR spectrum [40]. The peak at 5.3 ppm, assignable to C=C protons, also appeared for both acetone and methanol extracts, conforming with the literature [25]. Other than saponins, C=C is part of the chemical structure of natural products that showed positive qualitative phytochemical tests such as linoleic acid and/or terpenes.

1H NMR spectra of Acacia rigidula’s acetone, methanol, and acetic acid extracts in deuterated water (D2O)f
Signals between the regions of 6.5–7.5 ppm are associated with the aromatic protons and are present in both the acetone and methanol extracts. Furthermore, the results of our experiment are comparable to those of Buchanan et al. [41] where the presence of aromatic rings in A. confusa were detected using 1H NMR spectroscopy. The isolation and purification of the extracts is highly recommended in order to confirm the results of our experiment.
The results from our study identified potential chemical structures of secondary metabolites that were also qualitatively determined in the phytochemical analysis. The next two analytical techniques further support the presence of the secondary metabolites reported in all three extracts, as qualitatively determined by our phytochemical analysis.
2.4.2 Ultraviolet–visible spectroscopy
Ultraviolet–Visible (UV–Vis) spectroscopy allows for the determination of presence of conjugated organic compounds in unknown extracts. The qualitative UV–Vis spectroscopy profile of all three extracts of A. rigidula was selected at a wavelength between 200 to 450 nm. The UV–Vis profile of the acetone extracts showed peaks at 215, 235, 260, 270, 290, 310, and 325 nm with the absorption 4.523, 10.000, 10.000, 10.000, 10.000, 3.676, and 3.232, respectively (Fig. 3). The absorbance values of each peak were greater than 1.000 and this can be attributed to how concentrated the extracts were. As a result, in the UV–Vis spectra there is an appearance of one or more peaks in the region from 200 to 400 nm with high absorbance values, which is indicative of the presence of conjugation, unsaturated groups, and/or heteroatom such as oxygen [42]. The results of this profile provide characteristics for phenols and its derivatives such as flavonoids and saponins [43].

Ultraviolet–visible spectra of Acacia rigidula’s acetone, methanol, and acetic acid extracts
The spectrum for A. rigidula acetone extract shows multiple peaks within these exact parameters. More so, flavonoids typically consist of two absorption maxima in the ranges 240–285 nm (band I) and 300–350 nm (band II) for the aromatic rings present in flavonoids [43]. UV–Vis analysis reported on two species belonging to the Fabaceae family, Acacia raddiana and Meizotropis pellita, support the findings from this study [43, 44]. Results on these two species presented similar chemical characteristics that relate to those found in flavonoid derivatives [43, 44]. UV–Vis spectra of M. pellita displayed peaks at 235 and 270 nm, while the spectra for A. raddiana showed a peak at 272 nm, both of which are associated with the bands that correspond with flavonoids and are shown in the results of this study. The findings from UV–Vis analysis were further supplemented with the analytical technique known as infrared (IR) spectroscopy.
2.4.3 Infrared spectroscopy
Infrared spectroscopy was performed to determine the functional groups present in all three extracts of leaves of A. rigidula. The results obtained in the infrared region enable the identification of the chemical constituents and insights of the functional groups in the structures of compounds present. A stack of IR spectra is illustrated in Fig. 4. The acetone extract displayed an absorption at 3263 cm−1 that is due to the stretching of hydroxyl groups (O–H) present in the extract. The strong peak at 2927 cm−1 is common for hydrocarbons or organic compounds and due to C-H stretch. The peaks obtained at 1702 cm−1 indicates the presence of C=O stretching (carboxylic acids for linoleic acid and saponins, ketones for flavonoids, and ester for tannins), while the peak at 1031 cm−1 is associated with C-O of esters (e.g., tannins) and/or ether (flavonoids/flavone/isoflavone/flavonol). The peaks at 1606 cm−1 and 1442 cm−1 is due to C=C stretching associated with the unsaturated alkene and aromatic skeletal mode of the extracts, respectively .

Infrared spectra of Acacia rigidula extracts: acetone, methanol, and acetic acid. Peak values are given for distinct functional groups absorbed at their respective value
The results from other studies of M. pellita and A. raddiana both confirmed the presence of alcohols, carboxylic acids, and aromatic compound through the use of infrared spectroscopy to elucidate structural compounds in their extracts [43, 44]. IR spectra of both A. raddiana and M. pellita displayed stretches produced by hydroxyl groups between 3269.21 cm−1 and 2249.81 cm−1, respectively. Methylene stretches in both species were seen between the wavenumbers of 2927–2031 cm−1, and C = O stretches between 1649.80–1691.20 cm−1.
2.5 Antioxidant assay
Oxidative damage by free radicals plays a critical role in the pathogenesis of human diseases, including cancer, emphysema, and neurodegenerative diseases such as Alzheimer's disease [45]. Therefore, the identification of exogenous antioxidants to overcome the toxicity generated by free radicals is of high interest. In this experiment, the antioxidant activity in A. rigidula and A. berlandieri extracts were determined. The ferric thiocyanate method was used to assess the ability of all extracts to prevent oxidation. The antioxidant activity of all three extracts were determined using ferric thiocyanate by measuring the percent inhibition of linoleic acid oxidation (Fig. 5). Linoleic acid is a polyunsaturated fatty acid and under peroxidation, super radicals are formed that oxidizes Fe2+ to Fe 3+ which forms a complex with SCN− and its concentration can be determined by measuring its absorbance at 500 nm [46].

Antioxidant properties of A. berlandieri and A. rigidula extracts (5 mg/25 mL) determined by ferric thiocyanate method. Positive controls (BHT and ascorbic acid) and negative control (linoleic acid) were used to compare results from each extract. Duncan’s Multiple Range Test indicated a significant difference as shown by the non-overlapping confidence intervals. Mean values are from three replicates run in quadruplets
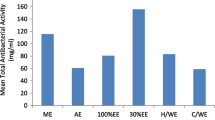
Our preliminary results from the antioxidant assays showed that the acetic acid extracts from neither A. rigidula nor A. berlandieri did not show promising antioxidant activity. Thus, only the comparison among A. rigidula’s and A. berlandieri’s acetone and methanol extracts were done. A. rigidula’s acetone extract exhibited the significantly (p < 0.001) highest inhibition of peroxidation, 42%. While a 25%, 19%, and 13% inhibition were observe for A. rigidula’s methanol extract, A. berlandieri ‘s acetone and A. berlandieri ‘s methanol extracts, respectively. All extracts were compared to the positive controls, BHT and ascorbic acid, where both showed inhibition of peroxidation of 70% and 69%, respectively. All extracts exhibited a significant difference (p < 0.001) of percent inhibition of oxidation compared to one another and the negative control, linoleic acid. A study performed by Sultana et al. [29] found a higher antioxidant activity in A. nilotica’s acetone and methanol extracts which reported inhibition of peroxidation of 75–86%. However, they used bark extracts, and our study used leaf extracts.
The observed antioxidant activity in our Acacia extracts could be attributed to secondary metabolites namely flavonoids, terpenes, saponins, and tannins that were determined to be present in these extracts. We can only assume that A. rigidula acetone extracts significant antioxidant activity could be attributed to terpenes and tannins that were qualitatively higher in A. rigidula compared to A. berlandieri. Terpenes, tannins, flavonoids, and saponins are secondary metabolites known to contribute to antioxidant activities. These secondary metabolites were also present in the Acacia extracts as shown by our phytochemical analysis, and were suggested to be the major secondary metabolites from the results of chemical analysis. Thus, the quantitative determination of terpenes and tannins, as well as flavonoids and saponins will potentially explain the higher antioxidant activity observed in A. rigidula. Likewise, a nuanced correlation analysis based on the total quantity of each of those secondary metabolites and results of antioxidant assays may also be able to tease out and explain the difference in antioxidant activities between the species and among extracts.
3 Experimental section
3.1 Chemicals
Acetone, methanol, acetic acid, dimethyl sulfoxide (DMSO), and ethanol were purchased from Fischer Scientific (Fair Lawn, New Jersey). Linoleic acid, butylated hydroxytoluene (BHT), ascorbic acid, hydrochloric acid, and ammonium thiocyanate were acquired from Sigma-Aldrich (St. Louis, Missouri). Ferrous chloride was purchased from Ricca Chemical Company (Arlington, Texas). Sodium phosphate monobasic dihydrate and sodium phosphate dibasic, 7-hydrate crystal were purchased from Avantor Performance Materials, Inc. (Center Valley, Pennsylvania).
3.2 Plant collection
Fresh leaves (the specimens) of A. berlandieri and A. rigidula were collected at Texas A&M International University (TAMIU) campus with respective coordinates (27°34′27′’N, 99°25′56′'W and 27°34′26′’N, 99°25′55′’W) in September to November, 2019. The specimens’ identities were confirmed by TAMIU Department of Biology’s ecologist, Dr. Neal McReynolds. Voucher specimens of A. berlandieri (accession number WSCO 004,009) and A. rigidula (accession number WSCO 004,010) were deposited at Mary Carver Hall Herbarium at Weber State University (WSCO). The specimens' identities were authenticated by WSU botanist and herbarium curator, Dr. James Cohen. For this study, two trees (one from each species) were sampled whereby each tree was sampled for its leaves on three distinct frontages (i.e., northern, western, and eastern frontage of the tree). For every sample, 30 g of plant leaves were collected and stored in labeled bags. In total, six samples were collected for sequential extraction. After plant samples were collected, they were stored at −20 °C, freeze dried and grounded into a powder consistency using a coffee blender.
3.3 Crude extraction
Each powdered plant sample of A. berlandieri and A. rigidula was extracted separately with different organic solvents based on the increasing polarity order of acetone (30%) < methanol (20%) < acetic acid (7%); and a 1:10 mass: volume ratio. One of the secondary metabolites likely present in our Acacia leaf extracts are phenolic compounds, and the previously used solvents for their extractions are methanol, acetone, ethanol and a form of acid [47, 48]. Thus, those three solvents were used in our extraction procedure. In addition, solvent polarity plays a key role in increasing phenolic solubility. The combination of water and acetic acid has been shown to extract bioactive compounds with antioxidant capacity, specifically flavonoids and alkaloids [49, 50].
Each sample was homogenized separately in each solvent with shaking (VWR S-500 Orbital Shaker) for 48 h at 350 rpm. Following the 48 h shaking, the homogenate was centrifuged at 3000 rpm for 20 min (Beckman GS-15 Series Centrifuge) and the supernatant was collected and evaporated for the respective solvent using a Hei-VAP Precision rotary evaporator and finally lyophilized (Labconco 2.5 L) [31]. The product after lyophilization was designated as the extract. The extract was stored in a −20 °C freezer for future use. After centrifugation, the remaining pellet was added to the next solvent for extraction. The extracts were then tested for their antimicrobial activity.
3.4 Microbial cell culturing
The microorganisms tested for antibacterial activity were purchased from Presque Isle Cultures namely: Yersinia enterocolitica (Cat# + 330), Escherichia coli (Cat#336), Staphylococcus aureus (Cat# + 4651), Providencia alcalifaciens (Cat#368), Enterobacter aerogenes (Cat#341), Serratia marcescens (Cat#3612), Klebsiella pneumonaie (Cat# + 344), Pseudomonas aeruginosa (Cat# + 99), and Enterococcus faecalis (Cat# + 522A). Following the instructions of the manufacturer, Mueller Hinton (MH) agar (HiMedia No. 2) was prepared and sterilized by autoclaving. Streak plating was performed to prepare master plates and a single colony of each microorganism was used to prepare overnight cultures for the disc diffusion assay.
3.5 Bacterial inoculum preparation
Miller’s LB Broth (Amresco) was prepared following the instruction of the manufacturer and 2 mL were added to each test tube that were sterilized by autoclaving. A single colony of each bacterium, grown in MH culture plates, was transferred into each LB broth tube. Cultures were incubated in a water bath shaker at 37 °C for 16–18 h. Following incubation, the turbidity of each LB Broth was diluted with sterile water to an absorbance level of 0.132 ± 0.005 at 625 nm. This level is optically comparable to the 0.5 McFarland standards. A spectrophotometer (Bausch and Lomb, Model Spectronic 20) was used to adjust the absorbance of the suspension. This yielded a bacterial suspension of approximately 0.5–1.0 × 108 CFU/mL [51].
3.6 Testing for antibacterial activity
3.6.1 Disc diffusion assay
Antibacterial activity of leaf extracts was tested by using the disc diffusion method. Using Mueller Hinton agar, the presence of a zone of inhibition (ZOI; mm) indicated antibacterial activity by the plant extracts. From diluted bacteria in the LB broth tubes, 100 uL were inoculated to Mueller Hinton agar plates. Bacteria were spread onto the plate using a sterilized L-shaped rod, while being rotated 15 times clockwise on a platform, to ensure equal distribution of the inoculum. Sterile Whatman 6 mm antibiotic assay discs were impregnated with 20 μL of the lyophilized plant extracts dissolved in dimethyl sulfoxide (DMSO) at a concentration of 0.5 mg/μL. DMSO is the solvent used to prepare the extract solutions. DMSO also served as the negative control, as it has no ability to inhibit bacterial growth.
The following antibiotic discs were used as the antibiotic positive controls for the following bacteria: (1) 10 μg of vancomycin (Carolina Biological) was used against P. aeruginosa, K. pneumonaie, S. aureus, and E. faecalis; while (2) 10 μg of ciprofloxacin (Carolina Biological) was used against E. coli, Y. enterocolitica, E. aerogenes, P. alcalifaciens and S. marcescens. After completing the disc diffusion assay, plates were incubated at 37 °C for 18–20 h. After incubation, presence of a ZOI and its diameter (largest and smallest) were measured in millimeters using a Vernier caliper. The test was replicated three times and each replicate with three trials for the determination of antibacterial activity.
3.6.2 Statistical analysis
For each of the nine bacteria, an analysis of variance associated with a 2 × 3 factorial experiment was performed. In this experiment, species (factor 1) had 2 levels, namely: A. berlandieri and A. rigidula; and solvents (factor 2) had 3 levels namely: acetone, methanol and acetic acid. To compare the 2 × 3 treatment combinations to both positive ( +) and negative (−) controls, a mean comparison was performed among the 2 × 3 = 6 treatment combinations and the two controls. The plant species x solvent extracts were randomly allocated within each of the three frontages. Each treatment was given a number and using a random number generator, this designated the placement of each extract. In all analyses, a type-I error rate of α = 0.01 (or 1%) was used.
3.7 Minimum inhibitory concentration
Minimum inhibitory concentrations (MIC) values of Acacia extracts were determined by broth micro-dilution method [31]. Glycerol stocks of S. aureus, E. faecalis, Y. enterocolitica and P. alcalifaciens were grown in sterile Mueller Hinton Broth (MHB) overnight and diluted to a McFarland Standard of 0.5 at OD625. Antibiotic serial dilutions (0–0.512 gµ/mL) were prepared in MHB from the antibiotic stock solution and pipetted into columns 1–10 in a 96-well plate. A two-fold serial dilution of acetone and methanol extracts for both species (0.29 mg/mL-150 mg/mL) were prepared in the 96-well plate from a stock of 300 mg extract per mL of DMSO. Seventy-five µL of the antibiotic and extracts were dispensed into all wells of a 96-well plate. Seventy-five µL of the diluted bacteria was added into wells of columns 1 to 11. Column 11 was utilized as a growth control while column 12 was used as a broth sterility control. Plates were incubated at 37 °C for 18–20 h. The lowest concentration that had no visible growth after 18–20 h of incubation was considered the MIC. MIC assays were conducted with 3 replications (blocks) Each replication/block experiment was conducted in triplicates.
3.8 Qualitative phytochemical analysis
Each extract was subjected to a standard protocol for phytochemical screening to identify the presence of alkaloids, phenols, tannins, diterpenes, triterpenes, saponins, and flavonoids by standard protocols [52,53,54]. The results were graded on a scale that ranges from the absence (symbolized by a “–– “) of the respective metabolite, to the presence + (which defines minimal or slightly present), + + (intermediate or fairly present) and + + + (definite or clearly present).
3.9 Chemical analysis
Reagents and deuterated NMR solvents were obtained from commercial sources and used as received. 1H NMR spectra were recorded on a Bruker 300 MHz AVANCE I spectrometer with a quattro nucleus probe (QNP). Proton chemical shifts were given in δ values (ppm) using D2O as solvent at room temperature. UV–Vis single beam spectrophotometer Agilent Cary 60 UV–Vis Spectrophotometer was used for the absorbance measurements in the range 200–800 nm using water as solvent at room temperature. All measurements were repeated at least thrice to ensure reproducibility. Solid state IR spectra were recorded on a Varian 640-IR with single bounce ZnSe ATR cell spectrometer and υmax are partially reported (cm–1). The solid samples were pressed against the diamond crystal using the attached pressure clamp.
3.10 Testing for antioxidant activity.
3.9.1 Ferric thiocyanate method
The antioxidant activity of leaf extracts was also determined in terms of measurement of percent inhibition of peroxidation in linoleic acid system following a reported method by Sultana et al. [29]. Extracts (5 mg) of each treatment were added to a solution mixture of linoleic acid (0.13 mL), 99.8% ethanol (10 mL) and 10 mL of 0.2 M sodium phosphate buffer (pH 7). Total mixture was diluted to 25 mL with distilled water. The solution was incubated at 40 °C for 15 days and the degree of oxidation was measured following ferric thiocyanate method with 10 mL of ethanol (75%), 0.2 mL of an aqueous solution of ammonium thiocyanate (30%), 0.2 mL of sample solution and 0.2 mL of ferrous chloride (FeCl2) solution (20 mM in 3.5% HCl) being added sequentially. After three minutes of stirring, the absorption values of mixtures measured at 500 nm were taken as peroxide contents. BHT and ascorbic acid were used as positive controls while a mixture without plant samples was used as the negative control. Percent inhibition of linoleic acid peroxidation, 100 – [(Abs. increase of sample at 360 h/Abs. increase of control at 360 h) × 100], was calculated to express antioxidant activity.
3.9.2 Statistical analysis
Statistical comparison was performed using a one-way analysis of variance followed by Duncan’s multiple range test (DMRT) for mean comparison. The three crude extracts of A. rigidula, acetone, methanol, and acetic acid, were used to compare the antioxidant activity against two positive controls, BHT and ascorbic acid, and a negative control, linoleic acid. Each analysis was used to determine the significant differences among means from three replicates run in quadruplets at p < 0.05.
4 Conclusion
This study has determined presence of antibacterial and antioxidant activities in A. berlandieri and A. rigidula leaf extracts. When comparing both species, extracts of A. rigidula have significantly (p < 0.001) higher antibacterial and antioxidant activities than extracts of A. berlandieri. NMR, UV–Vis, and IR spectroscopy confirmed the presence of the secondary metabolites that showed positive results in the qualitative phytochemical analysis. The secondary metabolites potentially contributing to the observed biological activities in the Acacia leaves understudy are terpenes, tannins, flavonoids and saponins. The results of this study provide a basis for further investigations of the A. rigidula leaf extracts. A. rigidula leaf extracts have potential to serve as a source of novel antimicrobial and antioxidant agents.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- 1H NMR:
-
Proton nuclear magnetic resonance
- BHT:
-
Butylated hydroxytoluene
- C6D6 :
-
Deuterated benzene
- CDCl3 :
-
Deuterated chloroform
- CI:
-
Confidence intervals
- D2O:
-
Deuterated water
- DMRT:
-
Duncan’s multiple range test
- DMSO:
-
Dimethyl sulfoxide
- Fe2 + :
-
Ferrous ion
- Fe3 + :
-
Ferric ion
- FeCl2 :
-
Ferrous chloride
- IR:
-
Infrared
- MeOH:
-
Methanol
- MIC:
-
Minimum inhibitory concentration
- mZOI:
-
Mean zone of inhibition
- NMR:
-
Nuclear magnetic resonance
- PM:
-
Primary metabolites
- Ppm:
-
Parts per million
- SCN− :
-
Thiocyanate
- SM:
-
Secondary metabolites
- UV–Vis:
-
Ultraviolet–visible
References
Gorlenko CL, Kiselev HY, Budanova EV, Zamyatnin AA, Ikryannikova LN (2020) Plant secondary metabolites in the battle of drugs and drug-resistant bacteria: new heroes or worse clones of antibiotics? Antibiotics. https://doi.org/10.3390/antibiotics9040170
Larayetan R, Ololade ZS, Ogunmola OO, Ladokun A (2019) Phytochemical constituents, antioxidant, cytotoxicity, antimicrobial, antitrypanosomal, and antimalarial potentials of the crude extracts of Callistemon citrinus. Evid-Based Complement Altern Med 2019:e5410923. https://doi.org/10.1155/2019/5410923
González-Burgos E, Carretero ME, Gómez-Serranillos MP (2011) Sideritis spp: Uses, chemical composition and pharmacological activities—a review. J Ethnopharmacol 135(2):209–225
Dias DA, Urban S, Roessner U (2012) A historical overview of natural products in drug discovery. Metabolites 2:303
Ben Amor IL, Neffati A, Ben Sgaier M, Bhouri W, Boubaker J, Skandrani I, Bouhlel I, Kilani S, Ben Ammar R, Chraief I, Hammami M, Ghoul M, Chekir-Ghedira L, Ghedira K (2008) Antimicrobial activity of essential oils isolated from phlomis crinite cav ssp mauritanica munby. JAOCS J Am Oil Chem Soc 85(9):845–849
Dzoyem JP, McGaw LJ, Eloff JN (2008) In vitro antibacterial, antioxidant and cytotoxic activity of acetone leaf extracts of nine under-investigated Fabaceae tree species leads to potentially useful extracts in animal health and productivity. BMC Complement Altern Med 14(1):1–7
Alves RRN, Rosa IML (2007) Biodiversity, traditional medicine and public health: where do they meet? J Ethnobiol Ethnomed 3:1
Hossain MK, Hassan MM, Parvin MN, Hasan MM, Islam MS, Haque MA (2012) Antimicrobial, cytotoxic and thrombolytic activity of Cassia senna leaves (family: fabaceae). J Appl Pharm Sci 2:186
Asghar R, Ahmad M, Zafar M, Akram A, Mahmood J, HassanPakistan M (2003) Antibacterial efficacy of Acacia modesta wall miswak against dental pathogen. J Biol Sci 6(24):2024–2025
Mazid M, Khan TA, Mohammad F (2011) Role of secondary metabolites in defense mechanisms of plants. Biol Med 3:232
Seema DN (2013) J Pharmacogn preliminary phytochemical analysis and in vitro investigation of antibacterial activity of Acacia nilotica against clinical isolates. Phytochem 1:23
Madjid AMO, Sanni A, Lagnika L (2020) Chemical diversity and pharmacological properties of genus Acacia. Asian J Appl Sci 13:40
Farzana MUZN, Sultana A (2014) A review of ethnomedicine, phytochemical and pharmacological activities of Acacia nilotica (Linn) Willd. J Pharmacogn Phytochem 3(1):84–90
Ali A (2012) Acacia nilotica: a plant of multipurpose medicinal uses. J Med Plants Res 6:1492
Mattana CM, Cangiano MA, Alcaráz LE, Sosa A, Escobar F, Sabini C, Sabini L, Laciar AL (2014) Evaluation of cytotoxicity and genotoxicity of acacia aroma leaf extracts. Sci World J 2014:e380850. https://doi.org/10.1155/2014/380850
Afsar T, Razak S, Shabbir M, Khan MR (2018) Antioxidant activity of polyphenolic compounds isolated from ethyl-acetate fraction of Acacia hydaspica R parker. Chem Cent J 12:1
Sadiq MB, Tharaphan P, Chotivanich K, Tarning J, Anal AK (2017) In vitro antioxidant and antimalarial activities of leaves, pods and bark extracts of acacia nilotica (L) del. BMC Complement Altern Med 17(1):1–8
Uzunuigbe EO, Osunsanmi FO, Masamba P, Mosa RA, Opoku AR, Kappo AP (2019) Phytochemical constituents and antioxidant activities of crude extracts from Acacia Senegal leaf extracts. Pharmacogn J 11:1409
Borges A, José H, Homem V, Simões M (2020) Comparison of techniques and solvents on the antimicrobial and antioxidant potential of extracts from Acacia dealbata and Olea europaea. Antibiotics. https://doi.org/10.3390/antibiotics9020048
Lin HY, Chang TC, Chang ST, Tradit J (2018) A review of antioxidant and pharmacological properties of phenolic compounds in Acacia confuse. Complement Med 8:443
Pawar RS, Grundel E, Fardin-Kia AR, Rader JI (2014) Determination of selected biogenic amines in Acacia rigidula plant materials and dietary supplements using LC-MS/MS methods. J Pharm Biomed Anal 88:457
Alanís-Garza BA, Arroyo JL, González GG, González EG, De Torres NW, Aranda RS (2017) Anti-fungal and anti-mycobacterial activity of plants of Nuevo Leon Mexico. Pak J Pharm Sci 30(1):17–21
Escárcega-González CE, Garza-Cervantes JA, Vázquez-Rodríguez A, Montelongo-Peralta LZ, Treviño-González MT, Castro EDB, Saucedo-Salazar EM, Morales RMC, Regalado Soto DI, Treviño-González FM, Rosales JC, Morones-Ramírez JR (2018) In vivo antimicrobial activity of silver nanoparticles produced via a green chemistry synthesis using Acacia rigidula as a reducing and capping agent. Int J Nanomedicine 13:2349
Seigler DS (2003) Phytochemistry of Acacia - Sensu lato. Biochem Syst Ecol 31:845
Mattana CM, Satorres SE, Sosa A, Fusco M, Alcaráz LE (2010) Brazilian Antibacterial activity of extracts of Acacia aroma against methicillin-resistant and methicillin-sensitive Staphylococcus. J Microbiol 41:581
Jain S, Sharma P, Jhade D, Sharma NK, Paliwal P, Ahirwar D (2011) Pharmacognostic screening and phytochemical evaluation of Acacia leucophloea root. Int J Green Pharm 5:155
Cock I (2012) Antimicrobial Activity of Acacia aulacocarpa and Acacia complanta methanolic extracts. Pharmacogn Commun 2:66
Lawrence R, Jeyakumar E, Gupta A (2015) Antibacterial activity of Acacia arabica (bark) extract against selected multi drug resistant pathogenic bacteria. Int J Curr Microbiol App Sci 1:213
Wink M (2015) Modes of action of herbal medicines and plant secondary metabolites. Medicines 2:251
Al-Ani I, Zimmermann S, Reichling J, Wink M (2015) Pharmacological synergism of bee venom and melittin with antibiotics and plant secondary metabolites against multi-drug resistant microbial pathogens. Phytomedicine 22:245
Sadiq MB, Tarning J, Cho TZA, Anal AK (2017) Antibacterial activities and possible modes of action of Acacia nilotica (L) del against multidrug-resistant escherichia coli and salmonella. Molecules 22(1):47
Mostafa AA, Al-Askar AA, Almaary KS, Dawoud TM, Sholkamy EN, Bakri MM (2018) Antimicrobial activity of some plant extracts against bacterial strains causing food poisoning diseases. Saudi J Biol Sci 25:361
Mujeeb F, Bajpai P, Pathak N (2014) Phytochemical evaluation, antimicrobial activity, and determination of bioactive components from leaves of Aegle marmelos. Biomed Res Int 2014:1–11. https://doi.org/10.1155/2014/497606
Mrudulakumari Vasudevan U, Lee EY (2020) Flavonoids, terpenoids, and polyketide antibiotics: Role of glycosylation and biocatalytic tactics in engineering glycosylation. Biotechnol Adv 41:107550
Sparg SG, Light ME, Van Staden J (2004) Biological activities and distribution of plant saponins. J Ethnopharmacol 94:219
Mandal P, Sinha Babu SP, Mandal NC (2005) Antimicrobial activity of saponins from Acacia auriculiformis. Fitoterapia 76:462
Nakashima H, Murakami T, Yamamoto N, Sakagami H, Tanuma SI, Hatano T, Yoshida T, Okuda T (1992) Inhibition of human immunodeficiency viral replication by tannins and related compounds. Antiviral Res 18:91
Akiyama H, Fujii K, Yamasaki O, Oono T, Iwatsuki K (2001) Antibacterial action of several tannins against Staphylococcus aureus. J Antimicrob Chemother 48:487
Sharma V, Paliwal R (2013) Isolation and characterization of saponins from Moringa oleifera (moringaeceae) pods. Int J Pharm Pharm Sci 5:179
Brown GD, Bauer J, Osborn HMI, Kuemmerle R (2018) A solution NMR approach to determine the chemical structures of carbohydrates using the hydroxyl groups as starting points. ACS Omega 3:17957
Buchanan M, Carroll A, Pass D, Quinn R (2007) Dynamics and conformations of PEO chains chemically bonded on silica: comparison between 1H and 2H NMR. Magn Reson Chem 45:488
Jain PK, Soni A, Jain P, Bhawsar J (2016) Phytochemical analysis of Mentha spicata plant extract using UV-VIS, FTIR and GC/MS technique. J Chem Pharm Res 8:1
Elgubbi H, Altajtal A, Elbath A (2016) Phytochemical screening study and anti-candida evaluation of Acacia raddiana leaves. EC Nutrition. https://www.researchgate.net/publication/336685391_Phytochemical_Screening_Study_and_Anti-Candida_Evaluation_of_Acacia_raddiana_Leaves
Rani N, Sharma S, Sharma M (2016) Phytochemical analysis of Meizotropis pellita by FTIR and UV- VIS spectrophotometer. Indian J Sci Technol 9:1
Subhaswaraj P, Sowmya M, Jobina R, Sudharshan SJ, Dyavaiah M, Siddhardha B (2017) Determination of antioxidant potential of Acacia nilotica leaf extract in oxidative stress response system of Saccharomyces cerevisiae. J Sci Food Agric 97:5247
Sultana B, Anwar F, Przybylski R (2007) Antioxidant activity of phenolic components present in barks of Azadirachta indica, Terminalia arjuna, Acacia nilotica, and Eugenia jambolana Lam trees. Food Chem 104:1106
Alothman M, Bhat R, Karim AA (2009) Antioxidant capacity and phenolic content of selected tropical fruits from Malaysia, extracted with different solvents. Food Chem 115:785
Lafka TI, Sinanoglou V, Lazos ES (2007) On the extraction and antioxidant activity of phenolic compounds from winery wastes. Food Chem 104:1206
Złotek U, Mikulska S, Nagajek M, Świeca M (2016) The effect of different solvents and number of extraction steps on the polyphenol content and antioxidant capacity of basil leaves (ocimum basilicum L.) extracts. Saudi J Biol Sci 23:628
Shoaib M, Shah SWA, Ali N, Shah I, Ullah S, Ghias M, Tahir MN, Gul F, Akhtar S, Ullah A, Akbar W, Ullah A (2016) Scientific investigation of crude alkaloids from medicinal plants for the management of pain BMC Complement. Altern Med 16:1
Ynalvez R, Compean K, Addo-Mensah A (2018) Qualitative determination of the secondary metabolites and evaluation of the antimicrobial activity of leaf extracts from different plant families (Boraginaceae, Fabaceae, Lamiaceae and Lauraceae) against microorganisms of clinical importance. J Pharm Res Int 23:1
Lu Y, Knoo T-J, Wiart C (2014) Phytochemical analysis and antioxidant activity determination on crude extracts of Melodinus eugeniifolus barks and leaves from Malaysia. Pharmacol Pharm 05:773
Panti AB, Aguda RM, Razal RA, Belina-Aldemita MD, Tongco JVV (2014) Proximate analysis, phytochemical screening and total phenolic and flavonoid content of the ethanolic extract of molave Vitex parviflora Juss leaves. J Chem Pharm Res 6:1538
Usman A, Abdulrahman FI, Usman A (2009) Qualitative phytochemical screening and in vitro antimicrobial effects of methanol stem bark extract of Ficus thonningii (Moraceae). African J Tradit Complement Altern Med 6:289
Acknowledgements
National Science Foundation Major Research Instrumentation Award#0959395 to RA Ynalvez
Funding
Texas A&M International University.
Author information
Authors and Affiliations
Contributions
Peter Cavazos: formal analysis, investigation, methodology, writing. David Gonzalez: investigation, review & editing Jocelyn Lanorio: formal analysis, methodology, resources, review & editing Ruby Ynalvez: conceptualization, formal analysis, funding acquisition, investigation, methodology, project administration, writing, review & editing
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cavazos, P., Gonzalez, D., Lanorio, J. et al. Secondary metabolites, antibacterial and antioxidant properties of the leaf extracts of Acacia rigidula benth. and Acacia berlandieri benth.. SN Appl. Sci. 3, 522 (2021). https://doi.org/10.1007/s42452-021-04513-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42452-021-04513-8