
Abstract
Introduction
Juvenile dermatomyositis (JDM) is characterized by persistent non-purulent inflammation in the muscle and skin. The underlying mechanisms still remain uncertain. This study aims to elucidate the mechanism of interleukin-6 (IL-6) activation of Janus kinase/signal transducer and activator of transcription 3 pathway (JAK/STAT3), contributing to the pathogenesis of JDM.
Methods
Serum IL-6 levels were compared between 72 newly diagnosed patients with JDM and the same patient cohort in treatment remission. Single-cell RNA sequencing (scRNA-seq) was employed to identify differential signaling pathway expression in muscle biopsy samples from two patients with JDM and healthy controls. Immunohistochemistry was used to examine differences in STAT3 phosphorylation between JDM and control muscle tissues. In vitro, skeletal muscle cell lines were stimulated with IL-6, and the transcription levels of genes related to mitochondrial calcium channels were quantified via reverse transcription-polymerase chain reaction (RT-PCR). Reactive oxygen species (ROS) production was measured in both IL-6 treated and untreated groups. ROS levels were then compared between IL-6 receptor antagonist pre-treated skeletal muscle cells and untreated cells.
Results
IL-6 levels in newly onset patients with JDM were significantly higher compared to the same patient cohort in remission states (p < 0.0001). Serum IL-6 was significantly increased in patients with negative myositis specific antibody (MSA), positive melanoma differentiation associated protein 5 (MDA5) and positive nuclear matrix protein 2 (NXP2), yet not for JDM with positive transcriptional intermediary factor γ (TIF1γ), based on subgroup analysis. ScRNA-seq analysis of muscle biopsies from patients with MDA5-positive JDM and patients with MSA negative JDM revealed abnormal activation of the JAK/STAT3 pathway in skeletal myocytes, macrophages, and vascular endothelial cells. The phosphorylation levels of STAT3 were elevated in active JDM cases. Transcription of the calcium channel modulation gene sarcolipin (SLN) was significantly higher in JDM primary skeletal muscle cells compared to normal cells. In vitro, IL-6 enhanced SLN transcription and induced ROS production, and blocking the IL-6 receptor resulted in decreased ROS generation in skeletal muscle cells.
Conclusions
IL-6/JAK/STAT3 signaling pathway was abnormally activated in patients with JDM. IL-6 may be involved in the pathogenesis of muscle damage by triggering the development of calcium overload and production of ROS. Blockade of the IL-6/JAK/STAT3 pathway can be a potential treatment option for JDM, especially MDA5-positive patients and those who are negative for MSA.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Why carry out this study? |
The detailed mechanism of muscle damage in juvenile dermatomyositis (JDM) remains largely unknown. Discrepancy in the treatment outcome indicates more than one major signaling pathway may be involved in the development of the disease. |
The serum interleukin-6 (IL-6) level is increased in patients with JDM. We propose that IL-6 may be involved in the muscle damage of JDM. |
What was learned from the study? |
Results from single-cell RNA sequencing (scRNA-seq) analysis of muscle biopsies, immunohistopathological study, and in vitro cellular experiments revealed abnormal activation of the Janus kinase/signal transducer and activator of transcription 3 (JAK/STAT3) pathway, which may lead to the onset of calcium overload and production of reactive oxygen species (ROS) in JDM. |
Due to a small sample size for scRNA-seq analysis and the lack of the muscle activity study, further studies including expanded sample study, muscle activity assessment in the patients with JDM to IL-6 level, and subgroup analysis of JDM are needed to confirm the profound mechanism of IL-6 on the pathogenesis of JDM. |
Introduction
Juvenile dermatomyositis (JDM) is a rare chronic connective tissue disease in children, which is mainly characterized by inflammation of both the muscles and skin. Classical manifestations of JDM are typical rashes (such as heliotrope rash, Gottron’s papules), proximal muscle weakness and multiple organ systems involvement which may include gastrointestinal tract, lungs, heart and rarely kidneys [1]. Notably, despite the advances in therapeutic strategies utilizing traditional agents like corticosteroids (CS) and immunosuppressive agents, JDM carries a disconcerting prognosis. Approximately 20% of afflicted patients progress to refractory stages, characterized by resistance to these standard interventions [2].
Central to the clinical phenotype of JDM is the debilitating muscle weakness, a direct offshoot of the non-suppurative inflammation afflicting striated muscle tissues. This inflammation is predominantly characterized by lymphocyte infiltration, which is frequently seen in tandem with elevated creatine kinase (CK) levels, a molecular signature of muscle injury. However, the detailed mechanism of muscle damage in JDM still remains largely unknown.
Research over the past decade has shed light on an interferon (IFN) signature that appears to be foundational to the disease molecular landscape, findings which resonate with both adult patients with dermatomyositis (DM) and patients with JDM studies [3,4,5]. Notably, in muscle biopsy specimens, there is an observable link between the presence of pro-inflammatory T and B cells and the upregulation of IFN-driven myxovirus resistance protein A (MxA). Moreover, tissue-resident macrophages in these biopsies showcase heightened IFN-γ expression, producing a diverse array of pro-inflammatory molecules, including IL-6 [6]. However, the molecular mechanism for the IFN pathway and the subsequent cascade of events leading to immune cell dysregulation in JDM remain shrouded in mystery. The evident heterogeneous responses to treatment among patients with JDM further reinforce the likelihood of other signaling cascades playing pivotal roles in the disease's progression.
An intriguing aspect that has been thrust into the research spotlight is the role of mitochondria. Beyond being the cellular powerhouses responsible for ATP synthesis, mitochondria become critical hubs for ROS production when under the duress of oxidative stress or inflammation. Recent discoveries hint at a plausible nexus between mitochondrial dysfunction and the muscle weakness intrinsic to JDM. This is further cemented by reports highlighting the detection of anti-mitochondrial autoantibodies in the sera of adult patients with DM [7]. However, the mechanism between inflammation of muscle and mitochondria’s ROS abnormal generation in JDM remains to be fully deciphered. For these reasons, we meticulously probed the gene expression dynamics in JDM muscle tissues, contrasting them with healthy controls. Our finding provided insight into cellular heterogeneity intrinsic to JDM and spotlighted the paramountcy of the IL-6/JAK/STAT3 signaling pathway in JDM-associated muscle pathology.
Methods
Human Skeletal Muscle Cells (HSMCs), Isolation and Culture
Skeletal muscle samples were obtained from two JDM donors during muscle biopsy. The control muscle samples were obtained from patients with bone tumors. The discarded paratumoral muscle tissues were used as subsequent experiments (control HSMCs isolation and immunohistochemical staining). Primary human skeletal muscle was isolated and expanded by tissue explant adherent method. After washing these samples with PBS, non-muscle tissues were removed and the samples were sectioned into 1-mm3 columnar tissue blocks. Utilizing ophthalmic forceps, these blocks were methodically spaced at 0.5-cm intervals in a culture flask (NEST, Jiangsu, China) and covered with a specific growth medium blend. This medium, composed of high-glucose Dulbecco’s modified Eagle’s medium (DMEM) (GIBCO, USA), 20% fetal bovine serum (ExCell Bio, Jiangsu, China), and 2% Penicillin–Streptomycin Solution (beyotime, Shanghai, China), nourished the cells incubated at 37 °C in a 5% CO2 environment. The growth medium was replenished as necessary. For long-term storage, HSMCs were cryopreserved in a 90% growth medium and 10% dimethylsulfoxide mixture. Generally, cultured cells were maintained in DMEM enhanced with 10% fetal bovine serum and 1 × penicillin/streptomycin at standard conditions. Only cells from passages 1–3 were used for experiments.
Preparation for Single-Cell RNA Sequencing
To initiate the single-cell RNA sequencing, cells were first prepared by passing them through a 40-µm cell strainer to ensure a uniform single-cell suspension. Each cell suspension, 10 × barcode gel magnetic beads, and an oil mix were meticulously pipetted into the chambers of the chromium chip G to produce Gel Beads-in-emulsion (GEM). Each GEM was then subjected to a reverse transcription, performed at 53 °C for 45 min in a specialized PCR instrument (Thermo Fisher Scientific, Waltham, MA, USA). Subsequent complementary deoxyribonucleic acid (cDNA) purification and amplification ensured the quality and quantity of starting material. Enzymatic fragmentation was adjusted to yield fragments primarily between 200 and 300 base pairs. These fragments then underwent magnetic bead selection, end-repair, A-tailing, and Read2 sequencing primer ligation. Adapter-ligated cDNA was finally indexed, and the library was constructed. Prior to sequencing, libraries were quantified using the Qubit 2.0 Fluorometer and validated for size distribution using the Agilent 2100 Bioanalyzer. The validated libraries were then sequenced on the NovaSeq6000 (Illumina, San Diego, CA, USA) platform with a 150-bp paired-end configuration.
10× Genomics single-nucleus RNA-Seq (snRNA-seq) experiment
The 10× Genomics library preparation and snRNA-seq were carried out by NovelBio Co., Ltd (Shanghai, China). The process employed the 10× Genomics Chromium Controller Instrument and the Chromium Single-Cell 3′ V3 Reagent Kits, sourced from 10× Genomics (Pleasanton, CA, USA). Initially, the nuclei were washed twice using PBS. Subsequently, they were left to incubate for 30 min at room temperature. With precision, approximately 1000 nuclei/μl were then loaded onto the 10× Chromium chips. These chips were pre-loaded with the 10× single-cell 3′ V3 chemistry and barcode, facilitating the creation of single-cell Gel Bead-In-Emulsions (GEMs). Post-GEMs creation, the synthesis of cDNA commenced. Once this step was completed, the GEMs were broken to release the barcoded cDNA. This cDNA then underwent an amplification process consisting of 14 cycles post library construction. The process continued with fragmentation and index PCR amplification. The resulting final libraries were then assessed quantitatively using the Qubit High Sensitivity DNA assay (Thermo Fisher Scientific, Waltham, MA, USA). Additionally, the size distribution of these libraries was ascertained with the assistance of a High Sensitivity DNA chip, utilized on a Bioanalyzer 2200 (Agilent Technologies, Santa Clara, CA, USA). The entire sequencing process was undertaken on the Novaseq. 6000 platform (Illumina, San Diego, CA, USA), deploying a 2 × 150-bp paired-end sequencing protocol.
Preprocessing and quality control of the snRNA-seq data
In our endeavor to quantify both exonic and intronic reads captured via snRNA-seq, we constructed a tailored "pre-mRNA" reference package. This was built upon the human reference genome dataset, specifically the version refdata-gex-GRCh38-2020-A, in alignment with the 10× Genomics protocol. The raw data from the sequencing was matched to this "pre-mRNA" reference employing the Cell Ranger toolkit (version v4.0.0). For the ensuing data preprocessing steps, the resulting nucleus-gene expression matrix was assimilated into the "Seurat" software package, version v3.2.3. We applied strict criteria during the preprocessing phase. Genes that manifested counts in less than three nuclei were pruned from the dataset. This filtration process was vital in nullifying the genes that might have been detected due to potential random noise. The quality control of the nuclei was paramount. They were subjected to a rigorous screening process based on unique molecular identifier (UMI) counts, ensuring a range between 500 and 50,000. Similarly, the gene count range was established between 300 and 7000. Any nuclei that surpassed a 0.05 proportion in mitochondrial genes or ribosomal genes were disregarded. These boundaries were essential to eliminate inferior-quality nuclei, which might have been indicative of doublets or perturbations from technical discrepancies. An additional layer of quality control was the exclusion of nuclei with doublet scores exceeding 0.35, as predicted by the Scrublet tool. Furthermore, any nuclei exhibiting enhanced expression across marker genes spanning multiple lineages were meticulously omitted to ensure data integrity in subsequent analyses.
Data Download and Processing
For our analysis, we sourced muscle cell datasets from healthy individuals as a comparative benchmark. These datasets were procured from the renowned Gene Expression Omnibus (GEO) repository, which can be accessed at https://www.ncbi.nlm.nih.gov/geo/. The control muscle tissues used in genetic analysis were distinct from the JDM samples in terms of its origin and processing. The healthy control individuals were adults aged 18–50 years who were recruited through advertisements at the university and local community based on information provided in the original text [8]. These samples were notably from individuals without any diagnosed muscular anomalies. Specifically, we utilized data under the accession numbers GSM6611295, GSM6611297, and GSM6611299. Notably, all three samples pertained to muscle tissue and were collected prior to any form of exercise. In our endeavor to maintain uniformity in our analysis, the filtering criteria employed for the snRNA-seq data of JDM were replicated for this control group. It is worth mentioning that the JDM samples originated in our laboratory, derived from biopsy specimens of patients diagnosed with the condition. For a comprehensive analysis, we integrated the JDM sample data with the snRNA-seq data of the aforementioned control samples. Given the multifaceted nature of the combined data, it was imperative to address and rectify potential batch effects. To achieve this, we harnessed the capabilities of the "harmony" package. This package boasts an algorithm specifically tailored to streamline and align intricate multi-sample datasets, ensuring uniformity and coherence across the board.
Normalization, Feature Selection, Integration, Scaling, and Clustering of the SnRNA-Seq Data
For each sample, the sum of the UMI counts for each nucleus was normalized to 10,000 followed by log-transformation. We used the FindVariableFeatures function of Seurat to select 2000 highly variable genes for each sample based on mean variability plot. Nuclei of all samples were integrated via canonical correlation analysis implemented in Seurat, which was set to identify the top 30 canonical correlations to correct for potential batch effects and identify shared cell states across samples. Unwanted sources of variation were mitigated by regressing out specific factors: the proportion of mitochondrial genes, UMI count, gene number, the proportion of mitochondrial genes, and the proportion of ribosomal genes with linear models using ScaleData. The data were then centered for each gene by subtracting the mean expression and scaled by dividing the centered expression by the standard deviation. Principal component analysis (PCA) was applied and the first 20 PCA components were used to compute a shared nearest-neighbor (SNN) graph of the nuclei. The SNN graph was visualized using uniform manifold approximation and projection (UMAP). Clustering was performed using the Louvain algorithm with the clustering parameter resolution set to 0.5, specifically to identify the subpopulation structures within the samples.
Gene Set Variation Analysis (GSVA)
Cell types were analyzed for genomic variation using the GSVA software package, with the objective to explore and interpret the biological pathways activated in each cell type. Gene sets for GSVA were obtained from the MSigDB database (c2. Cp. Kegg. V7.1. Symbols. gmt), including more than 5000 gene sets that cover various biological processes and pathways. A GSVA score was generated for each cell using the "GSVA" software package, with kcdf set to Poisson, reflecting the underlying distribution of the gene expression data. The “complexHeatmap” and “ggplot2”software packages were used to visualize the activated pathways, providing an interactive and informative view of the data.
Detection of Serum Helper T cell (Th) 1/Th2 Cytokine Levels by Flow Cytometry
The serum levels of interleukin (IL) -2, IL-4, IL-6, IL-10, tumor necrosis factor-α (TNF-α), and IFN-γ from patients with JDM (pre and post treatment) were determined by using the Cytometric Bead Array Human Th1/Th2 Cytokine Kit II, purchased from BD Pharmingen (San Jose, CA) (cat. no. 551809). Cytokine titers were expressed as pg/ml, as calculated by reference to standard curves based on known amounts of recombinant cytokines.
The diagnosis of JDM was based on the 2017 EULAR/ACR classification criteria [9] and disease remission should fulfil the Pediatric Rheumatology International Trials Organization (PRINTO) criteria for clinically inactive disease in JDM, meet at least three out of four of the following criteria on or off therapy: (i) physician global assessment of overall disease activity (PhyGloVAS) ≤ 0.2, (ii) creatine kinase (CK) ≤ 150 U/l, (iii) CMAS score ≥ 48, (iiii) MMT-8 score ≥ 78 [10]. Patients with complete cytokine information on pre-treatment and remission state were included in this study.
Immunohistochemical (IHC) Staining
IHC was applied to assess the expression of p-STAT3 in muscle tissues from both JDM and controls. Antibody specific to p-STAT3 was purchased from Cell Signaling Technology (Beverly, MA, USA), using a working dilution as 1:500. The slides were scanned by a Pannoramic MIDI scanner (3DHISTECH, Hungary) and viewed using CaseViewer software v2.2.
ROS Measurement
Primarily isolated human skeletal muscle cells from JDM and HSMC cell line were stimulated with IL-6 (25 ng/ml, Novoprotein, Jiangsu, China; suspended in PBS) or PBS (served as control) for 24 h and cells were then collected into tubes. Cells were stained with DCFH-DA (1 mM in 1 × PBS) for 30 min at 37 °C. ROS analysis of HSMCs was completed by using Reactive Oxygen Species Assay Kit (beyotime, S0033S, Shanghai, China) by a microplate reader (TECAN, Switzerland). HSMC treated with humanized recombinant anti-IL-6R antibody (Tocilizumab, ACTEMRA®, Roche, working concentration 100 ng/ml) was listed as intervention group. The concentration of IL-6 and tocilizumab was set based on previous studies and our pre-experiments performed by different concentration gradients [11, 12]. We decided on a final IL-6 stimulation concentration of 25 ng/ml, and tocilizumab treatment concentration of 100 ng/ml.
Quantitative Reverse Transcription PCR
Total RNA was extracted after HSMC (from JDM and normal cell line as control) were stimulated with IL-6 (25 ng/ml) using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). RNA was reverse transcribed into cDNA using the PrimeScript RT Master Mix (Takara, Shiga, Japan) according to the manufacturer’s protocol. Quantitative PCR was performed on the CFX Opus 96 Real-Time PCR System (Bio-Rad, Hercules, CA, USA) using SYBR Green reagents (Monad Biotech, Jiangsu, China); the corresponding primers are listed in Supplementary Table 1. The expression of target genes was calculated using the 2−ΔΔt method after normalizing to the housekeeping gene GAPDH and control group.
Western Blot
The protein extracted from both JDM and control muscle tissues as well as HSMCs (isolated from JDM or normal cell line) stimulated with IL-6 (25 ng/ml) were degenerated, electrophoresed, and transferred to the polyvinylidene fluoride membrane (Bioss, Beijing, China). Subsequently, the membrane was blocked using 5% non-fat milk and incubated with primary antibodies-rabbit stat3 (1:1000, ab68153, Abcam) and p-stat3 (1:1000, ab32143, Abcam) and mouse anti-β-actin (Bioss, Beijing, China) overnight at 4 °C. On the following day, the membranes were washed and incubated with the corresponding horseradish peroxidase-conjugated secondary antibodies (Bioss, Beijing, China) and detected with an enhanced chemiluminescence reagent (Bioss, Beijing, China) using the chemiluminescence system (GBOX-CHEMI-XRQ, Syngene, USA). The grayscale of the detected bands were assessed by ImageJ software (National Institutes of Health), and the results were expressed as fold changes after normalizing to β-actin.
Statistical Analysis
All statistical analyses were performed using GraphPad Prism 9 (GraphPad Software Inc., La Jolla, CA, USA). Continuous data were expressed as mean (standard deviation) and were tested by Student’s t test. Non-normal continuous data were expressed as median (interquartile ranges) and were tested by Wilcoxon test. Levels of IL-6 were compared using paired t test. The effect values and p values generated in these tests were used to map the volcano, which was visualized using the ggplot2 package 3.2.1 in R. A two-tailed t test 0.05 was used to test each term for significance. p values were adjusted for multiple testing using the “Benjamini–Hochberg” method or the “Holm” methods.
Ethical Approval
Informed consent was obtained from guardians as well as subjects aged over 8 years. The study was approved by institutional research ethics committee of Zhejiang University School of Medicine (2021-IRBAL-303).
Results
IL-6 and IL-10 were Significantly Increased in Treatment-Naïve Patients with JDM
IL-6 is a key upstream activator of JAK/STAT3 signal transcription pathway. Serum cytokine levels were detected from 72 newly diagnosed JDM and the same cohort of patients who were in a clinical remission state. The flow chart of patient enrollment in this study was shown in Supplementary Fig. 1. Characteristics of the study population and changes in metrics pre- and post-treatment in patients with JDM were displayed in Supplementary Table 2. The result showed that treatment-naïve patients with JDM [median 13.0, IQR (1.3, 66.4) pg/ml] had much higher IL-6 than patients in clinical remission state [median 3.7, IQR (1.1, 16.4) pg/ml] (R = 0.32, p < 0.0001). Patients with JDM were then divided into five subgroups: MSA negative JDM (n = 30), MDA5+ JDM (n = 12), NXP2+ JDM (n = 9), TIF1γ+ (n = 8) and other (any other MSA-positive JDM, n = 13). Subgroup analysis revealed that both MSA negative and MDA5+ patients had increased level of IL-6 and IL-10. However, there was no significant difference in IL-6 level for TIF1γ+ patients before and after treatment. Serum IL-6 was also found elevated in NXP2+ JDM and patients who had other MSA positive (Fig. 1).

Comparison of cytokine levels of patients with juvenile dermatomyositis in different clinical states. Helper T cells (Th) 1/Th2 cytokines (IL-2, IL-4, IL-6, IL-10, TNF-α, and IFN-γ) were tested in treatment-naïve patients with juvenile dermatomyositis (JDM) and the same cohort patients who met the inactive disease criteria after 12 months of treatment. IL-6 and IL-10 were found significantly increased in treatment-naïve patients. Analysis of subgroups indicated that the majority of patients with JDM, with the exception of those with anti-transcription intermediary factor 1-gama autoantibodies-positive exhibited increased levels of IL-6. IL-2 interleukin-2, IL-4 interleukin-4, IL-6 interleukin-6, IL-10 interleukin-10, TNF-α tumor necrosis factor-alpha, IFN-γ interferon-gamma, MSA myositis specific antibody, MDA5 melanoma differentiation associated protein 5, NXP2 nuclear matrix protein 2, TIF1γ transcriptional intermediary factor γ
IL-6 Enhances Transcription of Mitochondrial Calcium Overload-Related Genes In Vitro
Genes associated with mitochondrial calcium overload include lactate dehydrogenase A (LDHA), lactate dehydrogenase B (LDHB), mitochondrial calcium uniporter (MICU) 1, MICU2, SLN, phospholamban (PLN), phosphoglucomutase (PGM), cytochrome b5 reductase 1 (CYB5R1), Aldolase A (ALDOA) and mitochondrial Ca2+ uniporter (MCU). Through single-cell transcriptome analysis, we found a significant elevation in the expression of the calcium channel regulatory genes in JDM myoblasts compared to the control (average logFC = 3.6, p value < 0.001) (see Supplementary Fig. 2). The gene result was further verified by RT-PCR, to quantify the transcription levels of above genes. Sarcolipin (SLN) is a regulator of sarcoplasmic/endoplasmic reticulum Ca2+ transport ATPase. As a calcium channel regulatory gene, SLN displayed a markedly higher expression in JDM skeletal muscle cells compared to their normal counterparts (Fig. 2). Interestingly, in vitro study showed that IL-6 only boosted the transcription of SLN in inflammatory muscle cells, but not exhibit the same influence on the standard skeletal cell line.

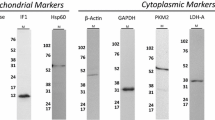
Interleukin-6 upregulates the transcription of mitochondrial calcium overload-related genes. The expression of calcium channel regulatory genes including PGM, CYB5R1, SLN and ALDOA were increased significantly in primary skeletal muscle cells from JDM (e, f, g, j) compared with normal cell lines. IL-6 upregulates the expression of above genes in vitro, especially in primary JDM skeletal muscle cells (e–g). Interestingly, IL-6 only boosted the transcription of SLN in inflammatory muscle cells, but not exhibited the same influence on the standard skeletal cell line (g). LDHA lactate dehydrogenase A, LDHB lactate dehydrogenase B, MICU1 mitochondrial calcium uniporter 1, MICU2 mitochondrial calcium uniporter 2, PGM phosphoglucomutase, CYB5R1 cytochrome b5 reductase 1, SLN sarcolipin, NCLX Na+/Ca2+ exchanger, ALDOA aldolase A, IL-6 interleukin-6, JDM juvenile dermatomyositis
Integrated scRNA-seq Analysis of Cellular Diversity in JDM
In our quest to discern gene expression alterations and to construct an intricate map of JDM muscle tissue at the cellular level, we subjected two JDM samples and three normal muscle tissue controls to scRNA-seq. An integrated bioinformatics method was employed in our analysis. With thorough quality control and clustering measures in place, our focus shifted to 44,300 cells for a more in-depth analysis. This meticulous process incorporated batch effect adjustments, downscaling, and graph-based clustering, facilitating the identification of 21 distinct clusters spanning five principal cell lineages. These identified clusters were categorized as follows: endothelial cells (VWF), fibroblasts (DCN), macrophages (CD163, LYN), myocytes (TNN), neuronal cells (CTNAP2), NK cells (NKG7, GZMA, GZMB), pericytes (TAGLN, ACTA2), and satellite cells (PAX7). Using a dot plot, we showcased the top five marker genes for each category (Fig. 3A, C). Also, our findings highlighted heterogeneity between JDM and healthy control, particularly in the cellular composition and gene expression profiles of fibroblasts, endothelial cells, and myocytes. Our result demonstrated significant differences between JDM and control samples: an increase in macrophages and myocytes, conversely a decrease in fibroblasts and endothelial cells in JDM, which may indicate potential vascular dysfunction and less fibrotic remodeling in JDM. Despite these changes, the proportion of satellite cells remains consistent (Fig. 3B). This cellular profiling highlights the complex pathological interplay in JDM.

Integrated single-cell RNA sequencing analysis of cellular diversity and gene set variation in juvenile dermatomyositis. Single-cell RNA sequencing analysis of muscle biopsies from two patients with juvenile dermatomyositis (control data were from references). Cell clusters (A), cell composition (B), and corresponding gene markers (C) implied in analysis
Gene Set Variation Analysis in Cell Subpopulations Between JDM and Control
Incorporating the GSVA on our integrated data, we identified enrichment in 225 distinct pathways. Among these, 39 were found to be downregulated in JDM compared to the controls (logFC > 0 and p value < 0.05) while 176 were upregulated (logFC < 0 and p value < 0.05) irrespective of the cell type. A breakdown of the mean GSVA score per cell type pinpointed the top 15 pathways boasting the highest mean scores. We also juxtaposed the GSVA scores of the JAK-STAT signaling pathway between the JDM and control groups across all cell types. Strikingly, JDM consistently recorded a higher GSVA score compared to the control across all cell subpopulations, a variance that was statistically significant (p < 0.001) (Fig. 4).

Comparison of JAK/STAT signaling pathway between the juvenile dermatomyositis and control group. In total, 225 distinct pathways were noticed in juvenile dermatomyositis compared to the control. A The top 15 pathways boasting the highest mean scores. Janus kinase/signal transducer and activator of transcription signaling pathway was found more active in juvenile dermatomyositis than the controls across all cell subpopulations (B, C). NK natural killer, JDM juvenile dermatomyositis, JAK/STAT Janus kinase/signal transducer and activator of transcription, STAT3 signal transducer and activator of transcription 3
IL-6 Induced Over-Expression of pSTAT3 and ROS in Skeletal Muscle Cells
Compared with controls (muscle tissues from patients with bone tumors), patients with JDM had significantly increased expression of pSTAT3, especially in intermuscular and perivascular areas (Fig. 5d). Results of Western blot convinced over-expression of STAT3 and pSTAT3 from primary isolated HSMC from JDM patients (Fig. 5b). Simultaneously, IL-6 enhanced phosphorylation of STAT3 in the muscular cell model, which could be blocked by STAT3 inhibitor (S31-201) (Fig. 5c).

Interleukin-6 induced over-expression of phospho-signal transducer and activator of transcription 3 and reactive oxygen species in skeletal muscle cells. a Interleukin-6 stimulus induces ROS production in vitro. Blockade of interleukin-6 receptor decreases the ROS production in primary human skeletal muscle cells. ROS tests were repeated twice for each donor specimens (two patients and two controls) and each experiment set triplicate wells. b, c Human skeletal muscle cells derived from patients with JDM exhibited higher STAT3 and pSTAT3 expression than normal cell line in vitro. Interleukin-6 stimulus enhanced the expression of pSTAT3 in both control cells and JDM skeletal muscle cells, while STAT3 inhibitor (S3I-201) significantly reversed it. The data represented the means ± standard deviation from three independent experiments. d Immunohistochemistry analysis of muscle biopsy from JDM displayed increased expression of pSTAT3 [Ia and Ib represented JDM muscle biopsy; IIa and IIb were from control samples, scale bars, 100 µm (Ia, IIa), 50 µm (Ib, IIb)]. ROS reactive oxygen species, JDM juvenile dermatomyositis, STAT3 signal transducer and activator of transcription 3, pSTAT3 phospho-signal transducer and activator of transcription 3, TCZ tocilizumab
The IL-6/STAT3 pathway has been theorized to play a pivotal role in the muscle weakness observed in patients with JDM. Upon co-culturing HSMCs with IL-6 (25 ng/ml) for a span of 24 h, with or without anti-IL-6R antibody (tocilizumab, Roche), it was determined that IL-6 treated HSMCs had increased ROS production comparing with untreated cells. While blocking IL-6R significantly curtailed ROS production in skeletal muscle cells (Fig. 5a).
Discussion
Biologics such as rituximab, abatacept, tocilizumab and TNF-ɑ antagonists have been used off-label for refractory DM in these two decades [13]. While most of these studies had limited sample sizes, and their effectiveness has yet to be verified. Tocilizumab, an anti-IL-6R antibody, was shown to be effective in some refractory rapidly progressive interstitial lung disease (ILD) related to anti-MDA5-positive DM and immune-mediated necrotizing myopathies in adults [14, 15]. However, current research still lacks depth on the mechanism of IL-6 in disease development, and reports on the effectiveness of anti-IL-6R antibody in JDM are scarce.
IL-6 binds to membrane-bound receptor or soluble IL-6R, activating STAT3 and triggering the phosphorylation of STAT3. Dimeric pSTAT3 or heterodimer with STAT1 transferred to the nucleus and activated subsequent transcription factor to regulate cell function [16]. A case-control study displayed that IL-6 174G/C polymorphism might be associated with muscle weakness and elevated muscle enzymes in patients with DM [17]. It was reported that serum IL-6 was a poor prognostic factor of clinically amyopathic dermatomyositis complicating with rapidly progressive ILD, however, multiple stepwise regression analysis failed to confirm this result [18]. In our study, comparing treatment-naïve patients with the same cohort patients in clinical remission, the serum level of IL-6 was significantly higher in active patients, reinforcing the notion that IL-6 might play a pathogenic role in JDM.
Skeletal muscle cells from patients with JDM showed increased transcription of SLN, (a regulator of sarcoplasmic/endoplasmic reticulum Ca2+ transport ATPase) compared to normal cells. When exposed to IL-6, JDM muscle cells exhibited heightened SLN transcription in vitro, a phenomenon absent in normal myocytes. SLN modulates ATP hydrolysis and Ca2+ cycling within cells. Its overproduction might lead to increased energy consumption in muscles, correlating with muscle dystrophy in various diseases [19]. Our findings indicated that inflamed muscle cells, might be more sensitive to inflammatory cytokines like IL-6, potentially boosting SLN production, causing energy and calcium imbalances in the mitochondria.
Amidst these findings, our integrated scRNA-seq analysis has offered unprecedented insights, illuminating distinct cellular states and intricate molecular signatures associated with JDM, highlighting potential pathways and targets for future therapeutic interventions. The IL-6/STAT3 signaling pathway has garnered significant attention in recent studies, being implicated in various conditions like hepatic dysfunction, oncogenesis, interstitial lung disease, and fibrosis, among others [20, 21]. What is interesting is IL-6 on the one hand can activate STAT3 signaling [22], while on the other hand, STAT3 signaling pathway can transcribe a number of cytokine genes which includes IL-6 [23]. In our study, immunohistochemical analysis of skeletal muscle tissues revealed heightened pSTAT3 expression in patients with JDM compared to controls, emphasizing that STAT3 signaling pathway may involve in the pathogenesis of muscle damage.
ROS was proposed as an upstream modulator of IL-6/STAT3 signaling in some inflammatory diseases [20]. In a mice model, acute exercise increased blood IL-6 level, while deficiency of ROS could impair exercise induced inflammation [24]. IL-6 knockout mice exhibited reduced skeletal muscle atrophy and a simultaneous decline in ROS production within these muscles [25]. Our findings demonstrated increased ROS production in muscle cells from patients with JDM compared to normal controls. Furthermore, in vitro studies revealed that IL-6 could trigger ROS production even in normal myocytes. Given these observations, we postulate that IL-6 might play a pivotal pathogenic role by stimulating ROS production via the JAK-STAT3 signaling pathway, creating a positive feedback loop in inflamed muscle cells, culminating in irreversible tissue damage. To ascertain the impact of IL-6/STAT3 inhibitors in patients with JDM, further studies on a larger patient cohort, alongside an in-depth exploration of the IL-6 pathway in JDM pathogenesis are paramount. Noteworthy, IL-6 was not elevated in all patients with JDM, especially in the group of TIF1-γ.
There are shortcomings of the present study such as limit sample size for scRNA-seq analysis of muscle tissue, and lack of study on cytokines and muscle activity. More in-depth research is needed to develop clinical or serum biomarkers that correlate with muscle activity to better study the relationship between cytokines and muscle damage and provide more accurate or effective treatment for those patients.
Conclusions
Our study investigated the activation of IL-6/STAT3 signaling in the muscle of JDM by scRNA-seq and in vitro cellular experiments. The results suggested that the IL-6/JAK/STAT3 signaling pathway was abnormally activated in the muscle of patients with JDM. Moreover, serum IL-6 was significantly increased in JDM with negative MSA, positive MDA5 and positive NXP2, yet not for JDM with positive TIF1γ. The limitation of this study was the small sample for scRNA-seq analysis and lack of the muscle activity study. Therefore, further studies including expand sample study, muscle activity assessment in JDM to IL-6 level, and subgroup analysis of JDM are needed to confirm the profound mechanism of IL-6 on the pathogenesis of JDM.
Data Availability
All data generated or analyzed during this study are included in this published article and its supplementary information files. Human JDM single-cell RNA-seq data is available at the GEO repository (accession number PRJNA1036392).
References
Papadopoulou C, Chew C, Wilkinson MGL, McCann L, Wedderburn LR. Juvenile idiopathic inflammatory myositis: an update on pathophysiology and clinical care. Nat Rev Rheumatol. 2023;19:343–62.
Ruperto N, Pistorio A, Oliveira S, et al. Prednisone versus prednisone plus ciclosporin versus prednisone plus methotrexate in new-onset juvenile dermatomyositis: a randomised trial. Lancet (London, England). 2016;387:671–8.
Kim H, Gunter-Rahman F, McGrath JA, et al. Expression of interferon-regulated genes in juvenile dermatomyositis versus mendelian autoinflammatory interferonopathies. Arthritis Res Ther. 2020;22:69.
Moneta GM, Pires Marafon D, Marasco E, et al. Muscle expression of type i and type ii interferons is increased in juvenile dermatomyositis and related to clinical and histologic features. Arthritis Rheumatol (Hoboken, NJ). 2019;71:1011–21.
Gitiaux C, Latroche C, Weiss-Gayet M, et al. Myogenic progenitor cells exhibit type i interferon-driven proangiogenic properties and molecular signature during juvenile dermatomyositis. Arthritis Rheumatol (Hoboken, NJ). 2018;70:134–45.
Nistala K, Varsani H, Wittkowski H, et al. Myeloid related protein induces muscle derived inflammatory mediators in juvenile dermatomyositis. Arthritis Res Ther. 2013;15:R131.
Duvvuri B, Pachman LM, Morgan G, et al. Neutrophil extracellular traps in tissue and periphery in juvenile dermatomyositis. Arthritis Rheumatol (Hoboken, NJ). 2020;72:348–58.
Lovrić A, Rassolie A, Alam S, et al. Single-cell sequencing deconvolutes cellular responses to exercise in human skeletal muscle. Commun Biol. 2022;5:1121.
Lundberg IE, Tjärnlund A, Bottai M, et al. 2017 European league against rheumatism/American college of rheumatology classification criteria for adult and juvenile idiopathic inflammatory myopathies and their major subgroups. Arthritis Rheumatol (Hoboken, NJ). 2017;69:2271–82.
Lazarevic D, Pistorio A, Palmisani E, et al. The PRINTO criteria for clinically inactive disease in juvenile dermatomyositis. Ann Rheum Dis. 2013;72:686–93.
Kudo M, Jono H, Shinriki S, et al. Antitumor effect of humanized anti-interleukin-6 receptor antibody (tocilizumab) on glioma cell proliferation. Laboratory investigation. J Neurosurg. 2009;111:219–25.
Han F, Li S, Yang Y, Bai Z. Interleukin-6 promotes ferroptosis in bronchial epithelial cells by inducing reactive oxygen species-dependent lipid peroxidation and disrupting iron homeostasis. Bioengineered. 2021;12:5279–88.
Spencer CH, Rouster-Stevens K, Gewanter H, et al. Biologic therapies for refractory juvenile dermatomyositis: five years of experience of the childhood arthritis and rheumatology research alliance in North America. Pediatr Rheumatol Online J. 2017;15:50.
Li S, Li W, Jiang W, et al. The efficacy of tocilizumab in the treatment of patients with refractory immune-mediated necrotizing myopathies: an open-label pilot study. Front Pharmacol. 2021;12: 635654.
Zhang X, Zhou S, Wu C, et al. Tocilizumab for refractory rapidly progressive interstitial lung disease related to anti-MDA5-positive dermatomyositis. Rheumatology (Oxford). 2021;60:e227–8.
Calabrese LH, Rose-John S. Il-6 biology: implications for clinical targeting in rheumatic disease. Nat Rev Rheumatol. 2014;10:720–7.
Hristova M, Dourmishev L, Kamenarska Z, et al. Role of the promoter polymorphism il-6 -174g/c in dermatomyositis and systemic lupus erythematosus. Biomed Res Int. 2013;2013: 315365.
Nara M, Komatsuda A, Omokawa A, et al. Serum interleukin 6 levels as a useful prognostic predictor of clinically amyopathic dermatomyositis with rapidly progressive interstitial lung disease. Mod Rheumatol. 2014;24:633–6.
Pant M, Bal NC, Periasamy M. Sarcolipin: a key thermogenic and metabolic regulator in skeletal muscle. Trends Endocrinol Metab. 2016;27:881–92.
Öztürk Akcora B, Vassilios Gabriël A, Ortiz-Perez A, Bansal R. Pharmacological inhibition of stat3 pathway ameliorates acute liver injury in vivo via inactivation of inflammatory macrophages and hepatic stellate cells. FASEB Bioadv. 2020;2:77–89.
Braicu C, Buse M, Busuioc C, et al. A comprehensive review on MAPK: a promising therapeutic target in cancer. Cancers. 2019;11:1618.
Yao Y, Ye H, Qi Z, et al. B7–h4(b7x)-mediated cross-talk between glioma-initiating cells and macrophages via the il6/jak/stat3 pathway lead to poor prognosis in glioma patients. Clin Cancer Res. 2016;22:2778–90.
Jarnicki A, Putoczki T, Ernst M. Stat3: linking inflammation to epithelial cancer—more than a “gut” feeling? Cell Div. 2010;5:14.
de Barcellos LAM, Gonçalves WA, de Oliveira MPE, et al. Effect of physical training on exercise-induced inflammation and performance in mice. Front Cell Dev Biol. 2021;9: 625680.
Yang B, Yang X, Sun X, Shi J, Shen Y, Chen R. Il-6 deficiency attenuates skeletal muscle atrophy by inhibiting mitochondrial ROS production through the upregulation of pgc-1α in septic mice. Oxid Med Cell Longev. 2022;2022:9148246.
Acknowledgements
The authors thank all participating patients and volunteers for their support.
Funding
This work was supported by Key R&D Program of Zhejiang (2023C03032). The journal’s Rapid Service Fee was funded by the authors.
Author information
Authors and Affiliations
Contributions
Qi Zheng: Study design and manuscript writing. Zhaoling Wang: Lab work and statistical analysis. Yejun Tan: ScRNA-seq analysis. Kun Zhu: Immunohistochemical staining; Meiping Lu: Review and modification of the article.
Corresponding author
Ethics declarations
Conflict of Interest
Qi Zheng, Zhaoling Wang, Yejun Tan, Kun Zhu, and Meiping Lu announce that there is no conflict of interests among them.
Ethical Approval
Informed consent was obtained from guardians as well as subjects aged over 8 years. The study was approved by institutional research ethics committee of Zhejiang University School of Medicine (2021-IRBAL-303).
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License, which permits any non-commercial use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc/4.0/.
About this article

Cite this article
Zheng, Q., Wang, Z., Tan, Y. et al. Over Activation of IL-6/STAT3 Signaling Pathway in Juvenile Dermatomyositis. Rheumatol Ther (2024). https://doi.org/10.1007/s40744-024-00699-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s40744-024-00699-6