
Abstract
The Ribota site (Ágreda, Soria, Spain) is a new locality in the Matute Formation (Tithonian–Berriasian) composed of several carbonate layers, outstandingly rich in macrovertebrate remains. Fossils show an unusual replacement of the original bioapatite by quartz, and are found as positive reliefs protruding from lacustrine limestone beds. This type of conservation has allowed the identification of around one hundred vertebrate bone accumulations in an outcrop of more than 10 hectares. Osteichthyans (articulated partial skeletons, cranial material, and isolated postcranial bones and scales), crocodylomorphs (disarticulated cranial material, isolated teeth, vertebrae and osteoderms), turtles (partial carapaces and plastra, but also isolated plates) and pterosaurs (cranial and appendicular elements) have been identified. Around 80 specimens have been collected and a preliminary study of part of the collection (35 specimens) has allowed the identification of at least 5 different taxa: Halecomorphi indet., Neoginglymodi indet., Goniopholididae indet., Testudinata indet., and Pterodactyloidea indet. This new site represents one of the few sites from this time interval preserved in a fully lacustrine environment, so these vertebrate assemblages are unique and composed of different animals that presumably lived around and within the lake. They are dominated by aquatic and amphibian vertebrates and was formed by attrition in this lacustrine environment, possibly far from the lake shoreline. These macrovertebrate assemblages provide new data about the diversity in the faunal ecosystems from the Jurassic/Cretaceous transition of the Iberian Basin Rift System.
Resumen
El yacimiento de Ribota (Ágreda, Soria, España) es una nueva localidad en la Formación Matute (Titoniense-Berriasiense) compuesta por varios estratos de calizas excepcionalmente ricos en restos de macrovertebrados. Los fósiles muestran una sustitución inusual del bioapatito original por cuarzo, y se encuentran como relieves positivos sobresaliendo de la superficie de las calizas lacustres. Este tipo de conservación ha permitido la preservación e identificación de alrededor de cien acumulaciones de huesos de vertebrados en una superficie de más de 10 hectáreas. Se han identificado peces osteíctios (esqueletos parciales articulados, material craneal y escamas y huesos postcraneales aislados), cocodrilomorfos (material craneal desarticulado, dientes aislados, vértebras y osteodermos), tortugas (caparazones y plastrones pasciales, pero también placas aisladas) y pterosaurios (elementos craneales y apendiculares). Se han recuperado alrededor de 80 especímenes y un estudio preliminar de parte de la colección (35 especímenes) ha permitido la identificación de al menos 5 taxones diferentes: Halecomorphi indet., Neoginglymodi indet., Goniopholididae indet., Testudinata indet. y Pterodactyloidea indet. Este nuevo yacimiento representa uno de los pocos yacimientos de este intervalo temporal conservados en un ambiente completamente lacustre, por lo que estas asociaciones de vertebrados son únicas y están compuestas por diferentes animales que presumiblemente vivieron alrededor y dentro del lago. Están dominadas por vertebrados acuáticos y anfibios, y se formaron por atrición en este entorno lacustre, posiblemente dentro del lago y lejos de la orilla. Estas asociaciones de macrovertebrados proporcionan nuevos datos sobre la diversidad faunística en los ecosistemas presentes durante la transición Jurásico/Cretácico del Sistema de Rift de la Cuenca Ibérica.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
1 Introduction
The sedimentary successions deposited during the Jurassic/Cretaceous transition in the eastern Cameros basin (north central Spain) are well known for their large number of tracksites. The diverse ichnofauna of the Berriasian deposits (Oncala Group) of the Huérteles Formation in the North of Soria province stands out. This region has such an ichnological richness, (containing more than 7,000 vertebrate footprints produced mainly by dinosaurs, but also from pterosaurs, crocodylomorphs and turtles [e.g., Fuentes Vidarte et al., 2005; Hernández Medrano et al., 2006; Castanera et al., 2018]), that it gave rise to the creation of the touristic project “Ichnite Route of Soria” (Barco et al., 2013; Castanera et al., 2018), where a selection of dinosaur tracksites are prepared for tourist visits. This contrasts with the very scarce osteological fossil record found in the geological units that span around the Jurassic/Cretaceous (Tithonian–Berriasian) transition of the Tera Group (Agreda, Magaña and Matute Fms) in this sector of the Cameros basin.
One of the Tithonian–Berriasian geological units from the eastern Cameros basin that has yielded some fossil bones is the Matute Fm (Salomon, 1982a, b;mez Fernández and Meléndez, 1994). The latter authors indicate that remains of fishes and reptiles are relatively frequent in this unit, however only two notable specimens have been described in this formation. One corresponds to an articulated ginglymodian actinopterygian fish described as Camerichthys lunae Bermúdez-Rochas & Poyato-Ariza, 2015, from the locality of San Andrés de San Pedro (northeast of Soria province). The second is the freshwater aquatic stem turtle Pleurosternon moncayensis Pérez-García et al., 2022 (represented by a partial skull and several postcranial remains of a pleurosternid), recovered from the surroundings of Ágreda (east of Soria province), and only two kilometers away from the site here studied. In addition, also within the eastern Cameros basin in the town of Tera (north of Soria), some dinosaur remains of similar age from the Magaña Fm have also been reported (Canudo et al., 2010), and an articulated fish assigned to Lepidotes sp. has been described from the Berriasian of the Valdeprado Fm (Pascual-Arribas et al., 2007). South of Soria, in the Torrelapaja subbasin, the vertebrate fossil site of La Atalaya (Bijuesca Fm) includes large sized sauropod and crocodylomorph remains preserved in palustrine facies, most probably formed during the Tithonian–Berriasian transition (Aurell et al., 2021). This scarce vertebrate fossil content contrasts with the relatively abundant osteological record (mostly sauropod dinosaurs) with similar age, found in the western Cameros basin (mainly in Burgos Province) (e.g., Torcida Fernández-Baldor et al., 2020), in other areas of the Iberian Range, such as the Maestrazgo and South Iberian basins (northwest of Valencia and southwest of Teruel provinces) (e.g., Ruiz-Omeñaca et al., 2004; Aurell et al., 2016; Campos-Soto et al., 2019), or in the northern Aquitaine Basin in SW France (Allain et al., 2022).
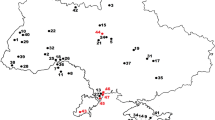
Here we present the new palaeontological site of Ribota (Ágreda, Soria province, Spain) (Fig. 1), which contains one of the most abundant vertebrate fossil assemblages of the whole Cameros basin. Until now, hundreds of bone remains (Figs. 2, 3, 4, 5, 6, 7 and 8) that correspond to osteichthyan fishes (Fig. 5), crocodylomorphs (Fig. 6), turtles (Fig. 7) and pterosaurs (Fig. 8) have been identified, and some of them, including all the remains described in this work, were recovered from the field. This paper aims to present a first approach to the genesis of this site (Figs. 3 and 4) and its palaeontological assemblage, in order to reconstruct the ecosystem of this area of the Cameros basin around the Jurassic/Cretaceous boundary.
2 Geographic and geological context
The Ribota fossil site is located 5 km northeast from the village of Ágreda (Soria Province, Castilla y León, north-central Spain), a town located about 10 km northwest of the Moncayo, the highest mountain of the Iberian Chain. The site outcrops in a sparsely vegetated area adjacent to the north of a meander of the Val River, about 300 m northwest of the “Pozo de las Truchas” waterfall.
Geologically, Ribota is located within the Cameros basin. This basin is found in the northwestern part of the Iberian Basin Rift System (Fig. 1A), which was formed due to extensional tectonics during the latest Jurassic–Early Cretaceous rifting stage. This rifting has been related to the opening of the Western Tethys Sea and the Northern Atlantic Ocean (see Salas et al., 2001; Mas et al., 2002, 2004, 2011; Aurell et al., 2019, 2021 and references therein). The Ribota site is located in the easternmost part of the eastern Cameros basin (Fig. 1B, C).

A) Geographic and geological location of the Ribota site (Soria, Castilla y León, Spain) within the Iberian Peninsula; B) geological location of the site within the Cameros basin, modified from Rodríguez-Barreiro et al. (2022); C) detailed geological map of the area and its stratigraphic framework, modified Gómez Fernández and Meléndez (1994)
The depocenter of the Cameros basin is filled with more than 8,000 m of Upper Jurassic–Lower Cretaceous lacustrine/coastal carbonates and evaporites alternated with fluvial siliciclastic deposits (Salas et al., 2001; Mas et al., 2004). Tischer (1996) divided this succession into five groups (Tera, Oncala, Urbión, Enciso, and Oliván). Successive genetic units were recognised, and the Tera Group was divided into three alloformations (Gómez-Fernández & Meléndez, 1994), that were subsequently used as lithostratigraphic units (e.g., Mas et al., 2004; Clemente, 2010; Martín-Chivelet el al., 2019). The lower Ágreda Fm consists of shallow marine, alluvial, lacustrine to palustrine deposits; the middle Magaña Fm is characterised by fluvial facies, and the upper Matute Fm (also known as the Sierra Matute Fm) is dominated by lacustrine deposits and includes the Ribota site studied in this work (Fig. 1C). This unit was originally interpreted as early Berriasian in age (Salomon, 1982a, b). The Berriasian age assignment for this unit, based on biostratigraphy, was also supported by Martín-Closas and Alonso Millán (1998), Salas et al. (2001), Mas and Salas (2002) or Clemente (2010). However, other authors considered the Matute Fm as Tithonian in age (Mas et al., 1993; Gómez-Fernández & Meléndez, 1994), although the top of the unit could even reach the beginning of the Berriasian (Gómez-Fernández, 1992; Meléndez & Gómez-Fernández, 2000). Accordingly, the precise age of this unit remains uncertain. The late Tithonian–earliest Berriasian age assumed here for the Matute Fm (Fig. 1C; see also Pérez-García et al., 2022) is based on the charophyte and ostracod content described in previous works (Gómez Fernández and Meléndez 1994; Tejero & Fernández-Gianotti, 2004; Schudack & Schudack, 2009).
The Ribota site is located in the lower part of the lacustrine carbonates of the Matute Fm (Fig. 1C), within a carbonate-dominated interval outcropping over a large area of about 10 hectares (Fig. 2A). Ribota is a rich locality where isolated bones are found in almost every square meter of the outcrop. Based on the closeness of several specific points with the higher bone concentrations, Ribota can be divided into three main areas (north, south, and east, Fig. 2A). The north and south areas are stratigraphically equivalent (almost the same strata), whereas the east area corresponds to slightly lower strata located about 150 m east from the other areas, but in the same formation and similar limestone facies. The division into 3 areas is provisional until a more exhaustive stratigraphic study is carried out. The most important fossiliferous accumulations are in the south area within the same stratigraphic level. Except for some articulated partial skeletons of fishes and turtles (exclusively represented by shell elements), most of the remains appear scattered and disarticulated along the different outcrops in the area.
The studied geological samples are very fossiliferous wackestone limestones, whereas the most common fossils are disarticulated ostracod valves, followed by vertebrate remains. All the samples were collected in the southern zone of Ribota, except for the RB72 sample that was recovered in the eastern area (Fig. 2A). All fossiliferous remains show a variable degree of recrystallization that might be related to the low-grade hydrothermal metamorphism that affected the Matute Formation (e.g., Barrenechea et al., 2001). The invertebrate remains have been completely replaced by silica (probably quartz), whereas the vertebrate remains partially retain their original apatite composition, being partially replaced by quartz and calcite. Isolated calcite and dolomite crystals can be found dispersed in a chlorite-dominated matrix, where clay mineral crystals show no preferred orientation. Secondary idiomorphic pyrite crystals of variable size, from sub-millimetric to centimetric, have grown both in the matrix and inside the bone elements. These pyrite crystals are partially oxidised, showing concentric rings of hematite and goethite, and present very small irregular inclusions of relics of apatite and calcite, incorporated into the crystal during growth.

A) Google Earth view of the Ribota site showing the different small outcrops with vertebrate remains; B) photograph and general view of the first site with vertebrate remains found in the east area, point “Fig. 1B” in the satellite photo in (A); C) fossiliferous point with a high concentration of disarticulated fish bones and a detail zoom of these remains (D) and a pyrite crystal pointed by a blue arrow; E) some pyrite crystals found in Ribota area
3 Materials and methods
To date, almost 80 vertebrate fossils have been recovered in the area. The material has been collected under the permit Exp. 37/2019-SO issued by the Dirección General de Patrimonio Cultural of the Junta de Castilla y León. Here we study only those samples that allowed a preliminary taxonomic identification (Table 1). The fossils, together with their surrounding rock matrix, were delimited in blocks cut with a radial saw and extracted with a chisel and hammer. Each collected sample has been labelled with the acronym RB (from Ribota) and the subsequent field number (RB1 to RB79), and they have been deposited (fossil numbers, nº 2023/6/RB1 to 2023/6/RB79) in the Museo Numantino, provincial de Soria (Soria, Spain). For simplicity, throughout the text, each specimen is just referred with the field number (RB1 to RB79).
Eight standard (30-micron thin) sections of selected bone specimens within limestone matrix (one Testudinata RB5; two Neoginglymodi RB11 and RB26; and four Crocodylomorpha RB6, RB52, RB72 and RB74) where prepared, in order to analyse the fossil diagenesis processes at the Ribota site. The thin sections were examined and photographed with an Olympus BX53M petrographic polarizing microscope equipped with a digital camera, housed at the IUCA Microscopy Lab at Universidad de Zaragoza. After this preliminary analysis, samples where carbon-coated and analysed in a Merlin Carl Zeiss Field Emission Scanning Electron Microscope housed at the Servicio de Apoyo a la Investigación (SAI) of the Universidad de Zaragoza. Chemical and textural information on the mineral phases, both in the fossils and in the encasing matrix, was obtained using Backscattered electron images (BSE) and Energy Dispersive X-Ray Analysis (EDS) and were acquired at 15 kV and an IProbe of 600 pA, at a working distance of 5.5 mm, using a Cobalt standard for calibration of the semiquantitative analysis.
4 Taphonomy
No meaningful differences concerning the lithology, abundance, and diversity of fossils, or taphonomic patterns have been observed between the sampled areas. The preliminary taphonomic analysis here reported, and supported by field and thin section observations, considers the working hypothesis that all of them represent a single bonebed (north and south areas), or successive bonebeds (e.g., east area), accumulated by similar non-differentiable processes. Thus, all the specimens will be analysed as coming from a single locality.
The fossil taphonomic history of the site is unusual and interesting. In the field, the fossils outcrop as dark brown to black positive reliefs protruding from the limestone, harder than the rock, and showing a very well-preserved external morphology. However, when broken, the bone tissue appears highly recrystallised, presenting conchoidal fractures, bright white and grey colours, and barely retaining its internal structure, making it almost indistinguishable from the encasing rock.
Bone remains appear distributed throughout the outcrop, without an appreciable distribution pattern, and no significant variation in the concentration of fossils is observed along the thickness of the studied section, within its several bone-bearing levels. Nevertheless, some areas show a relative increase in fossils elements, specifically of fossil fish bones (Fig. 2B-D) and some accumulations of crocodylomorph remains (Fig. 6F). Most fossils are bone fragments, but complete disarticulated elements are also frequent, and associated remains of crocodylomorphs and articulated remains of Testudinata and Osteichthyes have also been identified. No preferent orientation of long or flat bones has been observed on the field, and there is no apparent classification by size or shape of the specimens. Individual bones show various degrees of preservation, from bone splinters and fragments, to complete delicate elements such as a pterosaur limb and cranial bones. No evidence of subaerial exposure or weathering has been observed, including flaking or any presence of crusts or dissolutions. No plastic deformations have been identified in any of the specimens, and fractures caused by bone-to-bone contacts are also absent.
The fossil diagenetic history of the osteological elements analysed is of special interest, as all elements are heavily recrystallized. Concerning its geochemical composition, BSE images and EDX analyses show that the original bioapatite of the bones have been partially (Fig. 3 A, C, D) or totally (Fig. 4B, D, F) replaced by silica. Petrographic thin sections confirm that the SiO2 is actually crystalline quartz, which is characterized by white and brown interference colours and small to medium crystals with prismatic habits and blocky to undulating extinction, contrasting to the large, denim-blue fluorapatite crystals with fibrous habits. Contrary to what it is observed in hand samples, petrographic thin sections show that the internal structure of the bone tissue is relatively well preserved, despite the extensive mineralization. Bone growth lines can be observed, although are partially distorted. Occasional calcite and pyrite crystals have partially replaced the bone tissue (Fig. 3D) and are also present in the calcite matrix. Fractures, probably caused by lithostatic compaction of the specimens, are filled with calcite, quartz (which may be enriched with clay minerals), and occasionally iron oxides (Fig. 4E). The sediment infilling the bone cavities, such as medullary cavities or vascularization in osteoderms, is mainly micrite, and has abundant neo formed calcite crystals, chlorites, other silicate minerals, and euhedral pyrites. Some bones (especially those where the replacement by quartz has been completed) may show a chlorite coating.

Thin section of one scale of RB26 Neoginglymodi indet. (A) polarized transmitted light showing the preserved lamination of the histostructure of the scale, and diagenetic fractures infilled with quartz. (B) cross polarized transmitted light image of the same view as (A), showing that most of the bone has been replaced by quartz (brown and yellow interference colours), whereas only a small amount of the original fluorapatite is preserved in the bottom part of the scale (denim blue interference colours). Note that two different quartz phases can be observed in (A) but are indistinguishable in (B), one that is brown in (A) and another that is completely transparent suggesting the presence of an epitaxial growth of quartz after the bone replacement. White arrows point to recrystallized ostracod shells. (C) EDS map showing the elemental composition of the area in the white square in (A). Fluorapatite in light blue whereas quartz is shown in purple. Dark blue calcite infilled cracks can be observed in the top half of the map. Note the presence of small greenish pyrite crystal in the bottom part of the image. (D) BSE image of the squared area in 3D. The encasing sediment is a mixture of calcitic micrite and chlorite. Anatomical abbreviations: Cal., Calcite; Chl., Chlorite; F-Ap., Fluorapatite; Hl., Halite; Py., Pyrite; Qz., Quartz

A, C, D) RB6 Goniopholididae indet. thin section of the diaphysis of an undetermined long bone. B, D, F) RB52 Goniopholididae indet. thin section of a tooth cut at the base of the crown. (A) polarized transmitted light image showing the preserved lamination of the histostructure of the cortical and trabecular bone. Note both biomineralized tissues preserve the fibrous aspect of the bioapatite, and haversian channels can be observed in the trabecular bone. (B) polarized transmitted light image. Dentine of the teeth still shows lamination, whereas enamel is not present. Note fractures caused by diagenetic compression. (C) cross polarized transmitted light image. The general denim blue interference colour of the fluorapatite suggests the original composition of the bone is preserved, except for some relatively small areas were quartz, with a white interference colour, has replaced the structure. Large calcite crystals can be observed in the medullary cavity of the bone, together with indetermined silicate minerals of bright interference colours. (D) cross polarized transmitted light image, showing that the bone has been completely recrystallized by medium quartz crystals of brownish-yellowish interference colour. Drusy infillings are seen filling the fractures, both radially and around the teeth. The pulp cavity is filled with a calcite and chlorite matrix, similar to the encasing sediment. (E) BSE image. Note the presence of large areas preserving its original fluorapatite composition, interstratified with quartz, which has started the replacing process. Some cracks are filled with Iron oxides. Large calcite crystals with relics of fluorapatite grow in the medullary cavity. (F) BSE image of the teeth, completely replaced by quartz. A slightly lighter phase of quartz is filling the cracks, but no significative differences in composition were detected by EDS. The surface of the teeth, where the enamel should have been, has been replaced by chlorite. Anatomical abbreviations: Cal., Calcite; Chl., Chlorite; F-Ap., Fluorapatite; Fe-Ox., Iron oxides; Hl., Halite; Py., Pyrite; Qz., Quartz
No significative differences in mineralogical or chemical composition have been observed between the bones attributed to different taxa, suggesting that there is no taxonomic control of the diagenetic processes.
Preliminary taphonomic analysis allows the characterization of the assemblages as time-averaged, osteichthyan dominated, multitaxic bonebeds (especially the south area). The apparent lack of evidence of transport, with no preferred orientation or sorting of elements, together with the range of degrees of articulation of the specimens suggest that the assemblage formed by attrition and accumulation of bone fragments, and disarticulated and partially articulated skeletal remains in a lacustrine environment.
Despite the relatively small sample analysed, preliminary taphonomic and palaeoecological hypothesis about the composition of the assemblage can be postulated. The assemblage is dominated by obligate aquatic taxa (osteichthyan fishes), which represent the more articulated specimens. They are followed by amphibian organisms (Crocodylomorpha and Testudinata), which show partial articulation and some degree of association, suggesting that the environment was an extensive lake with a permanent water level, and the accumulation was produced far from the coast of the lake. This is also supported by the lack of fully terrestrial animals, outside of disarticulated elements of flying pterosaurs which would have been incorporated to the assemblage after falling to their deaths inside the lake.
The low degree metamorphism present in the Matute Fm has resulted in a very particular mode of preservation of the fossil bones in the Ribota bonebeds. Opalized vertebrate bones are known in the literature (e.g., Pewkliang et al., 2008), where the microstructure of the fossils is perfectly preserved at the level of the individual osteons. It seems that an analogue process may have occurred in the Ribota bonebed, which allowed the preservation of the detailed histostructure of the bone, but it was followed by a recrystallization of the opal into quartz. This process was also sufficiently slow to allow the survival of the bone laminations during this replacement. During the peak of low-grade metamorphism in the Cameros basin, the temperatures may have reached 350–370ºC and pressures of up to 1 Kbar (e.g., Casquet et al., 1992, Barrenechea et al., 2001), which probably caused this unusual preservation of the fossil bones. Further research is needed to explore the detailed mechanisms of the fossildiagenetic pathways in the context of low-grade metamorphism.
5 Vertebrate assemblage
In the outcrop, the most abundant remains correspond to isolated and disarticulated fish bones and scales (Fig. 2C, D), although some partial skeletons of indeterminate osteichthyans were also collected (Fig. 5). Other vertebrate remains include crocodylomorph isolated teeth, osteoderms, postcranial and a few cranial bones (Fig. 6); Testudinata isolated plates and partial shells (Fig. 7); and a few isolated cranial and appendicular pterosaur bones (Fig. 8). Interestingly, not a single dinosaur bone has been identified in the area so far.
5.1 Fishes
The Late Jurassic to the Early Cretaceous actinopterygian fish faunas from continental or transitional environments of Europe are commonly characterized by the presence of remains of ginglymodians and halecomorphs, together with other groups, such as pycnodontiforms or basal teleosts (e.g. Forey & Sweetman, 2011; Pouech et al., 2015; Ruiz-Omeñaca and Bermúdez-Rochas, 2010). The Early Cretaceous of the Cameros Basin is no exception and the presence of remains of these groups of fishes have also been recorded in the Enciso Group (Bermúdez-Rochas & Poyato-Ariza, 2007), with also abundant ginglymodian remains in the Urbion Group.
Fish faunas in the Ribota site are represented by abundant osteichthyan isolated remains (teeth, ganoid scales and bone fragments), and some incomplete articulated or semiarticulated remains. A preliminary study of these remains points to a low fish faunal diversity with specimens of two major groups of actinopterygians, namely: Halecomorphi and Neoginglymodi.
The Jurassic-Cretaceous boundary osteichthyan fishes record is scarce in Spain (Poyato-Ariza et al., 1999) and the data that the Ribota site could provide is potentially interesting in order to understand the distribution and evolution of these fish faunas during the development of the Iberian Basin Rift System.
Osteichthyes Huxley, 1880.
Actinopterygii Cope, 1887.
Neopterygii Regan, 1923.
Holostei Müller, 1844 (sensu Grande, 2010).
Halecomorphi Cope, 1872 (sensu Grande & Bemis, 1998).
Halecomorphi indet.

Fish remains from the Ribota fossil site. (A) left dentary RB13 of Halecomorphi indet. in lateral view; (B) possible right dentary RB27 of Neoginglymodi indet. in medial view; (C) possible dermopalatine fragment RB58 of Neoginglymodi indet. in ventral view; (D) slab of an articulated partial skeleton RB14 of Neoginglymodi indet. in lateral view; (E) articulated partial skeleton RB26 of Neoginglymodi indet. in lateral view; F and G) Isolated scales in the field assigned to Neoginglymodi indet
5.1.1 Description
Halecomorph fishes are represented by one isolated remain (RB13) corresponding to a partial left dentary with at least four incomplete teeth attached (Fig. 5A). The specimen was recovered in the east area (Fig. 2A) and partially exposed in lateral view, and as a result, the lateral surface of the dentary is quite damaged and abraded by erosion. Despite this, remains of the mandibular sensory canal can be observed. This canal traverses the bone parallel to its ventral margin, with three large pores visible, where the anterior and posterior ones are larger than the middle one. The dentary is elongated, robust and shallow, with a total length of 4.75 cm and a maximum height of approximately 1 cm at its posterior end. Teeth are only present in its anterior section. They are slightly conical and well-spaced, but poorly preserved (their tips are missing), without any presence of acrodine caps or potential carinate terminations. The two anterior teeth are slightly posteriorly curved.
Holostei Müller, 1844 (sensu Grande, 2010).
Ginglymodi Cope, 1872 (sensu Grande, 2010).
Neoginglymodi López-Arbarello & Sferco, 2018.
Neoginglymodi indet.
5.1.2 Description
Neoginglymodian fishes’ remains are abundant in the Ribota Site, with the presence of scales, teeth, and bones. Most of them have been located isolated and scattered all over the site, but several articulated or semiarticulated remains have been recovered. Among them, there are some cranial and postcranial bones, at least three mandibular bones with teeth attached, and some sets of scales in connection. Most of the specimens are partially covered by limestone and their exposed surfaces are damaged by meteorization, so further preparation is required in order to study them properly.
Two specimens (RB27 and RB58) are bone fragments with several teeth in connection (Fig. 5B, C). These specimens are incomplete and still partially covered by limestone, so it is difficult to identify them. One specimen seems to correspond to a right dentary (RB27, Fig. 5B) and the other one could be a dermopalatine (RB58, Fig. 5C). Both specimens show some degree of heterodonty, as the inner teeth are more robust and bigger than the marginal ones. In the dentary the posterior teeth are about twice the size of the anterior ones. All teeth are rounded to oval in section and present a conspicuous papilla in the centre of their occlusal surface. This character is also present in the teeth of Camerichthys lunae (Bermúdez-Rochas & Poyato-Ariza, 2015).
The abundant scales recovered are ganoid type scales, approximately rhomboidal in shape (RB14, RB26, Fig. 5D, E, F, G). There is no evidence of surface ornamentation or serration in their posterior borders, but the ganoine surface of exposed scales is heavily worn, and most of their posterior borders are broken. Despite this, a variation in their general morphologies, that corresponds to the different body region that they occupied, can be observed. Based on this morphology, in the Ribota site there are scales corresponding to all the regions of the body. Among the morphologies observed, at least one scale corresponds to the dorsal ridge, and seems to bear a short spine (Fig. 5F), as it occurs in the dorsal spines of C. lunae (Bermúdez-Rochas & Poyato-Ariza, 2015). The so-called peg-and-socket articulation (typical of ganoid scales) is not visible in the remains exposed. However, some specimens show a well-developed rostro-caudal articulation, with the corresponding two anteriorly oriented processes (Fig. 5G).
5.1.3 Remarks
Halecomorph fishes are common in the Jurassic and Cretaceous faunas of Spain (i.e., Wenz, 1995; Ruiz-Omeñaca et al., 2006; Martín-Abad & Poyato-Ariza, 2017) and, as commented above, remains of this group, assigned to Amiiformes, have been previously cited in the Early Cretaceous of the Cameros basin (Bermúdez-Rochas & Poyato-Ariza, 2007). Their finding in the Ribota site represents their first record in the Matute Formation, and their oldest record in the basin. Unfortunately, the preservation condition of the only specimen recovered does not allow a higher taxonomic precision to be achieved at this time.
The presence of isolated ganoid scales in the Matute Fm was recorded for the first time in the Virgen del Prado site (Inestrillas-Aguilar del Río Alhama, La Rioja, Spain) by Moratalla (1993), and assigned to the genus Lepidotes at the time. After the revised classification of the ginglymodian fishes made by López-Arbarello (2012), the assessment of these kind of isolated ganoid scales to the genus Lepidotes can no longer be considered valid (see Bermúdez-Rochas & Poyato-Ariza, 2015). More than twenty years later, the taxon Camerichthys lunae was erected in the San Andrés de San Pedro Site (Soria), based on an articulated specimen with the skull and the anterodorsal portion of the body preserved (Bermúdez-Rochas & Poyato-Ariza, 2015). The rostro-caudal articulation present in the Ribota site ganoid scales is known in several ginglymodian genera, such as Callipurbeckia or Scheenstia, belonging to the orders Semionotiformes and Lepisosteiformes, respectively (see López-Arbarello & Sferco, 2018). Therefore, although they present similarities with C. lunae, the attribution of these scales to this taxon cannot be assured, and a higher taxonomic assignment cannot be made until more complete material is recovered.
5.2 Crocodylomorpha
The crocodylomorphs of the Late Jurassic and Early Cretaceous of Europe are dominated by the Neosuchia, a clade of mesoeucrocodylian crocodyliformes, with their oldest record in the Early Jurassic (Sinemurian) of North America (Tykoski et al., 2002). The group survived to the present with the crown group Crocodylia as the clade that includes all extant species and their closest fossil relatives. In terrestrial to transitional environments from the Middle Jurassic to the Early Cretaceous of Europe, the most typical crocodylomorph fossil assemblage is dominated by the neosuchian clades Atoposauridae, Bernissartiidae, and Goniopholididae (e.g., Brinkmann, 1989; Buscalioni et al., 2008; Schwarz-Wings et al., 2009; Gasca et al., 2012; Puértolas-Pascual et al., 2015b; Tennant et al., 2016a; Guillaume et al., 2020). However, deposits with crocodylomorphs from the Jurassic–Cretaceous transition are still scarce, giving the Ribota site greater interest. At this site, the most abundant reptiles correspond to crocodylomorphs remains (Fig. 6), which represent more than half of the collected bones.
Crocodylomorpha Hay, 1930 (sensu Walker, 1970).
Crocodyliformes Hay, 1930.
Mesoeucrocodylia Whetstone & Whybrow, 1983.
Neosuchia Gervais, 1871 (sensu Benton & Clark, 1988).
Neosuchia indet.
5.2.1 Description
Most of these crocodylomorph remains correspond to isolated vertebrae (Fig. 6A-E), however, some of them were recovered associated in an area of less than 1 m2 and presenting similar sizes (Fig. 6F), so they could belong to the same individual. Since most of the vertebrae are partially eroded and/or contained within the rock matrix, some details have been hidden. However, their general morphology, shape of the vertebral centra, and the position of the ribs, diapophyses and parapophyses (when these are preserved) have allowed for assigning them to different regions of the vertebral column. Both in anterior and posterior views, the articular surfaces of the centra are sub-circular and gently concave due to the presence of a shallow central depression (amphicoelous condition), and they are almost as dorsoventrally tall as they are mediolaterally wide. The ventral and lateral surfaces of the centra are slightly concave, giving them the typical hourglass shape. At least one cervical vertebra has been recovered (RB57; Fig. 6A). This vertebra is identified by the shape and position of the diapophyses and parapophyses. The presence of parapophyses on the lateral surfaces of the centrum (but in its dorsal half), would indicate that it is one of the last posterior cervical vertebrae (e.g., Mook, 1921; Puértolas-Pascual et al., 2015a). The most abundant vertebrae correspond to the dorsal region of the trunk (RB44; Fig. 6B), and they have been identified based on the presence of long transverse processes located dorsally in the vertebral centrum and at the same height as the neural arch. A sacral vertebra has also been identified (RB69; Fig. 6C). It has been recognised as a sacral element due to the presence of a very stout fused rib (also referred to as the transverse process) (Gomes de Souza, 2018) attached to the lateral margin of the vertebral centrum (Mook, 1921). Whether it belongs to the first or second sacral vertebra cannot be confirmed with certainty since it is incomplete and partially covered by matrix. At least one caudal vertebra (RB21; Fig. 6D) from the anterior or middle region of the tail has been identified. Although it is not complete, this caudal vertebra is characterized by the presence of an elongated vertebral centrum with a more quadrangular articular surface, and transverse processes (= fused caudal ribs according to Gomes de Souza, 2018) arising from the dorsal region of the centrum rather than from the neural arch. In addition, the ventral region of this centrum appears to possess two articulation facets for the chevrons attached to two subtle anteroposterior parallel ridges (Mook, 1921). Interestingly, some small isolated centra (RB66; Fig. 6E), (tentatively assigned to caudal vertebrae) were found disarticulated by the opened neurocentral suture. If these vertebrae are confirmed to be caudal, they would indicate the presence of juvenile individuals in addition to adults, since juveniles fuse their caudal vertebrae very early in ontogeny (Brochu, 1996; Ikejiri, 2012).
Neosuchia Gervais, 1871 (sensu Benton & Clark, 1988).
Goniopholididae Cope, 1875.
Goniopholididae indet.

Crocodylomorph remains from the Ribota fossil site. (A) cervical vertebra RB57 in anterior view; (B) dorsal vertebra RB44 in anteroposterior view; (C) sacral vertebra RB69 in anteroposterior view; (D) caudal vertebra RB21 in lateral view; (E) caudal vertebra RB66 in lateral view; (F) disarticulated vertebrae (RB74-78) and osteoderms highlighted in red that could correspond to the same individual; (G) dorsal osteoderm RB61 in dorsal view; (H) ventral osteoderm RB62 in ventral view; (I) ventral osteoderm RB35 in ventral view; (J) isolated tooth RB39; K) disarticulated cranial remains RB17. Anatomical abbreviations: al., alveolus; ce., centrum; cr., caudal rib; de., dentary; di., diapophysis; nc., neural canal; ns., neural spine; p.a., parapophysis; pfm., posterior foramen for the mandibular ramus of cranial nerve V; pre., prezygapophysis; tr., transversal process; sp., splenial; su., surangular. Unnumbered scale bars = 2 cm
5.2.2 Description
The second most abundant crocodylomorph elements at the site are osteoderms (Fig. 6G-I). Although most of them appear isolated and fragmented, making their identification difficult, some characteristics of the most complete ones allow to assign them anatomically and taxonomically. At least three positional types of osteoderms have been identified: dorsal, ventral, and appendicular. One of the largest and most complete osteoderms (RB61) has been interpreted as belonging to the dorsal region of the trunk. This dorsal osteoderm (Fig. 6G) is twice as wide as it is long, with a smooth non-ornamented articular facet in the anterior margin, an anterolateral peg, and the lateral margin ventrally deflected with a high angle. These osteoderms are typical of crocodylomorphs with biseriate armour (two dorsal rows) and a closed paravertebral bracing system (Salisbury & Frey, 2001; Puértolas-Pascual et al., 2015a; Puértolas-Pascual & Mateus, 2020). At least two ventral osteoderms (RB62 and RB35) have been identified (Fig. 6H, I). They are flat, equidimensional (as wide as they are long), polygonal with pentagonal to hexagonal contours, with straight edges, and present the typical ornamentation of big circular pits evenly distributed over their ventral surface. Although they were isolated, their edges present some crenulation, indicating their sutures with adjacent osteoderms, which would form a rather rigid ventral armour (Wu et al., 1996; Salisbury & Frey, 2001). Some small, flat, elliptical to subcircular osteoderms have been interpreted as belonging to the appendicular region. This type of osteoderm can be confused with those of the neck region, however the latter usually have a keel, at least in some goniopholidids and extant crocodylians (Wu et al., 1996).
Neosuchia Gervais, 1871 (sensu Benton & Clark, 1988).
Neosuchia indet.
5.2.3 Description
Among the cranial material recovered at the site, the most abundant elements are isolated teeth (RB39; Fig. 6J). All the teeth belong to the same generalist morphotype, consisting of conical crowns, with smooth mesial and distal carinae, and ornamentation consisting of parallel longitudinal basiapical striae. Only a group of disarticulated (but associated) cranial bones (RB17) have been recovered from the site (Fig. 6K). Although they are partially covered by rock matrix, they can be recognized as part of a mandible. A dentary fragment, probably from the symphyseal region, with five circular dental alveoli (two of them preserving tooth fragments) has been identified. In addition, these associated bones also include an indeterminate cranial element (probably a surangular fragment) and a splenial fragment in which two posterior perforations for the mandibular ramus of cranial nerve V seem to be present.
5.2.4 Remarks
Although the crocodylomorph material at the Ribota site is very fragmentary and most elements are isolated, taking it as a whole has made it possible to make a series of taxonomic inferences. Regarding the vertebrae, most of them belong to individuals of medium to large size. In addition, the presence of amphicoelous vertebral centra points to the presence of non-eusuchian taxa. Considering the known taxa in this type of palaeoenvironment, age and palaeobiogeographical region, these vertebrae most probably belong to neosuchian crocodylomorphs (e.g., Guillaume et al., 2020; Puértolas-Pascual & Mateus, 2020). The osteoderms are more informative, since the combination of dorsal osteoderms that are twice as wide as they are long, with peg and groove articulation and a ventrally folded lateral margin, together with the presence of polygonal ventral osteoderms, points to the clade Goniopholididae (e.g., Wu et al., 1996; Salisbury & Frey, 2001; Puértolas-Pascual & Mateus, 2020). Furthermore, the presence of generalist conical teeth is also typical of this clade of neosuchian crocodylomorphs, among others (Guillaume et al., 2020). Therefore, given the palaeogeographic/chronological context, the size of the bones, the morphology of the vertebrae and teeth, and the characteristic shape of the osteoderms, most of these remains can be provisionally assigned to Goniopholididae indet.
5.3 Testudinata
The most abundant and diverse lineage of turtles in the Upper Jurassic levels of Europe is that of Thalassochelydia (Anquetin et al., 2017). Several representatives of the group of littoral thalassochelydians Plesiochelyidae have been recognized in the Iberian Peninsula, as well as the freshwater Hylaeochelys, the latter being the only thalassochelydian surviving after the Jurassic–Cretaceous transition (see Pérez-García, 2017; and references therein). The North American and European lineage of the paracryptodiran turtles Pleurosternidae is recognized within both the Late Jurassic and the Early Cretaceous. The Iberian record of this group of freshwater basal turtles (i.e., members of Testudinata not attributable to the crown Testudines) is currently identified as the most diverse in Europe, with several Iberian representatives having been defined from the Kimmeridgian to the Albian levels (see Pérez-García et al., 2022; and references therein). Another lineage identified in Europe from both Upper Jurassic sites (where its record is very limited) and throughout the Cretaceous is that of the terrestrial stem turtles Helochelydridae. Although the Iberian Late Jurassic record includes other lineages of turtles, the current limited availability of material does not allow us to know which lineages they belong to. However, a diverse fauna of freshwater eucryptodiran turtles is recognized in Europe, and especially in the Spanish record, after the Jurassic–Cretaceous transition, probably from the Berriasian, as a result of diachronic dispersals to this continent (see Pérez-García, 2017; and references therein).
Testudinata Klein, 1760.
Testudinata indet.
5.3.1 Description
All turtle specimens so far identified in Ribota correspond to shell elements (Fig. 7). They are represented by isolated plates, both from the carapace (Fig. 7A-C) and from the plastron (Fig. 7D). Although most of these remains are not complete, they generally show a well-preserved outer surface, with a rough pattern (Fig. 7E). The material currently found does not show evidence for the presence of more than one taxon in Ribota.
In visceral view, the proximal end of the dorsal ribs is well-developed (RB18 and RB24), being relatively robust (Fig. 7A, B). Both the ventro-medial margin of the bridge peripherals RB16 (Fig. 7C), and the lateral one of the only hypoplastron RB20 found (Fig. 6D), indicate that the contact between the carapace and the plastron was at least partially ligamentous. The hypoplastron corresponded to that of a taxon with inframarginals, with at least two of these scutes overlapping this plate. The anterior hypoplastral margin is not perpendicular to the axial plane (that plane being recognized by the suture of this plate with the other hypoplastron), but rather it is postero-medially directed. Thus, this taxon lacked mesoplastra between the hyoplastra and the hypoplastra. The abdominal-femoral sulcus is latero-posteriorly directed.

Selection of turtle remains from the Ribota fossil site. A) visceral view of an almost complete costal RB18; B) anterior or posterior view of the proximal region of a costal RB24; C) bridge peripheral RB16, in dorsal view; D-D’) partial left hypoplastron RB20, in ventral view. Dotted lines in D’ indicate broken edges; continuous black lines correspond to the margins of the plate; and the border of the scutes are represented by thicker gray lines. Scute abbreviations: ab., abdominal; fe., femoral; inf., inframarginal; m., marginal
5.3.2 Remarks
As has been indicated in the Introduction, a turtle was described in Ágreda. It corresponded to a member of the Pleurosternidae (Paracryptodira), and was attributed to the new species Pleurosternon moncayensis, exclusively represented by its holotype (Pérez-García et al., 2022). That turtle was defined by both cranial and shell remains. The turtle plates from Ribota cannot be attributed to Pleurosternidae because the taxon identified from there lacks mesoplastra, as well as an ornamental pattern composed by pits or tubercles, and by sulci on the margins of the plates (Pérez-García et al., 2022). The characters described here (i.e., absence of mesoplastra, present of inframarginal scutes), are compatible with those of the members of the lineage of Thalassochelydia, very abundant and diverse in the Late Jurassic levels of Europe, from the Oxfordian to the Tithonian (Anquetin at al., 2017; Pérez-García & Ortega, 2022). However, most of its representatives (including all members of Plesiochelyidae, a lineage exclusive to Europe and well-represented in the Iberian record) are recognized as littoral forms, which is not compatible with the sedimentary environment interpreted for the Matute Alloformation. The only freshwater member of Thalassochelydia currently known is Hylaeochelys which, furthermore, is the only one that survived until the Early Cretaceous (Pérez-García et al., 2023). The material analyzed here is not attributable to this genus represented in the Iberian Peninsula, considering the absence of a slightly fluted outer surface of the plates. The limited availability of characters in the turtle material found at Ribota is also compatible with that of another lineage of freshwater forms recognized in Europe: Xinjiangchelyidae. Although this group has an Asian origin, it reached Europe during the Lower Cretaceous, at the Berriasian or Valanginian. It was a diverse lineage in the European Lower Cretaceous record and especially in that of the Iberian Peninsula, where several forms have been identified (see Pérez-García, 2017; and references therein). Therefore, although the current limited information on the Ribota turtles does not allow us their precise systematic attribution, they could correspond to a single form, compatible with Thalassochelydia and with Xinjiangchelyidae. The potential future discovery of new remains in this fossil locality will be necessary to perform a more precise systematic attribution. However, the so far identified remains in Ribota are relevant since they allow the identification of two turtle lineages in Ágreda, as they are not compatible with Paracryptodira.
5.4 Pterosauria
The time period of the Jurassic–Cretaceous transition is a transformative time for the pterosaur faunal composition, where non-pterodactyloids were already in the process of global extinction, and the Pterodactyloidea had already began their diversification, radiating geographically and taxonomically throughout the remainder of the Cretaceous (although the exact early evolutionary drivers behind this transition are still largely undetermined). The specific time periods of the Tithonian–Berriasian, occurring just after the peak of this marked change, were therefore still typified by residual rhamphorhynchids, but also largely occupied by anurognathids, pterodactylids and ctenochasmatids (Paul, 2022).
Today, the known Late Jurassic–Early Cretaceous pterosaur occurrences in Spain have mainly been represented by trackways (e.g., Lockley et al., 1995, 2008; Wright et al., 1998; Hernández Medrano et al., 2006; Pascual Arribas et al., 2015; Piñuela, 2015; Pascual Arribas and Hernández-Medrano, 2016), but also by disparate occurrences of fragmentary fossil bone material (e.g. Fuentes Vidarte & Meijide Calvo, 1996, 2010; Buffetaut, 1999; Ruiz-Omeñaca et al., 2004; Vullo et al., 2009). Most known trackway localities are found in the north and northeast of Spain, in the provinces of Asturias, La Rioja, and Soria, with some fossil bones also occurring in the latter two, and additionally in the provinces of Aragón, Valencia and Cuenca (e.g., Holgado et al., 2011). Largely these fossils consist of postcranial remains of isolated bones (mainly wing elements) and teeth, although cranial material has also been recovered, most notably in the holotype fossils of Prejanopterus curvirostris Fuentes Vidarte and Mejide Calvo, 2010 (Pereda-Suberbiola et al., 2012), Europejara olcadesorum (Vullo et al., 2012), and Iberodactylus andreui (Holgado et al., 2019).
The Ribota pterosaur assemblage is thus far represented by six isolated elements: preliminary identifications designate them as a humerus (RB29, Fig. 8A), a radius fragment (RB37, Fig. 8B), a femur (RB28, Fig. 8C), a digit IV phalanx I (RB55, Fig. 8D), an epiphyseal fragment (RB34, Fig. 8E), and a mandibular fragment (RB19, Fig. 8F).
Order Pterosauria Owen, 1842 (sensu Andres & Padian, 2020a).
Suborder Pterodactyloidea Plieninger, 1901 (sensu Andres and Padian, 2020b).
Pterodactyloidea indet.

Pterosaur remains from the Ribota fossil site. (A) humerus RB29 in ventral view; (B) Partial radius RB37 in indeterminate view; (C) Femur RB28 in posterior view; (D) a digit IV phalanx RB55 in indeterminate view (note that they are two different pictures superimposed; E) epiphyseal fragment RB34 in lateral view; F) mandibular fragment RB19 in dorsal view. Anatomical abbreviations: dpc., deltopectoral crest. Scale bars = 2 cm
5.4.1 Description
RB29 is a mostly complete left humerus (Fig. 8A), visible in ventral view. Based on humerus scaling with P. curvirostris (the closest taxon geographically, and similar morphologically) (Pereda-Suberbiola et al., 2012), the Ribota specimen RB29 is about 30% smaller and would have achieved about one-meter wingspan. The humerus is severely eroded along both articular ends, and especially in the area of the deltopectoral crest, which is seemingly placed distal to the ulnar crest (a condition of pterodactyloids according to Unwin & Martill, 2018). At the proximal end, the ulnar crest faces anterolaterally, with a distinct concavity between it and the region of the deltopectoral crest. The humeral shaft is straight, with an ovoid cross-section at its midpoint, and moderately expanded mediolaterally towards both articular ends.
Mandibular fragment RB19 (Fig. 8F) is one of the most informative pterosaur bones in the Ribota assemblage, and is exposed dorsally and laterally, with sediment obscuring its ventral surface. The fragment is very slender, with almost parallel margins and a very thin midline rectilinear groove. The exact position of the fragment along the length of the entire mandible is indeterminate (due to being broken off both ends), but it is likely almost very near the anterior-most end of the dentary. In the mandible fragment, a total of two partial and eight complete alveoli have been preserved. Posterior to the partial alveoli (one in each side), four complete and well-developed raised alveoli are visible on each side, directed anterolaterally. The alveoli are all ovoid, elongated anteroposteriorly, although they vary in diameter, alternating between larger and smaller along the tooth row. Each pair of alveoli located laterally to each other also present asymmetry in size, with a larger alveolus in one side of the mandible and a smaller one on the opposite side. Furthermore, each alveolus is not aligned at the same anteroposterior height as its lateral counterpart, giving an extra degree of asymmetry to the dental arrangement. In transverse section, the broken ends of the symphysis show a distinctive rounded-triangle cross-section, compressed laterally, with the pointed apex facing ventrally.
Suborder Pterodactyloidea Plieninger, 1901 (sensu Andres and Padian, 2020b).
Pteranodontoidea Kellner, 2003.
Pteranodontoidea indet.
5.4.2 Description
RB28 is a left femur (Fig. 8C) only visible from its posterior face, as it is still embedded in matrix. It is highly eroded on its articular ends, and in particular in the region of the greater trochanter (which has been entirely eroded away) and in the asymmetrical distal epiphyseal region. The shaft is ovoid in cross-section, relatively straight anteroposteriorly, and slightly bowed mediolaterally, also exhibiting a slight expansion in mediolateral width to the distal end. The femoral head is strongly offset and proximally directed, capped by a mushroom-shaped constriction of the neck, and well differentiated from the femoral shaft by an 158° angle.
5.4.3 Remarks
Although likely belonging to different individuals, since they were isolated and separated by several meters, it is not possible at the moment to determine if the pterosaur elements from Ribota belong to the same species, and since there are no overlapping elements in the assemblage, more sampling would be needed to ensure whether one or more than one species is here represented. The bones were identified as belonging to pterosaurs due to the hollowness of the bones and a cortical thinness of between 0.3 and 1.6 mm (e.g., Bennet, 2003). The preservation of these elements is excellent in terms of three-dimensionality with almost no taphonomic distortion, however significant exposure to the elements over time has led to extensive erosional damage or breakage along the bone surfaces, rendering certain features indistinguishable. Thus, some elements are not currently identifiable beyond the level of Pterosauria indet., such as a distal left radius fragment RB37 (Fig. 8B) (visible in anterior view, and missing large sections of cortical bone throughout the shaft and articular end), right digit IV phalanx I RB55 (Fig. 8D) (visible in ventral view, and although nearly complete, sustained the extensor tendon process being completely eroded away anterior to the ventral cotyle), and epiphyseal fragment RB34 (Fig. 8E) (which is to too highly eroded to glean sufficient morphological information, precluding any assignation). The remaining better-preserved elements, however, are more informative.
The humeral shaft RB29 is straight (as in pterodactyloid pterosaurs), unlike the typical curved shaft of basal pterosaurs (Unwin & Martill, 2018). Humerus RB29 is therefore provisionally assigned to Pterodactyloidea indet. Regarding the femur RB28, the 158° angle of its femoral head with respect to the shaft, is consistent with the Ornithocheiridae (Unwin, 2003). Also, based on this femoral neck-shaft angle greater than 135°, it also corresponds to the Pteranodontoidea of Kellner (2003) in this analysis.
As previously stated, the cranial material is more informative. Although it is significantly smaller in size, on first glance, RB19 shows distinct morphological similarities with Prejanopterus curvirostris, particularly in the triangular cross-section of the symphyseal area, however the alveoli on P. curvirostris are organized quite differently and more symmetrical than in RB19. P. curvirostris was assigned to the Pterodactylidae (Pereda-Suberbiola et al., 2012), however some authors have implied that it belongs instead to the Lonchodectidae (Witton, 2003; Paul, 2022), a contentious group. However, the alveolar margins of the Ribota specimen do not appear as pronouncedly raised as most lonchodectids (whose alveoli usually are elevated above the medial portion of the occlusal surface), nor is the midline groove as pronounced as the sulcus present in most lonchodectids (Unwin, 2001; Averianov, 2020). Mandibular fragment RB19 also shows morphological similarities with Aetodactylus halli Myers, 2010, which was assigned to the Ornithocheiridae (as is Ribota femur RB28), particularly in the labiolingually-compressed alveoli, but also differing in RB19’s more lateral positioning of the alveoli along the symphysis (a feature more commonly associated with the Ctenochasmatidae [Unwin, 2003]). Therefore, mandibular fragment RB19 is here conservatively assigned to the Pterodactyloidea (the suborder inclusive of all the afore-mentioned groups), but due to the differences observed with these other taxa, could potentially be further representative of a new taxon. A more detailed study of the specimen and the possibility of more material being recovered in the future are necessary to clarify its phylogenetic position.
Although the majority of elements in this preliminary assemblage of pterosaurs have overall been informatively limited by their physical preservation, the potential of both the further preparation of these specimens, coupled with future excavation efforts will doubtlessly yield a significant and valuable picture of the pterosaur palaeodiversity of the Ribota locality, as well as the overall landscape for the Pterosauria during the transitional Tithonian–Berriasian time period worldwide.
6 Implications for the palaeobiodiversity in the Iberian ecosystems during the Jurassic/Cretaceous boundary
The Jurassic–Cretaceous transition is a significant geological period, and considerable faunal and environmental changes occurred across the Tithonian–Berriasian transition (see Tennant et al., 2016b, 2017; Allain et al., 2022). Instead of representing a significant faunal turnover between the Tithonian and the Berriasian, these faunal changes are more related with environment shifts associated to a sea level regression at the end of the Jurassic (Allain et al., 2022). Among the groups described in the present paper, Tennant et al. (2016b) indicate that marine osteichthyans and shallow marine turtles show a considerable decrease across the boundary (see also Pérez-García, 2017 and references therein) whereas freshwater and terrestrial turtles, and goniopholidid crocodylomorphs, seem to be unaffected. Pterodactyloids increasingly flourished right after the boundary (Butler et al., 2013; Tennant et al., 2016b). In this regard, the Iberian Peninsula has yielded many vertebrate localities from the end of the Jurassic (Kimmeridgian–Tithonian) and Jurassic–Cretaceous transition (Tithonian–Berriasian) in different areas and is becoming a key area to understanding how those faunal changes occurred. Significant sites have been found in areas such as the Lusitanian Basin in Portugal (e.g., Martin & Krebs, 2000; Malafaia et al., 2010; Mateus et al., 2017, and references therein), the Asturian Basin (Piñuela, 2015, and references therein), and several areas of the Iberian Basin Rift System (mainly South Iberian and Maestrazgo basins) in Spain (see Aurell et al., 2016, 2019; Campos-Soto et al., 2017;, 2019, and references therein). It is also worth noting that adjacent to the Iberian Peninsula, the Aquitaine Basin has also yielded rich and diverse Tithonian and Berriasian vertebrate assemblages that show strong similarities with Iberian faunas (Buffetaut et al., 1989; Mazin et al., 2008; Vullo et al., 2014; Allain et al., 2022).
Among the Lusitanian localities with abundant osteological vertebrate remains, Guimarota (Martin & Krebs, 2000) and Andrés (Malafaia et al., 2010) in Leiria (Portugal) stand out. These two localities are characterized by a highly diverse vertebrate fauna including fishes, lizards, crocodylomorphs, pterosaurs, and dinosaurs among others. There are several Spanish areas with abundant evidence of these groups of vertebrates within this time interval (Kimmeridgian–Berriasian) such as the Asturian Basin (e.g., Lockley et al., 2008; Piñuela 2015) or the Cameros basin (Hernández-Medrano et al., 2006; Castanera et al., 2018), but their records are mainly tracksites, and bone remains are comparatively scarce. In the Asturian basin, osteological remains from different groups of vertebrates have been reported and mainly represented by dinosaurs, pterosaurs, crocodylomorphs, turtles, and fishes (e.g., Ruiz-Omeñaca and Bermúdez-Rochas, 2010; Ruiz-Omeñaca et al., 2006;, 2012; Rauhut et al., 2018). In the eastern Cameros basin, dinosaurs, pterosaurs, crocodylomorphs, and turtles are represented in the ichnological record (Hernández-Medrano et al., 2006; Castanera et al., 2018), and osteological fossils are also represented by isolated remains from different sites and from different vertebrate groups, such as fishes (Pascual-Arribas et al., 2007; Bermúdez-Rochas & Poyato-Ariza, 2015), turtles (Pérez-García et al., 2022), dinosaurs (Canudo et al., 2010), and pterosaurs (Fuentes Vidarte & Meijide Calvo, 1996. In the western Cameros basin and in the closely located Torrelapaja subasin, new sites with different identified taxa (dinosaurs and crocodylomorphs) have been recently discovered, but their study is currently under way (Torcida Fernández-Baldor et al., 2020; Aurell et al., 2021).
Thus, the Ribota assemblage with a total minimum number of 5 taxa (Halecomorphi indet., Neoginglymodi indet., Goniopholididae indet., Testudinata indet. and Pterodactyloidea indet.) represents one of the highest macrovertebrate accumulations in the eastern Cameros basin, providing new data regarding vertebrate diversity in lacustrine ecosystems during the Jurassic–Cretaceous transition of the Iberian Basin Rift System (IBRS). In this regard, the southern basins of the IBRS (Maestrazgo and South Iberian basins) have yielded many of the important vertebrate Spanish localities (with multiple sites) from the Jurassic–Cretaceous transition (e.g., Ruiz-Omeñaca et al., 2004; Royo-Torres et al., 2009; Suñer & Martín, 2009; Cobos et al., 2020), but these localities are mainly known for their dinosaur remains, whereas other groups of vertebrates are comparatively scarce or not studied in detail. It is also noteworthy that the Kimmeridgian–Berriasian deposits of the IBRS have yielded a low vertebrate diversity (see supplementary material in Aurell et al., 2016; Campos-Soto et al., 2017, 2019) when compared with other Hauterivian–Barremian localities, especially those where microvertebrate remains are abundant (Ruiz-Omeñaca et al., 2004; Canudo et al., 2010). Accordingly, the Ribota assemblage in particular, and the Matute Formation in general, open a new window towards the understanding of the vertebrate fauna during the latest Tithonian–earliest Berriasian of the Iberian Peninsula, as it shows a high concentration of macrovertebrates and a moderate number of identified taxa (6 with Pleurosternon moncayensis, Pérez-García et al., 2022). Although the number of taxa is not higher than other coeval sites on the IBRS, the absence of more taxa could be produced by the preliminary nature of this work, since only some of the best fossil remains recovered from the site were selected for the study. It is quite likely that additional field work, laboratory preparation, and further detailed studies of the material from Ribota site may increase the number of identified taxa. In addition, the Ribota site is a key site to providing new data about the assemblages preserved in the fully continental realm (see palaeobiogeographical reconstructions of the basin in Aurell et al., 2021) since the deposits of the Matute Formation in the area represent the deposition in a shallow carbonate lake with intense evaporation, and far from the marine influence (Gómez-Fernández and Melendez, 1994). This palaeoenvironment is compatible with the fauna identified so far, where the assemblage is dominated by obligate aquatic taxa (Neoginglymodi indet. and Halecomorphi indet.), amphibian organisms (Testudinata indet. and Goniopholididae indet.) and flying pterosaurs (Pterodactyloidea indet.) This palaeoenvironment is also possibly the reason why the assemblage is different to many of the mentioned sites in the IBRS that are preserved in continental to transitional units and dominated by dinosaurs.
7 Conclusions
The Ribota site represents a new locality that shows the highest bone concentration of macrovertebrate remains of the Tithonian–Berriasian sites within the eastern Cameros Basin. The fossil diagenesis of the deposit is interesting and particular, since the bones have suffered considerable taphonomic modifications in which the original structure has been replaced by quartz. The assemblage is formed by attrition in a lacustrine environment, and dominated by fragmented bones, but with complete bones and associated and articulated specimens (mainly fishes and partial turtle shells) also present. The identified vertebrate assemblage is composed of fishes (Halecomorphi indet. and Neoginglymodi indet.), crocodylomorphs (Goniopholididae indet.), turtles (Testudinata indet.) and pterosaurs (Pterodactyloidea indet.). Counting with the turtle Pleurosternon moncayensis, described in the same formation and a few hundred meters northwest of Ribota, there is a total of 6 minimum taxa identified within the Matute Fm. The study of the vertebrate record of the Matute Fm has been neglected for years, considering the abundant vertebrate remains that have been previously mentioned (Gómez-Fernández and Melendez, 1994). Therefore, the potential of the Ribota locality in particular, and the geological unit in general (e.g., Bermudez-Rochas and Poyato-Ariza, 2014; Pérez-García et al., 2022), makes these deposits key towards the understanding of the vertebrate ecosystems in the Jurassic–Cretaceous transition of the Iberian Basin Rift System (especially because this studied assemblage is palaeoenvironmentally different from the known coeval sites described within the IBRS). Nonetheless, this is just a preliminary study and a first step towards the understanding of how the fossils have been preserved, how many bonebeds exist in the area, and how the vertebrate diversity was around the lake (for which the number of taxa will probably increase with further studies). Therefore, the preparation of the material already collected and/or new recovery may provide a considerable increase in the information about these lacustrine assemblages.
References
Allain, R., Vullo, R., Rozada, L., Anquetin, J., Bourgeais, R., Goedert, J., Lasseron, M., Martin, J. E., Pérez-García, A., De Fabrègues, C. P., Royo-Torres, R., Augier, D., Bailly, G., Cazes, L., Despres, Y., Gailliègue, A., Gomez, B., Goussard, F., Lenglet, T., Vacant, R., Mazan, & Tournepiche, J. F. (2022). Vertebrate Paleobiodiversity of the early cretaceous (Berriasian) Angeac-Charente Lagerstätte (southwestern France): Implications for continental faunal turnover at the J/K boundary. Geodiversitas, 44(25), 683–752.
Andres, B., & Padian, K. (2020a). Pterosauria, R. Owen 1842. In de K. Queiroz, P. D. Cantino, & J. A. Gauthier (Eds.), Phylonyms (pp. 1201–1204). CRC Press.
Andres, B., Padian, K., Pterodactyloidea, F., & Plieninger (2020b). 1901. In K. de Queiroz, P. D. Cantino, & J. A. Gauthier (Eds.), Phylonyms (pp. 1205–1207). CRC Press.
Anquetin, J., Püntener, C., & Joyce, W. G. (2017). A review of the fossil record of turtles of the clade Thalassochelydia. Bulletin of the Peabody Museum of Natural History, 58, 31–369.
Aurell, M., Bádenas, B., Canudo, J. I., Castanera, D., García-Penas, A., Gasca, J. M., Martín-Closas, C., Moliner, L., Moreno-Azanza, M., Rosales, I., Santas, L., Sequero, C., & Val, J. (2019). Kimmeridgian–berriasian stratigraphy and sedimentary evolution of the central Iberian Rift System (NE Spain). Cretaceous Research, 103, 104153.
Aurell, M., Bádenas, B., Castanera, D., Gasca, J. M., Canudo, J. I., Laita, E., & Liesa, C. L. (2021). Latest jurassic–early cretaceous synrift evolution of the Torrelapaja Subbasin (Cameros Basin): Implications for Northeast Iberia palaeogeography. Cretaceous Research, 128, 104997.
Aurell, M., Bádenas, B., Gasca, J. M., Canudo, J. I., Liesa, C. L., Soria, A. R., Moreno-Azanza, M., & Najes, L. (2016). Stratigraphy and evolution of the Galve sub-basin (Spain) in the middle Tithonian–early Barremian: Implications for the setting and age of some dinosaur fossil sites. Cretaceous Research, 65, 138–162.
Averianov, A. O. (2020). Taxonomy of the Lonchodectidae (Pterosauria, Pterodactyloidea). Proceedings of the Zoologicaal Institute RAS, 324(1), 41–55.
Barco, J. L., Castanera, D., Canudo, J. I., Pascual, C., Rubio, C. J., & Rubio, C. (2013). Aula Paleontológica y Ruta de las Icnitas de Soria: Un espacio paleontológico musealizado con fines didácticos y turísticos. HER&MUS, 12, 132–138.
Barrenechea, J. F., Rodas, M., Frey, M., Alonso-Azcárate, J., & Mas, J. R. (2001). Clay diagenesis and low-grade metamorphism of Tithonian and Berriasian sediments in the Cameros Basin (Spain). Clay Minerals, 36(3), 325–333.
Bennett, S. C. (2003). Morphological evolution of the pectoral girdle of pterosaurs: Myology and function. In E. Buffetaut, & J. M. Mazin (Eds.), Evolution and paleobiology of Pterosaurs (pp. 191–215). Geological Society.
Benton, M. J., & Clark, J. M. (1988). Archosaur phylogeny and the relationships of the Crocodylia. The Phylogeny and Classification of the Tetrapods, 1, 295–338.
Bermúdez-Rochas, D. D., & Poyato-Ariza, F. J. (2007). New fossiliferous sites with fish fauna from the basque-cantabrian and Cameros basins, early cretaceous of Spain. Journal of Vertebrate Paleontology, 27(supplement to Number 3), 47A–48A. 67th Annual meeting of the Society of Vertebrate Paleontology. Austin, Texas.
Bermúdez-Rochas, D. D., & Poyato-Ariza, F. J. (2015). A new semionotiform actinopterygian fish from the Mesozoic of Spain and its phylogenetic implications. Journal of Systematic Palaeontology, 13(4), 265–285.
Brinkmann, W. (1989). Vorläufige Mitteilung über die Krokodilier-Faunen aus dem ober-jura (Kimmeridgium) der Kohlegrube Guimarota, bei Leiria (Portugal) und der Unter-Kreide (Barremium) von Uña (Provinz Cuenca, Spanien). Documenta Naturae, 56, 1–28.
Brochu, C. A. (1996). Closure of neurocentral sutures during crocodilian ontogeny: Implications for maturity assessment in fossil archosaurs. Journal of Vertebrate Paleontology, 16(1), 49–62.
Buffetaut, E. (1999). Pterosauria from the Upper cretaceous of Laño (Iberian Peninsula): A preliminary comparative study. Estudios del Museo de Ciencias Naturales de Álava, 14(Número especial 1), 289–294.
Buffetaut, E., Pouit, D., Rigollet, L., & Archambeau, J. P. (1989). Poissons et reptiles continentaux du Purbeckien de la région de Cognac (Charente). Bulletin de la Société géologique de France, 5(5), 1065–1069.
Buscalioni, A. D., Fregenal, M. A., Bravo, A., Poyato-Ariza, F. J., Sanchíz, B., Báez, A. M., Moo, C., Martín, O., Closas, C., Evans, S. E., & Marugán Lobón, J. (2008). The vertebrate assemblage of Buenache de la Sierra (Upper Barremian of Serrania de Cuenca, Spain) with insights into its taphonomy and palaeoecology. Cretaceous Research, 29, 687–710.
Butler, R. J., Benson, R. B., & Barrett, P. M. (2013). Pterosaur diversity: Untangling the influence of sampling biases, Lagerstätten, and genuine biodiversity signals. Palaeogeography Palaeoclimatology Palaeoecology, 372, 78–87.
Campos-Soto, S., Benito, M. I., Cobos, A., Caus, E., Quijada, I. E., Suarez-Gonzalez, P., Mas, R., Royo-Torres, R., & Alcalá, L. (2019). Revisiting the age and palaeoenvironments of the Upper jurassic–lower cretaceous? Dinosaur-bearing sedimentary record of eastern Spain: Implications for Iberian palaeogeography. Journal of Iberian Geology, 45(3), 471–510.
Campos-Soto, S., Cobos, A., Caus, E., Benito, M. I., Fernández-Labrador, L., Suarez-Gonzalez, P., Van Den Berghe, Q., Mas, I. E., Royo-Torres, R., R., & Alcalá, L. (2017). Jurassic Coastal Park: A great diversity of palaeoenvironments for the dinosaurs of the Villar del Arzobispo formation (Teruel, eastern Spain). Palaeogeography Palaeoclimatology Palaeoecology, 485, 154–177.
Canudo, J. I., Barco, J. L., Castanera, D., & Torcida Fernández-Baldor, F. (2010). New record of a sauropod in the jurassic–cretaceous transition of the Iberian Peninsular (Spain): Palaeobiogeographical implications. Paläontologische Zeitschrift, 84(3), 427–435.
Casquet, C., Galindo, C., González-Casado, J. M., Alonso, A., Mas, R., Rodas, M., García, E., & Barrenechea, J. F. (1992). El metamorfismo en la Cuenca de Los Cameros. Geogaceta, 11, 22–25. Geocronología e implicaciones tectónicas.
Castanera, D., Pascual, C., Canudo, J. I., & Barco, J. L. (2018). Bringing together research, geoconservation and reaching a broad public in the form of a geotourism project: The Ichnite Route of Soria (Spain). Geoheritage, 10(3), 393–403.
Clemente, P. (2010). Review of the Upper jurassic-lower cretaceous stratigraphy in western Cameros Basin, Northern Spain. Revista de la Sociedad Geológica de España, 23(3–4), 101–143.
Cobos, A., Alcalá, L., & Royo-Torres, R. (2020). The dinosaur route in El Castellar (Teruel, Spain): Palaeontology as a factor of territorial development and scientific education in a sparsely inhabited area. Geoheritage, 12(3), 52.
Cope, E. D. (1872). Observations on the systematic relations of the fishes. Proceedings of the American Association for the Advancement of Science, 2(20), 317–343.
Cope, E. D. (1875). Check-list of North American Batrachia and Reptilia with a systematic list of the higher groups and an essay on geographical distribution based on the specimens in the U.S. National Museum. Bulletin of the United States National Museum, 1, 1–104.
Cope, E. D. (1887). Zittel’s manual of Palaeontology. American Naturalist, 21, 1014–1019.
de Gomes, L. (2018). Comments on the serial homology and homologues of vertebral lateral projections in Crocodylia (Eusuchia). The Anatomical Record, 301(7), 1203–1215.
Forey, P., & Sweetman, S. C. (2011). 18. Bony fishes. In: English Wealden fossils (Batten, D. J. ed.). The Paleontological Association, London, 225–235.
Fuentes Vidarte, C., & Meijide Calvo, M. (1996). Restos de pterosaurios en el Weald de Soria (España). Studia Geologica Salmanticensia, 32, 15–22.
Fuentes Vidarte, C., & Meijide Calvo, M. (2010). Un nuevo pterosaurio (Pterodactyloidea) en el Cretácico Inferior de La Rioja (España). Boletín Geológico y Minero, 121(3), 311–328.
Fuentes Vidarte, C., Meijide Calvo, M., Meijide Fuentes, F., & Meijide Fuentes, M. (2005). El conjunto faunístico de la base del Cretácico Inferior de Soria (cuenca de Cameros, grupo Oncala) a través del análisis icnológico. Celtiberia, 99, 367–404.
Gasca, J. M., Badiola, A., Canudo, J. I., Moreno-Azanza, M., Puértolas, E., Huerta, P., & Torcida, F. (2012). In P. Huerta, T. Fernández-Baldor, F., & J. I. Canudo (Eds.), Zaragoza, España). Actas de las V Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno (pp. 159–172). Colectivo Arqueológico y Paleontológico de Salas, Salas de los Infantes. La asociación de vertebrados fósiles del yacimiento Pochancalo 1 (Valanginiense-Hauteriviense, Villanueva de Huerva.
Gervais, P. (1871). Remarques sur les reptiles provenant des calcaires lithographiques de Cerin. Comptes Rendus Académie Sciences Paris, 73, 603–607.
Gómez-Fernández, J. C. (1992). Análisis de la cuenca sedimentaria de los Cameros durante sus etapas iniciales de relleno en relación con su evolución paleogeográfica. Unpublished PhD thesis, Universidad Complutense de Madrid, 343 pp.
Gómez-Fernández, J. C., & Meléndez, N. (1994). Estratigrafía de la Cuenca de los Cameros (Cordillera Iberica Noroccidental, N de España) durante el tránsito Jurásico-Cretácico. Revista de la Sociedad Geológica de España, 7, 121–139.
Grande, L. (2010). An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy. Th e resurrection of Holostei. Copeia, 2010(2A), iii–x1. supplement.
Grande, L., & Bemis, W. E. (1998). A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy. An empirical search for interconnected patterns of natural history. Memoir of Society of Vertebrate Paleontology, 4, 1–690.
Guillaume, A. R., Moreno-Azanza, M., Puértolas-Pascual, E., & Mateus, O. (2020). Palaeobiodiversity of crocodylomorphs from the Lourinhã formation based on the tooth record: Insights into the palaeoecology of the late jurassic of Portugal. Zoological Journal of the Linnean Society, 189(2), 549–583.
Hay, O. P. (1930). Second bibliography and catalogue of the fossil vertebrata of North America. Carnegie Institute Wash Publications, 390, 1–1074.
Hernández Medrano, N., Arribas, P., Latorre, C., Macarrón, P., & Sanz Pérez, E. (2006). Contribución de los yacimientos de icnitas sorianos al registro general de Cameros. Zubía, 23-24, 79–120.
Holgado, B., Martínez-Pérez, C., & Ruiz-Omeñaca, J. I. (2011). Revisión actualizada del registro fósil de pterosauria en la Península Ibérica. In A. Pérez-García, F. Gascó, J. M. Gasulla, & F. Escaso (Eds.), Viajando a Mundos Pretéritos (pp. 183–194). Ayuntamiento de Morella.
Holgado, B., Pêgas, R. V., Canudo, J. I., Fortuny, J., Rodrigues, T., Company, J., & Kellner, A. W. A. (2019). On a new crested pterodactyloid from the early cretaceous of the Iberian Peninsula and the radiation of the clade Anhangueria. Scientific Reports, 9, 1–10.
Huxley, T. H. (1880). On the applications of the laws of evolution to the arrangement of the Vertebrata and more particularly of the Mammalia. Proceedings of the Zoological Society of London, 649–662.
Ikejiri, T. (2012). Histology-based morphology of the neurocentral synchondrosis in Alligator mississippiensis (Archosauria, Crocodylia). The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 295(1), 18–31.
Kellner, A. W. A. (2003). Pterosaur phylogeny and comments on the evolutionary history of the group. Geological Society London Special Publications, 217(1), 105–137.
Klein, I. T. (1760). Klassification und kurze Geschichte der vierfüßigen Thiere. Jonas Schmidt.
Lockley, M. G., Garcia-Ramos, J. C., Pinuela, L., & Avanzini, M. (2008). A review of vertebrate track assemblages from the late jurassic of Asturias, Spain with comparative notes on coeval ichnofaunas from the western USA: Implications for faunal diversity in siliciclastic facies assemblages. Oryctos, 8, 53–70.
Lockley, M. G., Logue, T. J., Moratalla, J. J., Hunt, A. P., Schultz, R. J., & Robinson, J. W. (1995). The fossil trackway Pteraichnus is pterosaurian, not crocodilian: Implications for the global distribution of pterosaur tracks. Ichnos, 4(1), 7–20.
López-Arbarello, A., & Sferco, E. (2018). Neopterygian phylogeny: The merger assay. Royal Society Open Science, 5(3), 172337.
Malafaia, E., Ortega, F., Escaso, F., Dantas, P., Pimentel, N., Gasulla, J. M., Ribeiro, B., Barriga, F., & Sanz, J. L. (2010). Vertebrate fauna at the Allosaurus fossil-site of Andrés (Upper jurassic), Pombal, Portugal. Journal of Iberian Geology, 36(2), 193–204.
Martin, T., & Krebs, B. (2000). Guimarota. A jurassic ecosystem (p. 155). Verlag Dr. Friedrich Pfeil.
Martín-Abad, H., & Poyato-Ariza, F. J. (2017). A new genus and species for the amiiform fishes previously assigned to Amiopsis from the early cretaceous of Las Hoyas, Cuenca, Spain. Zoological Journal of the Linnean Society, 181, 604–637.
Martín-Chivelet, J., López-Gómez, J., Aguado, R., Arias, C., Arribas, J., Arribas, M. E., Aurell, M., Bádenas, B., Benito, M. I., Bover-Arnal, T., Casas-Sainz, A., Castro, J. M., Coruña, F., de Gea, G. A., Fornós, J. J., Fregenal-Martínez, M., García-Senz, J., Garóano, D., Gelabert, B., Giménez, J., González-Acebrón, J., Guimerà, J., Liesa, C. L., Mas, R., Meléndez, N., Molina, J. M., Muñoz, J. A., Navarrete, R., Nebot, M., Nieto, L. M., Omodeo-Salé, S., Pedrera, A., Peropadre, C., Quijada, I. E., Quijano, M. L., Reolid, M., Robador, A., Rodríguez-López, J. P., Rodríguez-Perea, A., Rosales, I., Ruiz-Ortiz, P. A., Sàbat, F., Salas, R., Soria, A. R., Suarez-González, P., & Vilas, L. (2019). In C. Quesada, & J. T. Oliveira (Eds.), The late jurassic–early cretaceous rifting (pp. 170–250). Springer. The Geology of Iberia: A Geodynamic Approach, The Alpine Cycle, Volume 3.
Martín-Closas, C., & Alonso Millán, A. (1998). Estratigrafía y biostratigrafía (Charophyta) del Cretácico Inferior en el sector occidental de la Cuenca de los Cameros (Cordillera Ibérica). Revista de la Sociedad Geológica de España, 11, 253–270.
Mas, J. R., Alonso, A., & Guimerá, J. (1993). Evolución tectonosedimentaria de una cuenca extensional intraplaca: La cuenca finijurásica-eocretácica de Los Cameros (La Rioja-Soria). Revista de la Sociedad Geológica de España, 6(3–4), 129–144.
Mas, R., Benito, M. I., Arribas, J., Alonso, A., Arribas, M. E., Lohmann, K. C., González-Acebrón, L., Hernán, J., Quijada, E., Suárez, P., Omodeo, S. Evolution of an intra-plate rift basin: the Latest Jurassic-Early Cretaceous Cameros Basin (Northwest Iberian Ranges, North Spain). In: Post-Meeting Field trips Guidebook (, Arenas, C., Pomar, L., & Colombo, F. (2011). 28th IAS Meeting, Zaragoza. Sociedad Geológica de España. Geo-Guías 8, Zaragoza, pp. 117–154.
Mas, R., Benito, M. I., Arribas, J., Serrano, A., Guimerà, J., Alonso, A., & Alonso-Azcárate, J. (2002). La Cuenca de Cameros: Desde la extensión finijurásica-eocretácica a la inversión terciaria - implicaciones en la exploración de hidrocarburos. Zubía, 14, 9–64.
Mas, R., García, A., Salas, R., Meléndez, A., Alonso, A., Aurell, M., Bádenas, B., Benito, M. I., Carenas, B., García Hidalgo, J. F., Gil, J., & Segura, M. (2004). El Rift Mesozoico Ibérico: Segunda fase de rifting: Jurásico Supe- rior-Cretácico Inferior. In J. A. Vera (Ed.), Geología de España (pp. 503–510). SGE-IGME.
Mas, R., & Salas, R. (2002). Lower cretaceous of the Iberian Basin. In W. Gibbons, & T. Moreno (Eds.), The geology of Spain (pp. 284–288). Geological Society.
Mateus, O., Dinis, J., & Cunha, P. P. (2017). The Lourinhã formation: The Upper jurassic to lower most cretaceous of the Lusitanian Basin, Portugal–landscapes where dinosaurs walked. Ciências da Terra/Earth Sciences Journal, 19(1), 75–97.
Mazin, J. M., Pouech, J., Hantzpergue, P., & Lenglet, T. (2008). The Purbeckian site of Cherves-de-Cognac (Berriasian, early cretaceous, SW France): A first synthesis. Travaux et Documents des Laboratoires de Géologie de Lyon, 164(1), 68–71.
Meléndez, N., & Gómez-Fernández, J. C. (2000). Continental deposits of the eastern Cameros Basin (northern Spain) during tithonian–berriasian time. American Association of Petroleum Geologists Studies in Geology, 46, 263–278.
Müller, J. (1844). Ueber den Bau und die Grenzen der Ganoiden und uber das naturliche System der Fische. Bericht uber die zur Bekanntmachung Geeigneten Verhandlungen der Akademie der Wissenschaften Berlin, 1846, 117–216.
Mook, C. C. (1921). Notes on the postcranial skeleton in the Crocodilia. Bulletin of the American Museum of Natural History, 44, 67–100.
Moratalla, J. J. (1993). Restos indirectos de Dinosaurios del registro español. Paleoicnología de la Cuenca de Cameros (Jurásico Superior-Cretácico Inferior) y Paleoología del Cretácico Superior, Vol. 1. Tesis doctoral inédita. Universidad Autónoma de Madrid, 421 pp.
Myers, T. S. (2010). A new ornithocheirid pterosaur from the Upper cretaceous (Cenomanian–Turonian) Eagle Ford Group of Texas. Journal of Vertebrate Paleontology, 30(1), 280–287.
Owen, R. (1842). Report on British Fossil Reptiles, Part II. Proceedings of the 11th Meeting of the British Association for the Advancement of Science, Plymouth, UK, 24 July 1842 (pp. 60–204).
Pascual-Arribas, C., Canudo, J. I., Perez, S., Hernández-Medrano, E., Castanera, N., D., & Barco, J. L. (2015). On the validity of Pteraichnus palacieisaenzi, Pascual Arribas and Sanz Pérez 2000: New data from the Huérteles formation. Paläontologische Zeitschrift, 89, 459–483.
Pascual-Arribas, C., & Hernández-Medrano, N. (2016). Huellas de Pteraichnus en la Muela (Soria, España): Consideraciones sobre el icnogénero y sobre la diversidad de huellas de pterosaurios en la Cuenca de Cameros. Revista de la Sociedad Geológica de España, 29(2), 89–105.
Pascual-Arribas, C., Sanz Pérez, E., Hernández Medrano, N., & Macarrón, L., P (2007). Lepidotes sp. en la Aloformación Valdeprado del Cretácico Inferior (Berriasiense) de la cuenca de cameros (Cordillera Ibérica, Soria, España). Studia Geologica Salmanticensia, 43(2), 193–206.
Paul, G. S. (2022). The Princeton Field Guide to Pterosaurs (p. 184). Princeton University Press.
Pereda-Suberbiola, X., Knoll, F., Ruiz-Omenaca, J. I., Company, J., & Fernandez-Baldor, T., F (2012). Reassessment of Prejanopterus curvirostris, a basal pterodactyloid pterosaur from the early cretaceous of Spain. Acta Gologica Sinica, 86(6), 1389–1401.
Pewkliang, B., Pring, A., & Brugger, J. (2008). The formation of precious opal: Clues from the opalization of bone. The Canadian Mineralogist, 46(1), 139–149.
Piñuela Suárez, L. (2015). Huellas de dinosaurios y otros reptiles del Jurásico Superior de Asturias. PhD Thesis, Universidad de Oviedo, 326 pp.
Plieninger, F. (1901). Beiträge Zur Kenntnis der Flugsaurier Paläontographica, 48, 65–90.
Pouech, J., Mazin, J. M., Cavin, L., & Poyato-Ariza, F. J. (2015). A berriasian actinopterygian fauna from Cherves-de-Cognac, France: Biodiversity and palaeoenvironmental implications. Cretaceous Research, 55, 32–43.
Poyato-Ariza, F. J., Buscalioni, A. D., & Cartanyá, J. (1999). The Mesozoic record of osteichthyan fishes from Spain In: Mesozoic Fishes 2. Systematics and Fossil Record. Proceedings of the international meeting (Buckow, 1997) (Arratia, G., Schultze, H.-P. (eds.). Verlag Dr. Friedrich Pfeil, München, Germany, 505–533.
Pérez-García, A. (2017). The Iberian fossil record of turtles: An update. Journal of Iberian Geology, 43, 155–191.
Pérez-García, A., Camilo, B., & Ortega, F. (2023). New data on the poorly known jurassic record of the turtle Hylaeochelys (Thalassochelydia), based on new finds from Portugal. Diversity, 15, 167.
Pérez-García, A., Martín-Jiménez, M., Aurell, M., Canudo, J. I., & Castanera, D. (2022). A new Iberian pleurosternid (jurassic-Cretaceous transition, Spain) and first neuroanatomical study of this clade of stem turtles. Historical Biology, 34(2), 298–311.
Pérez-García, A., & Ortega, F. (2022). New finds of the turtle Plesiochelys in the Upper jurassic of Portugal and evaluation of its diversity in the Iberian Peninsula. Historical Biology, 34, 121–129.
Puértolas-Pascual, E., Canudo, J. I., & Sender, L. M. (2015a). New material from a huge specimen of Anteophthalmosuchus cf. escuchae (Goniopholididae) from the Albian of Andorra (Teruel, Spain): Phylogenetic implications. Journal of Iberian Geology, 41(1), 41–56.
Puértolas-Pascual, E., & Mateus, O. (2020). A three-dimensional skeleton of Goniopholididae from the late jurassic of Portugal: Implications for the Crocodylomorpha bracing system. Zoological Journal of the Linnean Society, 189(2), 521–548.
Puértolas-Pascual, E., Rabal-Garcés, R., & Canudo, J. I. (2015b). Exceptional crocodylomorph biodiversity of ‘La Cantalera’ site (lower barremian; lower cretaceous) in Teruel, Spain. Palaeontologia Electronica, 18, 1–16.
Rauhut, O. W., Pinuela, L., Castanera, D., Garcia-Ramos, J. C., & Cela, I. S. (2018). The largest european theropod dinosaurs: Remains of a gigantic megalosaurid and giant theropod tracks from the Kimmeridgian of Asturias. Spain PeerJ, 6, e4963.
Regan, C. T. (1923). The skeleton of Lepidosteus, with remarks on the origin and evolution of the lower neopterygian fishes. Proceedings of the Zoological Society of London, 93, 445–461.
Rodríguez-Barreiro, I., Santos, A. A., Arribas, M. E., Mas, R., Arribas, J., Villanueva-Amadoz, U., Fernández-Baldor, T., F., & Diez, J. B. (2022). The Jurassic–Cretaceous transition in the West Cameros Basin (Tera Group, Burgos, Spain): Sedimentological and palynostratigraphical insights. Cretaceous Research, 139, 105300.
Royo-Torres, R., Cobos, A., Luque, L., Aberasturi, A., Espílez, E., Fierro, I., González, A., Mampel, L., & Alcalá, L. (2009). High european sauropod dinosaur diversity during jurassic–cretaceous transition in Riodeva (Teruel, Spain). Palaeontology, 52, 1009–1027.
Ruiz-Omeñaca, J. I., Bermúdez-Rochas, D. D. Vertebrados fósiles (restos directos). In: V Congreso de Jurásico de España. Guía de campo (excursión B). Las sucesiones litorales y marinas restringidas del Jurásico Superior. Acantilados de Tereñes (Ribadesella) y de la playa de La Griega (Colunga) (García-Ramos, J. C., & Aramburu, C. (2010). (coords.), Servitec, Oviedo: 47–50.
Ruiz-Omeñaca, J. I., Canudo, J. I., Aurell, M., Bádenas, B., Barco, J. L., Cuenca-Bescós, G., & Ipas, J. (2004). Estado de las investigaciones sobre los vertebrados del Jurásico Superior y Cretácico Inferior de Galve (Teruel). Estudios Geológicos, 60, 179–202.
Ruiz-Omeñaca, J. I., García-Ramos, J. C., Piñuela, L., Bardet, N., Bermúdez-Rochas, D. D., Canudo, J. I., & Pereda-Suberbiola, X. (2006). Restos directos de vertebrados del Jurásico de Asturias. En: Fernández-Martínez, E. (ed.), XXII Jornadas de la Sociedad Española de Paleontología y simposios de los proyectos PICG 493, 503, 499 y 467. Libro de resúmenes. Universidad de León, León, 171–173.
Ruiz-Omeñaca, J. I., Piñuela, L., & García-Ramos, J. C. (2012). New ornithopod remains from the Upper jurassic of Asturias (North Spain). Fundamental, 20, 219–222.
Salas, R., Guimerà, J., Mas, R., Martín-Closas, C., Meléndez, A., & Alonso, A. (2001). Evolution of the mesozoic central Iberian Rift System and its cainozoic inversion (Iberian chain). Peri-Tethys Memoir, 6, 145–185.
Salisbury, S. W., & Frey, E. (2001). A biomechanical transformation model for the evolution of semi-spheroidal articulations between adjoining vertebral bodies in crocodilians. In G. C. Grigg, F. Seebacher, & C. E. Franklin (Eds.), Crocodilian biology and evolution (pp. 85–134). Surrey Beatty & Sons.
Salomon, J. (1982a). El Cretácico Inferior. In A. García (Ed.), El Cretácico de España (pp. 345–387). Universidad Complutense de Madrid.
Salomon, J. (1982b). Les formations continentales du Jurassique Supérieur - Crétacé Inferieur (Espagne du Nord – Chaînes Cantabriques et NW Ibérique). Mémoires Géologiques de l´Université de Dijon, 6, 1–227.
Schudack, U., & Schudack, M. (2009). Ostracod biostratigraphy in the lower cretaceous of the Iberian chain (eastern Spain)/Bioestratigrafia de ostracodos en el Cretacico Inferior de la Cordillera Iberica (este de Espana). Journal of Iberian Geology, 35(2), 141–168.
Schwarz-Wings, D., Rees, J., & Lindgren, J. (2009). Lower cretaceous mesoeucrocodylians from Scandinavia (Denmark and Sweden). Cretaceous Research, 30(5), 1345–1355.
Suñer, M., & Martín, M. (2009). Un nuevo yacimiento del tránsito Jurásico-Cretácico de Alpuente (Los Serranos, Valencia, España): Resultados preliminares. Paleolusitana, 1, 441–447.
Tejero, R., & Fernández-Gianotti, J. (2004). CD-Rom appendix. In J. A. Vera (Ed.), Geología de España. SGE-IGME.
Tennant, J. P., Mannion, P. D., & Upchurch, P. (2016a). Environmental drivers of crocodyliform extinction across the Jurassic/Cretaceous transition. Proceedings of the Royal Society B: Biological Sciences, 283(1826), 20152840.
Tennant, J. P., Mannion, P. D., & Upchurch, P. (2016b). Sea level regulated tetrapod diversity dynamics through the Jurassic/Cretaceous interval. Nature Communications, 7(1), 12737.
Tennant, J. P., Mannion, P. D., Upchurch, P., Sutton, M. D., & Price, G. D. (2017). Biotic and environmental dynamics through the late jurassic–early cretaceous transition: Evidence for protracted faunal and ecological turnover. Biological Reviews, 92(2), 776–814.
Tischer, G. (1966). Über die Wealden Ablagerung und die Tektonik der östlichen Sierra de los Cameros in der nordwestlichen Iberischen Ketten (Spanien). Beihefte zum Geologischen Jahrbuch, 44, 123–164.
Torcida Fernández-Baldor, F., Canudo, J. I., & Huerta, P. (2020). New data on sauropod palaeobiodiversity at the jurassic-cretaceous transition of Spain (Burgos). Journal of Iberian Geology, 46(4), 351–362.
Tykoski, R. S., Rowe, T. B., Ketcham, R. A., & Colbert, M. W. (2002). Calsoyasuchus valliceps, a new crocodyliform from the early jurassic Kayenta formation of Arizona. Journal of Vertebrate Paleontology, 22(3), 593–611.
Unwin, D. M. (2001). An overview of the pterosaur assemblage from the Cambridge Greensand (cretaceous) of Eastern England. Fossil Record, 4, 189–221.
Unwin, D. M. (2003). On the phylogeny and evolutionary history of Pterosaurs. In E. Buffetaut, & J. M. Mazin (Eds.), Evolution and paleobiology of Pterosaurs (pp. 139–190). Geological Society.
Unwin, D. M., & Martill, D. M. (2018). Systematic reassessment of the first Jurassic pterosaur from Thailand. Geological Society, London, Special Publications, 455(1), 181–186.
Vullo, R., Abit, D., Ballèvre, M., Billon-Bruyat, J. P., Bourgeais, R., Buffetaut, É., Daviero-Gomez, V., Garcia, G., Gomez, B., Mazin, J. M., Morel, S., Néraudeau, D., Pouech, J., Rage, J. C., Schnyder, J., & Tong, H. (2014). Palaeontology of the Purbeck-type (Tithonian, late jurassic) bonebeds of Chassiron (Oléron Island, western France). Comptes Rendus Palevol, 13(5), 421–441.
Vullo, R., Buscalioni, A. D., Marugan-Lobon, J., & Moratalla, J. J. (2009). First pterosaur remains from the Early Cretaceous Lagerstätte of Las Hoyas, Spain: palaeoecological significance. Geological Magazine, 146(6), 931–936.
Vullo, R., Marugán-Lobón, J., Kellner, A. W. A., Buscalioni, A. D., Gomez, B., de la Fuente, M., & Moratalla, J. J. (2012). A New Crested Pterosaur from the Early Cretaceous of Spain: The First European Tapejarid (Pterodactyloidea: Azhdarchoidea). PLOS ONE, 7(7), e38900.
Walker, A. D. (1970). A revision of the jurassic reptile Hallopus victor (Marsh), with remarks on the classification of crocodiles. Philosophical Transactions of the Royal Society of London B Biological Sciences, 257, 323–372.
Wenz, S. (1995). Les Amiidés du Crétacé inferieur du Montsec (Province de Lleida, Espagne): Urocles sauvagei (Vidal, 1915) synonime de vidalamia catalunica (Sauvage, 1903). Treballs del Museu de Geologia de Barcelona, 4, 5–13.
Whetstone, K. N., & Whybrow, P. J. (1983). A cursorial crocodilian from the Triassic of Lesotho (Basutoland), Southern Africa. Occasional Papers of the Museum of Natural History The University of Kansas, 106, 1–37.
Witton, M. P. (2003). Pterosaurs: Natural history, evolution, anatomy. Princeton University Press.
Wright, J. L., Barret, P. M., Lockley, M. G., & Cook, E. (1998). A review of the early cretaceous terrestrial vertebrate track – bearing strata of England and Spain. New Mexico Museum of Natural History and Science Bulletin, 14, 143–153.
Wu, X. C., Brinkman, D. B., & Russell, A. P. (1996). Sunosuchus junggarensis sp. nov. (Archosauria: Crocodyliformes) from the Upper jurassic of Xinjiang, people’s Republic of China. Canadian Journal of Earth Sciences, 33(4), 606–630.
Acknowledgements
E.P-P was supported by a postdoctoral contract (María Zambrano) funded by the Ministry of Universities of the Government of Spain through the Next Generation EU funds of the European Union. D.C has been supported by the Beatriu de Pinós postdoctoral programme (BP2017-00195) of the Government of Catalonia’s Secretariat for Universities and Research of the Ministry of Economy and Knowledge. D.D.B-R was supported by the project PGC2018-094034-B-C21 of the Spanish Ministry of Science and Innovation. M.M-A is supported by the MCIN/AEI/https://doi.org/10.13039/501100011033 and co-financed by the NextGeneration EU/PRTR, Ramón y Cajal contract RYC2021-034473, Fundação para a Ciência e a Tecnologia (grant number PTDC/CTA-PAL/31656/2017 and research unit UIDB/04035/2020; GeoBioTec); Fieldwork has been funded thanks to and ERG Grant of the European Association of Vertebrate Palaeontologists (EAVP). The study was subsidized in part by Project PID2021-122612OB- I00 of the Spanish Ministerio de Economía y Competitividad-ERDF, as well as by the Aragón Regional Government (Grupos de referencia Aragosaurus: Recursos Geológicos y Paleoambientales and E04_20R_FOCONTUR) and Unidad de Dinosaurios de Teruel financed by the Ministerio de Ciencia e Innovación (Gobierno de España). Authors would like to acknowledge the use of Servicio General de Apoyo a la Investigación-SAI, Universidad de Zaragoza. Thanks to Marian Arlegui Sánchez, director of the Museo Numantino, provincial de Soria (Spain). Special thanks to the geologist Miguel Bartolomé for discovering us the first fossils of the Ribota site. The field assistance of the Aragosaurus members Jara Parrilla, Manuel Pérez, Eduardo Medrano, Julia Galán and Martín Linares is greatly appreciated. Thanks to the reviewers Romain Vullo and Elisabete Malafaia for their helpful comments that greatly improved the manuscript.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Puértolas-Pascual, E., Aurell, M., Bermúdez-Rochas, D. et al. A new vertebrate assemblage from the Matute Formation of the Cameros Basin (Ágreda, Spain): implications for the diversity during the Jurassic/Cretaceous boundary. J Iber Geol (2023). https://doi.org/10.1007/s41513-023-00220-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s41513-023-00220-y