
Abstract
Key message
The disturbance of a research plot by a windstorm allowed us to study the role of the seedling bank in the regeneration processes. The released advance regeneration dominated among the saplings; taller individuals retained their position until the end of the study. Pioneer species occurred sporadically. Seven years after the disturbance, the windthrow was covered by a dense thicket of young trees.
Context
The dominant role played by advance regeneration in natural regeneration processes after intense wind disturbances is still a matter of dispute.
Aims
We took advantage of a windstorm in one of our research plots to study the role of the seedling bank released by the disturbance in the regeneration processes.
Methods
We collected data in 70 plots, recording the survivorship of seedlings, annual height growth, and signs of browsing. The height ranking was analyzed with Kendall’s concordance coefficient, and the height growth rates were compared using Dunn’s test.
Results
The density of seedlings increased from 6.7/m2 in 2008 to 8.1/m2 in 2010 and then decreased to 1.2/m2 in 2015. The density of saplings increased continuously from 0.14 to 1.9/m2. The highest size differentiation occurred in sycamore maple; the individuals which were taller before the windstorm retained their position until the year 2015. The only species that was recruited mainly from germinants was European hornbeam.
Conclusion
The advance regeneration released by the windstorm played a major role in the regeneration process, while pioneer species occurred only sporadically. Seven years after the disturbance, the windthrow was already covered by a dense thicket of young trees.
Similar content being viewed by others
1 Introduction
Natural disturbances have been widely recognized as a driving force in forest ecosystem dynamics (Frelich 2002; Johnson and Miyanishi 2007). While small disturbances usually release the advance regeneration of late-successional species (Woods 2004), large and intense disturbances that can kill most of the canopy trees in the whole stand allow for colonization of the disturbed area by pioneer species (Bobiec 2007; Jaloviar et al. 2017). However, the mortality of canopy trees after a large and intense disturbance can be much lower than 100% and the role of pioneer trees can be limited by competition with the remnants of the old stand (Nagel et al. 2006).
Hurricanes, downbursts, and tornadoes are examples of large and intense disturbances (Turner et al. 1997, Greenberg and Collins 2015). Mortality rates of canopy trees after wind disturbance rarely exceed 50% (Wolf et al. 2004; Woods 2004). Partly damaged live trees belong to important legacies (Franklin et al. 2007) of wind disturbances. Therefore, the conditions for the development of a young generation of trees (both advance regeneration and new recruitment) vary according to the local density of mature trees that have survived the disturbance. Windthrows and breakage of stems brought about by severe winds do not necessarily lead to tree mortality (Peterson 2000; Canham et al. 2001). Some species are able to survive and rebuild damaged crowns or to develop secondary shoots from windthrown stems lying on the ground (Peterken 1996). A substantial role in the development of the new generation of trees could be played by vegetative reproduction of these species (Koop 1987).
The rates of regeneration of forest communities following intense wind disturbances vary among different forest types. In tropical forests, regeneration processes can last less than 10 years (Brokaw 1985; Marra et al. 2014), while in boreal forests, the regeneration processes span several decades (Rich et al. 2007). Rates of forest regeneration in temperate forests are difficult to determine, as in most cases large-scale wind disturbances in these forests are followed by salvage-logging and tree planting (Lindenmayer et al. 2012), and the opportunities to study natural processes are very few (Palik and Kastendick 2009; Buma and Wessman 2012; Vodde et al. 2015). According to the results of studies conducted in mixed temperate forest, regeneration processes can be very variable in terms of speed (Gill et al. 2017): in poor habitats regeneration can be relatively slow (Dobrowolska 2015), while under good site conditions regeneration processes can be very fast (Bartels et al. 2016).
In the case of wind disturbance, an important role could be played by the seedling bank, present under the canopy prior to the disturbance (Nagel et al. 2006; Franklin et al. 2007). Seedling banks usually consist of shade-tolerant species and thus can contribute to maintaining a big share of shade-tolerant species in post-disturbance stand development (Nagel et al. 2007; Palik and Kastendick 2009). They can play a significant role after wind disturbances, as little damage is done by wind to the young trees growing on the forest floor. Moreover, the spatial heterogeneity of light intensity following wind disturbance provided by slowly dying crowns of broken or uprooted trees allows for acclimation to the changing light conditions even for very shade-tolerant species (Čater and Diaci 2017).
One of the important problems associated with natural regeneration after disturbances is the disturbance interactions (Veblen et al. 1994; Buma and Wessman 2011). In coniferous forests, some blown down areas experience more severe wildfires (Kulakowski and Veblen 2007; Buma and Wessman 2011) or insect outbreaks (Stadelmann et al. 2013). In mixed deciduous temperate forests, wildfires and insect outbreaks are rare, but an important interaction can occur between wind disturbance and herbivory by ungulate browsers. Gaps created by windstorms are strongly preferred by foraging ungulates (Bobiec 2007), but accelerated growth rates of young trees in wind-disturbed areas (Kupferschmid et al. 2013) could allow the highly palatable species to regenerate successfully after windstorms (Keren et al. 2017). In our paper, we studied regeneration processes after a windstorm that blew down a patch of forest in the Roztoczanski National Park, SE Poland. One of the permanent research plots, which had been measured 1 year before, was located in the middle of the area affected by the windstorm. After disturbance of the entire site, the windthrow was set-aside for studying natural regeneration. The fact that field data were collected immediately before the disturbance allowed us to study the regeneration process in detail; we could follow the fate of young trees released by the disturbance along with the establishment of new cohorts of seedlings.
Advance regeneration plays an important role in the development of the new stands following large and intense disturbances (Veblen 1986, Nagel et al. 2006, Cater and Diaci 2017). This is especially true for windstorms, as wind does little harm to the seedlings and saplings (Franklin et al. 2007). That gives an advantage to shade-tolerant species, which usually form the advance regeneration and are in a better position compared to the seedlings recruited after the disturbance (Diaci 2017); in extreme cases, it can even contribute to the long-term dominance of late-successional species (Jaloviar et al. 2017).
In our study, we formulated five specific hypotheses concerning regeneration processes:
-
1.
Mortality rates of canopy trees after wind disturbance differ strongly among species, and some badly damaged trees of deciduous species survive and are able to regenerate vegetatively.
-
2.
Advance regeneration of shade-tolerant species suffers little damage during the windstorm and is most abundant in the regeneration process after disturbance.
-
3.
Sycamore maple and hornbeam will increase in relative abundance among the regenerating trees because they thrive under high light intensity.
-
4.
In mixed deciduous temperate forests, seedlings of pioneer tree species will establish only during the 3 years after the windstorm; after 4 years the density of saplings is so high that it will prevent further recruitment of shade-intolerant species.
-
5.
Fast growing seedlings and saplings of palatable species are able to reach safe heights despite intense browsing by ungulates in the area of the windthrow.
2 Methods
2.1 Study area
The Roztocze National Park (RNP; coordinates 50° 31′–50°40′ N, 22° 53′–23° 07′ E) is situated in the central part of the Roztocze Highlands in the South-East part of Poland. The landscape is characterized by long chains of Late Cretaceous limestone hills reaching 390 m a.s.l. surrounded by thick layers of postglacial deposits, sand, or loess. The most common soil types are podzols and cambisols. The climate of the Roztocze region is characterized by warm and usually dry summers (mean monthly temperature in July is 17.2 °C) as well as cold and snowy winters (mean monthly temperature in January is − 4.3 °C, long-term average number of frost days is 38.1 per year and the long-term average yearly duration of the snow cover is 71 days). Mean yearly temperature is about 7.3 °C. The yearly amplitude of the mean temperatures often exceeds 22 °C. The growing season lasts usually 210 days. The mean annual precipitation in the Roztocze region ranges from 650 to 750 mm (Kaszewski 2008).
The most common forest types are mixed European beech-silver fir-hornbeam stands (25% of the Park area), Scots pine stands with a small admixture of deciduous trees (19%) and almost pure silver fir stands (8.5%). Out of 40 tree species native to Poland, 32 occur naturally in the RNP. However, 90% of the forest stands are dominated by three species: European beech Fagus sylvatica L., Scots pine Pinus sylvestris L., and silver fir Abies alba Mill. (Izdebski et al. 1992). The current area of the RNP is 8483 ha, 95% of which is covered by forests. The majority of the forests have retained their natural character and they have been recognized as remnants of the European primary forests (Sabatini et al. 2018). The main types of natural disturbance in RNP are windstorms (winter storms and downbursts associated with summer storms) and accumulation of wet snow in the early autumn, before the leaves of deciduous trees are shed. No large wildfires or insect outbreaks have been recorded since the establishment of the park (Maciejewski and Szwagrzyk 2011).
2.2 Field data collection
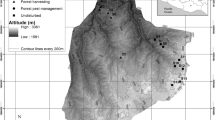
The original research plot, with a size of 0.5 ha (rectangle 50 × 100 m), was established in 1973 to study changes in species composition and the structure of secondary forest stands. All trees exceeding DBH of 7 cm were measured (DBH and height) and numbered within 10 × 10 m quadrats. A set of circular subplots of 0.5 m2 each was established with regular spacing (Fig. 1) to study the natural regeneration of trees and the composition of the herbaceous vegetation. The re-measurements of the plot were completed in 1997 and in 2008, by the end of July. The time between consecutive measurements (1997, 2008) was too long to provide reliable information about the demographic processes in seedlings and saplings.

Scheme of study area; mature trees were measured and mapped in entire 50 × 100 m plot, while circular sample plots (area—0.5 m2) were used for measuring forest regeneration. Filled (34) circles—plots established in 1973, hollow (36) circles—plots established in 2009; one 10 × 10 m quadrat represents basic unit, in which canopy trees were measured and numbered prior to windstorm
In August 2008, the area was hit by a windstorm, resulting in a blowdown with a size of about 2.5 ha. The entire research plot was within the limits of the area disturbed by the wind.
In the year 2009, the circular plots were re-measured again, and an additional 36 circular plots for monitoring tree regeneration and forest floor vegetation were added (Fig. 1). Seedlings and saplings growing in the small circular plots were numbered and labeled to follow their fate. In the years 2010, 2011, and 2015 the measurements of the heights of the seedlings and saplings were repeated in all 70 circular sample plots. Signs of ungulate browsing were recorded for all seedlings and saplings in every year throughout the study. The recording of browsing was conducted twice during the growing season (in spring and in summer), and for each individually tagged seedling or sapling, the location of the browsing (top of stem, side branches) was described.
New individuals established in the circular plots were also recorded and measured. The vitality of the mature trees in the wind-disturbed area was checked visually every year at the end of the growing season, and the time of death of each canopy tree was determined.
2.3 Methods of data analysis
First, we analyzed the distribution of the heights of the trees belonging to three major species (sycamore maple Acer pseudoplatanus L., European hornbeam Carpinus betulus L., and European beech) as well as their growth rates in consecutive periods. We checked the normality of these distributions by statistical tests: visualizing Q-Q (quantile-quantile) plot, Shapiro-Wilk test, and Bartlett’s test. The distributions were characterized by high skewness and data heteroscedasticity. However, because the sample sizes for the groups were large enough (Rutherford 2001), we employed Welch’s ANOVA followed by the Games-Howell post hoc test along with the non-parametric Kruskal-Wallis test followed by Dunn’s test.
To determine the differences in the height increments among the three species over the four studied time periods, we calculated the intra-species Gini coefficients as measures of their distribution inequalities. After arranging the data values in order from the lowest value to the highest value, we employed the following formula (Dixon et al. 1987):
where:
- GST:
-
Gini coefficient value for species S in time period T,
- n :
-
the number of individuals for species S in time period T,
- x i :
-
height increment value for ith individual,
- x :
-
mean value for all individuals of species S in time period T.
To determine whether the young trees maintain their initial height advantage, we compared the ranks of individual trees among the consecutive measurements. We tested the consistency of height rankings during the study period using Kendall’s coefficient of concordance (KCC) (Legendre 2005). In our analysis, we took into account only the trees for which we had height measurements in all years, i.e., in 2009, 2010, 2011, and 2015. We calculated the KCC for each pair of height rankings from the above-mentioned years along with chi-squared significance tests to identify which of them were concordant with one another and what was the strength of this association.
To compare the intensity of the ungulate browsing among tree species, we used the data showing how many times the individual trees had been browsed during the period of our study. We analyzed the number of browsing episodes along with the species identity of young trees. As the Cameron and Trivedi test (Cameron and Trivedi 1990) suggested that data may be underdispersed, we used a generalized Poisson regression (Hilbe 2014) to estimate the parameter values in the model where the response variable was the number of browsing events per individual and its species identity was an explanatory variable. The indicator variable was the expected difference in the log count between particular species and the reference group of sycamore maple. Hence, the incidence rate for particular species was the exponent of the coefficient value multiplied by the incidence rate for the reference.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
3 Results
3.1 Tree mortality after windstorm
Prior to the windstorm, the stand density was 534 trees/ha and the basal area (BA) was almost 40 m2/ha. According to tree numbers, Scots pine was less than 42%, hornbeam almost 30% and beech 24%. By BA, Scots pine was the dominant species, with almost 47% of BA, beech was slightly over 31%, and hornbeam had less than 16% (Table 1).
The total mortality in the study plot amounted to 113 trees, i.e., 40.45% of all trees prior to the windstorm. Mortality calculated on the basis of BA was even higher: 48.94%. The only species that was completely eliminated from the stand was wild cherry Prunus avium (L.) L. The highest mortality by tree number was recorded in Scots pine (59%). High mortality (48%) occurred also in European beech and in sessile oak Quercus petraea (Matt.) Liebl. (50%). The mortality rate calculated by BA was higher in European beech (61%) than in Scots pine (55%).
The mortality was distributed unevenly among the tree species in consecutive years after the windstorm. Most of the Scots pines that were broken or uprooted by the windstorm died in the first year, while in the case of European beech the mortality was concentrated in the second and third years after the windstorm (Table 2). In European beech, almost 30% of the uprooted trees survived until 2015. The extreme case was European hornbeam where out of 43 uprooted trees only five had died by the year 2015, and out of nine broken hornbeams, eight trees survived and rebuilt their crowns, so total mortality in that species amounted to less than 8%.
3.2 Changes in densities of seedlings and saplings
The density of seedlings prior to the windstorm amounted to 6.7 individuals per square meter (ind./m2), and the most abundant species among the seedlings was sycamore maple (56%) followed by European beech (33%). At the same time, the density of the saplings was much lower (0.14 ind./m2), and the most abundant species among the saplings was European beech (89%), sycamore maple had 9%, and the other species occurred only as single individuals (Fig. 2).

Densities of seedlings and saplings in study area in years 2008–2015
The total densities of seedlings of all species increased slightly in the first 2 years following the windstorm and reached 8.1 ind./m2 in 2011; then they decreased to 4.4 ind./m2 in 2011 and 1.2 ind./m2 in 2015 (Fig. 2). That decline was a result of two processes: height growth resulting in reaching the sapling stage (height larger than 0.5 m) and mortality increasing with time as the sapling layer grew denser. European beech was abundant among seedlings since the beginning of this study (33% of all seedlings in 2008), and it slightly increased its share among seedlings by 2015. Seedlings of Scots pine arrived in 2010 and were still present in 2011; however, by the year 2013, they had died.
The density of the saplings increased throughout the period under study: it reached 0.3 ind./m2 in the year 2009 and increased steadily to 1.9 ind./m2 in 2015. Already in 2009, sycamore maple was the most abundant species among the saplings (69%); in the next year, its share remained high and then gradually declined to 34% in 2015. In the same time, the share of European hornbeam among the saplings increased from zero in 2009 to 35% in 2015. European beech was dominant among the saplings prior to the disturbance (89% of saplings in 2008). In the years 2009–2015, it maintained an almost constant share among the saplings (Fig. 2).
3.3 Growth rates of young trees
The three main groups of young trees were the advance regeneration—released by the disturbance, the new recruits—established after disturbance and vegetative sprouts of the trees damaged by the wind. The advance regeneration was the most numerous group, attaining the greatest heights. Newly recruited young trees were much lower than the advance regeneration. The vegetative sprouts were less numerous, but they had the largest mean height of these three groups (Fig. 3).

Comparison of height distributions of advance regeneration, new recruits and vegetative sprouts 7 years after windstorm
The analysis was focused on the three most common species: European beech, hornbeam, and sycamore maple. The average heights of the young generation of trees in the wind-disturbed area ranged in the year 2015 from 190.6 cm for European hornbeam to 230.5 cm for sycamore maple. However, the height distributions were strongly right-skewed, with many low saplings and a few tall ones (Fig. 4). The tallest young tree recorded in our plots was sycamore maple with the height of 940 cm; the tallest European hornbeam reached 755 cm and the tallest European beech 600 cm. The tallest specimens of European hornbeam were vegetative shoots produced by uprooted mature trees, and their average height was 458.6 cm. The young trees of generative origin for hornbeam were on average 115.2 cm tall.

Height distributions of young trees in wind-disturbed area in years 2008–2015
Height increments for the entire period under study (2009–2015) were calculated only for the young trees present throughout the entire period under study, and that group amounted to 232 individuals. The height increments were strongly right-skewed (Fig. 5), with average values ranging from 165.5 cm in European beech through to 189.2 cm in European hornbeam and to 198.7 cm in sycamore maple. Height increments calculated for young European hornbeams of generative origin were much smaller (113.4 cm). The largest height increment (824 cm) was recorded in sycamore maple.

Comparison of height increments of young individuals: 1—in the first 2 years after disturbance, 2—in the third and fourth years after disturbance, 3—from the fourth to seventh years after disturbance, and 4—over period of 7 years after disturbance. x—median, filled circle—outliers
The results from comparing the mean height increments among the tree species using Welch’s ANOVA followed by the Games-Howell post hoc test were consistent with the results of a non-parametric Kruskal-Wallis test followed by Dunn’s test (Table 3). The European beech had significantly smaller height increments than sycamore maple and European hornbeam in the beginning of the study, but for the entire period, the differences among those three species were insignificant (Table 3). European beech was the least variable in height growth, while in European hornbeam and sycamore maple, apart from individuals with large height growth rates, there were also many slow growing individuals (Fig. 5). This was also supported by the results of calculating the Gini coefficient, where the highest values were ascertained for sycamore maple and the lowest values for beech (Table 4).
3.4 Size hierarchy among young trees
On average, the European beech had the lowest Gini coefficient, and sycamore maple had the largest one. The values of the Gini coefficient fluctuated among years, and the only exception was hornbeam, in which the initial value of the coefficient was relatively low and rapidly increased in the second year of the measurements (Table 4).
On the basis of Kendall’s concordance coefficient (Table 5), we rejected the null hypothesis that any two height rankings were uncorrelated at the 0.05 significance level. In general, trees that had been taller in the beginning of the study tended to be also taller in the year 2015, and the tendency among smaller trees was consistent in a similar way (Table 5).
3.5 Sum of height of young trees per area unit
The sum of heights of the young trees increased from 2 m/m2 in 2009 to almost 14 m/m2 in 2015. The most spectacular increase was recorded in the case of European hornbeam: the sum of the height of seedlings and saplings for that species in 2009 was 10 times lower than the sum of young individuals of European beech, and in the year 2015, the sum of the height of young European hornbeam was substantially higher than that of European beech (Fig. 6). The species with the greatest sums of height was sycamore maple.

Sum of heights of young trees [m/100 m2]
3.6 Natural regeneration and deer browsing
Browsing of seedlings and saplings by ungulate herbivores was common throughout the study period; out of 127 young individuals of European beech, European hornbeam, and sycamore maple present in the sample plots from 2009 to 2015, 102 were browsed at least once. Only 25 individuals present from 2009 to 2015 were not browsed at all. The most browsed species was sycamore maple and the least browsed was beech (Table 6).
The results of the generalized Poisson regression indicated that European beech was about 0.37 times less likely to be browsed than sycamore maple. The difference between sycamore maple and hornbeam (0.43 times less likely to be browsed) was insignificant (Table 7). A similar model run without sycamore maple suggested that European hornbeam was more frequently browsed than European beech (coefficient ~ − 0.843) and that the difference was statistically significant (p value ~ 0.02).
4 Discussion
This study fortuitously captured data immediately before wind disturbance, and then we were able to follow the dynamics of the regeneration processes from there on a yearly basis, until the young trees in the study area had formed a dense thicket. Despite the small size of the study area and lack of repetitions, this has provided a unique and clear picture of the regeneration processes.
4.1 Mortality rates and patterns in canopy trees
The tree mortality after a windstorm (40%) was similar to that in many other studies on wind disturbance (Peterson 2000; Woods 2004; Szwagrzyk et al. 2017). European hornbeam seems to be most adapted to regenerating in a vegetative way (Faliński and Pawlaczyk 1993), and sycamore maple also regenerated well, especially trees that had been uprooted by the wind. In European beech, the ability to rebuild the crown was very low, and that agrees well with the results of earlier work (Fichtner et al. 2013). The process of mortality following the windstorm was prolonged; only in the case of Scots pine did all broken and uprooted trees die within 1 year after the disturbance.
The tree species present in the study area responded to the disturbance in different ways. European beech was characterized by high mortality and abundant recruitment. Two species—European hornbeam and sycamore maple—displayed low mortality and high recruitment. Scots pine was a loser as it suffered high mortality and had no effective recruitment. According to the classification of Batista and Platt (2003), the trees present in the study area could be classified as resilient (European beech), usurpers (European hornbeam and sycamore maple), and susceptible (Scots pine). No tree species displayed the resistant syndrome, which means low mortality and low recruitment.
4.2 Role of advance regeneration
Before the windstorm, the understory of the analyzed stand was covered with abundant seedlings, mostly of sycamore maple; however, the growth of this species was suppressed, so among the saplings, the most abundant species was European beech. The strategy of increasing the likelihood of long-term survival by reducing height growth rates has been described by Woods (2008) for sugar maple Acer saccharum Marshall. Our observations suggest that sycamore maple responds to light conditions in a similar way. On the other hand, some individuals of European beech growing in a similar light environment were able to attain the sapling stage, and this agrees well with the well-known shade tolerance in beech, which is higher than in any other deciduous tree in the Central European temperate forests (Ellenberg 1988).
The released advance regeneration formed the largest group among the young generation of trees, and along with vegetative sprouts, it dominated among the tallest saplings. That leaves little room for the new recruits; however, they were still abundant, but their chances in competition for light with their taller neighbors are low (Čater and Diaci 2017). The dominance of advance regeneration is not surprising, as similar situations had been already described (Nagel et al. 2006; Vodde et al. 2015).
4.3 Advantages of fast growth rates
The growth of the advance regeneration was surprisingly fast. The maximum height growth rates in sycamore maple reached 163, 195, and 190 cm in the 3 years after the windstorm. In European hornbeam, the growth rates of new stems of vegetative origin were even higher. In the year 2015, 7 years after the disturbance, the sum of the heights of the young trees had reached 1380 m/100 m2; this value is almost twice as large as the sum of the heights of trees in 30-year thickets of European beech growing under good habitat conditions (Szymkiewicz 1971).
Individuals that were taller than their neighbors at the time of the windstorm maintained their dominant position throughout our study. Suppressed individuals present under the forest canopy as well as seedlings established after the disturbance played a minor role in the regeneration process. Similar findings were reported by Uriarte et al. (2004) from subtropical forests of the Caribbean affected by recurring hurricanes.
The extremely high growth rates displayed by sycamore maple and European hornbeam in the area affected by the windstorm indicated that these species are able take advantage of the resources (light, nutrients) that were made available by the mortality following the disturbance. A similar situation was described for sugar maple by Leak (1987). Thus, European hornbeam and sycamore maple seemed to benefit from the wind disturbance. Both species were classified as “competitors” within the framework of the Grime’s classification of life history strategies (Brzeziecki and Kienast 1994). However, in the context of the findings from recent studies (Maciejewski and Szwagrzyk 2011; Szwagrzyk et al. 2012), their life history strategies should be considered as a combination of “C” (competitor) and “R” (ruderal) strategies.
4.4 Marginal role of pioneer species
In the first year after the windstorm, the crowns that were still alive of broken or uprooted trees covered much of the blowdown area, thereby creating partly shady conditions on the forest floor; similar conditions have been reported from other wind-disturbed areas left without any salvage-logging (Peterson 2000). Seedlings that were already present prior to the disturbance took advantage of the release (Veblen 1986; Nagel et al. 2006) and grew at high rates; in addition, herbaceous vegetation developed rapidly in the blowdown area. Like in other studies (Peterson 2000; Vodde et al. 2015), the only places suitable for the establishment of pioneer tree species were in the pits and mounds created by the windfallen trees. The single species that germinated in large numbers after the windstorm was European hornbeam, and this was probably due to abundant seed rain (Jankowska-Błaszczuk and Grubb 1997). In addition, wild cherry germinated after the disturbance, but was then eliminated by browsing pressure. Typical pioneer species, like silver birch Betula pendula Roth., goat willow Salix caprea L., and European aspen Populus tremula L. (Ellenberg 1988), played minor roles. In the first year after the windstorm 97 seedlings of Scots pine were found in 70 circular sample plots, but the number of Scots pine germinants declined dramatically. Most of the pine seedlings died in only a few years; only one of two specimens growing on elevated root plates attained the sapling stage. Similar results have been reported in several studies (McClure and Lee 1993, Cook 2000), and their authors suggested that a mineral seedbed is a factor controlling the regeneration of shade-intolerant, opportunistic species, and such microsites are very limited in wind-disturbed areas not subjected to salvage-logging.
Shrubs, like European elder Sambucus nigra L., red-berried elder Sambucus racemosa L., common dogwood Cornus sanguinea L., or dog rose Rosa canina L., took advantage of the disturbance and established on the elevated root plates and on windthrow mounds. The study by Herrera and García (2009) suggested that birds, using elevated places as perches, can disperse the seeds to wind-disturbed areas.
4.5 Role of deer browsing in regeneration process
Prior to the windstorm, 40% of the sycamore maple and 21% of the European beech saplings had been browsed by deer. These figures are comparable with the levels of browsing pressure recorded in other national parks in southern Poland (Miścicki and Żurek 2015; Bodziarczyk et al. 2017). After the windstorm, the percentage of browsed saplings increased to 61%; however, the high growth rates of the young trees in most cases compensated for the losses resulting from browsing and allowed the saplings to escape browsing of terminal shoots by growing tall enough in one or two growing seasons. An additional factor was the rapid arrival of numerous saplings of European hornbeam, and that species is strongly preferred by deer (Faliński and Pawlaczyk 1993), but according to recent findings, it is also the most resilient species growing in the temperate deciduous forest of Central Europe (Kuijper et al. 2010). Apparently, in the few years after the windstorm, the browsing pressure by ungulates was not able to slow down the regeneration processes of the trees. The successful growth of the seedlings and saplings could be partly attributed to the rapid development of herbs and shrubs in the windthrow area, which provided an alternative food source for the ungulates. However, the ungulate herbivory probably contributed significantly to the elimination of the pioneer species: willows, aspens, and wild cherries.
5 Final remarks
In the 7 years since the windstorm, the analyzed stand was a patchwork of groups of mature trees that survived the disturbance and dense young thickets dominated by released advance regeneration. European beech, which used to be one of the dominant species in the tree stand prior to the windstorm, lost many trees in the canopy, but retained its position due to the ample advance regeneration released by the disturbance. European hornbeam survived the windstorm very well and regenerated in the disturbed area in generative and vegetative ways. The species that was much more abundant in the regeneration than in the mature stand prior to the windstorm was sycamore maple.
References
Bartels SF, Chen HYH, Wulder MA, White JC (2016) Trends in post-disturbance recovery rates of Canada’s forests following wildfire and harvest. For Ecol Manag 361:194–207. https://doi.org/10.1016/j.foreco.2015.11.015
Batista WB, Platt WJ (2003) Tree population responses to hurricane disturbance: syndromes in a south-eastern USA old-growth forest. J Ecol 91(2):197–212
Bobiec A (2007) The influence of gaps on tree regeneration: a case study of the mixed lime-hornbeam (Tilio-Carpinetum Tracz. 1962) communities in the Białowieża Primeval Forest. Pol J Ecol 55:441–455
Bodziarczyk J, Zwijacz-Kozica T, Gazda A, Szewczyk J, Frączek M, Zięba A, Szwagrzyk J (2017) Species composition, elevation, and former management type affect the browsing pressure on forest regeneration in Tatra National Park. For Res Pap 78(3):238–247. https://doi.org/10.1515/frp-2017-0026
Brokaw N (1985) Gap-phase regeneration in a tropical forest. Ecology 66:682–687
Brzeziecki B, Kienast F (1994) Classifying the life-history strategies of trees on the basis of the Grimian model. For Ecol Manag 69:167–187
Buma B, Wessman CA (2011) Disturbance interactions can impact resilience mechanisms of forests. Ecosphere 2:64. https://doi.org/10.1890/ES11-00038.1
Buma B, Wessman CA (2012) Differential species responses to compounded perturbations and implications for landscape heterogeneity and resilience. For Ecol Manag 266:25–33. https://doi.org/10.1016/j.foreco.2011.10.040
Cameron AC, Trivedi PK (1990) Regression-based tests for overdispersion in the Poisson model. J Econ 46(3):347–364
Canham CD, Papaik MJ, Latty EF (2001) Interspecific variation in susceptibility to windthrow as a function of tree size and storm severity for northern temperate tree species. Can J For Res 31:1–10
Čater M, Diaci J (2017) Divergent response of European beech, silver fir and Norway spruce advance regeneration to increased light levels following natural disturbance. For Ecol Manag 399:206–212. https://doi.org/10.1016/j.foreco.2017.05.042
Cook JE (2000) Disturbance history of two natural areas in Wisconsin: implications for management. Nat Areas J 20:24–35
Diaci J (2017) Challenges for uneven-aged silviculture in restoration of post-disturbance forests in Central Europe: a synthesis. Forests 8:378. https://doi.org/10.3390/f8100378
Dixon PM, Weiner J, Mitchell-Olds T, Woodley R (1987) Bootstrapping the Gini coefficient of inequality. Ecology 68:1548–1551
Dobrowolska D (2015) Forest regeneration in north-eastern Poland following a catastrophic blowdown. Can J For Res 45:1172–1182
Ellenberg HH (1988) Vegetation ecology of Central Europe, 4th ed. C. U. Press
Faliński JB, Pawlaczyk P (1993) The outline of ecology. In: Białobok S (ed) Grab zwyczajny Carpinus betulus L. Nasze drzewa leśne [Hornbeam Carpinus betulus L. Our forest trees] – Sorus Poznań – Kórnik Vol. 9, pp 157–263 [in Polish with English summary]
Fichtner A, Sturm K, Rickert C, von Oheimb G, Härdtle W (2013) Crown size-growth relationships of European beech (Fagus sylvatica L.) are driven by the interplay of disturbance intensity and inter-specific competition. For Ecol Manag 302:178–184
Franklin JF, Mitchell RJ, Palik BJ (2007) Natural disturbance and stand development principles for ecological forestry. USDA Forest Service Northern Research Station General Technical Report NRS-19
Frelich LE (2002) Forest dynamics and disturbance regimes. Cambridge University Press, Cambridge
Gill NS, Jarvis D, Veblen TT, Pickett STA, Kulakowski D (2017) Is initial post-disturbance regeneration indicative of longer-term trajectories? Ecosphere 8:e01924. https://doi.org/10.1002/ecs2.1924
Greenberg CH, Collins BS (2015) Natural disturbances and historic range of variation: type, frequency, severity, and post-disturbance structure in Central Hardwood Forests. Springer
Herrera JM, García D (2009) The role of remnant trees in seed dispersal through the matrix: Being alone is not always so sad. Biol Conserv 142:149–158
Hilbe J (2014) Modeling count data. Cambridge University Press, Cambridge
Izdebski K, Czarnecka B, Grądziel T, Lorens B, Popiołek Z (1992) Plant communities against the background of the Roztocze National Park habitat conditions. Wydawnictwo UMCS, Lublin [in Polish with English summary]
Jaloviar P, Saniga M, Kucbel S, Pittner J, Vencurik J, Dovciak M (2017) Seven decades of change in a European old-growth forest following a stand-replacing wind disturbance: a long-term case study. For Ecol Manag 399:197–205. https://doi.org/10.1016/j.foreco.2017.05.036
Jankowska-Błaszczuk M, Grubb PJ (1997) Soil seed banks in primary and secondary deciduous forest in Bialowieza, Poland. Seed Sci Res 7:281–292
Johnson EA, Miyanishi K (eds) (2007) Plant disturbance ecology. Academic Press, San Diego
Kaszewski BM (2008) Climatic conditions of the Lublin region. Wydawnictwo UMCS, Lublin [in Polish with English summary]
Keren S, Diaci J, Motta R, Govedar Z (2017) Stand structural complexity of mixed old-growth and adjacent selection forests in the Dinaric Mountains of Bosnia and Herzegovina. For Ecol Manag 400:531–541. https://doi.org/10.1016/j.foreco.2017.06.009
Koop H (1987) Vegetative reproduction of trees in some European natural forests. Plant Ecol 72(2):103–110
Kuijper DPJ, Jędrzejewska B, Brzeziecki B, Churski M, Jędrzejewski W, Żybura H (2010) Fluctuating ungulate density shapes tree recruitment in natural stands of the Białowieża Primeval Forest, Poland. J Veg Sci 21:1082–1098
Kulakowski D, Veblen TT (2007) Effect of prior disturbance on the extent and severity of wildfire in Colorado subalpine forests. Ecology 88(3):759–769
Kupferschmid AD, Zimmermann S, Bugmann H (2013) Browsing regime and growth response of naturally regenerated Abies alba saplings along light gradients. For Ecol Manag 310:393–404
Leak WB (1987) Fifty years of compositional change in deciduous and coniferous forest types in New Hampshire. Can J For Res 17(5):388–393
Legendre P (2005) Species associations: the Kendall coefficient of concordance revisited. J Agric Biol Environ Stat 10(2):226–245
Lindenmayer DB, Burton PJ, Franklin JF (2012) Salvage logging and its ecological consequences. Island Press, Washington
Maciejewski Z, Szwagrzyk J (2011) Long-term changes in stand composition of natural forest stands on the Roztocze Highlands. Pol J Ecol 59(3):285–297
Marra DM, Chambers JQ, Higuchi N, Trumbore SE, Ribeiro GHPM, dos Santos J, Negron-Juarez RI, Reu B, Wirth C (2014) Large-scale wind disturbances promote tree diversity in the central Amazon forest. PLoS One 9(8):e103711
McClure JW, Lee TD (1993) Small-scale disturbance in a northern hardwoods forests: effects on tree species abundance and distribution. Can J For Res 23:1347–1360
Miścicki S, Żurek Z (2015) Monitoring of the deer-related damage to the young stands and regeneration in the Gorczański National Park. Sylwan 159(6):505–515 [in Polish with English summary]
Nagel TA, Svoboda M, Diaci J (2006) Regeneration patterns after intermediate wind disturbance in an old-growth Fagus-Abies forest in southeastern Slovenia. For Ecol Manag 226(1-3):268–278
Nagel T, Levanic T, Diaci J (2007) A dendroecological reconstruction of disturbance in an old-growth Fagus-Abies forest in Slovenia. Ann For Sci 64:891–897
Palik B, Kastendick D (2009) Woody plant regeneration after blowdown, salvage logging, and prescribed fire in a northern Minnesota forest. For Ecol Manag 258:1323–1330
Peterken GF (1996) Natural woodland. Cambridge University Press, Cambridge
Peterson CJ (2000) Damage recovery of tree species after two different tornadoes in the same old growth forest: a comparison of infrequent wind disturbances. For Ecol Manag 235:237–252
Rich RL, Frelich L, Reich PB (2007) Wind-throw mortality in the southern boreal forest: effects of species, diameter and stand age. J Ecol 95:1261–1273
Rutherford A (2001) Introducing ANOVA and ANCOVA: a GLM approach. SAGE Publications Ltd, London
Sabatini FM, Burrascano S, Keeton WS, et al (2018) Where are Europe’s last primary forests? Divers Distrib 1–14
Stadelmann G, Bugman H, Wermelinger B, Meier F, Bigler C (2013) A predictive framework to assess spatio-temporal variability of infestations by the European spruce bark beetle. Ecography 36:1208–1217
Szwagrzyk J, Szewczyk J, Maciejewski Z (2012) Shade-tolerant tree species from temperate forests differ in their competitive abilities: a case study from Roztocze, south-eastern Poland. For Ecol Manag 282:28–35
Szwagrzyk J, Gazda A, Dobrowolska D, Chećko E, Zaremba J, Tomski A (2017) Tree mortality after wind disturbance differs among tree species more than among habitat types in a lowland forest in northeastern Poland. For Ecol Manag 398:174–184
Szymkiewicz B (1971) Growth and yield tables for tree stands. PWRiL, Warszawa [in Polish]
Turner MG, Dale VH, Everham EH III (1997) Fires, hurricanes and volcanoes: comparing large disturbances. BioScience 47(11):758–768
Uriarte M, Canham CD, Thompson J, Zimmerman JK (2004) A maximum-likelihood, neighborhood analysis of tree growth and survival in a tropical forest. Ecol Monogr 74:591–614
Veblen TT (1986) Treefalls and the coexistence of conifers in subalpine forests of the central Rockies. Ecology 67(3):644–649
Veblen TT, Hadley KS, Nel EM, Kitzberger T, Reid M, Villalba R (1994) Disturbance regime and disturbance interactions in a Rocky Mountain subalpine forest. J Ecol 82:125–135
Vodde F, Jogiste K, Engelhart J, Frelich LE, Moser WK, Sims A, Metslaid M (2015) Impact of wind-induced microsites and disturbance severity on tree regeneration patterns: results from the first post-storm decade. For Ecol Manag 348:178–185. https://doi.org/10.1016/j.foreco.2015.03.052
Wolf A, Moller PF, Bradshaw RHW, Bigler J (2004) Storm damage and long-term mortality in a semi-natural, temperate deciduous forest. For Ecol Manag 182:197–210. https://doi.org/10.1016/j.foreco.2003.07.009
Woods KD (2004) Intermediate disturbance in a late-successional hemlock-northern hardwood forest. J Ecol 92:464–476. https://doi.org/10.1111/j.0022-0477.2004.00881.x
Woods KD (2008) Living long by staying small: stem layering as an adaptive life-history trait in shade-tolerant tree seedlings. Can J For Res 38:480–487. https://doi.org/10.1139/X07-136
Acknowledgements
We are also grateful to the authorities of the Roztoczański National Park for setting aside the wind-disturbed area for studying natural processes. We thank the two anonymous reviewers for the helpful comments that improved the quality of the paper.
Funding
This study was conducted as a part of research project 2012/07/B/NZ8/01908 “Patterns and drivers of the regeneration processes following catastrophic wind disturbances in forests” funded by the Polish National Science Foundation (NCN) and research project Poland N N304 048934 “Comparison of the dynamics of woody species and vascular plants in active and passive protected areas of the Roztoczański National Park” funded by the Ministry of Science and Higher Education of the Republic of Poland. This work was also partially supported by the Ministry of Science and Higher Education of the Republic of Poland (DS 3421/ZBL).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interests.
Additional information
Handling Editor: Andreas Bolte
Contribution of the co-authors JS and ZM designed the study, ZM and EM collected the field data, AG and AT conducted data analyses, and JS and AG wrote the text.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Szwagrzyk, J., Maciejewski, Z., Maciejewska, E. et al. Forest recovery in set-aside windthrow is facilitated by fast growth of advance regeneration. Annals of Forest Science 75, 80 (2018). https://doi.org/10.1007/s13595-018-0765-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13595-018-0765-z