
Abstract
Logging of natural forests can directly alter landscape structure, defined here as the spatial pattern of patches of overstory forest cover of different ages. Logging can also alter landscape structure through interactions with other disturbances such as wildfire. Here, I briefly outline interactions between logging of wet forests, altered fire dynamics and landscape structure, with particular emphasis on the wet ash eucalypt forests of south-eastern Australia. There is compelling evidence for increased fire proneness of logged and regenerated wet forest stands around the world. There is also evidence of accumulated effects on fire dynamics, so-called “landscape traps” that are associated with multiple, spatially dispersed cutblocks in wood production landscapes. That is, changes in stand-level flammability may accumulate over larger areas, thereby influencing patterns of spatial contagion in fire behavior. The impacts of altered patterns of landscape structure on biodiversity and ecosystem processes are often poorly understood. New science is required to better understand, quantify, and predict biotic responses to the new landscape dynamics and the spatial and temporal patterns of forest cover now occurring in many landscapes. Altered patterns of landscape structure have the potential to trigger ecosystem collapse in some forested environments. However, predicting ecosystem collapse is difficult and may even be impossible from a practical perspective. Managers and researchers need to become better informed about the risks of negative effects of combinations of ecosystem stressors that lead to novel spatial patterns in landscape structure and may make forest ecosystems more prone to landscape traps, regime shifts, and ecosystem collapse.
Similar content being viewed by others


Introduction
A large proportion of the world’s terrestrial environment is subject to human land use. This includes forests, large areas of which are subject to land conversion for agriculture and urbanization, industrial plantation establishment, and various kinds of logging [1]. Logging can result in significant short- and long-term changes in forest environments at several spatial scales, ranging from (but not restricted to) the size of individual trees [2] and the structure and composition of stands [3], to the composition and heterogeneity of landscapes [4]. Documenting these many and varied changes is a highly complex, multi-faceted task that is beyond the scope of this review. Hence, the focus of this paper is restricted to interactions between logging regimes and the spatial structure of forest landscapes (which is crudely defined as the spatial pattern of patches of cover of overstory forest of different ages).
Logging has the potential to directly alter landscape structure as a result of cutting patterns that transform the spatial pattern of forest patches [5]. Indeed, there is a long history of research in landscape ecology on interactions between human disturbances such as logging and altered landscape structure. For example, one of the seminal early papers in the journal Landscape Ecology examined the potential effects on the spatial pattern of remaining forest and on key ecological processes resulting from a spatially dispersed (or checkerboard) pattern of logging versus a harvest schedule based on aggregating cutblocks [5]. Logging also has the potential to directly alter landscape structure in more subtle ways that can be harder to quantify. For example, many forest landscapes are characterized by marked variations in site and stand-level productivity with logging often focused on high productivity areas. Altering the structure of forest landscapes in this way can have profound impacts on biodiversity, particularly given strong associations of some species and groups with high productivity areas [6].
Logging may alter landscape structure not only as a result of cutting patterns but also indirectly through interactions with other kinds of disturbances such as fire regimes [7–9]. In this report, I provide some perspectives on relationships between forest landscape structure and natural resource management, with a particular focus on the effects of logging in wet forests where fire regimes (sensu [10, 11]) are typically rare, high-severity stand-replacing conflagrations. Interactions between logging and fire in these kinds of forest ecosystems can have profound effects on: (1) spatial and temporal patterns of forest landscape cover, (2) key ecosystem processes (including disturbance regimes), and (3) various elements of the biota. On this basis, I first summarize some of the empirical evidence for relationships between logging and fire proneness within harvested and subsequently regenerated stands, with a particular emphasis on the wet ash eucalypt forests of south-eastern Australia. I then explore the potential for elevated stand-level fire risk to accumulate across multiple stands and landscapes, thereby potentially altering the spatial contagion in fire dynamics at scales spanning multiple stands and entire landscapes. Finally, I briefly outline some of the considerable challenges which remain in rigorously quantifying the cumulative effects of logging and fire (and their interaction) on biodiversity and key ecosystem processes as mediated through spatio-temporal changes in landscape cover.
Much has been learned about relationships between logging and spatial patterns of forest landscape cover since the early studies by Franklin and Forman [5], but much also remains to be learned. Such new knowledge is important as forest landscapes worldwide are increasingly subject to major, often rapid, and novel combinations of natural and human disturbances [12•]. These novel combinations of disturbances are leading to novel spatial and temporal patterns of vegetation cover. Yet, how such combinations of disturbances and novel spatial and temporal patterns of landscape cover affect biodiversity and key ecological processes remains poorly understood. Moreover, work on interactions between logging, landscape structure, and fire remains highly contentious and controversial [13–16]. Given this, considerable empirical work is required to develop a strong body of scientific evidence that can help resolve debates about relationships between the type and spatial pattern of logging, landscape structure, and responses of biodiversity and key ecosystem processes.
Logging and Fire Interactions
Logging and Stand-Level Fire Proneness
A key question in the management of some kinds of forest is: Does logging make forests more fire prone? A review on this topic concluded that strategic entries into dry forest stands can reduce the risks of high severity fire [17]. Conversely, logging can make moist or wet forests more fire prone [17]. Indeed, clear evidence of such inter-relationships can be found in tropical rainforests [7, 8], wet temperate forests in western North America [9], and wet temperate forests in Australia [18••, 19, 20], although some moist forests may be an exception to this general response [21].
Several factors may underpin the increased flammability of post-logged and regenerated moist forest stands. First, the high densities of trees in young forest stands may make them more flammable than mature forests [19, 20]. Second, changes in the stand structure of post-logged and regenerated stands such as the losses of mesic understory plants and rainforest cover may alter microclimatic conditions and make them less wet and more prone to high-severity fire. Third, the debris or slash left after logging (e.g., discarded lateral branches, crowns, bark, and unmerchantable stems) may contribute to fuel loads in young regenerating forests [18••].
The relationships between logging and fire proneness in the wet forests of south-eastern Australia have been challenged by Attiwill et al. [13], who argued there is no relationship between logging and fire proneness. However, the work of these authors was flawed for several reasons [14], but, most importantly, it failed to account for non-linearities in responses between forest age and fire proneness [18••] (Fig. 1) which resulted in logging-fire proneness relationships being overlooked. This was demonstrated by Taylor et al. [18••], who found that in Australian wet forests the probability of crown-scorching fire was almost zero for the first 7 years after logging, but it then increased dramatically and highly significantly for the following 40+ years and did not approximate the relatively low levels characteristic of older forests until a century or more after logging (Fig. 1). Such relationships between stand age and the probability of crown-scorching fire shown in Fig. 1 are important because the spatial structure of many wood production landscapes is dominated by young, spatially clustered fire-prone stands, typically aged 40–75 years old or younger. Figure 2 shows one of many example landscapes characterized by such spatial patterns of landscape structure.

Non-linear statistically-derived relationships between stand age and the probability of crown-scorching wildfire in the wet forests of the Central Highlands of Victoria, south-eastern Australia (redrawn from Taylor et al. [18••]).

An extensively logged wood production landscape in the wet forests of Victoria, south-eastern Australia showing the year and location of a subset of cut blocks where the young regenerating forests are significantly more fire prone than older forests (see text). The underlying hypothesis of the landscape trap model is that an array of more fire-prone regenerating stands collectively add to the spatial contagion of fire at larger (landscape) scales. (Photo by Dave Blair)
It is important to note there are some moist forest ecosystems where strategic interventions in wood production forests may reduce fire risk. For example, in the boreal forests of Canada, strategic interventions in logged areas to promote greater areas of less flammable broadleaf trees in place of conifers may mitigate fire risks [21]. Roads and cutblocks in these forests may impede the spread of fire, although roads also may lead to an increase in ignition points [22]. Moreover, in Canadian mixedwood boreal forests, fire initiation following lightning strikes is more likely to occur in harvested areas because of increased fine fuels resulting from logging slash and this effect can remain for 10–30 years following logging [23].
Logging and the Landscape Trap Concept
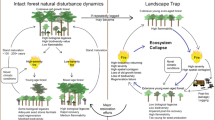
In intensively logged forest landscapes, changes in stand-level flammability have the potential to accumulate over larger areas, thereby influencing patterns of spatial contagion in fire behavior at landscape scales (Fig. 2). That is, the number and pattern of logged areas in a landscape (and the associated elevated fire severity within each logged stand) may lead to altered landscape-wide fire behavior and dynamics. Lindenmayer et al. [24] developed a theoretical conceptualization of this hypothesized inter-relationships between logging and fire and termed it a “landscape trap”. They defined a landscape trap as occurring when entire landscapes are shifted into and then maintained in a highly compromised state, typically young regrowth forest regenerating after logging. Young, densely stocked stands of post-logging regrowth produce large amounts of fine and medium fuels and are at risk of reburning at high severity before they reach a more mature state where trees are sexually mature and are able to self-regenerate (see Fig. 3). In addition, cutblocks in wood production landscapes often tend to be spatially and temporally clustered, further contributing to landscape-scale change.
Lindenmayer et al. [24] explored the landscape trap concept for the wet montane ash eucalypt forests of Victoria in which significant areas of forest have been subject to widespread industrial logging over the past 40 years and where there have been extensive stand-replacing wildfires in recent decades [25] (Fig. 3). Such repeated fires within extensively altered landscapes can eliminate the development of key stand structural attributes like large old cavity-bearing trees which are a critical habitat for a wide range of forest-dependent species [26]. Moreover, repeated fire in “trapped” landscapes may eventually result in a regime shift (sensu [27]), with wet forest being replaced by understory scrub [24].

Interactions between logging and fire that underpin the development of a landscape trap and, through feedbacks, affect landscape structure and landscape-wide patterns of key components of vegetation structure such as the abundance of large old cavity-bearing trees
Restoring Spatial Landscape Structure to Tackle the Landscape Trap Problem
The Australian wet forest environments where the landscape trap concept was first developed have been heavily altered by past logging operations (Fig. 2). This has directly and indirectly led to a dramatically altered landscape structure in these forests. Historically, relatively small areas of regrowth montane ash forest were embedded within large areas of old growth or mixed age forest [28]. Now the reverse is true; small patches of old growth (<1.2 % of the montane ash estate) are embedded within extensive areas of structurally simplified young, highly fire-prone regrowth forest [26]. Indeed, the altered spatial patterns and limited amount of old growth forest cover are likely well outside the bounds of historic range of natural variability (sensu [29, 30]) in montane ash ecosystems. Such changes in spatial patterns of forest cover are known to have significant problems in other ecosystems such as the boreal forests of Canada [31].
Although no old growth forest has been logged in Victorian montane ash ecosystems for almost 20 years, the limited current size of the old growth forest estate is a legacy of past logging, past recurrent wildfires, and a combination of both (i.e., post-fire salvage logging) [25]. Old growth forest is important because, in comparison with young logged and regenerated forest, it is less fire-prone [18••], stores significantly more biomass carbon [32], yields more water for human consumption [33], generates more (higher value) sawlogs relative to pulp logs [34], and provides significantly more key nesting resources for an array of cavity-dependent vertebrate species [28]. Thus, the negative ecological and economic impacts of a landscape trap in which forests are trapped at a young age are very large [25]. Preventing the development of a landscape trap is therefore critically important. The challenge is to regrow the old growth forest estate in ways that transform current novel patterns of landscape structure to more ecologically appropriate patterns which more closely resemble the amounts and spatial patterns of the old growth stands that used to characterize these forests. That is, use the concepts of the historic range of natural variability (see Keane et al. [29]) in spatial patterns of old growth forest cover to recreate appropriate future spatial of old growth coverage (see also Kuuluvainen [35]). Historically between 30 % and 60 % of the montane ash forest estate was old growth [25], with this age class typically occurring on flat plateaus or in deep, sheltered gullies [36]. However, key forest ecosystem restoration questions remain. For example: Given a particular target amount of old growth cover (e.g., 30 % of a particular region), how much forest needs to be set aside to reach this target? For example, if the long-term goal is 30 %, should 50 % be set aside because 20 % of retained forest will be damaged by future fires? How much old growth forest is required to prevent landscape traps from occurring? What spatial patterns are required – can old forest be spatially dispersed or should it be aggregated? How large should individual old growth patches be? In many respects, these questions are the flipside of the simulation study on forest cutting patterns by Franklin and Forman [5] for determining the amounts and spatial patterns of old growth forest cover that need to be restored and underpin ecologically sustainable ecosystem processes (particularly fire regimes).
Challenges in Quantifying the Effects on Biodiversity of Multiple Drivers of Change in Forest Landscape Patterns
Many studies have shown that spatial patterns of landscape cover can have profound effects on biota (e.g., [16, 37, 38]). However, as outlined above, forest landscapes worldwide are increasingly subject to rapid transformation [31, 39••], with novel combinations of drivers producing novel patterns of landscape structure [12•]. The response of biodiversity and key ecological processes to novel spatial and temporal patterns of landscape structure is poorly known. This is, in part, because many past studies have been snapshot investigations of spatial patterns of landscape structure at one point in time and largely of one kind of disturbance (e.g., fire), rather than time-series studies of multiple temporal and spatial drivers that characterize changes in landscape structure in many ecosystems [12•, 24]. New approaches are needed that link and extend traditional approaches in landscape ecology (which often focus on spatial landscape processes) and traditional approaches in disturbance ecology (which often focus on temporal landscape processes and history at a single location). This is important because changes in disturbance regimes have spatial implications for the amount and configuration of suitable habitat in a landscape [5, 40•], the spatial patterns of abundance of species [41], and population viability [42]. For example, a critical knowledge gap is how the cumulative spatial patterns of logging (see Figs. 2 and 4) affect biodiversity and key ecosystem processes [43]. Lessons from the effects of fire on the landscape structure of forests [37, 44] may not be readily tranferable to studies of the cumulative effects on biodiversity resulting from continuing to add logged cutblocks to harvested landscapes over successive years in a cutting rotation and which can cumulatively cover a large proportion of a landscape [5]. Moreover, spatial patterns and habitat suitability created by logging are very different to those resulting from fire [4, 45]. Furthermore, natural disturbances like fire occur in addition to human disturbances such as logging and the cumulative effects of both need to be considered in terms of their combined impacts on landscape structure (see Fig. 4), key ecosystem processes, and on biodiversity. An additional challenge with such kinds of spatio-temporal work will be to determine if there are threshold effects in relationships between various measures of biodiversity and the amount of cover of different forest age classes in forest landscapes. Most studies of threshold effects have been in agricultural areas characterized by remnant patches of native vegetation surrounded by largely cleared pastures and cropland (e.g., [46]). Indeed, threshold responses have been found in some agricultural landscapes [47], but not in others [48, 49•]. Similar work on critical breakpoints in measures like species richness and/or the occurrence of individual species is comparatively rare in native forests where disturbed areas are not removed but instead regenerated following harvesting or fire (but see [50–52]).

Spatio-temporal patterns of disturbance in two sample landscapes over 30 years in the montane ash forest of the Central Highlands of Victoria. Marked changes have occurred over time in the forest landscape in the top series as a result of fire (light gray) and logging (dark gray). Few changes have occurred in the bottom series because there has been limited or no fire or logging. Circles show nominal scales at which landscape analyses of the effects of changing spatial structure of the landscape can be quantified (radii 500 m, 1,000 m, and 2,000 m) centered around long-term monitoring sites where data on fauna have been gathered. Major fires occurred in 1939 and 2009
Approaches borrowed from medical sciences show considerable promise for application in landscape ecology to better quantify the effects of both spatially dynamic and temporally dynamic patterns of landscape structure on temporal changes in various measures of biodiversity as well as on key ecological processes. As an example, methods such as Functional Data Analysis [53, 54] that have previously been used in spatio-temporal studies of brain scan imaging can now be applied in studies of forest landscape ecology. Functional Data Analysis facilitates inclusion of 3D spatial and temporal maps (see Fig. 4) and images as predictors in statistical models by creating meaningful single and composite sets of covariates to describe key relevant features of the maps and images. This will greatly improve understanding and capability to predict biotic responses to spatio-temporal change in forest landscape cover. Preliminary testing of spatio-temporal changes in landscape structure effects on biodiversity resulting from logging, fire, and a combination of both have begun in 170 landscapes in the wet forests of south-eastern Australia. Figure 4 shows an example of two of the 170 landscapes characterized by high contrast in the spatio-temporal change in landscape structure over a period of 20 years. Studies employing Functional Data Analysis have coupled 32 years of repeated measures of biodiversity with changes in landscape spatial structure over the corresponding period and are being used to: (1) determine whether there are simple additive or more complex multiplicative (synergistic) cumulative effects (sensu [55, 56•]) on biodiversity resulting from adding cutblocks to harvested landscapes over successive years in a cutting rotation, (2) quantify the combined effects of logging and fire on biodiversity (both in space and over time), and (3) identify where to locate cutblocks in the landscape (and at what time in a rotation) to best integrate wood production and conservation values.
An important component of future work on the intersection between logging, spatio-temporal changes in landscape structure, and the responses of biodiversity and key ecosystems will be the development of computer visualization tools to assist forest, wildlife, and fire managers to better manage wood production forests for multiple values in space and over time. These tools would enable managers to simulate the spatial patterns of landscape cover through time and in response to different kinds of (logging and fire-mediated) disturbances. This would allow managers to compare spatio-temporal patterns of future forest cover with those under historic range of variability in cover. Spatio-temporal simulation tools also would assist managers to visualize the spatial and temporal implications for biodiversity resulting from management decisions and, in turn, help tackle potential problems with increased fire spread and elevated fire severity that might otherwise be associated with timber harvesting.
Altered Landscape Structure and Challenges in Predicting Forest Ecosystem Collapse
The intertwined concepts of altered landscape structure, landscape traps, and regime shifts imply risks of ecosystem collapse in some forest environments. Ecosystem collapse can be broadly defined as an abrupt change in ecosystem state [57] such as a rapid change from a rainforest to an exotic grassland (e.g., see [7]). Ecosystem collapse can be associated with large losses of biodiversity and key ecosystem services [57]. Ecosystem collapse may be irreversible [58–61], or alternatively it can be costly and time consuming to reverse (e.g., [62]). The concept of ecosystem collapse is now part of the IUCN Red List ecosystem process developed by Keith et al. [63•] and pivots around the ideas of significant alterations in ecosystem state and landscape cover, modification or loss of key ecosystem processes, and substantial losses of key ecosystem structures or critical species (e.g., see [64]). Despite this formalized approach to codify ecosystem collapse and the extensive literature on the topic of ecological collapse, it is clear that it is extremely difficult to predict if and when collapse might occur, including in forest ecosystems.
Some largely theoretical work (e.g., [65]) suggests that increasing variability in, and impaired recovery of, some ecological attributes like plant regeneration and populations of keystone biota are a useful early indicator of subsequent ecosystem collapse [66, 67]. However, this kind of work is hard to apply in practical management. This is because: (1) it is often not clear which entities are appropriate surrogates for ecosystem collapse, and (2) the need to gather extremely long-term datasets that target particular surrogates as early warning signals but that can be difficult and costly to monitor. Moreover, some ecosystems that have collapsed have not exhibited these early warning signals [68, 69]. Indeed, as noted by Boettiger and Hastings: “… no one has yet managed to use the theory on early warning signals to predict a natural catastrophe” (p. 157 [70•]). These authors further suggest that: “generic early warning signals of tipping points are unlikely to exist.” This is a major problem as significant changes in the ecosystem state will be a “wise-after-the-fact” situation – retrospective rather predictive – and therefore of limited value for guiding the on-the-ground management of natural resources.
Despite the difficulty in predicting ecosystem collapse, there are nevertheless significant potential risks of it occurring in some forest ecosystems, in part because of altered landscape structure. This is acknowledged under the formalized approach to ecosystem assessment development by the IUCN [63•]. Progress toward managing landscapes to limit the risks of ecosystem collapse may be made if managers and researchers more carefully define what they mean by ecosystem collapse and plan for this as a possibility. What constitutes possible ecosystem collapse needs to be carefully defined for a given ecosystem. This is because how ecosystem collapse manifests will be ecosystem specific [70•], and therefore managing to prevent it will likewise be ecosystem specific [63•]. Ecosystem collapse also may need to be defined relative to some benchmark or reference conditions or in relation to some well-defined goods and services that are expected from that ecosystem.
As part of the attempts to avoid ecosystem collapse arising from altered landscape structure, managers and researchers need to become more attuned to the risks of negative effects of multiple and novel combinations of ecosystem stressors that lead to novel spatial patterns in landscape structure (such as major changes in the amount and distribution of old growth forest). This is important because co-occurring disturbances can interact to produce different outcomes than those predicted by summing individual effects [55, 56•, 71]. In addition, forest managers and researchers need to develop trigger points in management programs that precipitate rapid change to avoid deterioration in ecosystems which might then be difficult and/or very expensive to reverse. This requires “management thresholds” [72], which when approached, trigger a change in management and associated policy setting. As an example, amounts and rates of cutting might be reduced and harvest rotation times increased directly after a wildfire if the spatial extent of burned forest in a landscape exceeds a given level (e.g., 30 % of a landscape or region) to avoid overcutting of the remaining unburned forest.
Finally, whilst it is clear that research on the theory of ecosystem collapse and predictive capacity lags well behind the needs of forest landscape managers, this does not mean research efforts attempting to predict ecosystem collapse should be abandoned. Rather, redoubled effort is required, particularly given the parlous state of many of the planet’s environments [73].
Conclusions
This paper has focused on interactions between logging of wet forests and landscape structure with a particular emphasis on altered fire dynamics at the stand level and altered spatio-temporal patterns of fire dynamics at the landscape level. There is compelling evidence of increased fire proneness of many kinds of logged and regenerated wet forests around the world. There is also some evidence of accumulated effects on fire dynamics – landscape traps – associated with multiple, often spatially clustered cutblocks in wood production landscapes. Novel combinations of disturbances leading to novel patterns of landscape structure both in space and over time are characterizing many forest landscapes globally. However, the impacts of such altered patterns of landscape structure on biodiversity and key ecosystem processes is often poorly understood. New science is required to better understand, quantify, and predict biotic responses to the new landscape dynamics and the new spatial and temporal patterns of forest cover now occurring in many landscapes. Altered patterns of landscape structure have the potential to trigger ecosystem collapse in some forested environments. However, predicting ecosystem collapse is notoriously difficult to do – and may be impossible from a practical perspective. Managers and researchers need to become more attuned to the risks of negative effects of multiple and novel combinations of ecosystem stressors that lead to novel spatial patterns in landscape structure. They also need to develop trigger points in management programs that precipitate rapid change to avoid deterioration in ecosystems.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
Mackey B, DellaSala DA, Kormos C, Lindenmayer DB, Kumpel N, Zimmerman B, et al. Policy options for the world’s primary forests in multilateral environmental agreements. Conserv Lett. 2015;8:139–47.
Linder P, Östlund L. Structural changes in three mid-boreal Swedish forest landscapes, 1885–1996. Biol Conserv. 1998;85:9–19.
Lindenmayer DB, Franklin JF. Managing stand structure as part of ecologically sustainable forest management in Australian Mountain Ash forests. Conserv Biol. 1997;11(5):1053–68.
Lindenmayer DB, Franklin JF. Conserving forest biodiversity: a comprehensive multiscaled approach. Washington DC: Island Press; 2002.
Franklin JF, Forman RT. Creating landscape patterns by forest cutting: ecological consequences and principles. Landsc Ecol. 1987;1:5–18.
Braithwaite LW, Turner J, Key J. Studies on the arboreal marsupial fauna of eucalypt forests being harvested for woodpulp and Eden, New South Wales. III. Relationships between the faunal densities, eucalypt occurrence and foliage nutrients and soil parent materials. Aust Wildl Res. 1984;11:41–8.
van Nieuwstadt MG, Shiel D, Kartawinata D. The ecological consequences of logging in the burned forests of east Kalimantan, Indonesia. Conserv Biol. 2001;15:1183–6.
Cochrane MA, Laurance WF. Synergisms among fire, land use, and climate change in the Amazon. Ambio. 2008;37:522–7.
Thompson JR, Spies TA, Ganio LM. Reburn severity in managed and unmanaged vegetation in a large wildfire. Proc Natl Acad Sci U S A. 2007;104:10743–8.
Gill AM. Fire and the Australian flora: a review. Aust For. 1975;38:4–25.
Keeley JE. Fire intensity, fire severity and burn severity: a brief review and suggested usage. Int J Wildland Fire. 2009;18:116–26.
Hobbs RJ, Higgs ES, Hall CM. Novel ecosystems: intervening in the new world order. Oxford: Wiley; 2013. This excellent edited book discusses the controversial topic of novel ecosystems. It explores different kinds of novel ecosystems and outlines how they might be defined, recognized and managed.
Attiwill PM, Ryan MF, Burrows N, Cheney N, McCaw L, Neyland M, et al. Timber harvesting does not increase fire risk and severity in wet eucalypt forests of southern Australia. Conserv Lett. 2014;7:341–54.
Bradstock R, Price OF. Logging and fire in Australian forests: errors by Attiwill et al. (2014). Conserv Lett. 2014;7(4):419–20.
Donato DC, Fontaine JB, Campbell JL, Robinson WD, Kauffman JB, Law BE. Post-wildfire logging hinders regeneration and increases fire risk. Science. 2006;311:352.
Taylor RS, Watson SJ, Nimmo DG, Kelly LT, Bennett AF, Clarke MF. Landscape-scale effects of fire on bird assemblages: does pyrodiversity beget biodiversity? Divers Distrib. 2012;18:519–29.
Lindenmayer DB, Hunter ML, Burton PJ, Gibbons P. Effects of logging on fire regimes in moist forests. Conserv Lett. 2009;2:271–7.
Taylor C, McCarthy MA, Lindenmayer DB. Non-linear effects of stand age on fire severity. Conserv Lett. 2014;7:355–70. This paper provides compelling evidence of the significant non-linear statistical relationships between logging and the elevated risk of crown-scorching fire, with a focus on the wet montane ash forests of south-eastern Australia. The work has major implications for forest management and biodiversity conservation and their interaction with fire regimes that have been altered by human activities like logging.
Florence RG. Ecology and silviculture of eucalypt forests. Melbourne: CSIRO Publishing; 1996.
Jackson WD. Fire, air, water and earth - an elemental ecology of Tasmania. Proc Ecol Soc Aust. 1968;3:9–16.
Girardin M, Terrier A. Mitigating risks of future wildfires by management of the forest composition: an analysis of the offsetting potential through boreal Canada. Clim Chang. 2015;130(4):587–601.
Bradshaw CJ, Warkentin IG, Sodhi NS. Urgent preservation of boreal carbon stocks and biodiversity. Trends Ecol Evol. 2009;24:541–8.
Krawchuk MA, Cumming SG. Disturbance history affects lightning fire initiation in the mixedwood boreal forest: observations and simulations. For Ecol Manag. 2009;257:1613–22.
Lindenmayer DB, Hobbs RJ, Likens GE, Krebs C, Banks SC. Newly discovered landscape traps produce regime shifts in wet forests. Proc Natl Acad Sci U S A. 2011;108:15887–91.
Lindenmayer DB, Blair D, McBurney L, Banks S. Mountain Ash: fire, logging and the future of Victoria’s giant forests. Melbourne: CSIRO Publishing; 2015.
Lindenmayer DB, Blanchard W, McBurney L, Blair D, Banks S, Likens GE, et al. Interacting factors driving a major loss of large trees with cavities in an iconic forest ecosystem. PLoS One. 2012;7:e41864.
Folke C, Carpenter S, Walker B, Scheffer M, Elmqvist T, Gunderson L, et al. Regime shifts, resilience, and biodiversity in ecosystem management. Annu Rev Ecol Syst. 2004;35:557–81.
Lindenmayer DB. Forest pattern and ecological process: a synthesis of 25 years of research. Melbourne: CSIRO Publishing; 2009.
Keane RE, Hessburg PF, Landres PB, Swanson FJ. The use of historical range and variability (HRV) in landscape management. For Ecol Manag. 2009;258:1025–37.
Kuuluvainen T, Grenfell R. Natural disturbance emulation in boreal forest ecosystem management - theories, strategies, and a comparison with conventional even-aged management. Can J For Res. 2012;42:1185–203.
Cyr D, Gauthier S, Bergeron Y, Carcaillet C. Forest management is driving the eastern North American boreal forest outside its natural range of variability. Front Ecol Environ. 2009;7:519–24.
Keith H, Lindenmayer DB, Macintosh A, Mackey B. Under what circumstances do wood products from native forests benefit climate change mitigation? PLoS One. 2015;10:e0139640.
O’Shaughnessy P, Jayasuriya J. Managing the ash-type forest for water production in Victoria. In: McKinnell FH, Hopkins ER, Fox JED, editors. Forest management in Australia. Chipping Norton: Surrey Beatty and Sons; 1991. p. 341–63.
Flint A, Fagg P. Mountain ash in Victoria’s state forests. Melbourne: Department of Sustainability and Environment; 2007.
Kuuluvainen T. Natural variability of forests as a reference for restoring and manageing biological diversity in boreal Fennoscandia. Silva Fennica. 2002;36:97–125.
Mackey B, Lindenmayer DB, Gill AM, McCarthy MA, Lindesay JA. Wildlife, fire and future climate: a forest ecosystem analysis. Melbourne: CSIRO Publishing; 2002.
McKenzie D, Miller C, Falk DA, editors. The landscape ecology of fire. Dordrecht: Springer; 2011.
Nimmo DG, Kelly LT, Spence-Bailey LM, Watson SJ, Haslem A, White JG, et al. Predicting the century-long post-fire responses of reptiles. Glob Ecol Biogeogr. 2012;21:1062–73.
Watson SJ, Luck GW, Spooner PG, Watson DM. Land-use change: incorpoarting the frequency, sequence, time span, and magnitude of changes into ecological research. Front Ecol Environ. 2014;12:241–9. This excellent and thought-provoking research paper discuss multiple temporal changes in spatial patterns and types of landscape cover and its profound implications for biodiversity conservation and landscape management.
Banks SC, Cary GJ, Smith AL, Davies ID, Driscoll DA, Gill AM, et al. How does ecological disturbance influence genetic diversity? Trends Ecol Evol. 2013;28:670–9. This paper highlights how spatial changes in patterns of forest cover can alter not just biotic assemblages and populations of individual species but also genetic variability.
Nimmo DG, Kelly LT, Spence-Bailey LM, Watson SJ, Taylor RS, Clarke MF, et al. Fire mosaics and reptile conservation in a fire-prone region. Conserv Biol. 2013;27:345–53.
Amarasekare P, Possingham H. Patch dynamics and metapopulation theory: the case of successional species. J Theor Biol. 2001;209:333–44.
Spies TA, Johnson KN, Burnett KM, Ohmann JL, McComb BC, Reeves GH, et al. Cumulative ecological and socioeconomic effects of forest policies in Coastal Oregon. Ecol Appl. 2007;17(1):5–17.
Lindenmayer DB, Blanchard W, McBurney L, Blair D, Banks SC, Driscoll DA, et al. Complex responses of birds to landscape-level fire extent, fire severity and environmental drivers. Divers Distrib. 2014;20:467–77.
McCarthy MA, Burgman MA. Coping with uncertainty in forest wildlife planning. For Ecol Manag. 1995;74(1–3):23–36.
Ochoa-Quintero JM, Gardner TA, Rosa I, de Barros Ferraz SF, Sutherland WJ. Thresholds of species loss in Amazonian deforestation frontier landscapes. Conserv Biol. 2015;29(2):440–51.
Radford JQ, Bennett AF, Cheers GJ. Landscape-level thresholds of habitat cover for woodland-dependent birds. Biol Conserv. 2005;124:317–37.
Cunningham RB, Lindenmayer DB, Barton PS, Ikin K, Crane M, Michael D, et al. Cross-sectional and temporal relationships between bird occupancy and vegetation cover at multiple spatial scales. Ecol Appl. 2014;24:1275–88.
Cunningham RB, Lindenmayer DB, Crane M, Michael DR, Barton PS, Gibbons P, et al. The law of diminishing returns: woodland birds respond to native vegetation cover at multiple spatial scales and over time. Divers Distrib. 2014;20:59–71. This research highlights the existence of non-threshold relationships between vegetation cover and species occurrence in landscapes and has implications for landscape management programs that seek to increase cover levels to promote biodiversity conservation.
Guenette J-S, Villard MA. Thresholds in forest bird response to habitat alteration as quantitative targets for conservation. Conserv Biol. 2005;9:1168–80.
Villard MA. Land use intensification and the status of focal species in managed forest landscapes of New Brunswick, Canada. In: Lindenmayer D, Cunningham S, Young A, editors. Land use intensification: effects on agriculture, biodiversity and ecological processes. Melbourne and London: CSIRO Publishing and CRC Press; 2012. p. 85–92.
Lindenmayer DB, Fischer J, Cunningham RB. Native vegetation cover thresholds associated with species responses. Biol Conserv. 2005;124:311–6.
Reiss PT, Ogden RT. Functional principal component regression and functional partial least squares. J Am Stat Assoc. 2007;102:984–96.
Reiss PT, Ogden RT. Functional generalized linear models with images as predictors. Biometrics. 2010;66:61–9.
Didham R, Tylianakis JM, Gemmell NJ, Rand TA, Ewers RM. Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol Evol. 2007;22:489–96.
Foster CN, Sato CF, Lindenmayer DB, Barton PS. Integrating theory into disturbance interaction experiments to better inform ecosystem management. Glob Chang Biol. 2015;22(4):1325–35. This paper highlights the importance of various kinds of experimental designs in developing robust studies that aim to identify the interacting effects of multiple stressors on ecosystems and assemblages of species within ecosystems.
MacDougall AS, McCann KS, Gellner G, Turkington R. Diversity loss with persistent human disturbance increases vulnerability to ecosystem collapse. Nature. 2013;494:86–9.
Freilich JE, Emlen JM, Duda JJ, Freeman DC, Cafaro PJ. Ecological effects of ranching: a six-point critique. Bioscience. 2003;53:759–63.
Young JA, Clements CD. Cheatgrass: fire and forage on the range. Reno: University of Nevada Press; 2009.
D’Antonio CM, Vitousek PM. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu Rev Ecol Syst. 1992;23:63–87.
Yelenik SG, D’Antonio CM. Self-reinforcing impacts of plant invasions change over time. Nature. 2013;503:517–20.
Frank KT, Petrie B, Fisher JA, Leggett WC. Transient dynamics of an altered large marine ecosystem. Nature. 2011;477:86–9.
Keith DA, Rodríguez JP, Rodríguez-Clark KM, Nicholson E, Aapala K, Alonso A, et al. Scientific foundations for an IUCN red list of ecosystems. PLoS One. 2013;8:e62111. This seminal paper outlines the structure for assessing the conservation status of ecosystems and thereby taking formal IUCN assessment processes beyond species.
Burns EL, Lindenmayer DB, Stein JA, Blanchard W, McBurney L, Blair D, et al. Ecosystem assessment of mountain ash forest in the Central Highlands of Victoria, south-eastern Australia. Aust Ecol. 2014;40(4):386–99.
Carpenter SR, Brock WA. Early warnings of unknown nonlinear shifts: a nonparamteric approach. Ecology. 2011;92:2196–201.
Drake JM, Griffen BD. Early warning signals of extinction in deteriorating environments. Nature. 2010;467:456–9.
Dai L, Korolev KS, Gore J. Slower recovery in space before collapse of connected populations. Nature. 2013;496:355–8.
Schreiber S, Rudolf HW. Crossing habitat boundaries: coupling dynamics of ecosystems through complex life cycles. Ecol Lett. 2008;11:576–87.
Hastings A, Wysham DB. Regime shifts in ecological systems can occur with no warning. Ecol Lett. 2010;13:464–72.
Boettiger C, Hastings A. From patterns to predictions. Nature. 2013;493:157–8. This short article highlights the extreme difficulty of predicting ecosystem collapse.
Tylianakis JM, Didham R, Bascompte J, Wardle DA. Global change and species interactions in terrestrial ecosystems. Ecol Lett. 2008;11:1351–63.
Hunter ML, Bean MJ, Lindenmayer DB, Wilcove DS. Thresholds and the mismatch between environmental laws and ecosystems. Conserv Biol. 2009;23:1053–5.
Steffen W, Richardson K, Rockström J, Cornell S, Fetzer I, Bennett EM, et al. Planetary boundaries: guiding human development on a changing planet. Science. 2015;347:736–45.
Acknowledgments
I thank Dr Rebecca Tittler for inviting me to write this review and providing insightful comments on earlier versions of the manuscript. Comments by two anonymous referees improved an earlier of this manuscript. Claire Shepherd provided editorial support for manuscript submission.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest Statement
Professor Lindenmayer has no conflicts of interest to declare.
Additional information
This article is part of the Topical Collection on Interaction of Landscape Structure and Natural Resource Management
Rights and permissions
About this article

Cite this article
Lindenmayer, D. Interactions between Forest Resource Management and Landscape Structure. Curr Landscape Ecol Rep 1, 10–18 (2016). https://doi.org/10.1007/s40823-016-0002-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40823-016-0002-0