
Abstract
Rice (Oryza sativa L.) roots play important role in the absorption of water, nutrients, and also in stress tolerance such as desiccation, salt and drought stress. The root’s anatomical and histological features showed large xylem vessels and reduced aerenchyma formation in resistant genotypes under water stress. Correlation of anatomical changes in parenchymatous cortex region, starch accumulation in the cortex, and outer unicellular layer of epiblema was also found in rice var. Nagina 22, the most acclaimed drought-resistant variety in Asia. Additionally, electrophoresis of proteins extracted from the roots after water stress showed putative induction of 4–5 proteins putatively induced in var. Nagina 22. These findings suggest that a complex network of regulatory pathways of large xylem vessels with lesser aerenchyma formation and higher starch content in tolerant rice varieties are required for the maintenance of water potential and energy storage under water stress.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Production of rice, the world’s most important staple food crop [17], is threatened worldwide by drought [11]. Drought is a multidimensional stress that affects different plant growth stages in about 20 % of the total rice-growing area in Asia [29]. Understanding the mechanisms of drought resistance is an important issue for rice-breeding programs and development of drought-resistant varieties. Different phenotypic traits associated directly with drought tolerance are unclear; however, it has been observed that deep rooting [44] and osmotic adjustment [16] are associated with drought tolerance characteristics. Roots are the principal plant organ for nutrient and water uptake. Therefore, improving our understanding of the interactions between root function and drought in rice could have a significant impact on global food security and production [37]. Upland rice genotypes usually have deep and thick root systems, which allow the crop to satisfy its water requirements [27]. Varieties that have deep root systems are needed for acquiring water and nutrition from the relatively wet deep soil layer to obtain a stable yield under rainfed conditions [24]. The mechanism controlling internal plant water status may be involved in the water uptake or water conservation by the plant and also internal plant water conductance under drought. Fukai and Cooper [10] proposed that the genotypic variations in the root system in order to enhance the water capture ability of the plant could be an avenue for improving the drought tolerance of rice. Other root traits, such as seminal and crown root developments, which are effective for rice plants to grow under drought and/or upland conditions, have been widely reported [22, 38]. It is reported that anatomical features of root, like xylem diameter, are likely to be related to maintenance of conductivity [39] and increased xylem vessel size has been hypothesized to be a useful trait for improving water extraction from deeper soil layers [47]. Another important factor having a correlation to drought is differential accumulation of carbohydrates in various parts of the rice under stress conditions. During the stress, grain growth is largely dependent on reserved food material mainly accumulated carbohydrates before stress and the actual amount of its remobilization. For avoiding such a situation, rice also accumulates a significant amount of carbohydrates before heading, and wide variability exists among genotypes for these characters [46]. Improved rice cultivars have a higher content of total non-structural carbohydrates at heading than the low-yielding traditional varieties [8].
As a biochemical signal, protein synthesis widely responds to environmental stresses such as water stress, heat shock, and osmotic shock. Changes in proteins induced due to drought play a pivotal role in the adaptive response of plants [34]. The stress-induced proteins allow plants to make biochemical and structural adjustment that enables them to cope with stress. Proteins that show significant down-regulation under drought stress were observed for photosynthesis-related function [26]. Among the stress-induced proteins identified, are those implicated in the biosynthesis of osmolytes [14], in the uptake and compartmentation of ions [19], in hydroxyl-radical scavenging [13], and protection of cellular structure [20]. Water deficit induces the expression of proteins that are directly or indirectly related to stress and some functions have been assigned to some of the sequenced proteins. Proteomics is a recent addition to the molecular tools used to analyze drought-affected plants, and has been applied to the study of drought response of several crop plants including barley [26, 35] and maize [34]. In addition to the physiological responses of rice to water stress, the information on the biochemical and molecular mechanisms of drought adaptation could be useful for the genetic improvement of drought-tolerant varieties. Thus, understanding the biochemical basis of drought tolerance of rice root proteins may be helpful in developing new strategies for improving drought tolerance in rice. In line with earlier findings, drought stress initiated at different growth stages may induce quantitative and qualitative changes in proteins in different plant parts. Although numerous genes and proteins, which potentially contribute to drought tolerance in rice, have been reported [48], most of these studies have been focussed on lowland rice genotypes. Salekdeh et al. [36] revealed a large number of differences in root proteins between salt-tolerant and salt-sensitive varieties, including proteins with antioxidant properties and proteins involved in lignification. The comparison of gene expression and proteome profiles in roots of drought-tolerant and drought-sensitive upland rice varieties revealed the tolerant variety to have increased expression of genes or proteins involved in turgor, cell integrity, and oxidative stress [32]. Currently, very little is known about the gene(s) and their expression in upland rice [45]. Thus, information on the biology of root anatomy, histology, and protein patterns during drought in upland rice is limited. So the present investigation was taken up to study and compare anatomical changes, starch accumulation patterns, and protein induction in rice roots under drought conditions. The findings of this study would assist in developing new screening methods and markers for the management of water stress in upland environments.
Materials and Methods
Plant Materials and Growth Conditions
Four drought-resistant rice varieties, Nagina 22, Azucena, OS-6, and NDR-1025-2 and four drought susceptible varieties, IR64, Saita, BPT-5204, and Suryamukh were investigated for the study of different drought-linked characters in the year 2004–2005 and 2005–2006. PVC pipes (100 cm long × 15 cm wide, attached with plastic membrane inside) were filled with upland soil (oxidizing soil, texture sandy loam, pH 7.8, EC 1.9 (ds/m), CEC Me/100 mg of soil 13.6, organic carbon 0.48 %, extractable nitrogen 126.3 kg/ha, and water saturation 19.20 %). Nitrogen, phosphorus, and potash were added at a rate of 60:40:40 kg/ha respectively, half of nitrogen, total phosphorus, and potash were added in two equal split doses, one at tillering and another at the time of panicle emergence. After burring the PVC pipes to the soil, five seeds of each rice variety were directly sown in each pipe.
Water Regime Treatments
Two water regimes, saturated and drought condition, were artificially created in each PVC pipe. The rain out shelter was manually pushed to cover the PVC pipe area to avoid the rainfall. The level of water in each PVC pipe was maintained by normal irrigation throughout the growing period of the crop in control condition, whereas in the drought treatment, water was not applied after 60 days of sowing for the next 15 days to the rice varieties.
Determination of Morphological and Yield Attributes
Three plants were randomly selected from each variety for the determination of root length (RL), sterility percentage (SP), and grain yield per plant (GY) under control and drought conditions. The rice plants were safely pulled out along their roots from the PVC pipe in which they were planted. Root lengths of each of the rice varieties were individually measured.
Leaf Water Potential
Leaf water potential was measured using a pressure chamber (Soil Moisture Equipment, Santa Barbara, CA, USA). Midsection of the second youngest fully expanded leaf blade was used in all experiments. The detached leaf was sealed in a steel chamber with only the cut end (petiole) protruding out. Pressure was applied to the chamber from a compressed nitrogen cylinder until a sap meniscus appeared at the xylem surface. The pressure reading was recorded as the leaf water potential.
Anatomical and Histochemical Studies
Root samples from control and drought were collected and washed with distilled water. Further samples were fixed in formalin aceto alcohol (FAA) solution [ethyl alcohol (50 %) 90 ml + glacial acetic acid 5 ml + commercial formalin 5 ml (total 100 ml volumes)] for 4 h. The roots were then transferred to fresh FAA solution, kept in vials, and stored at 40 °C for further use. For studying, section cutting roots were fixed into potato pith and a very thin section was cut with the help of s single-edge blade. Root sections were stained as described by Johanson [15] and transverse sections were examined under stereoscopic microscope (Lica, Germany). Finally, photographs were taken with a digital camera (Canon) attached to a microscope.
Protein Isolation and SDS-PAGE Profiling
Fresh root samples were homogenized in pre-chilled pestle and mortar in phosphate buffer. The homogenates were centrifuged at 10,000 × g for 15 min. After centrifugation, clear supernatants were collected, which were lyophilized up to 1 ml and protein content in each sample was determined as described by Bradford [3]. The lyophilized homogenates of each sample (~5 μg) of control and drought condition were electrophoresed on 12 % SDS-PAGE [18]. Initially, electrophoresis was started at 10 mA of constant current until the tracking dye entered to the separating gel and then continued at 25 mA until the tracking dye reached the end of the gel. The electrophoresed gel was finally stained with silver nitrate [7]. Relative molecular weights of proteins were determined by using a standard curve generated from the standard proteins marker (Merck India Limited).
Statistical Analysis
Data on dry tiller numbers, root lengths, sterility percentage, grain yield per plant, and water potential were analyzed using two-way analysis of variance (ANOVA). The subsequent multiple comparisons among the means of treatments, genotypes, and treatments by genotype interactions were examined based on Panse and Sukhatme [30].
Results
Morphological Studies
Root Length
In control conditions, the maximum root length was recorded in Suryamukh (70.30 cm) followed by IR 64 (60.00 cm) and minimum in Saita (30.00 cm). Under the drought conditions, root length increased in all the rice varieties (Table 1). However, maximum elongation in root length was recorded in var. Nagina 22 (35.81 %) followed by OS-6 (25.12 %) and minimum root elongation was recorded in BPT-5204 (11.11 %) followed by IR 64 (14.28 %).
Sterility Percentage
In control conditions, maximum sterility was recorded in var. BPT-5204 (19.96 %) followed by var. Saita (17.90 %), whereas minimum sterility was recorded in var. Nagina 22 (8.23 %). Water stress significantly increased sterility percent in all the rice varieties. After drought, maximum sterility was also recorded in var. BPT-5204 (53.73 %) and minimum in var. OS6 (29.33 %). However, maximum percent increase in sterility after drought was recorded by var. BPT-5204 (33.77 %). The resistant rice varieties had less increase in sterility after drought than the susceptible rice varieties. Minimum sterility was recorded in var. Nagina 22 (11.77 %). The interaction among the varieties and treatments was found to be significant (Table 1).
Grain Yield per Plant
Under the drought conditions, mean grain yield of all the tested rice varieties was significantly reduced. Maximum yield reduction was observed in var. IR64 (38.40 %), followed by var. BPT-5204 (38.07 %). The drought-tolerant rice var. Nagina 22 showed minimum yield reduction (9.82 %) after drought treatment (Table 1). Positive regression coefficient (r2 = 0.0459) was obtained between the root length and grain yield (Fig. 1b).

Regression coefficient relation between root length and grain yield in the 13 different rice varieties under water stress conditions
Leaf Water Potential
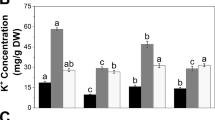
Leaf water potential declined significantly in all the tested varieties after drought treatment. The decline in leaf water potential was minimum in var. Nagina 22 followed by var. OS-6 and was maximum in var. Saita after drought stress (Table 1).
Anatomical and Histochemical Analysis
Anatomical features of the transverse sections of rice roots clearly distinguished resistant and susceptible varieties under the control and drought conditions. The outermost layer contained unicellular epiblema followed by the cortex region made of multilayer cortical parenchymatous tissue and finally vascular bundles containing xylem and phloem. The drought resistant varieties had denser aerenchyma compared to the susceptible varieties both under control and drought conditions. Among the resistant varieties, Nagina 22 had the largest cortex, vessels and closely linked parenchyma with fewer intercellular spaces compared to the susceptible varieties after drought treatment (Fig. 2a, b).

a Anatomical characteristics of aerenchyma formation in the root of drought-susceptible rice variety Saita. b Anatomical characteristics of aerenchyma formation in the root of drought-resistant rice variety Nagina 22
Histological observation of starch accumulation in roots was seen after staining with the IKI solution. The starch accumulation pattern also varied in resistant and susceptible rice varieties in control as well as in drought conditions. Accumulation of starch was mainly observed in secondary xylem parenchyma and secondary phloem parenchyma of the roots. Drought-susceptible varieties Saita and BPT-5204 showed least level of starch accumulation in their roots. Rice var. Nagina 22 showed highest level of starch accumulation under drought conditions among all the tested varieties.
SDS-PAGE Profiling
Changes in protein bands in SDS-PAGE gel (i.e., up-regulation/down-regulation) under drought conditions were observed in resistant and susceptible varieties. Protein bands of 35 ± 2, 40 ± 2, 50 ± 2, and 56 ± 2 kDa, were observed in vars Nagina 22 and Ratna under drought conditions. A novel protein band of 39 ± 2 kDa was found only in var. Nagina 22. About 3–4 protein bands in the range of 66–90 kDa were absent in the drought-susceptible rice varieties under drought conditions, whereas a common protein band of 55 ± 2 kDa was found in all the tested rice varieties under drought conditions.
Discussion
To study the root growth of rice, under controlled environments [24], a protocol of drought treatment by growing plants in polyvinyl chloride (PVC) pipes was developed. In PVC pipe test, drought avoidance laid to the deep root systems and small plant sizes as distinguished by applying a uniform severity of drought stress to each plant [49]. Elongation in root length, under drought conditions, was observed in all the rice varieties, though the drought-tolerant rice varieties showed greater root length elongation than the susceptible varieties.
Root development is fundamentally involved in the response to plant stress, particularly drought and mineral stress. It has been hypothesized that coarse roots have a direct role in drought resistance because of their penetration ability [6]. O’Toole [28] also suggested that under a large soil water reservoir and without root growth barriers, rooting should be dense and deep, while with a small soil water reservoirs, increased root penetration of physical and chemical impedance may be required. The ability to access deep water from the soil profile is an important drought-resistance trait in upland rice and substantial genetic variation exits for this character [10]. Limited efforts have been made to investigate the input side of the water balance, i.e., the acquisition of water from the soil. Basic hydraulic properties of roots are not yet adequately understood, simply because roots in soil are much less accessible than shoots. During the stress conditions, the anatomy of roots changes largely due to stress (such as water deficit), which induces the development of apoplastic barriers for water and ion flow [43].
Mambani and Lal [23] reported that good water potential in plants, having deeper root system, improves the water uptake from the deeper soil strata. Significant reductions in grain yield was observed in all the varieties tested, however, in resistant varieties, the reduction of grain yield was less. The less reduction in grain yield in resistant varieties showed that water status was maintained to some extent under drought stress. Drought stress significantly increased sterility in all the varieties tested; however, the degree of sterility was less in drought-resistant varieties. Liu et al. [21] also suggested that water status probably has a more direct effect on spikelet fertility, grain filling, and, finally, yield. It has been reported that water potential of well-watered plants can decline with ontogeny due to increasing demand for water as the leaf area increases [9]. In this study, water potential in the watered plants was maintained in the range that well-watered plants are expected to maintain [40]. Thus, the possibility that water deficits developed in watered plants to levels that could substantially reduce the magnitude of osmotic adjustment was minimal. Rice var. Nagina 22 had minimum reduction in the leaf water potential status that showed the maintenance capability of leaf water potential under stress conditions. Maintenance of plant water status is a fundamental phenomenon for the maintenance of normal growth of plants under stressful environment [2].
The transverse sections of rice roots (viz., upper, middle, and lower), revealed specific characteristics such as well-developed aerenchyma in mature parenchymatous cortex and vascular bundle with xylem and phloem. Large number of aerenchyma formation with shrinkage of cortex were observed in drought-susceptible varieties when subjected to drought treatment. However, aerenchyma formation has been observed under drought conditions in both aerobic and lowland genotypes, although to a lesser extent than under flooded conditions [42].
Resistance var. Nagina 22 had larger cortex and less aerenchyma in control as well as after drought treatment among all the tested rice varieties. Sibounheuang et al. [39] reported that xylem diameter is related to maintenance of xylem water conductivity. An increased xylem vessel size has been hypothesized to be a useful trait for improving water extraction from deeper soil layers [47]. Rice roots have also been reported to be highly susceptible to cavitation (the collapse of a continuous water column in the xylem due to air bubble formation). Stiller et al. [41] reported that cavitation occurs when axial water flow in the xylem vessels cannot keep up with the transpiration rate. Although drought condition has been observed to affect aerenchyma formation, it is unknown whether aerenchyma formation affects water uptake in rice. The ideal upland rice root system is thought to be composed of only a few thick and long roots with large xylem vessels capable of extracting water in the deep soil layers [1, 10].
Starch is the major constituent and energy reserve in many important foods, including the seeds of cereal grains and the potato tuber. In plants, it also plays important roles in the development and tolerance against various abiotic stresses. Different intensity of starch staining pattern was observed in root sections. However, the drought-tolerant varieties had a higher starch content than the susceptible varieties around the vascular bundles and periphery region of roots after drought. When rice suffers drought at the flowering stage, substantial quantities of carbohydrates accumulate in different parts in rice plants [31]. Other cereals also accumulate carbohydrates that are of significance when plants experience water deficit [33, 5].
The accumulation of medium molecular weight proteins in roots after drought stress in tolerant varieties, especially in var. Nagina 22 were observed. These proteins might be stress-responsive proteins [12]. Our finding of extra novel protein induction after drought stress in Nagina 22 is very similar to the finding observed by Chandra and Tyagi [4] who reported a higher number of polypeptide in var. Nagina 22 under ABA treatment. Mushtaq et al. [25] also reported an interesting finding with the correlation of drought and starch activity after drought stress in rice seed endosperm. The up-regulated and down-regulated proteins in response to starch content accumulation in rice pith of drought stressed roots may be useful additional tools for the screening and understanding of drought stress, especially in rice.
References
Abd Allah AA, Shimaa AB, Zayed BA, Gohary AAE (2010) The role of root system traits in the drought tolerance of rice (Oryza sativa L.). Int J Agric Biol Sci 1(2):83–87
Ali Q, Ashraf M (2011) Induction of drought tolerance in maize (Zea mays L.) due to exogenous application of trehalose: growth, photosynthesis, water relations and oxidative defense mechanism. J Agron Crop Sci 194(4):258–271
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem 7:248–254
Chandra A, Tyagi A (2004) Protein profiles of two rice varieties by 2-D gel electrophoresis under moisture stress. Indian J Biochem Biophys 41:191–194
Chaturvedi GS, Ingram KT (1988) Assimilate translocation of lowland rice in response to water deficit and shade. Philipp J Crop Sci 13(1):55–59
Clark LJ, Price AH, Steele KA, Whalley WR (2008) Evidence from near-isogenic lines that root penetration increases with root diameter and bending stiffness in rice. Funct Plant Biol 35:1163–1171
Damerval C, Guillox ML, deVienne BD (1987) A simplification of Heukeshovven and Dernick’s silver staining of proteins. Electrophoresis 8:158–159
Fisher KS, Fukai S (2003) How rice responds to drought. In: Fisher KS, Lattife R, Fukai S, Atlin G, Hardy B (eds) Breeding rice for drought prone environments. International Rice Research Institute, Los Banos, pp 32–36
Flower DJ, Usha Rani A, Peacock JM (1990) Influence of osmotic adjustment on the growth, stomatal conductance and light interception of contrasting sorghum lines in a harsh environment. Aust J Plant Physiol 17:91–105
Fukai S, Cooper M (1995) Development of drought resistant cultivars using physiomorphological traits in rice. Field Crop Res 40:67–86
Hanson AD, Peacock WJ, Evans LT, Arntzen CJ, Khush GS (1990) Drought resistance in rice. Nature 345:26–27
Hashimoto M, Kisseleva L, Sawa S, Furukaw T, Komats S, Koshiba T (2004) A novel rice PR10 protein, RSOSPR10, specifically induced in roots by biotic and abiotic stresses, possibly via the Jasmonic acid signaling pathway. Plant Cell Physiol 45(5):550–559
Ingram J, Bartels D (1996) The molecular basis of dehydration tolerance in plants. Annu Rev Plant Physiol 47:377–403
Ishitani M, Nakamura T, Han SY, Takabe T (1995) Expression of the betaine aldehyde dehydrogenase gene in barley in response to osmotic stress and abscisic acid. Plant Mol Biol 27:307–315
Johansan DA (1940) Plant microtechnique, 5th edn. McGraw-Hill, New York, p 523
Jongdee B, Cooper M (1998) Genetic variation for grain yield of rice under water deficit condition. In: Proceedings of the 9th Australian Agronomy Conference, Wagga Wagga, Australia, pp 403–406
Khush GS (2005) What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol Biol 59:1–6
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685
Lisse T, Bartels D, Kalbitzer HR, Jaenicke R (1996) The recombinant dehydrin-like desiccation stress protein from the resurrection plant Craterostigma plantagineum displays no defined three-dimensional structure in its native state. Biol Chem 377:555–561
Liu H, Mei H, Yu X, Zou G, Liu G, Luo L (2007) Towards improving the drought tolerance of rice in China. Plant Genet Resour 4(01):47–53
Liu G, Mei HW, Yu XQ, Zou GH, Liu HY, Hu SP, Li MS, Wu JH, Chen L, Luo LJ (2008) QTL analysis of panicle neck diameter, a trait highly correlated with panicle size, under well-watered and drought conditions in rice (Oryza sativa L.). Plant Sci 174:71–77
Luo LJ, Zhang QF (2001) The status and strategy on drought resistance of rice (Oryza sativa L.). Chin J Rice Sci 15:209–214
Mambani B, Lal R (1983) Response of upland rice varieties to drought stress III. Relation between root system development and leaf water potential. Plant Soil 73:59–72
Matsuo N, Mochizuki T (2009) Genotypic differences in root traits of rice (Oryza sativa L.) seedlings grown under different soil environments. Plant Root 3:17–25
Mushtaq R, Katiyar S, Bennett J (2008) Proteomic analysis of drought stress-responsive proteins in rice endosperm affecting grain quality. J Crop Sci Biotechnol 11(4):227–232
Neslihan-Ozturk Z, Talame V, Deyholos M, Michalowski CB, Galbraith DW, Gozukirmizi N, Tuberosa R, Bohnert HJ (2002) Monitoring large-scale changes in transcript abundance in drought and salt stressed barley. Plant Mol Biol 48:551–573
Nguyen HT, Babu RC, Blum A (1997) Breeding for drought resistance in rice: physiological and molecular genetics considerations. Crop Sci 37:1426–1434
O’Toole JC (1982) Adaptation of rice to drought prone environments. In: Drought resistance in crops with emphasis on rice. IRRI, Los Banos, pp 195–213
Pandey S, Bhandari H (2009) Drought: economic costs and research implications. In: Serraj R, Bennet J, Hardy B (eds) Drought frontiers in rice: crop improvement for increased rainfed production. World Scientific Publishing, Singapore, pp 3–17
Panse VG, Sukhatme PV (1967) Statistical methods for agricultural workers. Indian Council of Agricultural Research, New Delhi, p 381
Perez CM, Palmino EP, Baun CL, Julliano BO (1971) Starch metabolism in leaf sheaths and culm of rice. Plant Physiol 47:404–408
Rabello AR, Guimaraes CM, Rangel PHN, daSilva FR, Seixas D, deSouza E, Brasileiro AC, Spehar CR, Ferreira ME, Mehta A (2008) Identification of drought responsive genes in roots of upland rice (Oryza sativa L.). BMC Genomics 9:485–498
Rahman MS, Yashida S (1985) Effect of water stress on grain filling in rice. Soil Sci Plant Nutr 31(4):497–511
Riccardi F, Gazeau P, de Vienne D, Zivy M (1998) Protein changes in response to progressive water deficit in maize. Quantitative variation and polypeptide identification. Plant Physiol 117:1253–1263
Salekdeh GH, Siopongco J, Wade LJ, Ghareyazie B, Bennett J (2002) Proteomic analysis of rice leaves during drought stress and recovery. Proteomics 2(9):1131–1145
Salekdeh GH, Siopongco J, Wade LJ, Ghareyazie B, Bennett J (2002) A proteomic approach to analyzing drought- and salt-responsiveness in rice. Field Crops Res 76:199–219
Serraj R, McNally KL, Slamet-Loedin I, Kohli A, Haefele SM, Atlin G, Kumar A (2011) Drought resistance improvement in rice: an integrated genetic and resource management strategy. Plant Prod Sci 14:1–14
Sharp RE (2002) Interaction with ethylene: changing views on the role of abscisic in root and shoot growth responses to water stress. Plant Cell Environ 25:211–222
Sibounheuang V, Basnayake J, Fukai S (2006) Genotypic consistency in the expression of leaf water potential in rice (Oryza sativa L.). Field Crops Res 97:142–154
Sinclair TR, Ludlow MM (1986) Influence of water supply on the plant water balance of four tropical grain legumes. Aust J Plant Physiol 13:329–341
Stiller V, Lafitte HR, Sperry JS (2003) Hydraulic properties of rice and the response of gas exchange to water stress. Plant Physiol 132:1698–1706
Suralta RR, Yamauchi A (2008) Root growth, aerenchyma development, and oxygen transport in rice genotypes subjected to drought and water logging. Environ Exp Bot 64:75–82
Taleisnik E, Peyrano G, Cardoba A, Arias C (1999) Water retention capacity in root segments differing in the degree of exodermis development. Ann Bot 83:19–27
Wade LJ, Fukai S, Samson BK, Ali A, Mazid MA (1999) Rainfed lowland rice: physical environment and cultivar requirements. Field Crops Res 64:3–12
Wang H, Zhang H, Li Z (2007) Analysis of gene expression profile induced by water stress in upland rice (Oryza sativa L. var. IRAT109) seedlings using subtractive expressed sequence tags library. J Int Plant Biol 49(10):1455–1463
Weng JW, Agata Takeda T (1986) Studies on dry matter and grain production in rice plants, i.e., varietal differences of total carbohydrates content of heading stage. Jpn J Crop Sci 55(2):201–207
Yambao EB, Ingram KT, Real JG (1992) Root xylem influence on the water relations and drought resistance of rice. J Exp Bot 43:925–932
Yang L, Zheng B, Mao C, Qi X, Liu F, Wu P (2004) Analysis of transcripts that are differentially expressed in three sectors of the rice root system under water deficit. Mol Genet Genomics 272(4):433–442
Yue B, Xue W, Luo L, Xing Y (2008) Identification of quantitative trait loci for four morphologic traits under water stress in rice (Oryza sativa L.). J Gen Genomics 3:569–575
Acknowledgments
We are highly thankful to Dr. G.S. Chaturvedi, Department of Plant Physiology, and Dr. A.C. Verma, Department of Nematology, N.D. University of Agriculture & Technology, Kumarganj, Faizabad, India, for providing the water rain out shelter and photography facility during the course of investigation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Singh, A., Shamim, M. & Singh, K.N. Genotypic Variation in Root Anatomy, Starch Accumulation, and Protein Induction in Upland Rice (Oryza sativa) Varieties Under Water Stress. Agric Res 2, 24–30 (2013). https://doi.org/10.1007/s40003-012-0043-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40003-012-0043-5