
Abstract
A comparative evaluation of grain legumes is essential for the effective planning of legume-based agricultural systems in a given environment. The goal of this work was to contrast the growth, translocation of assimilates, and grain yield of spring-planted common vetch (Vicia sativa L.), red pea (Lathyrus cicera L.), lentil (Lens culinaris Medik.), chickpea (Cicer arietinum L.), and field pea (Pisum sativa L.) under rainfed Mediterranean conditions. Two cultivars of each species were cultivated on a silty clay soil in northeastern Greece for 2 years (2014 and 2015) with contrasting rainfall patterns. Chickpea and field pea exhibited better early crop growth rate than any other legume. Species differences in assimilates availability prior to grain filling affected the remobilization of assimilates to seed, which increased by 45% for every kg ha−1 rise in early dry matter accumulation. Dry matter translocation efficiency varied from 9 to 51% depending on species and year. Red pea was the best option in terms of seed yield, regardless of the seasonal rainfall. Chickpea in the drier year (2015) and field pea in the wetter year (2014) produced seed yields that were comparable to that of red pea. Lentil and common vetch were generally less productive species in terms of seed yield. Species seed yield was associated with their ability to accumulate biomass either before podding (r = 0.52, P < 0.05) or at maturity (r = 0.51, P < 0.05), but not with harvest index or translocation of dry matter. Findings provide new knowledge regarding growth attributes and reallocation of assimilate in five legume species grown simultaneously in the same environment, which has never been studied before. In addition, results highlight that selecting species with enhanced early or final biomass potential as well as adopting cultural practices that promote biomass accumulation in the growing season appear to be effective management strategies for improving seed yield of the tested grain legumes under Mediterranean conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
European cropping systems are dominated by cereals, while the cultivation of grain legumes is restricted to less than 3% of the total arable land (Eurostat, 2019). This situation has raised several concerns regarding the lack of crop diversity (Plaza-Bonilla et al., 2017) and the increased dependence on chemical fertilizers (Sainju et al., 2012), which has been linked to serious environmental risks (Bouwman et al., 2013). Seed yields of grain legumes vary across species and European sub-regions, being generally more unstable compared with cereals grain yields (Cernay et al., 2015). Aside from production uncertainty, European farmers are not encouraged to grow legumes in favorable environments and on high-quality soils; instead, they prefer to grow more profitable major crops in such situations than legumes (Cernay et al., 2015).
Legumes, when grown as part of a rotation scheme, contribute to the long-term environmental improvement because of their capacity to fix the atmospheric nitrogen, their beneficial effects on the following crop, as well as the services they provide to other agroecosystem components (Stagnari et al., 2017). Legume-wheat cropping systems, in particular, is an appealing choice for increasing soil nitrogen and organic carbon, as well as improving crop productivity, both in conventional and low-input cropping systems (Geng et al., 2023; Virk et al., 2022), regardless of whether the seeds of plants are harvested or not (Geng et al., 2023). Several factors, including growing season environmental conditions (Hungria & Vargas, 2000) and agronomic management strategies (Guinet et al., 2020), influence the extent of soil nutrient improvement achieved by legume cultivation. Furthermore, because grain legumes are used in animal feed, expanding their cultivation in Europe can reduce the dependency of livestock sector on imports of soybean (de Visser et al., 2014). Finally, grain legumes can also emerge as a group of crops that mitigate the threads of climate change, which arise by the increase of temperature and water scarcity (Dutta et al., 2022).
Seed yield of grain legumes is influenced by a wide range of physiological and biochemical functions taking place throughout the crop growth, as well as by crop management practices. In general, growth and productivity of crops depend on how effectively they convert solar energy into dry matter (DM), assuming that other resources are not limited (Sinclair & Muchow, 1999). In grain legumes, the supply of assimilate for the growing seed comes from two separate sources, i.e., the current assimilation and the remobilization of stored reserves. These sources can occur simultaneously and independently of one another, even though their significance relies on environmental factors and crop developmental stages (Turner et al., 2005). The accumulation and remobilization of DM, as well as seed yield are controlled by the association between source and sink (Zhang et al., 2020). The percentage of seed weight deriving from photosynthetic products during seed filling varies according to legume species and growing environment (Davies et al., 2000; Mohapatra et al., 2003; Turner et al., 2005). In European Mediterranean areas, the performance of several grain legumes, such as soybean and chickpea, in terms of seed yield is primarily associated with increases in plant DM following the onset of grain filling (Fotiadis et al., 2019; Koutroubas et al., 1998). Additionally, it has been observed that the supply of assimilates prior to the start of grain filling has less impact on variation in assimilate translocation in chickpea than dry matter translocation efficiency (DMTE) does (Fotiadis et al., 2017).
Proper sowing time is a crucial management decision in the cultivation of grain legumes. In terms of seed yield, autumn sowing performs better than spring one in several environments (López-Bellido et al., 2008; Neugschwandtner et al., 2013). Growing grain legumes in the winter rather than the spring has also been considered a promising option for adapting to future climate changes (Falconnier et al., 2020). However, in inland Mediterranean areas sowing of grain legumes is typically shifted from autumn to spring to avoid crop failure due to frost over the winter. Dry matter translocation to seeds is considered an adaptive response of plants to unfavorable conditions during the later stages of growth, such as terminal drought (Turner et al., 2005). Even though the reallocation of assimilates to seeds has been extensively studied in grain legumes, the majority of the studies has centered on comparisons within species. In this context, great variation in pre-podding translocation of nitrogen and assimilates has been reported in several legumes, such as chickpea (Fotiadis et al., 2017), soybean (Gaspar at al., 2017), and lentil (Kurdali et al., 1997), depending on genotype and growing conditions. Additionally, agronomic practices, like intercropping with maize, can modulate DM translocation in peanut (Gao et al., 2022). Yet, there is a lack of understanding regarding the differences in growth attributes and reallocation of assimilates between legume species grown simultaneously in the same environment. Such information is necessary for the successful design of legume-supported agricultural systems across a range of various agro-ecological zones and management types (Virk et al., 2022). The goal of the current work was to compare how different spring grain legumes responded to seasonal environmental variations with respect to growth, assimilation translocation and partitioning, and grain yield under rainfed Mediterranean conditions. This knowledge can be used to improve spring legumes’ adaptive response, implement appropriate agronomic practices in leguminous cultivation, and establish indirect selection criteria for seed yield.
Materials and Methods
Experimental Procedure
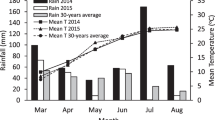
Common vetch (Vicia sativa L., cvs Pigasos and Tembi), red pea (Lathyrus cicera L., cvs Rodos and Argos), lentil (Lens culinaris Medik., cvs Thessalia and Samos), chickpea (Cicer arietinum L., cvs Andros and Kassos), and field pea (Pisum sativum L., cvs Vermio and Ithomi) were grown for two consecutive years (2014 and 2015) in field experiments at Democritus University of Thrace’s farm in Orestiada (41°33ʹN, 26°31ʹE, 33 m elevation), Greece. The region, which is in northeastern Greece, has a Mediterranean climate with hot, dry summers and cool, rainy winters. The crops were rotated each year. The soil was silty clay with the following characteristics: pH (1:2 water) 6.68, organic matter 10.1 g kg−1, electrical conductivity 0.04 S m−1, total N 1.11 g kg−1, NO3-N 9.0 mg kg−1, Olsen P 10.2 mg kg−1, and available K 495.3 g kg−1. Weather parameters (mean temperature and rainfall) during each experimental season, as well as the long-term (30 years) average are given in Table 1.
A trial with four replications was arranged in a randomized complete block design. Plots were 4 m wide (12 rows, 25 cm apart) and 3 m long. The sowing was done by hand on 17 March 2014 and 6 March 2015. Each year, 26 kg P ha−1 (in the form of superphosphate) was applied prior to sowing. Following the emergence, the density of the seedlings were reduced to 40 plants per m2 for common vetch, 60 plants per m2 for red pea, 150 plants per m2 for lentil, 50 plants per m2 for chickpea, and 100 plants per m2 for field pea. Weeds were controlled by hand weeding when necessary.
Samplings, Measurements, and Calculations
The time of the main growth stages [seedling emergence, beginning of seed filling (growth stage R5), and full maturity (growth stage R8)] were noted. At R5 and R8, plants from a row (1-m-long) were cut at the ground level in each plot and then split into distinct plant parts (i.e., leaf and stem, seeds, and pod walls). Samples of plants were weighted after being oven dried at 70 oC until constant weight. The total number of nodes (TNN) and the number of nodes under the first pod (NFP) at growth stage R5 were recorded on the main stems of 10 nearby plants randomly chosen by each plot. Crop growth rate (CGR) was determined by dividing the rise in DM across two samplings by the time (days) between these samplings. At growth stage R8, seed yield was determined by harvesting the six middle rows of each plot. The parameters related with translocation and partitioning of dry matter were calculated using the following equations (Koutroubas et al., 1998):
Statistical Analysis
Data were analyzed using the MSTAT-C (Michigan State University, East Lansing, MI) statistical software package. A year-combined ANOVA was used to analyze differences among species when averaged across all cultivars within each species. In addition, an over-years combined ANOVA was conducted separately for each species to compare cultivars within a particular species. Year, species, and cultivars as well as the interactions between them were regarded as fixed effects. Year was regarded as a variable with fixed effects, due to the considerable variation in precipitation between the research years. Means were compared with Fisher’s protected least significant difference (LSD). Simple correlation analysis was used to evaluate relationships between traits.
Results
Weather Data and Plant Development
Growing season weather varied considerably between years (Table 1). The total amount of seasonal precipitation in 2014 (483 mm) was significantly higher than that in 2015 (197 mm), indicating a noticeable wetter growing season. Total seasonal precipitation in 2014 was 114% higher compared with that in 2015 and 13% below the long-term average (226 mm). When the grain filling of legumes took place (June and July), total rainfall was 227 mm in 2014 but only 57 mm in 2015. The average temperature of each growing season (18.7 °C in 2014 and 18.9 °C in 2015) was consistent between years. However, between March and April, when plants are first beginning to grow, the mean monthly temperature in 2014 was marginally higher than in 2015. On the other hand, starting with seed growth, the average monthly temperature in 2015 was a little bit higher than in 2014.
Regarding the timing of crop emergence, there were no differences between species in either year. Crop emergence was, on average, 10 days later in 2014 than in 2015 (35 DAS vs. 25 DAS, respectively). Common vetch, red pea, and field pea all started podding earlier in 2014 (75 DAS vs. 89 DAS for lentil and chickpea). For common vetch, red pea, lentil, and field pea, the total crop duration was 120 days, while for chickpea it was 141 days. The time needed to start podding and the length of the entire crop, which averaged 74 and 116 DAS, respectively, in 2015, were the same for all species.
Crop Growth and Dry Matter Accumulation
Except for lentil, which had higher values in 2014 than in 2015, species TNN was consistent between years (Table 2). In both years, chickpea displayed the highest TNN, followed by lentil and field pea in 2014 and field pea in 2015. Species performance varied across years in terms of NFP, but the response pattern was inconsistent. In 2014, red pea had the lowest and field pea had the highest NFP. In 2015, red pea and lentil had the lowest NFP, while chickpea had the highest.
Crop growth rate up to the beginning of seed growth was significantly higher in 2014 (106 kg ha−1 day−1) than in 2015 (55 kg ha−1 day−1). Early crop growth rates varied by species, with chickpea and field pea outperforming common vetch, red pea, and lentil (Fig. 1).

Crop growth rate (CGR) up to the onset of seed growth as affected by a year of the experimentation and b legume species. Data for each year are means of five species and two cultivars for each species; data for each species are means of two cultivars and 2 years. Vertical bars represent the standard errors of the means. Bars with the same letter indicate means that are not significantly different at P < 0.05
Year, species, and their interactions had a significant impact on accumulation of DM at both samplings (Table 3). On average, the total amount of DM accumulated up to the onset of seed growth was 45% lower in 2014 compared with that in 2015. This trend was consistent across species, but the percentage of decrease between years was above average for lentil (62%) and chickpea (55%), and below average for common vetch (40%), field pea (33%), and red pea (23%). Species variation in DM accumulation at R5 was more evident in 2014, where chickpea produced the highest early DM (6770 kg ha−1). In 2015, field pea had the highest DM at R5 (3235 kg ha−1), which was comparable to chickpea (3055 kg ha−1) or red pea (2750 kg ha−1). The picture for DM accumulation at maturity was similar to that observed at R5, showing a decline in 2015 compared to 2014, which averaged 31%. In both years, chickpea had the highest DM at maturity and common vetch had the lowest. Early and final DM accumulation were significantly correlated (r = 0.77, P < 0.01). Differences in early DM accumulation between cultivars within a particular species were observed for common vetch and lentil; however, these variations did not persist across years (Table 4). In particular, there were no differences between cultivars in 2015, but DM accumulation at R5 was significantly higher in common vetch cv. Pigasos and lentil cv. Thessalia compared to cvs Tempi and Samos, respectively.
Dry Matter Translocation and Partitioning, and Seed Yield
Dry matter translocation from vegetative plant parts to seed between R5 and R8 was significantly higher in 2014 (2001 kg ha−1) than in 2015 (1025 kg ha−1) (Table 5). In 2014, lentil, chickpea, and field pea had similar DMT, which was significantly higher than that of common vetch and red pea. In 2015, there were no observable differences in DMT between species. Differences in DMT between cultivars within a particular species were observed for common vetch, red pea, and chickpea, but these differences were not always consistent across years (Table 4). In particular, DMT of red pea cv. Rodos was significantly higher than that of cv. Argos in both years. On the other hand, DMT was significantly higher in common vetch cv. Pigasos and in chickpea cv. Kassos in 2014 compared to cvs Tempi and Andros, respectively. However, this was not the case in 2015.
Dry matter translocation efficiency was significantly higher in 2014 (37%) compared with that in 2015 (20%) (Table 5). In 2014, lentil, field pea, and chickpea had similar DMTE that was significantly higher than that of common vetch and red pea. In 2015, field pea showed the highest DMTE, which was comparable to that of red pea but higher than that of common vetch, chickpea, and lentil. Dry matter translocation was positively correlated with DMTE (Fig. 2). Both DMT and DMTE were positively correlated with DM accumulation at R5 (Fig. 3).

Relationship between dry matter translocation efficiency (DMTE) and dry matter translocation (DMT) in 2014 (black symbols, solid line) and 2015 (open symbols, dashed line). Each relationship was based on means derived from five legume species (common vetch, red pea, lentil, chickpea, and field pea) and two cultivars for each species

Relationship (a) between dry matter accumulation (DMA) before the onset of grain filling (R5) and dry matter translocation (DMT), and (b) between DMA at R5 and dry matter translocation efficiency (DMTE). Each relationship was based on means derived from five legume species (common vetch, red pea, lentil, chickpea, and field pea), two cultivars for each species, and 2 years (2014, black symbols; 2015, open symbols)
Seed yield was significantly affected by year and species, as well as by the year × species interaction (Table 5). In 2014, the average seed yield (1742 kg ha−1) was much greater than it was in 2015 (1158 kg ha−1). This pattern between years was common for all species, except for lentil that had similar seed yield in both years. In 2014, lentil produced the lowest seed yield, whereas field pea produced the highest seed yield, which was comparable to that of red pea. In 2015, chickpea showed the highest seed yield that was comparable with that of field pea and red pea, but higher than that of common vetch and lentil. Seed yield showed a positive correlation with early and final DM accumulation (Fig. 4). No correlation between seed yield and DMT or DMTE was observed (r = 0.31, P > 0.05 and r = 0.29, P > 0.05, respectively).

Relationship (a) between dry matter accumulation (DMA) before the onset of grain filling (R5) and seed yield (SY), and (b) between DMA at maturity (R8) and SY. Each relationship was based on means derived from five legume species (common vetch, red pea, lentil, chickpea, and field pea), two cultivars for each species, and 2 years (2014, black symbols; 2015, open symbols)
Harvest index remained constant between years in red pea and chickpea, while it increased in common vetch, lentil, and field pea in 2015 compared to 2014 (Table 5). No species differences in HI were observed in 2014, except for chickpea, which had the lowest value. In 2015, chickpea had the highest HI, which was comparable to those of red pea and field pea. Similar HI values were observed in common vetch and lentil, which were lower than those of the other species. No significant correlation between seed yield and HI existed (r = 0.35, P > 0.05).
Discussion
Early Crop Growth
Crop agronomic performance was influenced by legume species and year of experimentation, reflecting the variation in genotypic constitution of the species as well as the seasonal variation of the weather parameters. Most legumes produced large biomass, which was particularly noticeable the year with above-average rainfall, confirming their adaptation to the area even at spring sowing. Variation between species in accumulation of DM could be attributed to differences in intercepted photosynthetically active radiation (Estrada-Campuzano et al., 2012) along with the formation of green area (Thomson & Siddique, 1997), both of which influence grain legumes adaptation in Mediterranean conditions. Regardless of growing environment, chickpea ranked first in terms of final DM production, primarily owing to its rapid crop growth rate during the early stages. Vigorous early growth benefits crop in a variety of ways, including superior crop establishment (Huang et al., 2017), enhanced water use efficiency (Siddique et al., 2001), and weed competition (Damalas & Koutroubas, 2022). However, the advantages of rapid early growth in a given environment rely on the water availability, as rapid early growth may reduce soil water reserves throughout the reproductive establishment and filling period, particularly in late flowering environments (Turner et al., 2001). In the current study, chickpea and field pea showed better crop establishment, as evidenced by their higher early DM than the other legumes. This rapid growth during the early stages set the potential for a high final biomass production.
Assimilate Translocation
Early DM accumulation has been linked to the capacity of source to translocate photosynthetic products to developing sink (French & Turner, 1991; Koutroubas et al., 1998; Leport et al., 1999), enabling plants to continue providing assimilates to the grain even when current assimilate supply is limited (Wardlaw, 1990). Such a situation was supported by the present study, in which species-specific variation in parameters related with assimilate translocation to seeds followed that of biomass accumulation before the onset of seed filling. Actually, for every kilogram per hectare increase in DM accumulation up to the growth stage R5, DMT increased by 45% and DMTE increased by 0.51%. However, apart from the amount of the available carbohydrates for remobilization before seed filling begins, the reallocation of carbon from vegetative organs to the seed can be influenced also by the seed size (Koutroubas et al., 2021), as well as by the ability of the seed to incorporate the translocated carbon in the embryo or endosperm (Turner et al., 2001). In this context, plants that produced a high number of seeds were found to remobilize more DM than plants with a lower number of seeds (Davies et al., 2000). Such an association was not confirmed in the current study, suggesting that species differences in DMT and seed size were unrelated. It seems that differences in seed size were probably only able to account for the variation in DMT within species, but not between species, given that different species had different DMTE. Indeed, in the present study, DMTE varied greatly between species, ranging from 24% to 51% the favorable year and from 9% to 32% in the drier year. As the DMTE increased, so did the DMT, indicating that the efficiency of legumes to remobilize assimilates to seeds was a decisive parameter in determining the net amount of DMT.
Although the ability of grain legumes to remobilize stored assimilates may be an adaptation to rainless periods (Leport et al., 1999), photosynthetic products assimilated before podding may improve the availability of assimilates to the seed during the filling stage in both well-watered plants and plants experiencing water stress (Davies et al., 2000). In the current study, high legume yields in the favorable year appeared to be more reliant on assimilate translocation to seeds compared with low yields in the less favorable year. In addition, the drier year, species with large DMT (such as field pea, red pea, and chickpea) produced high seed yields, which highlights the importance of translocation in maintaining the supply of assimilates to the seeds under adverse conditions.
Seed Yield and Harvest Index
Species showed substantial variation in seed yield, but their ranking fluctuated throughout the years because of the year by species interaction. Actually, species accounted for nearly half (50.65%) of the overall seed yield variation, while the year of the experimentation contributed about one-third (36.35%) and the year by species interaction made a small contribution (6.53%). Regardless of the growing conditions, red pea produced a comparatively high seed yield. Additionally, chickpea in the drier growing season and field pea in the more favorable season both produced high seed yields, which were comparable to that of red pea. Finally, in terms of seed yield, lentil was ranked last and common vetch was ranked second from the end in both growing seasons. The superior seed yield of red pea agrees with previous studies showing that this crop has a substantial potential in Mediterranean environments with low annual rainfall (250–350 mm), owing to its resistance to water shortage and its suitability to poor soils (Hanbury et al., 2000). Furthermore, red pea had the highest individual seed weight (> 200 mg) among the legumes used in the present study, a yield component that probably contributed to its superior yield. On the other hand, lentil plants, as opposed to the other legumes, had a smaller and more determinate growth habit (Erskine & Goodrich, 1991), which led to a poor seed yield, in line with previous research in both subhumid (Ayaz et al., 2004) and Mediterranean environments (Siddique et al., 1999). Regarding the superiority of chickpea in the drier year, it was probably due to its deep roots that provide high level of drought resistance (Jacobsen et al., 2012). From this point of view, chickpea could be a good option in Mediterranean environments even as spring crop. Early planting was crucial for crop success in this case; depending on the growing season, March sowing yielded 48 to 80% greater seed yield than April sowing (Fotiadis et al., 2017). It has also been calculated that late spring sowing decreased the total seed number by 7–18.2 seeds per square meter per day of planting delay (Fotiadis et al., 2019). Concerning common vetch, a seed yield of approximately 160 kg ha−1 has been reported under Mediterranean conditions (Siddique & Loss, 1996), a value comparable to that of the present study. Despite its poor seed yield, common vetch appears to be capable of producing a satisfactory silage yield, based on DM accumulation data, giving the crop an interesting alternative use.
Seed yield of legumes increased linearly with their capacity to accumulate biomass either before podding or at maturity. Therefore, selecting legume species with enhanced potential to produce biomass and improving management strategies favoring the accumulation of DM could boost seed yield of the tested grain legumes. Actually, an increase in seed yield by 1 kg ha−1 went along with an increase in vegetative DM of only 0.26 kg ha−1, resulting in a gradual increase of HI. However, no significant correlation between seed yield and harvest index existed. It has been demonstrated that the cultivating environment influences the association of seed yield with DM accumulation and partitioning. While in subcontinental environments seed yield was associated with higher aboveground biomass, it was connected with greater harvest indices in Mediterranean environments (Annicchiarico, 2008).
Yearly and Cultivar Differences
Seed yield of grain legumes fluctuates largely between growing seasons, a fact that explains why farmers in Europe do not cultivate systematically these crops (Von Richthofen et al., 2006). In the current research, growth attributes and seed yield of grain legumes varied substantially between years because of the seasonal variations in environmental conditions, which are typical for Mediterranean environments. In general, grain legumes grew better and produced more seed yield in the year with high growing season rainfall. However, the reduction of seed yield due to the lack of rainfall varied by species, being above average (34%) for field pea (45%) and common vetch (36%), and bellow average for lentil (21%), chickpea (29%), and red pea (30%). In most studied traits, larger differences were observed between species than those within species. The cultivar differences in DMT for common vetch, red pea, and chickpea were noteworthy. From this point of view, the ability of some genotypes to remobilize more assimilates to growing seeds could be exploited for improving the overall crop adaptation. However, because in the current study each legume was represented by only two cultivars, additional research involving a large number of cultivars is necessary to examine within-species variation and the potential for enhancing seed yield through breeding.
Conclusions
Data from the present research offer helpful evidence on the agronomic performance and assimilate remobilization in five spring grain legumes under Mediterranean conditions, whereas most previous studies have focused only on comparisons within single species. Therefore, the study improves our understanding of the differences in growth attributes and reallocation of assimilate between legume species grown simultaneously in the same environment, which has never been studied before. A broader understanding of how different legume species perform under specific conditions provides farmers with more tools for environmental and economic decision-making that could increase the adoption of more profitable production practices. Growth attributes and seed yield varied greatly by legume species and cultivation year, reflecting the differences in adaptation of species in the study area, as well as the fluctuations in seasonal patterns of rainfall. Early dry matter accumulation was critical in establishing the base for the final dry matter production. Between the evaluated species, red pea was the best option as a spring crop for cold Mediterranean areas in terms of seed yield, regardless of the seasonal pattern of rainfall. Furthermore, chickpea in the drier year and field pea in the wetter year appeared to be good options generating seed yields comparable to that of red pea. Less promising was the lentil and common vetch, which exhibited low seed yields in both years. Differences between species in seed yield appeared to be closely associated with biomass production rather than harvest index. Translocation of assimilates to seed was dependent on the availability of assimilates prior to grain filling. When rainfall was limited, species with greater dry matter translocation tended to produce higher seed yield. The ability of some genotypes to remobilize more assimilates to growing seeds could be exploited for improving the overall crop adaptation.
Data Availability
The data that support the findings of this study are available from the corresponding author, upon reasonable request, subject to restrictions and conditions.
References
Annicchiarico, P. (2008). Adaptation of cool-season grain legume species across climatically‐contrasting environments of southern Europe. Agronomy Journal,100, 1647–1654.
Ayaz, S., McKenzie, B. A., Hill, G. D., & McNeil, D. L. (2004). Variability in yield of four grain legume species in a subhumid temperate environment I. Yields and harvest index. The Journal of Agricultural Science,142, 9–19.
Bouwman, A. F., Beusen, A. H. W., Griffioen, J., Groenigen, J. W. V., Hefting, M. M., Oenema, O., Van Puijenbroek, P. J. T. M., Seitzinger, S., Slomp, C. P., & Stehfest, E. (2013). Global trends and uncertainties in terrestrial denitrification and N2O emissions. Philosophical Transactions of the Royal Society B,368, 91–97.
Cernay, C., Ben-Ari, T., Pelzer, E., Meynard, J. M., & Makowski, D. (2015). Estimating variability in grain legume yields across Europe and the Americas. Scientific Reports,5, 11171.
Damalas, C. A., & Koutroubas, S. D. (2022). Weed competition effects on growth and yield of spring-sown white lupine. Horticulturae,8, 430.
Davies, S. L., Turner, N. C., Palta, J. A., Siddique, K. H. M., & Plummer, J. A. (2000). Remobilisation of carbon and nitrogen supports seed filling in chickpea subjected to water deficit. Australian Journal of Agricultural Research,51, 855–866.
de Visser, C. L. M., Schreuder, R., & Stoddard, F. (2014). The EU’s dependency on soya bean import for the animal feed industry and potential for EU produced alternatives. OCL-OL Corps Gras Li,21, D407.
Dutta, A., Trivedi, A., Nath, C. P., Gupta, D. S., & Hazra, K. K. (2022). A comprehensive review on grain legumes as climate-smart crops: Challenges and prospects. Environmental Challenges,7, 100479.
Erskine, W., & Goodrich, W. J. (1991). Variability in lentil growth habit. Crop Science,31, 1040–1044.
Estrada-Campuzano, G., Slafer, G. A., & Miralles, D. J. (2012). Differences in yield, biomass and their components between triticale and wheat grown under contrasting water and nitrogen environments. Field Crops Research,128, 167–179.
Eurostat. (2019). Crop production in EU standard humidity. European Commission, Brussels, Belgium. https://ec.europa.eu/ euros tat. Accessed 22 Feb 2021.
Falconnier, G. N., Vermue, A., Journet, E. P., Christina, M., Bedoussac, L., & Justes, E. (2020). Contrasted response to climate change of winter and spring grain legumes in southwestern France. Field Crops Research,259, 107967.
Fotiadis, S., Koutroubas, S. D., & Damalas, C. A. (2017). Sowing date and cultivar effects on assimilate translocation in spring Mediterranean chickpea. Agronomy Journal,109, 2011–2024.
Fotiadis, S., Koutroubas, S. D., & Damalas, C. A. (2019). Nitrogen utilization and yield determination of spring Mediterranean chickpea as influenced by planting date and environmental conditions. International Journal of Plant Production,13, 59–72.
French, R. J., & Turner, N. C. (1991). Water deficits change dry matter partitioning and seed yield in narrow-leafed lupins (Lupinus angustifolius L). Australian Journal of Agricultural Research,42, 471–484.
Gao, H., Zhang, C., van der Werf, W., Ning, P., Zhang, Z., Wan, S., & Zhang, F. (2022). Intercropping modulates the accumulation and translocation of dry matter and nitrogen in maize and peanut. Field Crops Research,284, 108561.
Gaspar, A. P., Laboski, C. A., Naeve, S. L., & Conley, S. P. (2017). Dry matter and nitrogen uptake, partitioning, and removal across a wide range of soybean seed yield levels. Crop Science,57, 2170–2182.
Geng, S., Tan, J., Li, L., Miao, Y., & Wang, Y. (2023). Legumes can increase the yield of subsequent wheat with or without grain harvesting compared to Gramineae crops: A meta-analysis. European Journal of Agronomy,142, 126643.
Guinet, M., Nicolardot, B., & Voisin, A. S. (2020). Nitrogen benefits of ten legume pre-crops for wheat assessed by field measurements and modelling. European Journal of Agronomy,120, 126151.
Hanbury, C. D., White, C. L., Mullan, B. P., & Siddique, K. H. M. (2000). A review of the potential of Lathyrus sativus L. and L. Cicera L. grain for use as animal feed. Animal Feed Science and Technology,87, 1–27.
Huang, M., Zhang, R., Chen, J., Cao, F., Jiang, L., & Zou, Y. (2017). Morphological and physiological traits of seeds and seedlings in two rice cultivars with contrasting early vigor. Plant Production Science,20, 95–101.
Hungria, M., & Vargas, M. A. T. (2000). Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crop Research,65, 151–164.
Jacobsen, S. E., Jensen, C. R., & Liu, F. (2012). Improving crop production in the arid Mediterranean climate. Field Crops Research,128, 34–47.
Koutroubas, S. D., Damalas, C. A., & Fotiadis, S. (2021). Safflower assimilate remobilization, yield, and oil content in response to nitrogen availability, sowing time, and genotype. Field Crops Research,274, 108313.
Koutroubas, S. D., Papakosta, D. K., & Gagianas, A. A. (1998). The importance of early dry matter and nitrogen accumulation in soybean yield. European Journal of Agronomy,9, 1–10.
Kurdali, F., Kalifa, K., & Al-Shamma, M. (1997). Cultivar differences in nitrogen assimilation, partitioning and mobilization in rain-fed grown lentil. Field Crops Research,54, 235–243.
Leport, L., Turner, N. C., French, R. J., Barr, M. D., Duda, R., Davies, S. L., Tennant, D., & Siddique, K. H. (1999). Physiological responses of chickpea genotypes to terminal drought in a Mediterranean-type environment. European Journal of Agronomy,11, 279–291.
López-Bellido, F. J., López-Bellido, R. J., Khalil, S. K., & López-Bellido, L. (2008). Effect of sowing date on winter kabuli chickpea growth and yield under rainfed Mediterranean conditions. Agronomy Journal,100, 957–964.
Mohapatra, P. K., Turner, N. C., & Siddique, K. H. M. (2003). Assimilate partitioning in chickpea (Cicer arietinum L.) in drought prone environment. Management of agricultural drought: Agronomy and genetic options (pp. 173–188). Science Publishers Inc.
Neugschwandtner, R. W., Wichmann, S., Gimplinger, D. M., Wagentristl, H., & Hp, K. A. (2013). Chickpea performance compared to pea, barley and oat in Central Europe: Growth analysis and yield. Turkish Journal of Field Crops,18, 179–184.
Plaza-Bonilla, D., Nolot, J. M., Raffaillac, D., & Justes, E. (2017). Innovative cropping systems to reduce N inputs and maintain wheat yields by inserting grain legumes and cover crops in southwestern France. European Journal of Agronomy,82, 331–341.
Sainju, U. M., Lenssen, A. W., Caesar-TonThat, T., Jabro, J. D., Lartey, R. T., Evans, R. G., & Allen, B. L. (2012). Dryland soil nitrogen cycling influenced by tillage, crop rotation, and cultural practice. Nutrient Cycling in Agroecosystems,93, 309–322.
Siddique, K. H. M., & Loss, S. P. (1996). Growth and seed yield of vetches (Vicia spp.) in south-western Australia. Australian Journal of Experimental Agriculture,36, 587–593.
Siddique, K. H. M., Loss, S. P., Regan, K. L., & Jettner, R. L. (1999). Adaptation and seed yield of cool season grain legumes in Mediterranean environments of south-western Australia. Australian Journal of Agricultural Research,50(3), 375–388.
Siddique, K. H. M., Regan, K. L., Tennant, D., & Thomson, B. D. (2001). Water use and water use efficiency of cool season grain legumes in low rainfall Mediterranean-type environments. European Journal of Agronomy,15, 267–280.
Sinclair, T. R., & Muchow, R. C. (1999). Radiation use efficiency. Advances in Agronomy,65, 215–265.
Stagnari, F., Maggio, A., Galieni, A., & Pisante, M. (2017). Multiple benefits of legumes for agriculture sustainability: An overview. Chemical and Biological Technologies in Agriculture,4(1), 1–13.
Thomson, B. D., & Siddique, K. H. M. (1997). Grain legume species in low rainfall Mediterranean-type environments II. Canopy development, radiation interception, and dry-matter production. Field Crops Research,54, 189–199.
Turner, N. C., Davies, S. L., Plummer, J. A., & Siddique, K. H. M. (2005). Seed filling in grain legumes under water deficits, with emphasis on chickpeas. Advances in Agronomy,87, 211–250.
Turner, N. C., Wright, G. C., & Siddique, K. H. M. (2001). Adaptation of grain legumes (pulses) to water-limited environments. Advances in Agronomy,71, 193–271.
Virk, A. L., Lin, B. J., Kan, Z. R., Qi, J. Y., Dang, Y. P., Lal, R., & Zhang, H. L. (2022). Simultaneous effects of legume cultivation on carbon and nitrogen accumulation in soil. Advances in Agronomy,171, 75–110.
Von Richthofen, J. S., Pahl, H., Bouttet, D., Casta, P., Cartrysse, C., Charles, R., & Lafarga, A. (2006). What do European farmers think about grain legumes. Grain Legumes,45, 14–15.
Wardlaw, I. A. (1990). The control of carbon partitioning in plants. New Phytologist,116, 341–381.
Zhang, D. S., Sun, Z. X., Feng, L. S., Bai, W., Yang, N., Zhang, Z., Du, G. J., Feng, C., Cai, Q., Wang, Q., Zhang, Y., Wang, R. N., Arshad, A., Hao, X. Y., Sun, M., Gao, Z. Q., & Zhang, L. Z. (2020). Maize plant density affects yield, growth and source-sink relationship of crops in maize/peanut intercropping. Field Crops Research,257, 107926.
Funding
Open access funding provided by HEAL-Link Greece.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Koutroubas, S.D., Damalas, C.A. & Fotiadis, S. Assimilate Remobilization in Five Spring Grain Legumes Under Mediterranean Conditions. Int. J. Plant Prod. 18, 121–130 (2024). https://doi.org/10.1007/s42106-023-00279-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42106-023-00279-8