
Abstract
The asymmetric grandparental investment in humans may ultimately be explained by the paternity uncertainty hypothesis. The proximate mechanisms leading to grandparental bias in investment in grandchildren are, however, unclear. In a study of 233 males and females with an opposite sexed sibling, we examined whether comments on resemblance regarding one’s own child, or one’s sibling’s child, changed in frequency after both siblings became parents. We found that comments among siblings on resemblance of children occurred more frequently after both became parents, compared to when only one of the siblings had children, suggesting that resemblance descriptions may become more important after both siblings have children. Furthermore, and in line with the suggestion that mothers may mentally exploit the alloparenting environment by holding a stronger belief about resemblance, brothers reported that their sisters commented on resemblance concerning their own child more often and more intensely. Additionally, sisters corroborated this finding by self-reporting that they were the most proactive during resemblance descriptions of their brothers’ child. Thus, sisters might, through more frequent voicing of stronger opinions on parent–child resemblance than their brothers, influence alloparents’ perception of resemblance to their children and thus influence alloparental investments.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Evolutionary theory recognizes that it is not the species or even the individual, but the gene itself that is the unit of selection. In other words, a gene’s, and in turn an organism’s fitness, is determined by how large its (relative) genetic contribution is to the next generation. Thus, rather than investing in unrelated individuals, parents and grandparents should invest in offspring with whom they share genes (Hamilton, 1964). Parents are equally related to each of their biological children (i.e., they share 50% of the same genes) whereas full siblings, on average, share 50% of their genes with each other (i.e., in theory from 0 to 100%). Each child, on the other hand, is 100% related to itself (Hamilton, 1964). Thus, in general, the parent’s best fitness decision is to invest equally in all offspring, while each offspring gains more by redirecting resources toward themselves at the expense of their siblings (for details, see Trivers, 1974). This conflict in resource distribution between parents and offspring (i.e., the “parent–offspring conflict”) is thus a result of clear genetic constraints (Trivers, 1974).
The long period of parental care and the long post-reproductive lifespan of females (Hawkes et al., 1998) in humans are unique among primates. Long post-reproductive lifespans are rare (Ellis et al., 2018) and evolve when the old individuals still can benefit their younger relatives (Hamilton, 1966). According to Hamilton’s (1964) rule, grandparents should switch investments from their children to their grandchildren (in order to maximize their inclusive fitness) when the fitness outcome from investing in a grandchild is more than twice the fitness outcome from investing in their own child. Although one might expect the grandparents to invest in each grandchild equally, this is not the case. As Euler and Weitzel suggested in 1996, grandparents’ investment in grandchildren might be influenced by paternal uncertainty.
Parental certainty differs between males and females, and male mate guarding, rather than paternal care, most likely fueled our tendency for monogamy (Schacht & Bell, 2016). The explanation for such evolution can be found by looking at human reproductive strategies, with risks of cuckoldry from promiscuous rival infanticidal males (Opie et al., 2013) and hidden ovulation, internal fertilization, and infidelity in females (Benshoof & Thornhill, 1979). Consequently, while the mother can be completely certain that she is the mother of her child, it is impossible under natural circumstances for the father to know if he is the biological father of his putative child (Neel & Weiss, 1975). This paternity uncertainty also includes grandparents (Smith, 1988), and it has repeatedly been invoked when explaining skewed grandparental investments (Bishop et al., 2009; Euler & Weitzel, 1996; Fischer, 1983; Gibson & Mace, 2005; Michalski & Shackelford, 2005; Pashos & McBurney, 2008; Pollet et al., 2009). The maternal grandmother is the most certain of the kinship to their grandchildren, whereas the paternal grandfather is the least certain. This may cause the well-documented, unequal distribution of grandparental investments. That is, the maternal grandmother (with no uncertain genetic links between herself and her grandchildren) shows the highest investment in her grandchildren whereas the maternal grandfather and paternal grandmother (both with one uncertain genetic link) invest at similar intermediate levels. Investing the least in his grandchildren is the paternal grandfather, with two uncertain genetic links (Bishop et al., 2009; Euler & Weitzel, 1996; Fischer, 1983; Gibson & Mace, 2005; Laham et al., 2005; Pashos, 2000; Pollet et al., 2009; Smith, 1988). Moreover, physiological constraints may on a proximate level add to these findings as old males have the option to increase their fitness by mating with additional partners, while post-menopausal females of the same age are no longer able to produce children on their own. Thus, females are, unlike males, physiologically constrained to increase their fitness by investing in existing children or grandchildren when they get old. Therefore, grandmothers in general are expected to invest more in grandchildren than grandfathers (Euler & Weitzel, 1996). Grandparental investment, as well as parental investment, is a limited resource, and grandparental investment has proven to have positive effects—even on the grandchildren’s survival chances (Gibson & Mace, 2005; Sear, 2008; Sear et al., 2000).
Natural selection will favor fathers who can separate genetic kin from non-genetic kin, and the ability to detect phenotypic resemblance, especially facial resemblance, seems to have evolved as a kin recognition cue (Alvergne et al., 2007, 2009, 2010; Apicella & Marlowe, 2004; Daly & Wilson, 1982; Heijkoop et al., 2009). That is, the father’s investment decisions of importance for the child’s fitness are influenced by the father’s experience of facial resemblance to the child (Alvergne et al., 2009; Apicella & Marlowe, 2004; Heijkoop et al., 2009; Platek et al., 2002, 2003). Moreover, the underlying brain domains for facial resemblance show sex-specific differences in activation with males responding stronger to self-morphed child pictures than females (Platek et al., 2004; Wu et al., 2013). Even facial specifics, such as eye-color, which may show a direct concordance between genotype and phenotype, have been invoked as a cue of paternity insurance for blue-eyed males preferring blue-eyed females in populations where genes for blue eye-color exist in low frequencies. That is, blue-eyed parents (i.e., fathers) should get blue-eyed children (Laeng et al., 2007) and thus eye color could ensure genetic relatedness as a sort of “green beard” (popularized by Dawkins, 1976). These observations on associations between paternity certainty and eye-color are also well established in folklore, as for example by the Norwegian Nobel Prize-winning author Knut Hamsun’s descriptions of the wealthy land-lord Mack’s many extramarital brown-eyed children in the local fishing community, and the infertile Oliver Anderson’s frantic reaction to his blue-eyed daughter in the novel “The Women at the Pump” (see Busch, 2012). However, maintaining an accurate mental representation of one’s own many facial features might be difficult (Epley & Whitchurch, 2008; White et al., 2016) and, as the use of mirrors for facial self-identification is a relatively recent invention, fathers might for long have been dependent on facial features of putative close kin for evaluating facial familiarity with children (Bressan & Zucchi, 2009; DeBruine, 2002; Smith, 1988). Additionally, verbal descriptions from external judges—a “verbal mirror”—might have been important for fathers, and perhaps also grandfathers, for perceiving self-resemblance to offspring (Burch & Gallup, 2000; Busch et al., 2018; Daly & Wilson, 1982; Regalski & Gaulin, 1993). Yet, such “verbal mirrors” may not be entirely accurate.
Mothers, who seem to wish for their babies to look like their fathers (Leifer, 1977), are repeatedly found to ascribe resemblance of their infants to putative fathers rather than to themselves, while the opposite does not seem to occur (Alvergne et al., 2007; Daly & Wilson, 1982; McLain et al., 2000; Regalski & Gaulin, 1993). For example, McLain et al. (2000), who showed such biased descriptive behavior among mothers, also documented that resemblance descriptions of newborns to putative fathers was accentuated when these fathers were present and reduced when they were absent. Yet, impartial judges of pictures of the same newborns more often correctly paired newborns with their mothers than with their putative fathers, suggesting that the mother’s verbal ascription of resemblance was not consistent with actual resemblance (McLain et al., 2000). Thus, the alleged resemblance from mothers may not always correspond to the perceived resemblance among fathers and the assertions from mothers are also something fathers often negatively react to (Regalski & Gaulin, 1993). While paternal skepticism complies with the observations of the impartial judges of the facial photographs (Alvergne et al., 2007; McLain et al., 2000), and consequently might be parsimoniously understood on empirical grounds, maternal persuasion does not. That is, the overemphasis on paternal resemblance among mothers may be an evolved deceptive manipulative strategy to counter paternal uncertainty in the service of encouraging fathers to invest in child-rearing (Daly & Wilson, 1982; McLain et al., 2000; Regalski & Gaulin, 1993) rather than harming them (Alexandre et al., 2011; Bellis et al., 2005; Burch & Gallup, 2000). Is it possible that mothers, in the same way as they exaggerate their child’s resemblance to the putative father, have evolved behaviors to influence grandparents’ investment decisions in grandchildren?
Recently, it has been shown that grandparents, with grandchildren among both sons and daughters, report that their daughters more frequently and also more intensely than their sons verbally address the subject of resemblance (Busch et al., 2018). It was consequently not surprising that the majority of grandparents, in the latter study, also reported their daughters’ children as most similar to themselves, both physically and psychologically. Grandparents’ investment in grandchildren is positively influenced by grandchildren’s resemblance to them, and the largest positive effect of resemblance on grandparental investment can be seen among the grandparents least certain of genetic relatedness—the paternal grandfathers (Schlee & Kirchengast, 2015). Thus, the manipulative and biased resemblance descriptions, repeatedly documented at large effect sizes (Alvergne et al., 2007; Daly & Wilson, 1982; McLain et al., 2000; Regalski & Gaulin, 1993), may not be limited to mothers’ vocalization of father-child similarities. Mothers may also mentally exploit the alloparenting environment by expressing biased resemblance descriptions when accompanied by their parents (their children’s grandparents), i.e., the “manipulative mother hypothesis” (Busch et al., 2018). Such verbal influence of grandparents’ need not be premeditated as mothers also seem to believe that their children more resemble their grandmother than sons do (Busch et al., 2018).
Here, we extend the evaluation of the “manipulative mother hypothesis” and examine whether there is a difference in how brothers and sisters perceive and self-report vocal expressions about resemblance or dissimilarities between themselves and their child. More specifically, we explore (i) whether comments on resemblance become more frequent after both siblings become parents, compared to when only one of them had children; and (ii) whether there is a difference in the frequency and intensity of comments made about parent–child resemblance between opposite sexed siblings. The former question relates to the intensity of sibling rivalry, which is expected to increase after both siblings become parents, whereas the latter relates to sex-specific differences in potentially manipulative activities. That is, as grandparents in the examined population report that daughters more often and more intensely than sons express opinions about resemblance between their children and their parents, i.e., about grandchild-grandparent resemblance (Busch et al., 2018), we expect that brothers perceive sisters being most active in resemblance vocalization. We also added specific evaluations about dissimilarity descriptions and about the social context of resemblance descriptions, i.e., whether grandparents were present when children were described. The study was done among siblings with young children in a well-informed population exposed to long-term conscience rising about gender equality in a society relatively abundant in resources—making the study conservative.
Methods
Participants
The administration of private and public kindergartens in the largest populated towns in the northern parts of Norway (i.e., in the counties of Trøndelag, Nordland and Troms and Finnmark) were indiscriminately contacted by email and asked to forward an email with a link to a questionnaire (see supplementary file 1) to all employees and the parents of all children attending the kindergarten. The questionnaire, made by us, was sent to kindergartens because parents with young children might have a recent memory of comments concerning their child’s resemblance. Only individuals that had one sibling of the opposite sex, where either one or both had at least one biological child, could participate in the study. That is, the individuals included were unilaterally reporting about their one sibling of the opposite sex. After excluding 12 parents (11 females and one male), because the participant reported having more than one sibling, or inconsistencies in their answers to the questionnaire, a total of 233 people (see Table 1 for descriptive statistics): 177 females (mean age in years = 34.6 ± 8.1 SD) and 56 males (mean = 37.2 ± 7.1 SD) were included in the analysis. The recruitment of a larger number of female respondents than males might simply be explained by the questionnaire reaching out to more females than males when sending it from kindergartens. That is, the administrators of kindergartens reported that approximately 50% of the parents had registered the email addresses of both mother and father, while 40% only had the mothers and 10% only had the fathers (personal observations, GLHF). Thus, the skewed sex distribution of participants does not need to indicate unrepresentative sampling or biased respondance rate among the sexes. The age of the children was rounded off to the nearest 0.5 year, with a mean age of 8 (SD = 7.5) years for the participants’ children and 11 (SD = 8.3) years for the children of their sibling. The participants were informed that the author (GLHF) was writing a master thesis on family resemblance, and were asked to answer a questionnaire about the frequency and intensity of comments on resemblance to their children, among themselves and their sibling.
Questionnaire
A questionnaire (in Norwegian, translated in the supplementary file) consisting of 42 questions was made using “Google Forms”, An electronic questionnaire was chosen to make it easy to ensure anonymity and to reach out to a large number of people (Norway has a well-developed communication standard with a high frequency of regular users among the targeted age segment). Participants received the questionnaire by email from their kindergarten and answered it on the Internet. All data were collected between February and November 2016.
In the first part of the questionnaire, the participants were asked to report the age and sex of themselves, their opposite sexed sibling, their firstborn child, and/or the firstborn child of their sibling. If the participant and/or their sibling had more than one child, they were asked to answer all questions with regard to the firstborn child. They were also asked about possible confounders, such as whom they considered the most easily offended, themselves or their sibling, the travel time between themselves and their parents, and about how often their children were in contact with their grandparents.
In the second part of the questionnaire, most questions were asked in pairs. The participants were first asked to answer a question about themselves (e.g., how many children do you have?), followed by the same question regarding their sibling (e.g., how many children does your sibling have?). In the last part of the questionnaire, the participants were, in separate questions, asked to score the physical (e.g., facial and body characteriatics) and psychological (e.g., personality traits, interests and skills) resemblance between themselves and their child on a scale from one (i.e., the lowest resemblance) to seven (i.e., the highest resemblance). Afterwards, the participants answered questions about the occurrence and frequency of comments made on resemblance or dissimilarities between themselves and their children, and who made these comments. They were then asked if they could remember whether their sibling ever had made comments on resemblance or dissimilarities between themselves and their own child, or between the participant and his or her own child. In cases were both siblings had commented on child resemblance or dissimilarity, they were asked who had done so most often and most intence. In a forced-choice manner, the participants were thereafter asked whom they thought showed the highest resemblance; themselves and their own child, or their sibling and their own child. In the last question, they were asked if they could remember any change in the frequency of comments on resemblance from the time when only one of them had a child until both became parents (see supplementary file 1).
After finishing the questionnaire, the participants were given the choice to participate in the draw of two VISA gift certificates of 500 NOK (approximately 56 USD), by sending their contact information by email. The contact information could not be connected to the answers of the participant, so their answers remained anonymous. Consequently, the study needed no ethical approval under Norwegian regulations.
Statistical Analysis
The sample size for the different analyses varies as some respondents reported that only one of the siblings had children. That is, some participants could only answer questions regarding either their own child or their sibling’s child. Additionally, not all respondents answered all questions. Chi-square tests were used to examine whether there was any difference between the sexes in how they commented on resemblance or non-resemblance. For example, when asking: Have you or your sibling ever commented that your child resembles you (question 30 in supplementary file 1), there are five alternative answers (i.e., “both”, “only I”, “only my sibling”, “no one”, and “I do not have children”). In the subsequent analysis of the response to this question, only individuals that responded “both” ,“only I”, or “only my sibling” were included, and the test result is consequently reported with 2 degrees of freedom. All statistical tests were two-tailed and the null hypothesis was rejected at an α-level of 0.05. They were all run in R (R Development Core Team, 2018).
Results
Change in Frequency of Comments Before and After Both Siblings Became Parents
The participants were asked whether there was any change in the frequency of comments made on resemblance between parent and the child from the time when only one of the siblings had children until both siblings had become parents. While 40% of the siblings that had an opinion on this question reported that no change had occurred, 60% reported that they remembered a change in the frequency of comments (χ2 = 4.2, df = 1, p = 0.04, 42 reported no change, while 63 reported change). Furthermore, among the subsample of siblings reporting a change in frequency of comments after both siblings became parents, it was most often reported that resemblance vocalizations became more frequent, rather than less frequent (χ2 = 32.14, df = 1, p < 0.01, nine reported more comments before and 54 reported more comments after). There was, however, no difference between brothers and sisters in whether they reported increased or decreased frequency of comments (χ2 = 2.14, df = 1, p = 0.14, 13 males and 50 females). Among the respondents that became parents prior to their sibling in this subsample, 92.6% of the sisters and 66.7% of the brothers reported that the frequency of comments increased after their sibling became a parent. Among respondents that became parents after their sibling, 90.9% of the sisters and 70% of the brothers reported the same. There was no significant difference in reporting between brothers that became parents first, compared to brothers that became parents last (χ2 < 0.01, df = 1, p = 1, three had children first and 10 had children last), nor were there any differences among sisters (χ2 < 0.01, df = 1, p = 1, 28 had children first and 22 had children last).
Comments Made by Parents on Resemblance and Dissimilarity
More sisters (10.4%) than brothers (3.8%) reported that their own child had been described as dissimilar by another family member, yet this difference was not statistically significant (χ2 = 1.46, df = 1, p = 0.23, 53 males and 163 females). Moreover, 11.2% of sisters reported that their brother’s child had been described as dissimilar, while none of the brothers could remember to have ever heard a family member describe the sister’s child as dissimilar. This difference between the sexes was, however, not significant (χ2 = 1.55, df = 1, p = 0.21, 22 males and 107 females).
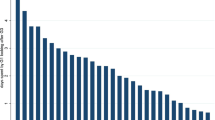
When asked if they or their opposite sexed sibling had commented on resemblance between parent and child, there was a tendency for sisters to be more outspoken about resemblance than brothers (Fig. 1). That is, sisters were more often than brothers reported to be the only sibling to comment on resemblance between both themselves and their own child (5.4% of brothers and 13.8% of sisters; χ2 = 19.21, df = 3, p < 0.01, 56 males and 174 females), and between their sibling and the sibling’s child (9.8% of brothers and 18.3% of sisters; χ2 = 19.18, df = 3, p < 0.01, 43 males and 126 females). When both brothers and sisters had made comments on resemblance, both brothers (24.8%) and sisters (53.5%) reported that the sister was the one who had commented on the resemblance most often and most intensely (Fig. 1A; Table 1). A similar pattern emerged regarding comments on resemblance between the opposite sexed siblings and his or her child. That is, both brothers (35.5%) and sisters (44.7%) reported that females (i.e., sisters or themselves, respectively) commented on the resemblance most often and most intensely (Fig. 1B; Table 1).

Bar plots showing the sex-difference in frequency and intensity of comments on child resemblance. That is, when interviewees (The Participant) answered that both themselves and their sibling of the opposite sex (The Sibling) had made comments on resemblance, they were subsequently asked about who had made such comments most often and most intense. (A) Comments about one’s own child: In these cases, brothers (Males) reported that their sister (The Sibling) had commented most often and most intense, while sisters (Female) not found more proactive reporting among brothers (The Sibling). (B) Comments about their niece’s/nephew’s resemblance to their sibling: In these cases sisters (The Participant, Female) self-reported to have commented most often and most intense, wheras brothers (The Participant, Male) reported that their sibling sister commented most often and intence. Interviewees that reported no difference between themselves and the opposite sexed sibling are included in “Both the same.”
The same tendency was not apparent when the participants were asked if they or their sibling ever had commented on dissimilarity between the parent and the child. When both siblings had made comments on dissimilarities to the respondent’s child, only 16.7% of the brothers and 17.7% of the sisters reported that their sibling was the one who commented on dissimilarities most often and most intensely (Table 1). Yet, when both the respondent and the sibling had commented on dissimilarities of their sibling’s child, none of the brothers while 17.6% of the sisters reported they did so most often and most intensely (Table 1).
Comments on the Resemblance in the Presence of Grandparents
When reporting about comments on resemblance or dissimilarities between parent and child in the presence of grandparents, there was a tendency for sisters to do so more often than brothers. That is, 21.4% of the brothers had never made comments on resemblance between their sister and her child, while only 10.9% of the sisters reported to have never commented on their brother and his child. Sisters also more often reported to have commented on dissimilarities to the brother’s child (6.4%) in the presence of grandparents, than what brothers reported for their sister’s child (3.6%). Yet, most of the comments that were made were pinpointing resemblance. The majority of both brothers (79.5%) and sisters (79.1%) reported having commented on resemblance between themselves and their own child in the presence of grandparents, while few sisters (14%) and brothers (6.8%) made comments on dissimilarities. This difference was however not significant (Table 2). Moreover, sisters commented on resemblance concerning their brother’s child more often than brothers commented on resemblance of their sister’s child in the presence of grandparents, but also this difference was not statistically significant (Table 2). When asked if their sibling had commented on resemblance between the sibling and the sibling’s child in the presence of grandparents, brothers (85%) more often than the sisters (70%) reported that their sibling had made comments on resemblance. This difference was however not significant (Table 2). There was also no difference in how often siblings made comments on resemblance concerning the participant and the participants’ child (Table 2).
When the sister of a male participant did not have children of her own, she tended to make fewer comments on the resemblance of her brother and his child. That is, 20% of brothers whose sister did not have children reported that the sister had never made comments on resemblance in the presence of grandparents, while only 7.4% reported that the sister did not comment on resemblance when the sister had children of her own (χ2 = 0.24, df = 1, p = 0.62, 27 sisters had children and 10 sisters did not have children). Conversely, 15.4% of sisters whose brother did not have children reported that the brother had never made comments on resemblance in the presence of grandparents, while 18.9% reported that the brother had not commented on resemblance when the brother had his own children (χ2 = 0.02, df = 1, p = 0.90, 79 brothers had children and 26 brothers did not have children).
Limitations
There seem to be few biases in our sample, e.g., there was no difference in estimated travel time to grandparents between siblings, and brothers and sisters did not systematically differ in how often their children met their grandparents (see supplementary file 2 for details). Although a larger proportion of participants in this study were females, this does not need to be caused by self-selection. As there are mostly female employees in Norwegian kindergartens (Digre, 2016) with access to contact information of more mothers than fathers, more females should receive our questionnaire. Additionally, it cannot be excluded that females, even in a relatively gender-equal society such as Norway, are more interested in participating in our study because they are more involved in their children’s everyday life than males (Lorentzen & Lappegård, 2009). Still, our possibly skewed recruitment procedure produces samples of brothers and sisters, with relatively young children, at quantities that give statistical significance to meaningful behavioral effects. Most importantly, however, there is a strong consensus in the independent reports from both brothers and sisters, suggesting that our results do not stem from a biased sampling of one sex.
Discussion
The majority of siblings reported a change in the frequency of comments on child resemblance after both siblings became parents, and among those remembering such change, most reported that resemblance vocalizations had increased. Brothers also reported that sisters made comments on resemblance more often and more intensely than themselves, something corroborated by the unilateral reporting by sisters. Yet, there was no clear difference between brothers and sisters in resemblance reporting when accompanied by their parents (i.e., their children’s grandparents) or when specifically asked about their reporting on children’s dissimilarities to parents.
Increased Talk About the Resemblance
Regardless of who first became a parent, both brothers and sisters who remembered a change in frequency of comments on resemblance report that it tended to occur more often after both siblings had become parents. The most parsimonious explanation for such change is that more children (and grandchildren) lead to more conversation about children and children’s resemblance. Yet, the self-reported change in the frequency of comments on resemblance might also indicate that the siblings’ competition for alloparental investments gets more intense after both siblings become parents. As the parent–offspring conflict is predicted to increase with increasing needs among offspring, and so also the offspring’s employment of psychological weapons (Trivers, 1974), one should expect siblings’ competition for the alloparental investment to intensify when both siblings become parents. It is, however, impossible for us to single out which of these explanations that might have the largest impact in our sample.
Sensitivity
Both brothers and sisters frequently make comments on resemblance, yet sisters are reported to make these comments more often and more intensely than what brothers do. Kin resemblance can be a sensitive issue, and males and females may differ in sensitivity to comments made on parent–child resemblance. Thus, if brothers were more easily offended (i.e., “touchier”) than their sisters, it could explain why brothers report that sisters comment on resemblance more often and more intensely than themselves. Yet, there was no difference in reported “touchiness” between brothers and sisters (see supplementary file 2 for details), suggesting that differences in sensitivity to comments were not causing the sex-difference in the perception of comments on parent–child resemblance. Sisters even self-reported that they most often and most intensively talked about the resemblance. Thus, sex-specific differences in perceptional biases seem to be an unlikely explanation for our results.
Similarity and Dissimilarity
We have not been able to disentangle the precise wording in the resemblance descriptions and so far only know (as reported by grandparents (Busch et al., 2018), brothers and sisters themselves) that females are more proactive than males in descriptions of resemblance. This, alone, is interesting. Yet, in order to figure out the exact nature of these resemblance descriptions, one probably need to conduct in-depth interviews and not surveys—as we currently have done (both methods have their pros and cons). Comments about resemblance can be of various forms (e.g., “my child resembles our father more than your child”) without pinpointing either dissimilarity or similarity. Moreover, descriptions of dissimilarity between parents and children can, in some contexts, be considered inappropriate and consequently not something easily admitted to by either parents or grandparents. This latter point may have reduced our ability to acquire samples giving appropriate statistical power on our most specific analysis—that of differences in dissimilarity vocalizations. That is, males and females rarely report their sibling to have made comments on dissimilarities between the parent and the child, neither in the presence of their sibling nor in the presence of grandparents. However, females more than males voice their stronger opinions about the parent–child resemblance, also about brothers’ children. The latter corresponds with our hypothesis and with results from the many studies on parents with newborns (Alvergne et al., 2007; Daly & Wilson, 1982; Regalski & Gaulin, 1993), some of which also reports that the resemblance reporting is context-dependent (McLain et al., 2000). That is, they report that the frequency of mothers’ descriptions about resemblance between father and child increase in the presence of putative fathers. These predominantly female vocalizations may, during our evolution, have served to increase putative fathers resource investment in children, to reduce the negative effects related to paternity uncertainty among fathers (e.g., child neglect and abuse) and also to reduce the risks related to infidelity among mothers.
Grandparents
The siblings in our study do not report sex-differences in verbal communication about child resemblance when accompanying parents. Yet, grandparents from the same population have reported their daughters to argue this resemblance more often and more intensively than their sons (Busch et al., 2018). Why this inconsistency? One possible explanation could be that the Busch et al. (2018) study, as any one unreplicated study, might have produced results out of random or some form of unknown context dependence (O’Dea et al., 2021). Yet, in the study, both maternal grandmothers and maternal grandfathers concurrently reported daughters, rather than sons, more often and more intensely talking about resemblance. Consequently, this inconsistency may rather stem from mothers arguing familiar resemblance to their parents when they are not accompanied by their brothers. As males have a tendency of ranking their child’s resemblance to themselves higher than what females do (Platek et al., 2002, 2005; Wu et al., 2013) and clearly recognize their sisters’ proactive verbal resemblance descriptions (this study), it seems unlikely that sisters’ resemblance descriptions in the company of parents would have gone unnoticed by brothers. Thus, given the present empirics, our best explanation for a more proactive description of resemblance by the daughter, as reported by grandparents, seems to be a context-dependent sex-specific bias in resemblance descriptions, i.e., that sisters voice their opinions about resemblance more often when brothers are absent.
Proactive Sisters
Our observation of a rather more proactive sister than brother in verbal descriptions of their children’s resemblance fit the earlier observations from research on parental resemblance descriptions of newborns (Alvergne et al., 2007; Daly & Wilson, 1982; McLain et al., 2000; Regalski & Gaulin, 1993) and that of parental activity in resemblance descriptions of grandchildren as observed by grandparents (Busch et al., 2018). Some of this maternal behavior has since 1982 (Daly and Wilson) been termed “manipulative”, and proactive verbal resemblance descriptions might in our “environment of evolutionary adaptedness” (Bowlby, 1969), yet after language evolved, have enabled females in a very vulnerable position to acquire resources (e.g., food and protection) for themselves and their newborns (see Campbell, 1999). That is, in a hunter-gatherer community where the net positive societal caloric income most likely was positive (and large) for males (i.e., putative fathers and grandfathers aged 20 to 60 years) and (though smaller) older females (i.e., grandmothers approximately 45 to 70 years-old) (Kaplan et al., 2000), biased verbal communication of children’s resemblance might have given increased access to vital needs and infanticidal protection. Although relatively naïve male targets might have coevolved scepticism to counter mother’s alleged resemblance (e.g., Regalski & Gaulin, 1993), novice female alloparents, on the other hand, might have more easily recognized mothers’ vocal behavior as manipulative. The latter recognition might be accepted by maternal grandmothers, with large fitness investments at stake from loosing paternal investments, but might also have caused resentment, particularly from doubtful paternal grandmothers. Thus, it should not be surprising that the paternal grandmothers are repeatedly found to invest less in grandchildren compared to that of the equally uncertain maternal grandfathers (Euler & Weitzel, 1996; Laham et al., 2005; Pashos & McBurney, 2008; Pollet et al., 2009), that the relationship between daughter-in-law and mother-in-law has repeatedly been reported to be the least healthy of all parent–grandparent relationships (Ayers et al., 2022; Euler et al., 2009; Fingermann, 2004; Lee et al., 2003; Sherlip & Stricker, 1998), that wives of brothers have been considered “classic rivals” (Hammel & Gullickson, 2004), and, last, why parenthood improves emotional closeness and contact rate between daughters and mothers (Danielsbacka et al., 2015; Tanskanen, 2017) while decreasing emotional closeness and intimacy between sons and mothers (Tanskanen, 2017). Furthermore, manipulations by mothers and a subsequent identification of manipulative behavior by other females might be important for generating the often observed reproductive competition among females (Vaillancourt, 2013), a competition that might have large negative fitness effects (Cant & Johnstone, 2008; Hammel & Gullickson, 2004; Lahdenpera et al., 2012; Pettay et al., 2016) even within kin groups (Ji et al., 2013; Mace & Alvergne, 2012; Sear, 2008; Willfuhr et al., 2018).
General Perspectives
Given fitness differences from behaviors in our “environment of evolutionary adaptedness”, it is no surprise that brains of males and females have evolved to differ (Chekroud et al., 2016; Del Giudice et al., 2016; Geary, 2017; Prager, 2017 entire volume), something observed even before sex-specific environments impact brain development (Wheelock et al., 2019), and the work of understanding how these genetically determined differences map onto sexual differences in behavior have started (Del Giudice et al., 2012; Ingalhalikar et al., 2014). For example, not surprisingly females seem to have larger connectivity between brain regions associated with motivation for obtaining social rewards and avoidance of social punishment than males (Tunc et al., 2016). These findings within neuroscience correspond to many of the findings in behavioral biology (Björkqvist et al., 1992; Gur et al., 2012; Ritchie et al., 2018), and it is in this landscape of observed anatomical sex-differences one could understand our observations of sex differences in behavior on a proximate level. While we may speculate that the manipulative behavior itself may be unconscious and not premeditated by mothers, the recognition and response to it may tap into consciousness. Yet, it might be that these differences are quite malleable, but their malleability will first be tested after we have “seen” them, unbiasedly examined their importance for family dynamics, and then confronted them.
Transparency and Reproducibility
The data has been uploaded for common use from UiT Open Research Data at: https://dataverse.no/dataset.xhtml?persistentId=doi:10.18710/CV1NQB&version=DR%20603%20AFT.
Change history
29 August 2022
Missing Open Access funding information has been added in the Funding Note.
References
Alexandre, G. C., Nadanovsky, P., Wilson, M., Daly, M., Moraes, C. L., & Reichenheirn, M. (2011). Cues of paternal uncertainty and father to child physical abuse as reported by mothers in Rio de Janeiro, Brazil. Child Abuse & Neglect, 35(8), 567–573. https://doi.org/10.1016/j.chiabu.2011.04.001
Alvergne, A., Faurie, C., & Raymond, M. (2007). Differential facial resemblance of young children to their parents: Who do children look like more? Evolution and Human Behavior, 28(2), 135–144. https://doi.org/10.1016/j.evolhumbehav.2006.08.008
Alvergne, A., Faurie, C., & Raymond, M. (2009). Father–offspring resemblance predicts paternal investment in humans. Animal Behaviour, 78, 61–69. https://doi.org/10.1016/j.anbehav.20
Alvergne, A., Faurie, C., & Raymond, M. (2010). Are parents’ perceptions of offspring facial resemblance consistent with actual resemblance? Effects on parental investment. Evolution and Human Behavior, 31(1), 7–15. https://doi.org/10.1016/j.evolhumbehav.2009.09.002
Apicella, C. L., & Marlowe, F. W. (2004). Perceived mate fidelity and paternal resemblance predict men's investment in children [Article]. Evolution and Human Behavior, 25(6), 371–378. https://doi.org/10.1016/j.evolhumbehav.2004.06.003
Ayers, J. D., Krems, J. A., Hess, N., & Aktipis, A. (2022). Mother-in-law daughter-in-law conflict: An evolutionary perspective and report of empirical data from the USA. Evolutionary Psychological Science. https://doi.org/10.1007/s40806-021-00312-x
Bellis, M. A., Hughes, K., Hughes, S., & Ashton, J. R. (2005). Measuring paternal discrepancy and its public health consequences [Review]. Journal of Epidemiology and Community Health, 59(9), 749–754. https://doi.org/10.1136/jech.2005.036517
Benshoof, L., & Thornhill, R. (1979). The evolution of monogamy and concealed ovulation in humans. Journal of Social and Biological Structures, 2(2), 95–106. https://doi.org/10.1016/0140-1750(79)90001-0
Bishop, D. I., Meyer, B. C., Schmidt, T. M., & Gray, B. R. (2009). Differential investment behavior between grandparents and grandchildren: The role of paternity uncertainty. Evolutionary Psychology, 7, 66–77. https://doi.org/10.1177/147470490900700109
Björkqvist, K., Lagerspetz, K. M., & Kaukiainen, A. (1992). Do girls manipulate and boys fight? Developmental trends in regard to direct and indirect aggression. Aggressive Behavior, 18(2), 117–127. https://doi.org/10.1002/1098-2337(1992)18:2
Bowlby, J. (1969). Attachment (Vol. 1). Basic Books.
Bressan, P., & Zucchi, G. (2009). Human kin recognition is self-rather than family-referential. Biology Letters, 5(3), 336–338. https://doi.org/10.1098/rsbl.2008.0789
Burch, R. L., & Gallup, G. G. (2000). Perceptions of paternal resemblance predict family violence. Evolution and Human Behavior, 21, 429–435. https://doi.org/10.1016/S1090-5138(00)00056-8
Busch, H. V. (2012). “Og hun bluedes ikke” En analyse av Knut Hamsuns Edvarda Mack University of Tromsø]. https://hdl.handle.net/10037/4750
Busch, M. V., Olaisen, S., Bruksas, I. J., & Folstad, I. (2018). Do mothers also “manipulate” grandparental care ? Peerj, 6, Article e5924. https://doi.org/10.7717/peerj.5924
Campbell, A. (1999). Staying alive: Evolution, culture, and women’s intrasexual aggression. Behavioral and Brain Sciences, 22(2), 203–214. https://doi.org/10.1017/S0140525X99001818
Cant, M. A., & Johnstone, R. A. (2008). Reproductive conflict and the separation of reproductive generations in humans. Proceedings of the National Academy of Sciences of the United States of America, 105(14), 5332–5336. https://doi.org/10.1073/pnas.0711911105
Chekroud, A. M., Ward, E. J., Rosenberg, M. D., & Holmes, A. J. (2016). Patterns in the human brain mosaic discriminate males from females. Proceedings of the National Academy of Sciences of the United States of America, 113(14), E1968-E1968. https://doi.org/10.1073/pnas.1523888113
Daly, M., & Wilson, M. I. (1982). Whom are newborn babies said to resemble. Ethology and Sociobiology, 3, 69–78. https://doi.org/10.1016/0162-3095(82)90002-4
Danielsbacka, M., Tanskanen, A. O., & Rotkirch, A. (2015). Impact of genetic relatedness and emotional closeness on intergenerational relations. Journal of Marriage and Family, 77(4), 889–907. https://doi.org/10.1111/jomf.12206
Dawkins, R. (1976). The selfish gene. Oxford University Press.
DeBruine, L. M. (2002). Facial resemblance enhances trust. Proceedings of the Royal Society B-Biological Sciences, 269(1498), 1307–1312. https://doi.org/10.1098/rspb.2002.2034
Del Giudice, M., Booth, T., & Irwing, P. (2012). The distance between Mars and Venus: Measuring global sex differences in personality. Plos One, 7(1), Article e29265. https://doi.org/10.1371/journal.pone.0029265
Del Giudice, M., Lippa, R. A., Puts, D. A., Bailey, D. H., Bailey, J. M., & Schmitt, D. P. (2016). Joel et al.’s method systematically fails to detect large, consistent sex differences. Proceedings of the National Academy of Sciences of the United States of America, 113(14), E1965-E1965. https://doi.org/10.1073/pnas.1525534113
Digre, K. (2016). Utdanningsspeilet 2016. udir.no/utdanningsspeilet.
Ellis, S., Franks, D. W., Nattrass, S., et al. (2018). Postreproductive lifespans are rare in mammals. Ecology and Evolution, 8, 2482– 2494. https://doi.org/10.1002/ece3.3856
Epley, N., & Whitchurch, E. (2008). Mirror, mirror on the wall: Enhancement in self-recognition. Personality and Social Psychology Bulletin, 34(9), 1159–1170. https://doi.org/10.1177/0146167208318601
Euler, H. A., Hoier, S., & Rohde, P. (2009). Relationship-specific intergenerational family ties: An evolutionary approach to the structure of cultural transmission. In U. Schönpflug (Ed.), Cultural Transmission: Psycological, developmental, social, and methodological aspects (pp. 70–91). Cambridge University Press.
Euler, H. A., & Weitzel, B. (1996). Discriminative grandparental solicitude as reproductive strategy [journal article]. Hu Nat, 7(1), 39–59. https://doi.org/10.1007/bf02733489
Fingermann, K. L. (2004). The role of offspring and in-laws in grandparents’ ties to their grandchildren. Journal of Family Issues, 25(8), 1026–1049. https://doi.org/10.1177/0192513X04265941
Fischer, L. R. (1983). Transition to grandmotherhood [Article]. International Journal of Aging & Human Development, 16(1), 67–78. https://doi.org/10.2190/gu3r-506f-2umw-6l8r
Geary, D. C. (2017). Evolution of human sex-specific cognitive vulnerabilities. Quarterly Review of Biology, 92(4), 361–410. https://doi.org/10.1086/694934
Gibson, M. A., & Mace, R. (2005). Helpful grandmothers in rural Ethiopia: A study of the effect of kin on child survival and growth [Article]. Evolution and Human Behavior, 26(6), 469–482. https://doi.org/10.1016/j.evolhumbehav.2005.03.004
Gur, R. C., Richard, J., Calkins, M. E., Chiavacci, R., Hansen, J. A., Bilker, W. B., Loughead, J., Connolly, J. J., Qiu, H. J., Mentch, F. D., Abou-Sleiman, P. M., Hakonarson, H., & Gur, R. E. (2012). Age group and sex differences in performance on a computerized neurocognitive battery in children age 8–21. Neuropsychology, 26(2), 251–265. https://doi.org/10.1037/a0026712
Hamilton, W. D. (1964). The genetical evolution of social behaviour. 1. Journal of Theoretical Biology, 7(1), 1–16. https://doi.org/10.1016/0022-5193(64)90038-4
Hamilton, W. D. (1966). The moulding of senescence by natural selection. Journal of Theoretical Biology, 12(1), 12–45. https://doi.org/10.1016/0022-5193(66)90184-6
Hammel, E. A., & Gullickson, A. (2004). Kinship structures and survival: Maternal mortality on the Croatian-Bosnian border 1750–1898. Population Studies-a Journal of Demography, 58(2), 145–159. https://doi.org/10.1080/0032472042000213703
Hawkes, K., O'Connell, J. F., Jones, N. G. B., Alvarez, H., & Charnov, E. L. (1998). Grandmothering, menopause, and the evolution of human life histories [Article]. Proceedings of the National Academy of Sciences of the United States of America, 95(3), 1336–1339. https://doi.org/10.1073/pnas.95.3.1336
Heijkoop, M., Dubas, J. S., & van Aken, M. A. G. (2009). Parent-child resemblance and kin investment: Physical resemblance or personality similarity? European Journal of Developmental Psychology, 6, 64–69. https://doi.org/10.1080/17405620802642306
Ingalhalikar, M., Smith, A., Parker, D., Satterthwaite, T. D., Elliott, M. A., Ruparel, K., Hakonarson, H., Gur, R. E., Gur, R. C., & Verma, R. (2014). Sex differences in the structural connectome of the human brain. Proceedings of the National Academy of Sciences of the United States of America, 111(2), 823–828. https://doi.org/10.1073/pnas.1316909110
Ji, T., Wu, J. J., He, Q. Q., Xu, J. J., Mace, R., & Tao, Y. (2013). Reproductive competition between females in the matrilineal Mosuo of southwestern China. Philosophical Transactions of the Royal Society B-Biological Sciences, 368(1631), Article 20130081. https://doi.org/10.1098/rstb.2013.0081
Kaplan, H., Hill, K., Lancaster, J., & Hurtado, A. M. (2000). A theory of human life history evolution: Diet, intelligence, and longevity. Evolutionary Anthropology, 9(4), 156–185.
Laeng, B., Mathisen, R., & Johnsen, J.-A. (2007). Why do blue-eyed men prefer women with the same eye color? Behavioral Ecology and Sociobiology, 61(3), 371–384. https://doi.org/10.1007/s00265-006-0266-1
Laham, S. M., Gonsalkorale, K., & von Hippel, W. (2005). Darwinian grandparenting: Preferential investment in more certain kin [Article]. Personality and Social Psychology Bulletin, 31(1), 63–72. https://doi.org/10.1177/0146167204271318
Lahdenpera, M., Gillespie, D. O. S., Lummaa, V., & Russell, A. F. (2012). Severe intergenerational reproductive conflict and the evolution of menopause. Ecology Letters, 15(11), 1283–1290. https://doi.org/10.1111/j.1461-0248.2012.01851.x
Lee, E., Spitze, G., & Logan, J. R. (2003). Social support to parents-in-law: The interplay of gender and kin hierarchies. Journal of Marriage and Family, 65(2), 396–403. https://doi.org/10.1111/j.1741-3737.2003.00396.x
Leifer, M. (1977). Psychological changes accompanying pregnancy and motherhood. Genetic Psychology Monographs, 95(1), 55–96. Go to ISI. https://wos.A1977CX99600002
Lorentzen, M., & Lappegård, T. (2009). Likestilling og deling av omsorgsoppgaver for barn (Vol. 42). https://www.ssb.no/a/publikasjoner/pdf/notat_200942/notat_200942.pdf
Mace, R., & Alvergne, A. (2012). Female reproductive competition within families in rural Gambia. Proceedings of the Royal Society B-Biological Sciences, 279(1736), 2219–2227. https://doi.org/10.1098/rspb.2011.2424
McLain, D. K., Setters, D., Moulton, M. P., & Pratt, A. E. (2000). Ascription of resemblance of newborns by parents and nonrelatives [Article]. Evolution and Human Behavior, 21(1), 11–23. https://doi.org/10.1016/s1090-5138(99)00029-x
Michalski, R. L., & Shackelford, T. K. (2005). Grandparental investment as a function of relational uncertainty and emotional closeness with parents [Article]. Human Nature-an Interdisciplinary Biosocial Perspective, 16(3), 293–305. https://doi.org/10.1007/s12110-005-1012-5
Neel, J., & Weiss, K. (1975). Genetic structure of a tribal population, Yanomama Indians. 12. Biodemographic studies. American Journal of Physical Anthropology, 42, 25–52.
O’Dea, R. E., Parker, T. H., Chee, Y. E., Culina, A., Drobniak, S. M., Duncan, D. H., Fidler, F., Gould, E., Ihle, M., Kelly, C. D., Lagisz, M., Roche, D. G., Sánchez-Tójar, A., Wilkinson, D. P., Wintle, B. C., & Nakagawa, S. (2021). Towards open, reliable, and transparent ecology and evolutionary biology. BMC Biology, 19(1), 68. https://doi.org/10.1186/s12915-021-01006-3
Opie, C., Atkinson, Q. D., Dunbar, R. I. M., & Shultz, S. (2013). Male infanticide leads to social monogamy in primates. Proceedings of the National Academy of Sciences of the United States of America, 110(33), 13328–13332. https://doi.org/10.1073/pnas.1307903110
Pashos, A. (2000). Does paternal uncertainty explain discriminative grandparental solicitude? A cross-cultural study in Greece and Germany [Article]. Evolution and Human Behavior, 21(2), 97–109. https://doi.org/10.1016/s1090-5138(99)00030-6
Pashos, A., & McBurney, D. H. (2008). Kin relationships and the caregiving biases of grandparents, aunts, and uncles - A two-generational questionnaire study [Article]. Human Nature-an Interdisciplinary Biosocial Perspective, 19(3), 311–330. https://doi.org/10.1007/s12110-008-9046-0
Pettay, J. E., Lahdenpera, M., Rotkirch, A., & Lummaa, V. (2016). Costly reproductive competition between co-resident females in humans. Behavioral Ecology, 27(6), 1601–1608. https://doi.org/10.1093/beheco/arw088
Platek, S. M., Burch, R. L., Panyavin, I. S., Wasserman, B. H., & Gallup, G. G. (2002). Reactions to children’s faces - Resemblance affects males more than females. Evolution and Human Behavior, 23, 159–166. https://doi.org/10.1016/S1090-5138(01)00094-0
Platek, S. M., Critton, S. R., Burch, R. L., Frederick, D. A., Myers, T. E., & Gallup, G. G. (2003). How much paternal resemblance is enough? Sex differences in hypothetical investment decisions but not in the detection of resemblance. Evolution and Human Behavior, 24(2), 81–87, Article Pii s1090–5138(02)00117–4. https://doi.org/10.1016/s1090-5138(02)00117-4
Platek, S. M., Keenan, J. P., & Mohamed, F. B. (2005). Sex differences in the neural correlates of child facial resemblance: An event-related fMRI study. Neuroimage, 25(4), 1336–1344. https://doi.org/10.1016/j.neuroimaging.2004.12.037
Platek, S. M., Raines, D. M., Gallup, G. G., Mohamed, F. B., Thomson, J. W., Myers, T. E., Panyavin, I. S., Levin, S. L., Davis, J. A., Fonteyn, L. C. M., & Arigo, D. R. (2004). Reactions to children’s faces: Males are more affected by resemblance than females are, and so are their brains [Article]. Evolution and Human Behavior, 25(6), 394–405. https://doi.org/10.1016/j.evolhumbehav.2004.08.007
Pollet, T. V., Nelissen, M., & Nettle, D. (2009). Lineage based differences in grandparental investment: Evidence from a large British cohort study. Journal of Biosocial Science, 41, 355–379. https://doi.org/10.1017/s0021932009003307
Prager, E. M. (2017). Addressing sex as a biological variable. Journal of Neuroscience Research, 95(1–2), 11–11. https://doi.org/10.1002/jnr.23979
R Development Core Team. (2018). R: A language and environment for statistical computing. In R Foundation for Statistical Computing. https://www.R-project.org/
Regalski, J. M., & Gaulin, S. J. (1993). Whom are Mexican infants said to resemble? Monitoring and fostering paternal confidence in the Yucatan. Ethology and Sociobiology, 14(2), 97–113.
Ritchie, S. J., Cox, S. R., Shen, X. Y., Lombardo, M. V., Reus, L. M., Alloza, C., Harris, M. A., Alderson, H. L., Hunter, S., Neilson, E., Liewald, D. C. M., Auyeung, B., Whalley, H. C., Lawrie, S. M., Gale, C. R., Bastin, M. E., McIntosh, A. M., & Deary, I. J. (2018). Sex differences in the adult human brain: Evidence from 5216 UK Biobank participants. Cerebral Cortex, 28(8), 2959–2975. https://doi.org/10.1093/cercor/bhy109
Schacht, R., & Bell, A. V. (2016). The evolution of monogamy in response to partner scarcity. Scientific Reports, 6, Article 32472. https://doi.org/10.1038/srep32472
Schlee, J., & Kirchengast, S. (2015). Differential grandparental investment - The impact of phenotypic resemblance [Article]. Anthropologischer Anzeiger, 72(4), 411–425. https://doi.org/10.1127/anthranz/2015/0523
Sear, R. (2008). Kin and child survival in rural Malawi - Are matrilineal kin always beneficial in a matrilineal society? Human Nature-an Interdisciplinary Biosocial Perspective, 19(3), 277–293. https://doi.org/10.1007/s12110-008-9042-4
Sear, R., Mace, R., & McGregor, I. A. (2000) Maternal grandmothers improve nutritional status and survival of children in rural Gambia. Proceedings of the Royal Society of London. Series B: Biological Sciences, 267(1453), 1641–1647. https://doi.org/10.1098/rspb.2000.1190
Sherlip, D., & Stricker, G. (1998). Expectant granparent’s relationship with the middle generation and their grandchildren. 106th annual confvention of the American Psycological Association., San Francisco, CA.
Smith, M. S. (1988). Research in developmental sociobiology: Parenting and family behavior. In K. B. MacDonald (Ed.), Sociobiological Perspectives on Human Development (pp. 271–292). Springer New York. https://doi.org/10.1007/978-1-4612-3760-0_9
Tanskanen, A. O. (2017). Intergenerational relations before and after offspring arrive: A within-person investigation [Article]. Social Science Research, 67, 138–146. https://doi.org/10.1016/j.ssresearch.2017.08.001
Trivers, R. L. (1974). Parent-offspring conflict. American Zoologist, 14(1), 249–264. Go to ISI. https://wos.A1974T008500016
Tunc, B., Solmaz, B., Parker, D., Satterthwaite, T. D., Elliott, M. A., Calkins, M. E., Ruparel, K., Gur, R. E., Gur, R. C., & Verma, R. (2016). Establishing a link between sex-related differences in the structural connectome and behaviour. Philosophical Transactions of the Royal Society B-Biological Sciences, 371(1688), Article 20150111. https://doi.org/10.1098/rstb.2015.0111
Vaillancourt, T. (2013). Do human females use indirect aggression as an intrasexual competition strategy? Philosophical Transactions of the Royal Society B-Biological Sciences, 368(1631), Article Unsp 20130080. https://doi.org/10.1098/rstb.2013.0080
Wheelock, M. D., Hect, J. L., Hernandez-Andrade, E., Hassan, S. S., Romero, R., Eggebrecht, A. T., & Thomason, M. E. (2019). Sex differences in functional connectivity during fetal brain development. Developmental Cognitive Neuroscience, 36, 100632. https://doi.org/10.1016/j.dcn.2019.100632
White, D., Burton, A. L., & Kemp, R. I. (2016). Not looking yourself: The cost of self-selecting photographs for identity verification. British Journal of Psychology, 107(2), 359–373. https://doi.org/10.1111/bjop.12141
Willfuhr, K. P., Johow, J., & Voland, E. (2018). When the mother-in-law is just as good-Differential mortality of reproductive females by family network composition. Plos One, 13(3), Article e0193252. https://doi.org/10.1371/journal.pone.0193252
Wu, H., Yang, S., Sun, S., Liu, C., & Luo, Y.-J. (2013). The male advantage in child facial resemblance detection: Behavioral and ERP evidence. Social neuroscience, 8(6), 555–567. https://doi.org/10.1080/17470919.2013.835279
Acknowledgements
Thanks to two anonymous referees for constructive comments, and to Nora Eline Slåttebrekk, Mari Veierud Busch, Sissel Kaino, and Torunn Moe for constructive criticism, support, and considerable help in collecting data.
Funding
Open Access funding provided by UiT The Arctic University of Norway. UiT – The Arctic University of Norway supported IF and GLHF during GLHF’s master study. GLHF was later partially funded by “ReiGN: Reindeer Husbandry in a Globalizing North—Resilience, Adaptations and Pathways for Actions,” which is a Nordforsk-funded Nordic Centre of Excellence (project number 76915) and Fram Centre funded project “Cooperative solutions to common problems” (project number 369902).
Author information
Authors and Affiliations
Contributions
GLHF collected all data, did the initial statistical analysis, and wrote the first draft. BJB helped with the statistical analysis. IF initiated the study as a master thesis for GLHF. All authors edited the many final drafts.
Corresponding author
Ethics declarations
Ethics Approval
The data were collected anonymously and the study needed no ethical approval under Norwegian regulations.
Consent for Publication
All authors approved this submission.
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Significance Statement
Parental care is a limited resource and mothers are repeatedly documented to communicate deceptive judgements about putative fathers’ resemblance to newborns, a behavior termed “manipulative”. From questioning parents with a sibling of the opposite sex, we document that resemblance vocalizations about offspring become more common when both siblings become parents, compared to when only one of them is a parent. More important, brothers report that their sisters more often and more intensely than themselves voice their opinions about parent-child resemblance—a finding also corroborated by sisters self-reporting. Thus, even in our sample from a population strongly influenced by ideas concerning gender equality, a potentially manipulative sex-specific bias in resemblance reporting can not be excluded. This behavior might affect resource distributions among caregivers, for example, if grandparental care was a limited resource, conflicts between siblings in need of such resources may have been instigating the evolution of sexual differences in tactics between sons and daughters.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fisktjønmo, G.L.H., Bårdsen, BJ. & Folstad, I. Resemblance Reporting on Children: Sisters Are More Proactive than Brothers. Evolutionary Psychological Science 9, 1–12 (2023). https://doi.org/10.1007/s40806-022-00322-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40806-022-00322-3