
Abstract
As a part of a larger study concerning the carbon sequestration capability by hazelnut orchards in Italy, we analyzed the total amount of carbon dioxide (CO2) removed over the year from the atmosphere through the net assimilation rates in two hazelnut orchards in Piedmont (i.e., the second region in Italy for surface and production). In particular, considering the key role played by the structural traits in affecting carbon sequestration potential, we assessed the impact of two different training systems widely diffused in the region: single trunk in orchardA and bush-like in orchardB. The results showed that plants in orchardA and orchardB sequestered 10.6 ± 1.8 and 25.7 ± 4.2 kg (CO2) plant−1 month−1, respectively. Higher CO2 sequestration in the plants in orchardB was due to their higher leaf area index relative to plants in orchardA. The mean CO2 sequestration from orchardA and orchardB per area was 4.25 ± 1.72 and 8.57 ± 3.41 Mg (CO2) ha−1 month−1, respectively. We also estimated the total amount of CO2 emission by the management over the entire production season in 157.335 kg CO2eq ha−1 by summing the contribution of diesel fuel, machinery and fertilization practices and considering that the total amount of CO2 sequestered by the two hazelnut orchards over the entire study period was estimated in 26 Mg (CO2) ha−1 in orchardA, and in 51 Mg (CO2) ha−1 in orchardB, they had an effective positive role as carbon sink at this local level.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
In the scenario of expected climatic change (temperature increases and water deficits) related to the greenhouse effect, there has been an increasing interest on the carbon (C) cycle in terrestrial ecosystems (Don et al. 2012). The global warming is now clearly recognized as a major threat to natural and socioeconomic systems, and the global community is searching for cost-effective ways to slow the build-up of atmospheric carbon dioxide (CO2) concentrations and minimize its impact (Kirschbaum 2006). The Kyoto protocol to the United Nations framework convention on climate change (1997) establishes the principle that carbon sequestration can be used by participating nations to help meet their respective net emission reduction targets for carbon dioxide (CO2) and other greenhouse gases. In fact, by sequestering a large amount of atmospheric C, terrestrial ecosystems are thought to offer a mitigation strategy for reducing global warming (Schimel et al. 2001). Specifically, the Article 3.4 of the Kyoto Protocol recommended, among the allowed activities, forest management, cropland management, grazing land management and re-vegetation (Smith 2004). In particular, among the cropland context, orchards in comparison with annual crops have structural characteristic allowing them to sequester significant quantities of atmospheric C for longer periods with smaller annual fluctuations (Smaje 2015). This different performance of the orchards is due to, on the one hand, their long life cycle, which allows them to accumulate C in permanent organs such as trunk, branches and roots and in the soil through rhizo-deposition and, on the other hand, to a low or null soil tillage, which preserves soil organic matter from mineralization (Scandellari et al. 2016). Nevertheless, in spite of the strategic role of orchards, their C potential has been only partially explored, with studies conducted regarding olive (Nardino et al. 2013; Sofo et al. 2005), palm (Navarro et al. 2008), apple (Zanotelli et al. 2015), peach (Sofo et al. 2005) and pear (Zhang et al. 2013).
The landscape of Mediterranean region is largely dominated by orchards (Aguilera et al. 2015), and one of the most widely diffused is represented by hazelnut orchard (Corylus avellana L). Among the Mediterranean region, Italy is one of the major hazelnut productions worldwide, being second only to Turkey. In Italy, the hazelnut production surface is located mainly in four regions: Lazio, Piedmont, Campania and Sicily, covering 98% of the entire Italian production area (Istat 2019).
The present work is part of a larger study on the carbon sequestration capability of hazelnut orchards, which first covered orchards in Lazio region, with data that have been extended to include the nearby Campania region (Granata et al. 2020). The current study is focalized on the role of carbon sinks in hazelnut orchards in Piedmont, that based on the latest updates on the surface devoted to this culture, ranks as the second region in Italy for surface and production (24,555 ha and 39,997 tons, respectively, data from Istat 2019). In Piedmont, the hazelnut cultivation became really important and expanded in the late 1980s (Godone et al. 2014). Over time, there has been a significant increase in hazelnut surfaces in Piedmont, with a stronger trend than in the rest of Italy which reflects an increased interest of landowners toward hazelnut and its economical potential (Valentini and Me 2002). In particular, the nut production is based on a single cultivar ‘tonda gentile delle Langhe,’ selected directly by farmers for its good environmental adaptability to Piedmont climate and for the excellent quality of the kernel (Valentini et al. 2014). In Piedmont, the traditional cultivation of this cultivar expects different training systems in open forms such as the bush or single trunk. In both cases, choosing the right distance between plants is necessary to ensure adequate ventilation and lighting as well as allowing the mechanized cultivation practices such as pruning and harvesting. Thus, based on the soil fertility, water availability and the training systems, the planting distance inside the hazelnut orchards may vary from 5 × 4 m (plant density 500 plants ha−1) to 6 × 6 m (plant density 278 plants ha−1) (Comunità Montana Alta Langa, 2009). The aim of this work was to analyze the CO2 sequestration capability of mature hazelnut orchards in Piedmont in two areas where hazelnut plants were subjected to two different training systems (bush vs single trunk), but similar in terms of plants densities (333 vs 400 plants ha−1). The objectives of the study were 1) to compare the CO2 sequestration in the two orchards with respect to the different training systems, 2) to measure the seasonal pattern of CO2 sequestration during the year and 3) to contextualize the obtain CO2 sequestration capability in the Italian scenario of hazelnut orchards and in respect to others agro-ecosystems.
2 Material and methods
2.1 Study site and plant materials
The study was carried out in the period May–October 2019 in two areas in Piedmont (north-western Italy) where C. avellana (cv. ‘tonda gentile della langhe’) orchards were widely cultivated. In particular, one orchard was located in Alba (CN, 44°42′ 13′′ N; 08°01′ 13′′ E) at 172 m a.s.l. and hereafter called orchardA, with plants planted at a 5 × 5 m spacing resulting in a density of 400 plants ha−1. The other orchard was located in Lequio Berra (CN, 44° 36′ 49′′ N; 08°06′ 11′′ E) at 715 m a.s.l. and hereafter called orchardB, with plants planted at a 6 × 5 m spacing resulting in a density of 333 plants ha−1. Regarding the training systems, plants were grown on a single trunk in orchardA, while the training system was bush-like (multi-stemmed plants with 4–6 separate stems) in orchardB. Ordinary horticultural care was applied in both production systems, in terms of floor and pest management, pruning and fertilization.
Three plots (400 m2 each) were randomly selected in each orchard and measurements carried out on four trees randomly selected in the central part of each plot, being sure that their apparent cropping performance and overall development fully reflected orchard conditions.
The climate of Alba was characterized by a yearly minimum air temperature (Tmin) of 8.3 ± 7.3 °C, a yearly maximum air temperature (Tmax) of 18.8 ± 8.8 °C, with the hottest months July and August (mean Tmax 30.2 ± 0.9 °C) and the mean annual air temperature (Tm) of 13.2 ± 8.2 °C. Total annual rainfall was 673 mm. The climate of Lequio Berra was characterized by a yearly minimum air temperature (Tmin) of 9.2 ± 6.7 °C, a yearly maximum air temperature (Tmax) of 17.0 ± 8.3 °C, with the hottest months July and August (mean Tmax 28.2 ± 0.7 °C) and the mean annual air temperature (Tm) of 12.8 ± 7.4 °C. Total annual rainfall was 816 mm (data from meteorological station of Alba and Rodello (for Lequio Berra), Arpa Piemonte, for the period 2004–2018).
2.2 Structural measurements
All structural measurements were carried out in May in the two considered orchards. Tree height (H, m) was measured by clinometers (HEC, Haglöf electronic clinometer, Sweden). Projected canopy area (Pc, m2) for each of the trees under study was measured by projecting the edge of the canopy to the ground and measuring the length along an axis from edge to edge through the canopy center (Gratani et al. 2011). Tree leaf area index (LAI) was measured monthly during the study period by the ‘LAI 2000 Plant canopy analyzer’ (LICOR Inc., Lincoln, USA). During the study period, the total leaf surface area of the canopy (SLT, m2 tree−1) was determined by multiplying the mean monthly LAI by Pc according to Gratani and Varone (2006).
2.3 Net photosynthesis and CO 2 sequestration capability
Net photosynthetic rate [NP, µmol(CO2) m−2 s−1] was measured monthly from May to October (i.e., when leaves are completely structured until the beginning of leaf senescence) by an open infrared CO2 gas analyzer (LC-Pro + , ADC, UK), equipped with a leaf chamber (PLC, Parkinson Leaf Chamber) in the two considered orchards. Measurements were carried out every two hours from 08:00 to 18:00 h on fully expanded sun leaves at the top of the canopy (n = 2 leaves per tree per plot in each area per sampling occasion).
Hourly CO2 sequestration capability per bush [CSh, kg (CO2) plants−1 h−1] during the study period was calculated by multiplying the obtained monthly SLT by the average value of NP (i.e., mean NP measured every two hours in the month) and the photosynthetic activity time (i.e., 2 hours) according to Gratani and Varone (2006). Daily CO2 sequestration capability per tree [CSd, kg (CO2) plants−1 day−1] was obtained by summing every value of CSh measured in each day. Then, the monthly CO2 sequestration capability per tree [CSm, kg (CO2) plants−1 month−1] was obtained by multiplying the CSd value of each month by the respective days and subsequently expressed in hectare based on the number of tree per hectare [Mg (CO2) ha−1 month−1]. Lastly, the total yearly CO2 sequestration capability per hectare [TCS, Mg (CO2) ha−1 year−1] was calculated by summing the CSm obtained in each month.
2.4 Data analysis
Differences in the considered traits were determined by the analysis of variance (ANOVA) and the Tukey test for multiple comparisons. Kolmogorov–Smirnov and Levene tests were used to verify the assumption of normality and homogeneity of variance, respectively. All data were shown as mean ± standard error (S.E). All statistical tests were performed using a statistical software package (PAST, Version 3.10).
3 Results
3.1 Structural measurements
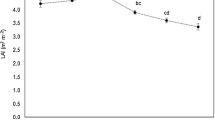
The average LAI value, during the sampling months, was 2.82 ± 0.34 and 3.77 ± 0.21 in orchardA and in orchardB, respectively. In both the orchards, the highest LAI values were measured in July (3.43 ± 0.05 and 4.05 ± 0.08, in orchardA and orchardB, respectively) (Fig. 1). The Pc value was halved in orchardA compared to orchardB due to the different training system, and then, the SFT in orchardB was more than 100% higher than orchardA.

Leaf area index (LAI, m2 m−2) values during the sampling months (May–October). Mean value and standard error are shown (n = 3)
3.2 Net photosynthesis and CO 2 sequestration capability
The average seasonal PN value during the study period was 13% higher (T test, p < 0.05) in orchardA than in orchardB, both having the significant highest PN rates in May (8.3 ± 0.1 and 7.6 ± 0.2 µmol m−2 s−1, mean daily value, in orchardA and orchardB, respectively) and the lowest ones in October (2.6 ± 0.3 and 1.9 ± 0.1 µmol m−2 s−1, mean daily value, in orchardA and orchardB, respectively) due to the approaching leaf senescence phase (ANOVA, p < 0.05; Fig. 2). The average CSm value per individual tree was 10.6 ± 1.7 and 25.7 ± 4.2 kg (CO2) plant month−1 in orchardA and orchardB, respectively (Fig. 3), and when expressed on hectare base, the mean CSm value in orchardA and orchardB was 4.25 ± 1.72 and 8.57 ± 3.41 Mg (CO2) ha−1 month−1, respectively (Fig. 4). Overall, during the study period, orchardA sequestered a total amount of CO2 equal to 26 Mg (CO2) ha−1 and orchardB equal to 51 Mg (CO2) ha−1. Table 1 shows the measured H, PC and monthly LAI and NP rates in each of the three plots considered in orchardA and orchardB.

Net photosynthetic rates [NP, µmol (CO2) m−2 s−1] of the average value measured during the day (8.00–18.00 h) in each sampling months (May–October). Mean value and standard error are shown (n = 3). Mean with different letters indicates significant differences, lowercase letters indicated significant differences between months in each orchard (ANOVA, p < 0.05), and capital letters indicate significant differences between the orchards in each month (T test, p < 0.05)

Monthly CO2 sequestration capability per plant in each orchard [CSm, kg (CO2) plant−1 month−1] during the sampling months (May–October)

Monthly CO2 sequestration capability per hectare of hazelnut orchard [CSm, Mg (CO2) ha−1 month−1] during the sampling months (May–October). Note mean value and standard error are shown (n = 2)
4 Discussion
On the whole, the two different training systems (i.e., single trunk vs bush) applied in hazelnut orchards in Piedmont had significant effects on carbon sequestration. This last depends, on the one hand, directly on the PN rates and, on the other hand, on the plant structural traits (Granata et al. 2019). However, although the nocturnal leaf respiration could adversely affect the plant’s sequestration capability, our estimation of its rates at leaf level reported an average value of around 1.02 µmol m−2 s−1 of emitted CO2 (mean value of the two orchards) resulting in lower magnitude compared to that removed through the net assimilation rates. Thus, its contribution in reducing the carbon sink capabilities can be overlooked. Unfortunately, we could not estimate the CO2 emissions from the orchard soil, although its contribution can be considered one of the major terrestrial sources of the greenhouse gases CO2 in the atmosphere (Schaufler et al. 2010). Comparing the PN rates, hazelnut in orchardA showed on average a significant higher value during the study period than hazelnut in orchardB, but the greater differences can be seen at the structural level and justified by the two applied training systems. In particular, plants in orchardB showed a more than 100% higher SFT which was reflected in a 33% higher LAI than plants in orchardA. This structural trait represents the projected area of foliage per unit ground area resulting in the most useful comparative measure of foliage quantity (Kram 1998) and of fundamental importance for quantifying the exchange rates of carbon between vegetation canopy and atmospheric interface (Spanner et al. 1994).
Thus, this higher LAI, as one of the major determinants of productivity, compensated with the lower PN rates so much that the value of carbon sequestration per plant was, on average, more than 100% higher (mean value during the study period) in orchardB than in orchardA. This result confirmed the role of LAI as indicator of carbon cycle in terrestrial ecosystem (Jonckheere et al. 2004), and its utility in the research focused on the plant carbon sequestration potential. Therefore, the training system like bush widespread at the higher elevation in the Piedmont hills (i.e., Alta Langa), with its larger size of the plants, seems to better contribute to the removal of the atmospheric CO2 by its higher carbon sequestration potential. The larger tree size in orchardB required a higher planting distance among the plants which resulted in a 17% lower plant density compared to orchardA, necessary to allow the mechanized actions inside the orchard, as well as for lighting and ventilation. When expressed on hectare basis, the average carbon sequestration capability during the study period resulted in 4.25 ± 0.70 and 8.57 ± 1.39 Mg (CO2) ha−1 month−1 in orchardA and orchardB, respectively, confirming the key role of the plant structural traits (i.e., LAI) in determining the amount of carbon removal from the atmosphere. Regarding the seasonal trends of the carbon sequestration potential, the association between the significant highest PN rates with higher LAI values caused the highest CSm value in the period May–July in both the orchards (5.64 ± 0.27 and 11.20 ± 0.42 Mg(CO2) ha−1 month−1, mean value, in orchardA and orchardB, respectively). This trend was explained, on the one hand, by the more suitable environmental conditions for PN both at the lower and higher altitude in Piedmont with a favorable mean air temperature. These last ranged from 18.6 °C to 27.4 °C in orchardA and from 17.3 °C to 25.9 °C in orchardB both following in the favorable range of air temperature for hazelnut according to the results of Schulze and Küppers (1979) and also associated with enough water availability being hazelnut a sensitive species to water deficit (Catoni et al. 2017, 2019). On the other hand, the seasonal LAI variations reported higher value in May and June and the highest one in July (3.43 ± 0.05 and 4.05 ± 0.05 in orchardA and orchardB, respectively). This highest LAI value in July was the consequence of achieving the complete maturation of the new leaves in the canopy reaching a total leaf surface of 35 ± 1 m2 and 84 ± 2 m2 in orchardA and orchardB, respectively. Following this period with the highest CSm, we observed a decline (by 37 and 32%, in orchardA and orchardB, respectively) in the carbon sequestration capability in the months August–September, where the important harvesting process takes place in piedmont because this cultivar is early ripening compared to others (Comunità Montana Alta Langa, 2009). The harvesting operations take place in both the orchards with a mechanized procedure, which guarantees a good hourly yield, but at the same time contribute to the emission of a significant quantity of greenhouse gases. In particular, based on the results of Nayeri et al. (2014) on 36 hazelnut farms in Iran, the amount of greenhouse emissions was estimated at 28.98 kg CO2eq ha−1 for diesel fuel and 0.355 kg CO2eq ha−1 for use of machineries.
However, these values of CO2 emissions were substantially lower compared to that removed in the same period by the hazelnut orchards, making them an active carbon sink. Finally, in October, due to the approaching of the senescence phase at physiological and structural traits there were the lowest CSm values. Regarding the contribution in the greenhouse emission given by the use of fertilizers, we have been able to calculate it through the use of specific CO2 emission coefficients according to Nayeri et al. (2014). Based on the data published by the Comunità Montana Alta Langa (2009) for the amount of the typical fertilizers, we could estimate a maximum amount of CO2 release equal to 104 kg CO2eq ha−1 by nitrogen fertilizer, 8 kg CO2eq ha−1 by phosphorus fertilizer and 16 kg CO2eq ha−1 by potassium fertilizer. Overall, the total amount of CO2 emission by hazelnut management over the entire production season can be estimated in 157.335 kg CO2eq ha−1 by summing the contribution of diesel fuel, machinery and fertilization practices. Considering that the total amount of CO2 sequestered by the two hazelnut orchards over the entire study period (May–October) was estimated at 26 Mg (CO2) ha−1 in orchardA and in 51 Mg (CO2) ha−1 in orchardB, they had an effective role as carbon sink at this local level (Fig. 5). These values corresponded to an amount of C sequestered of 7 Mg (C) ha−1 year−1 and 14 Mg (C) ha−1 year−1 in orchardA and in orchardB, respectively. The comparison between these values and that obtained for the hazelnut orchards in Lazio region (16 Mg of C ha−1 year−1, Granata et al. 2020) showed us a similar amount with orchardB, and indeed, the structural traits, meaning LAI values, were similar in these orchards in Lazio and Piedmont and related to the choice of the same training systems like bushes. On the other hand, the value in orchardA was beyond the lower limit of the range reported by Scandarelli et al. (2016) for others agro-ecosystems in Italy and represented by olive orchard (9.2 ± 1.1 Mg of C ha−1 year−1), while the value in orchardB was quite similar to that monitored in the apple orchards (14.6 ± 2.8 Mg of C ha−1 year−1) placed in the maximum limit of this range and according to the same authors similar to those reported for temperature forest in Luyssaert et al. (2007). Thus, on the one hand, we can highlight the higher amount of C sequestered by hazelnut orchards in Piedmont and, on the other hand, emphasize the significant role of the two training systems in affecting carbon sequestration potential.

The contribution of CO2 emissions from the farming activities carried out in the two hazelnut orchards compared with the total yearly amount of CO2 sequestered in orchardA and orchardB (small box)
5 Conclusion
Extending the average value of carbon sequestration potential, estimated in these two orchards, over the entire surface dedicated to this culture in Piedmont (24,555 ha, data from Istat 2019) we obtained a total CO2 sequestration of 944,476 Mg CO2) year−1 confirming that this species could create a sustainable ecosystem with favorable carbon sequestration potential. Thus, at the present state of our knowledge about the carbon sequestration potential of hazelnut orchard in Italy only the fourth region in terms of hazelnut production surface remains excluded (i.e., Sicily). However, with the current data in hand we could estimate a total amount of CO2 sequestered in Italy at 3,546,607 Mg (CO2) year−1 by summing the actual contribution of Lazio, Campania and Piedmont.
References
Aguilera E, Guzmán G, Alonso A (2015) Greenhouse gas emissions from conventional and organic cropping systems in Spain II Fruit tree orchards. Agron Sustain Dev 35:725–737
Catoni R, Gratani L, Bracco F, Granata MU (2017) How water supply during leaf development drives water stress response in Corylus avellana saplings. Sci Hort 214:122–132
Catoni R, Bracco F, Gratani L, Granata MU (2019) Physiological, morphological and anatomical leaf traits variation across leaf development in Corylus avellana. Mediterr Bot 40:185–192
Comunità Montana Alta Langa (2009) La coltivazione del nocciolo in Alta Langa. Linee guida per una corilicoltura sostenibile, Bra
Don A, Osborne B, Hastings A, Skiba U, Carter MS, Drewer J, Flessa H, Freibauer A, Hyvonen N, Jones MB, Lanigan GJ, Mander U, Monti A, Djomo SN, Valentine J, Walter K, Zegada-Lizarazu W, Zenone T (2012) Land-use change to bioenergy production in Europe: implications for the greenhouse gas balance and soil carbon. Glob Change Biol Bioenergy 4:372–391
Godone D, Garbarino M, Sibona E, Garnero G, Godone F (2014) Progressive fragmentation of a traditional Mediterranean landscape by hazelnut plantations: The impact of CAP over time in the Langhe region (NW Italy). Land Use Policy 36:259–266
Granata MU, Gratani L, Bracco F, Catoni R (2019) Carbon dioxide sequestration capability of an unmanaged old-growth broadleaf deciduous forest in a strict nature reserve. J Sustain For 38:85–89
Granata MU, Bracco F, Catoni R (2020) Carbon dioxide sequestration capability of hazelnut orchards: daily and seasonal trends. Energ Ecol Environ 5:153–160
Gratani L, Varone L (2006) Carbon sequestration by Quercus ilex L. and Quercus pubescens Willd. and their contribution to decreasing air temperature in Rome. Urban Ecosyst 9:27–37
Gratani L, Catoni R, Varone L (2011) Quercus ilex L. carbon sequestration capability related to shrub size. Environ Monit Assess 178:383–392
Istituto nazionale di statistica (Istat) (2019) Istat.it, Agricultura e Zootecnia, coltivazioni legnose: frutta fresca,TavC19
Jonckheere I, Fleck S, Nackaerts K, Muys B, Coppin P, Weiss M, Baret F (2004) Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agric For Meteorol 121:19–35
Kirschbaum MUF (2006) Temporary carbon sequestration cannot prevent climate change. Mitig Adapt Strat Glob Chang 11:1151–1164
Kram KJ (1998) Influence of species composition and forest age on leaf area index. Pol J Ecol 46:75–88
Luyssaert S, Inglima I, Jung M, Richardson AD, Reichstein M, Papale D, Piao SL, Schulze ED, Wingate L, Matteucci G, Aragao L, Aubinet M, Beer C, Bernhofer C, Black KG, Bonal D, Bonnefond JM, Chambers J, Ciais P, Cook B, Davis KJ, Dolman AJ, Gielen B, Goulden M, Grace J, Granier A, Grelle A, Griffis T, Grunwald T, Guidolotti G, Hanson PJ, Harding HDY, Hutyra LR, Kolari P, Kruijt B, Kutsch W, Lagergren F, Laurila T (2007) CO2 balance of boreal, temperate and tropical forests derived from a global database. Global Change Biol 13:2509–2537
Nardino M, Pernice F, Rossi F, Georgiadis T, Facini O, Motisi A, Drago A (2013) Annual and monthly carbon balance in an intensively managed Mediterranean olive orchard. Photosynthetica 51:63–74
Navarro MNV, Jourdan C, Sileye T, Braconnier S, Mialet-Serra I, Saint-Andre L, Dauzat J, Nouvellon Y, Epron D, Bonnefond JM et al (2008) Fruit development, not GPP, drives seasonal variation in NPP in a tropical palm plantation. Tree Physiol 28:1661–1674
Nayeri M, Firouzan AH, Azarpour E (2014) Greenhouse gas emissions for hazelnut production in forest north of Iran. Adv Environ Biol 8(24):289–292
Scandellari F, Caruso G, Liguori G, Meggio F, Palese Assunta M, Zanotelli D, Celano G, Gucci R, Inglese P, Pitacco A, Tagliavini M (2016) A survey of carbon sequestration potential of orchards and vineyards in Italy. Eur J Hortic Sci 81(2):106–114
Schaufler G, Kitzler B, Schindlbacher A, Skiba U, Sutton MA, Zechmeister-Boltenstern S (2010) Greenhouse gas emissions from European soils under different land use: effects of soil moisture and temperature. Eur J Soil Sci 61(5):683–696
Schimel DS, House JI, Hibbard KA, Bousquest P et al (2001) Recent patterns and mechanisms of carbon exchange by terrestrial ecosystems. Nature 414:169–172
Schulze ED, Küppers M (1979) Short-term and long-term effects of plant water deficits on stomatal response to humidity in Corylus avellana L. Planta 146:319–326
Smaje C (2015) The strong perennial vision: a critical review. Agroecol Sustain Food Syst 39:471–499
Smith P (2004) Monitoring and verification of soil carbon changes under Article 3.4 of the Kyoto Protocol Soil Use Manage 20: 264–270
Sofo A, Nuzzo V, Palese AM, Xiloyannis C, Celano G, Zukowskyj P, Dichio B (2005) Net CO2 storage in Mediterranean olive and peach orchards. Sci Hortic 107:17–24
Spanner M, Johnson L, Miller J, McCreight R, Freemantle J, Runyon J, Gong P (1994) Remote sensing of seasonal leaf area index across the oregon transect. Ecol Applic 4:258–271
UNFCCC (1997) Kyoto protocol to the United Nation framework convention on climate change. https://unfccc.int/
Valentini N, Me G (2002) Attualità e problematiche della coltura del nocciolo in Italia: la situazione piemontese. Atti del 2◦Convegno Nazionale sul Nocciolo, Giffoni Valle Piana (SA), pp. 133–140
Valentini N, Calizzano F, Boccacci P, Botta R (2014) Investigation on clonal variants within the hazelnut (Corylus avellana L.) cultivar ‘Tonda Gentile delle Langhe.’ Sci Hort 163:303–310
Zanotelli D, Montagnani L, Manca G, Scandellari F, Tagliavini M (2015) Net ecosystem carbon balance of an apple orchard. Eur J Agron 63:97–104
Zhang Y, Shen Y, Xu X, Sun H, Li F, Wang Q (2013) Characteristics of the water-energy-carbon fluxes of irrigated pear (Pyrus bretschneideri Rehd) orchards in the North China Plain. Agric Water Manag 128:140–148
Acknowledgements
This work was supported by the ‘Natural Reserve Bosco Siro Negri’ funded by the ministry of the environment and protection of land and Sea of Italy.
Funding
Open access funding provided by Università degli Studi di Pavia within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Informed consent
The authors give informed consent.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Granata, M.U., Catoni, R. & Bracco, F. The role of two different training systems in affecting carbon sequestration capability in hazelnut orchards. Energ. Ecol. Environ. 6, 285–291 (2021). https://doi.org/10.1007/s40974-020-00202-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40974-020-00202-1