
Abstract
Although honey bee brood does not need to seek shelter or food and restricts its movements to small wax cells, larvae have some degree of motility. Previously, other studies described how honey bee larvae showed analogous behaviours to the wandering period in holometabolous insects. The current research aimed to measure locomotion of the honey bee brood at different conditions of food supply and larval stadia. Besides, we developed an actometry assay to describe the larval behaviour under laboratory conditions. Our results suggested that the satiety and developmental program of larvae modulated their locomotion. Before they pupated, larval speed increased sharply and then it dropped until quiescence. However, starvation also induced an increase in angular velocity of brood. Starved larvae were between three and five times faster than the satiated ones. Moreover, fifth instars left their wax cells after 2 h of starvation without nurse bees. In the actometry assay, larvae showed behaviours of dispersion and changes in their kinematic parameters after detecting a tactile stimulus like the edge of arenas.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
As eusocial holometabolous insects, the honey bees (Apis mellifera L.) live within a nest as a colony with a single reproductive female (queen) and cooperative brood care among sisters (Winston 1987). In the darkness of the nest, chemical communication is crucial to take care of larvae. Brood pheromones and odour cues inform the presence of larvae, their developmental stage and food needs (Pankiw 2004). This complex brood rearing system is distinctive of certain eusocial hymenopteran species and termites (Wilson and Hölldobler 2005). However, other species of Hymenoptera and Coleoptera give also parental care to larvae (Wong et al. 2013). These behaviours evolved from an ancestral state of no brood care in the evolutionary history of holometabolous insects (Wong et al. 2013). Accordingly, the great majority of them are solitary species and consequently, the survival of brood depends on itself.
For this reason, locomotion and searching behaviours in most larvae are relevant traits allowing them to avoid predators and to seek food or shelter. The different kinds of locomotion in larvae are the consequence of different motor patterns which depend on the type of larva and the survival strategy (Brackenbury 2000; van Griethuijsen and Trimmer 2014). There are three main types of body design in insect larvae: oligopodous (grub-like), polypodous (caterpillar-like) and apodous (maggot-like) (Chapman 1998). Apodous larvae are legless, and most of them receive parental care or have a short foraging range. They belong to species of various orders as Diptera and Hymenoptera such as the honey bee.
An insect grows only during larval development as a consequence, a larva maximises its opportunities of feeding during the foraging period (Kause et al. 1999). Crawling in legless larvae is achieved by serial peristaltic contractions of muscles in the body wall starting posteriorly and continuing to the head (Brackenbury 2000; Heckscher et al. 2012; Simon et al. 2010; Quillin 1999). All these patterns, which occur because of motor programs defined in central pattern generators, are the product of rhythmic activity in the ventral nerve cord (Mantziaris et al. 2020; Lahiri et al. 2011). Although honey bee larva has numerous muscles, their role has been restricted by researchers to the ecdysis in moulting and the abdominal pumping in active ventilation ignoring a potential role in locomotion (Chapman 1998; Mänd et al. 2006; Oertel 1930). Nevertheless, solitary bees present locomotor and searching behaviours at larval stadia (Nakamatsu et al. 2006; Smirnoff 1960; Voigt and Gorb 2012).
Most insect pupae are immobile and vulnerable as a consequence larvae pupate in a cocoon or a shelter. The period after larvae cease the feeding and start the active seeking of a suitable site for pupation is called the wandering period (Ainsley et al. 2008). This is a stereotypic behaviour in which a dipteran larva can crawl as large distances as ten meters (Berrigan and Pepin 1995; Wang et al. 1997). In Drosophila and likely in the honey bee, the transition from foraging to wandering occurs by precisely timed developmental events which require coordination between the larval weight and gene expression (Callier and Nijhout 2013; Mirth and Riddiford 2007). This transition is fundamentally a change of innate response from attraction to aversion in the guided orientation to food-associated stimuli (Ainsley et al. 2008; Bell 1990; Gomez-Marin and Louis 2012; McIndoo 1929).
The study of larval locomotion and related behaviours has had great attention and development, particularly in D. melanogaster, in the last decades (Ainsley et al. 2008; Gerber and Stocker 2007; Heckscher et al. 2012; Loveless et al. 2019). However, little research has been focused on behaviours of honey bee larva, thus much remains to be done. In this sense, a few studies have measured and described how honey bee larvae move intensely and spin before pupation analogous to the wandering period in other insects (Jay 1963, 1964a). However, a honey bee larva does not need to seek shelter along large distances and restricts its movements inside a sealed cell in a period called spinning. Apart from that, Heimken et al. (2009) showed a brief experiment where the honey bee larva appeared to modify its motility depending on the duration of starvation.
Although larvae in brood frames are apparently immobile, they certainly have some degree of movement. Therefore, the current study aimed to describe and measure the motility of honey bee brood at different ages during larval development. For this purpose, we assessed the larval behaviour with video recordings under different environments (in a brood frame or in vitro) with or without nursing care. Furthermore, we developed a larval actometry assay to describe their locomotion and searching behaviour under laboratory conditions.
2 Materials and methods
2.1 Study site and animals
The experiments were performed from October to December during the spring season of the southern hemisphere (2016 and 2017). Female larvae (from cells of 5 mm of diameter) were monitored and sampled from three free-disease colonies (henceforth: A, B and C) of western honey bees (Apis mellifera ligustica Spinola) based on breeder information. These colonies were housed in Langstroth hives at the experimental apiary of the University of Buenos Aires, Argentina (34° 32′ S, 58° 26′ W).
2.2 Monitoring larval behaviour in a brood frame
2.2.1 Experiment 1: video-recording of nursing care of adults and locomotion of larvae
An empty frame was introduced in each source colony (A–C). Then, the queen was limited to lay eggs in this frame for 4 h to obtain a brood cohort with approximately known age (Vázquez and Farina 2020). The following day at the same time, once again the queen was limited to lay eggs in the same frame for 4 h. This procedure was repeated three more times. After 172 h since the empty frame was initially introduced, this was withdrawn from the hive to monitor it. The whole procedure was carried out twice in each colony. The brood frames generated with this procedure had larvae of various stadia and food in their periphery (honey and bee bread). All of them were very similar in their composition with approximately the same number of brood cells and storage cells. On average each brood frame contained around 800 storage cells (ratio of bee bread 1:3), 250 immature larvae (i.e., between stadium I and III) and 400 mature larvae (i.e., between stadium IV and V) on each side. The five stadia in the larval development were identified by their morphological traits (Human et al. 2013; Vázquez and Farina 2020). From each source colony, two brood frames were withdrawn: one fully covered with adult bees (larvae with nursing care and continuous feeding) and another without adult bees (larvae without nursing care and deprived of new food depositions). Both brood frames were monitored on different days but the same week inside an incubator (34 °C and 60% RH; chamber size: 49.5 × 30 × 30 cm). Here was video-recorded each brood frame with red light (LED A60, 5 W, 150 lm, wavelength: 660 nm). Adult honey bees are blind to light with wavelengths above 650 nm consequently the experiment was virtually in darkness to the insects (Avarguès-Weber et al. 2012). The monitoring was carried out for 3.5 h after 10 min of acclimatisation starting at 12:00 pm. Keep in mind that the experiment did not replicate the whole environment conditions inside a hive such as the attendance of the queen or the frequency of nursing visits. On average each brood frame with nursing care contained around 1000 adult honey bees with a ratio of brood to adults of approximately 1:1 (not all the adults were nurse bees). These conditions did not disrupt the rate of food supply during the experiment because food in the storage cells did not decrease significantly at the end. The video camera (Sony® HDR-CX260; video resolution: full HD 1920 × 1080 pixels, 50p) was positioned 13 cm above the chamber floor with 10 degrees of inclination. Besides, a front focal distance to the brood frame of 10 cm was set. The larvae at the base of the cells were digitally focused. An area of 80 × 56 cm (around 108 cells) in the middle of the brood frame was captured on one side. The brood in this area was stratified into the following age groups: young larvae (instars I and II, < 48 h old post-hatching), middle-aged larvae (instars III and IV ranged 48–96 h old post-hatching) and old larvae (instars V ranged 96–100 h old post-hatching). In each brood frame, 8 larvae of each age group were monitored randomly as focal individuals. Interactions between adult bees and larvae were classified by their duration but also with the body posture and movements of adult bees inside brood cells (Heimken et al. 2009; Huang and Otis 1991; Lindauer 1952; Siefert et al. 2021). On one hand, an interaction was classified as an inspection visit (short inspection < 10 s or long inspection > 10 s) when an adult bee entered partially into a brood cell with its abdomen on the outside. Frequently, an adult bee changed repeatedly its body position during an inspection with rotations. On the other hand, an interaction was classified as a feeding visit when an adult bee entered completely into a brood cell with its whole body remaining almost motionless. However, feedings were always preceded by a few seconds of inspection with changes in body position. Moreover, a new deposition of liquid food was usually observed in the bottom of the cell after the visit. Most of the time, a feeding visit lasted at least 10 s or more time (Brouwers et al. 1987). Nurse bees can also visit brood cells for thermoregulation, but larvae are mainly warmed at night and the incubator kept a steady optimum temperature during the experiment (Bujok et al. 2002; Lindauer 1954). In addition, dark combs were used for better visual contrast so adult bees did not build wax cells during the experiment. Nevertheless, other behaviours inside brood cells like resting or cleaning could not be excluded during long interactions. Therefore, the frequency and duration of the two main types of nursing visits (feeding and inspection) were approximately measured. Also, the duration of each inter-feeding period was measured in each focal larva. Finally, larvae within an unsealed cell mainly move in circles with transversal turns. Therefore, the angular velocity of each larva was quantified using its head as a reference point and concerning its centre of rotation. By convention, a positive angular velocity indicates counterclockwise rotation.
2.2.2 Experiment 2: quantification of searching behaviour in a brood frame
The procedure to obtain a brood cohort with approximately known age in the former experiment was repeated. Once again, a brood frame of each source colony (A–C) was withdrawn with the same properties as before. This was stored inside the same incubator in darkness, 34 °C and 60% RH at the same hour (12:00 pm). Unlike the earlier methodology, the whole brood frame was monitored on both sides with the naked eye every 5 min for 3.5 h. First, the number of larvae emerging partially outside their cells (at least their head) was counted. Then, the number of larvae leaving completely their cells and lying on the chamber floor was counted also. Besides, we identified their larval stadium by its morphological traits (Human et al. 2013; Vázquez and Farina 2020). As we stated in the former experiment, we monitored from each source colony a brood frame with nursing care and another without nursing care. Both brood frames were monitored on different days but the same week. For that, a metallic mesh was installed at the top entrance of the incubator. This was carried out to prevent adult bees from escaping the incubator when the door was opened. In each monitoring observation, larvae lying on the chamber floor were collected and weighed using an electronic balance (Mettler Toledo AG285, ± 0.1 mg).
2.3 Monitoring of larval behaviour in vitro
2.3.1 Experiment 3: video-recording of larval development in plastic cups
In the next experiment, honey bee larvae were reared in vitro because the end of larval development occurs hidden within sealed cells inside a hive. Hence, the conventional procedure of in vitro rearing was employed to study the motility of larvae during their complete development (10 days in vitro after hatching) (Crailsheim et al. 2013; Schmehl et al. 2016; Vázquez and Farina 2020). For that purpose, an empty frame was introduced in each source colony (A–C) and monitored for 8 h until the queen had laid enough eggs. Three days later, the brood frame was withdrawn and carried to a room with environmental conditions suitable for grafting. Here were grafted 21 first stadium larvae (0–8 h old post-hatching) from the brood frame to plastic cups with food (Crailsheim et al. 2013; Schmehl et al. 2016). Each larva was fed with one aliquot of 160 µl of food with unrestricted access. The composition of this food was: 6% D-glucose, 6% D-fructose, 1% yeast extract (provided by Sigma-Aldrich), and 50% commercial royal jelly (Kaftanoglu et al. 2011; Vandenberg and Shimanuki 1987). During the whole larval stage until pupation, the brood cohort was reared inside a sterile desiccator with a supersaturated salt solution to set humidity (Vázquez and Farina 2020). Then, this desiccator was kept inside an incubator (34.5 °C and ~ 90% RH). Only those larvae which survived to the whole larval development were monitored. The larvae were classified as dead when their pigmentation was unusual or lost cuticle turgor (Crailsheim et al. 2013; Schmehl et al. 2016). The brood cohort of each source colony was video-recorded for 10 days with the same video camera and red light of experiment 1. The video camera was positioned with a front focal distance to the plastic cups of 5 cm. The larvae at the base of the plastic cells were digitally focused. In each brood cohort, 8 larvae of each age group (see experiment 1) were monitored randomly as focal individuals. In each focal larva, its angular velocity was measured to compare with the behaviour of larvae in experiment 1. The angular velocity was quantified for 3.5 h in time intervals of 30 min defined by the mean duration of the inter-feeding period in experiment 1.
2.3.2 Experiment 4: larval actometry assay for honey bees
Actometer. An ad hoc video-recording device was developed for a novel actometry assay for honey bee larvae called henceforth the actometer. This device consisted of an incubator with a dark chamber treated with matte black damp-proofing paint (34 °C and 70% RH; chamber size: 61 × 42 × 46 cm). In addition, a removable plastic tray (45 × 45 cm) manufactured with black PLA was in the middle of the chamber floor. The tray could hold sixteen experimental arenas (glass Petri dishes of 10 cm Ø) in a setup of 4 × 4 arenas. This setup of arenas was recorded with a video camera located in the chamber roof with a front focal distance of 36.5 cm (Microsoft® LifeCam VX-1000 without its IR filter and with a wide-angle lens. Video resolution: 640 × 480 pixels). The arenas were illuminated with infrared light (eight 5-mm LEDs in each wall, wavelength: 940 nm). The LEDs were installed at the same level of the tray floor and parallel to the arena ground minimising reflections on the glass. Plastic Petri dishes or wax in the ground of arenas were not used because these materials reflect a great amount of infrared light. Therefore, the cuticle of larvae and the edge of arenas were the only objects in the actometer which reflected infrared light. The white larvae with black background in the video recordings simplified the computational tracking a posteriori. Lastly, the actometer had a ventilation system (flow rate: 60 m3/h) to prevent the accumulation of odours and carbon dioxide in the chamber and the experimental arenas.
Data acquisition. To carry out the assay, larvae were transferred into the arenas with careful handling and an adequate tool for each age as it was recommended in the study of Vázquez and Farina (2020) within a horizontal laminar flow. Larvae should lie in the centre of the arena with the same side facing up as they were in the wax cells or plastic cups. Previously, each larva was weighed using an electronic balance (Mettler Toledo AG285, ± 0.1 mg). The sixteen larvae inside the covered Petri dishes were located altogether with the tray into the actometer. And lastly, the larvae were manually focused on the ground of the arenas. The sixteen larvae in the tray were video-recorded simultaneously throughout 24 h starting at 12:00 pm. The larvae were weighed again at the end of the assay. In each video, larvae were analysed individually a posteriori with a tracking software ad hoc (see Supplementary). This software applied a motion mask to track the positions of a moving object inside the arena based on foreground detection. After this, the tracking software gave us an output file with positions of the centre point of the larva in the arena over time. Then, the data were normalised and the noise was removed from each signal (see Supplementary). With these denoised signals, different kinematic parameters were calculated for each larva describing its locomotor pattern and shape of its trajectory (see Supplementary). Brood was classified as dead when their colour changed to brownish at the end of the assay, their tracked area became zero during the assay or both.
Test and treatments. To describe the locomotion of brood according to larval age, a brood cohort of 60 larvae from each source colony (A–C) was reared in vitro. The conventional procedure of in vitro rearing was performed with standardised nutrition as experiment 3. Aside from this, larvae reared in the hives were also sampled because of the differences in pre-imaginal development depending on the rearing context (Vázquez and Farina 2020). For that purpose, after the grafting procedure in vitro, another group of 60 cells with newly hatching larvae in the same brood frame was monitored the following days. These procedures enabled us to obtain larvae with approximately known age. For each actometry assay, 8 larvae at the same age and stadium from both rearing contexts were sampled and video-recorded during 24 h. A video recording was carried out every day with new larvae during the larval development until 144 h of age post-hatching (Vázquez and Farina 2020). Finally, the last assay with 144-h-old larvae was video-recorded for 96 h until pupation. The large size of the arenas allowed us to identify changes in larval behaviour because of the stimulus of a vertical surface like the edge. Therefore, we analysed the kinematic parameters of larvae before, during and after to contact the edge. Sterile conditions were maintained during the in vitro rearing and the larval actometry assay to prevent bacterial or fungal infections and subsequent death.
2.4 Statistics
Data analysis and graphics were performed in R software (for details see Supplementary). All the variables were analysed with generalised linear models with or without mixed effects (GLMM or GLM) according to the experimental design. Data were fitted to negative binomial, beta, gamma or Gaussian distributions when it was appropriate. Kinematic parameters were also analysed with principal components analysis (PCA). Correlation tests were performed with the Pearson or Spearman’s rho coefficients depending on the normality of the data. Means were reported with their standard error (± s.e.m.) while the dispersion of data for each variable was reported with its coefficient of variation (CV). The alpha level was set at 0.05 and the P value was corrected with the Bonferroni procedure for multiple post hoc pairwise comparisons (Tukey test).
3 Results
3.1 Behavioural study of honey bee larva inside its cell
In the results of experiment 1 (Sect. 2.2), we measured the adult bee-larva interaction in random larvae inside of unsealed cells from three brood frames (colonies A-C) (Figure 1). These interactions were stratified according to age and developmental stage of the focal larvae in young brood (instars I and II, < 48 h old post-hatching), middle-aged brood (instars III and IV, 48–96 h old post-hatching) and old brood (instars V, 96–100 h old post-hatching). Because of the body postures and movements of the adult bees and the experimental conditions, these interactions were defined mainly as inspection and feeding visits. Young larvae could not be tracked in brood frames due to technical restrictions such as their smaller size, a different focus and less bright cuticle than other instars.

Larval behaviour was affected by the adult bee-larva interaction. Nursing interactions (inspection and feeding visits) were measured in random larvae from three colonies (A–C) inside unsealed cells of brood frames. Larvae were classified according to their age: instars III and IV in middle-aged group and instars V in old group. A The dispersion of the frequencies of nursing interaction per larva was plotted. Boxplots are displayed with their mean (square) and median (thick line). Bar plots were plotted for mean duration of B the feeding interaction and C the inter-feeding period (mean ± s.e.m.). The number of assessed larvae and p-values for each test are shown in the graphs. D The correlation between the cumulative time of nursing interaction with larval angular velocity among larvae is displayed. The best-fit lines are plotted with their confidence interval (95%) and different shapes per colony (orange for the middle-aged group and red for the old group). Regression equations: middle-aged group no significant correlation; old group, y = 0.01x + 1.51, R2 = 0.46
As a first remark, 56% of the monitored larvae showed locomotion in circles at the bottom of the wax cells with transversal turns in the clockwise direction. Meanwhile, the remaining half of larvae showed locomotion in the opposite direction. Furthermore, adult bees interacted with average middle-aged brood 1.5% of the whole recording time (3.5 h) and 5.1% with average old brood. The mean frequency of interaction was 8.63 ± 0.90 visits/h (CV = 51%) for middle-aged brood and 11.7 ± 0.98 visits/h (CV = 41%) for old brood. In this sense, adult bees visited significantly more frequent old brood than middle-aged brood [Figure 1a. GLM: count of visits ~ age group + colony + log(recording time); χ2 (2) = 17.38, P < 0.001, N = 48. Age group term: χ2 (2) = 5.76, P = 0.016]. The mean duration of the interactions was similar in middle-aged and old brood (middle age: 12.4 ± 3.3 s, CV = 130.5%. Old: 14.8 ± 4.3 s, CV = 141.2%).
If we discriminate the interactions between inspections or feedings, around 87% of them are to inspect larvae in both age groups. Then, the old brood with the greatest number of interactions (Figure 1a) also had the greatest number of inspections. Additionally, 90% of inspections for middle-aged brood and 86% of inspections for old brood were of short duration (< 10 s). Most of the long interactions (> 10 s) measured in middle-aged and old brood were for feeding (64 and 53%, respectively) and the remaining for long inspections.
The adult bees visited to feed average middle-aged and old brood a similar percentage of the total time of interaction (11.7 and 12.6%, respectively). In addition, the mean frequency of feedings was 1.38 feeding/h for middle-aged brood and 1.65 feeding/h for old brood. Meanwhile, the mean duration of the feeding interactions was shorter in middle-aged brood (32.6 ± 6.5 s, CV = 97.6%) than old brood (52.8 ± 7.7 s, CV = 71.3%) [Figure 1b. GLMM: duration of feeding interaction ~ age group + colony + (1|bee). Variance structure: 2.94% among bees. LR (5,8) = 20.69, P < 0.001, N = 43. Age group term: LR (7,8) = 15.46, P < 0.001]. Besides, there were no significant differences between age groups in the duration of the inter-feeding period (middle age: 17.47 ± 3.99 min, CV = 111.8%. Old: 23.29 ± 5.06 min, CV = 106.8%) [Figure 1c. GLMM: inter-feeding period ~ colony + (1|bee); Variance structure: < 1% among bees. χ2 (2) = 6.20, P = 0.045, N = 36. Age group term: χ2 (2) = 2.29, P = 0.130].
Lastly, a link between the larval behaviours and the nursing interactions was evaluated. Thus, a moderate positive correlation was found for old larvae between their angular velocity and the cumulative time of interaction with the adult bees [Figure 1d: Middle age: ρ = 0.04, S = 2205.3, P = 0.849, N = 24. Old: ρ = 0.57, S = 989, P = 0.004, N = 24].
Besides that, data acquired in experiments 1, 2 and 3 (Sects. 2.2 and 2.3) in combination enabled us to analyse if the locomotion of larvae was affected by their satiety, developmental program or both. Three conditions of food supply were tested for larvae from three colonies (A–C) inside wax cells or plastic cups: food ad libitum (larvae reared in vitro), food on request (larvae reared in a brood frame with nursing interaction) and food deprivation (larvae reared in a brood frame without nursing interaction). Besides, the larval behaviour was stratified in groups according to the age of focal larvae throughout the two main larval periods: feeding period and post-feeding/spinning period (Figure 2).

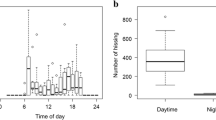
The locomotor pattern of honey bee larvae inside brood cells was measured. Larval behaviour according to food supply and age group was analysed. Food supply: ad libitum (larvae reared in vitro), on request (larvae reared in a brood frame with nursing interaction) and deprivation (larvae reared in a brood frame without nursing interaction). Age group: instars I and II (young group), instars III and IV (middle-aged group), instars V during feeding period/unsealed cell (old group), instars V during post-feeding period/sealed cell before defecation (early spinning: S1) and after defecation (late spinning: S2). Mean angular velocity of larvae from three colonies (A–C) inside wax or plastic cells was measured per age group according to A the food supply and B the developmental stage. Bars indicate means ± s.e.m. The number of assessed larvae is shown in the graphs. Groups with different letters had significantly different means (P < 0.05). Multiple pairwise comparisons were performed with Tukey test. C The cumulative number of larvae outside their wax cells in brood frames from three colonies (A–C) was plotted over 3.5 h according to the food supply. Stacked bars per colony are displayed with white colour for larvae partially outside their cells and grey for larvae completely outside their cells
The results showed that the larval behaviour of early instars was independent of the frequency of food supply, in other words with an invariable mean angular velocity. Also, in all instars (I–V) with ad libitum food, larvae showed transversal turns with the same mean angular velocity which ranged from 1.16 to 1.57°/min regardless of age (Figure 2a and S1). Meanwhile, old larvae increased significantly their angular velocity when there was a growing restriction in the food supply. In food deprivation, the mean angular velocity of the transversal turns was 6.61 ± 0.54°/min (CV = 40.3%) for old larvae [Figure 2a; GLMM: angular velocity ~ age group + food supply + colony + age group × food supply + (1|bee). Variance structure: 8.65% among bees. LR (5,12) = 114.91, P < 0.001, N = 144. Post hoc pairwise comparisons were performed with Tukey test: Table S1]. In this circumstance, old brood showed a gradual increase of their mean angular velocity over time reaching a maximum value after 100 min of food deprivation, i.e., 4–6 times an average inter-feeding period (Figure S1). Also, old larvae showed signs of fatigue around 9–10 times an average inter-feeding period.
However, the mean angular velocity rose sharply when we compared in the same fifth stadium the two different developmental periods, feeding and spinning (Figure 2b). At the beginning of the behavioural program of spinning, the old larvae showed a high increase of their mean angular velocity (22.89 ± 1.91°/min, CV = 40.9%) [Figure 2b. GLMM: angular velocity ~ age group + colony + (1|bee). Variance structure: 11.39% among bees. LR (8,14) = 92.39, P < 0.001, N = 120. Post hoc pairwise comparisons were performed with Tukey test: Table S2]. Besides that, in the larval locomotion during the spinning period predominated the transversal turns interrupted by a few longitudinal turns in the plastic cells. These longitudinal turns enabled larvae to change their direction of rotation from clockwise to counter-clockwise or vice versa. Then, the larvae decreased their motility until quiescence when they started to defecate (Figure S1).
Lastly, when larvae in brood frames were isolated without nursing interaction and with food deprivation, certain individuals showed increasing locomotion and movements outside their wax cells (Figure 2c). These behaviours implied ultimately longitudinal movements inside wax cells. A significant increase over time in the cumulative number of larvae emerging partially outside their cells was measured in food deprivation after 80 min (4 times an average inter-feeding period) with a peak in number around 140 min. Furthermore, after 120 min, these larvae started to leave completely their cells. The proportion of larvae laid on the chamber floor of the incubator was growing over time [Figure 2c. GLMM: count of larvae ~ food supply + behaviour + colony + (1|time). χ2 (2) = 267.26, P < 0.001, N = 147. Zero-inflated term (~ food supply): χ2 (1) = 13.18, P = 0.001. Random term (~ time): χ2 (1) = 300, P < 0.001. Food supply term: χ2 (2) = 151.03, P < 0.001].
No association was found between the time of escape from wax cells and the weight of larvae which left their cells [ρ = 0.18, S = 20,228, P = 0.186, N = 53]. Nevertheless, all individuals were identified as instars V with weights greater than 140 mg approximately. It is important to note that less than 1% of all monitored brood cells of the fifth instars (100-h-old larvae) were sealed during the experiment in the group with nursing interaction. At the same time, around 12% of all monitored brood cells of the fifth instars without nursing interaction presented 100-h-old larvae which moved partially or completely outside their cells.
3.2 Behavioural study of larval locomotion with an actometry assay
The locomotor pattern of larvae from three colonies (A–C) reared in vitro or a hive was tracked inside Petri dishes at different ages during starvation in experiment 4 (Sect. 2.3). The early instars (I, II and III) did not survive (0%) to the 24 h of testing with food deprivation. They were immobile in the starting point in the middle of arenas. Meanwhile, the remainder instars (IV and V) tolerated completely the whole testing and showed locomotion. Despite there was a high survival at the end of the assay regardless of age or rearing context (more than 92%), fourth instars reared in a hive showed a little lower survival (68.2%) than the others (Figure S2).
At the beginning of the assay, larvae were motionless with latency to start the locomotion which decreased significantly regarding age [Figure 3a. GLM: Box-Cox(latency) ~ age. F (3,182) = 6.57, P < 0.001, N = 187. Rearing context term: F (1,182) = 2.86, P = 0.093. Post hoc pairwise comparisons were performed with Tukey test: Table S3]. Although latency was not significantly different between rearing contexts, there was a trend to start the locomotion early in larvae reared in vitro. If we compare the results of experiment 1 and the mean latency of larvae reared in a hive, we will find that the larval behaviour belonging to the old brood matches in both experiments. In 120-h-old larvae, the mean latency was 29.32 ± 9.23 min (CV = 154.1%). In other words, their latency lasted around one average inter-feeding period. Meanwhile, in 96-h-old larvae, the mean latency was 54.08 ± 18.78 min (CV = 166.6%). In other words, their latency lasted around 3 times the average inter-feeding period.

Honey bee larvae showed locomotor and searching behaviours in the actometry assay. Larvae reared in vitro (iv) or in a hive (H) from three colonies (A–C) were sampled at different ages (hours post-hatching). Then, the locomotor pattern of larvae was tracked inside Petri dishes in an assay with controlled environmental conditions. Brood in feeding period (F) were tracked over 24 h in each age (72-h-old larvae in stadium IV, 96-h-old and 120-h-old larvae in stadium V). Meanwhile, larvae in spinning period (S, 144-h-old larvae in stadium V) were tracked over 48 h. A Mean latency to start locomotion and B mean cumulative displacement inside the arena were plotted according to rearing context and age. Bars indicate means ± s.e.m. The number of assessed larvae and P values for each test are shown in the graphs. Groups with different letters had significantly different means (P < 0.05). Multiple pairwise comparisons were performed with Tukey test. C The correlation between initial larval weight with cumulative displacement among larvae is displayed. The best-fit lines are plotted with their confidence interval (95%), different shapes per colony and different colours per rearing context and developmental stage. Regression equations: S + H and S + iv no significant correlations; F + H, y = 2.32x, R2 = 0.10; F + iv, y = 5.12x + 133.25, R2 = 0.25
The tested larvae (instars IV and V) showed movements inside the arena with transversal turns generating spiral trajectories with loops (Figure S3–S4). Sometimes, they also showed straight movements in short trajectories. In any case, the larvae always presented signs of dispersal with random orientation regardless of their age or rearing context (Figure S5). Furthermore, the cumulative displacement inside the arena showed a bell-shaped distribution according to age with a significant interaction between rearing context and age [Figure 3b. GLMM: cumulative displacement ~ age + rearing context + colony + age group × rearing context + (1|bee). Variance structure: < 1% among bees. LR (13,24) = 221.14, P < 0.001, N = 232. Post hoc pairwise comparisons were performed with Tukey test: Tables S4, S5]. Therefore, larvae reared in a hive showed a maximum displacement around 144 h of age post-hatching in concordance with the spinning period. Meanwhile, larvae reared in vitro overtook in-hive larvae with a peak around 120 h of age post-hatching. However, there were no significant differences between both peaks in the cumulative displacement with 763.54 ± 57.52 mm (CV = 36.9%) for the in-hive larvae and 776.76 ± 55.54 mm (CV = 35%) for the in vitro larvae.
If the cumulative displacement is relativized with the time of displacement and the larval weight, a steady decrease in locomotion can be noticed throughout the larval development with a trend to quiescence. Moreover, this pattern was more gradual for the in vitro larvae (Figure S6). Subsequently, we evaluated the relationship between the magnitude of the total displacement and the locomotor capability (biomass as an indicator) of larvae. Thus, a moderate positive correlation was found for larvae in the feeding period between the total cumulative displacement and the larval weight regardless of rearing context [Figure 3c: F + H: ρ = 0.37, S = 8330, P = 0.015, N = 67. F + iv: ρ = 0.56, S = 65,335, P < 0.001, N = 72. S + H: ρ = 0.06, S = 1069, P = 0.800, N = 47. S + iv: r = 0.36, t = 1.81, P = 0.084, N = 46]. Therefore, the magnitude of the larval activity was not associated with the weight of larvae during the spinning period. In addition, the proportion of weight loss in tested larvae was about the same on average among ages and rearing contexts at the end of the actometry assay (Table S6). Nevertheless, a positive low correlation was detected between the weight loss and the magnitude of the locomotion during the spinning period regardless of rearing context [S + H: ρ = 0.51, S = 846, P = 0.014, N = 24. S + iv: r = 0.47, t = 2.41, P = 0.026, N = 22. F + H: ρ = 0.14, S = 47,094, P = 0.252, N = 67. F + iv: ρ = − 0.19, S = 62,206, P = 0.126, N = 68].
The locomotor patterns of larvae were described with different kinematic parameters measured in the trajectories inside the arenas (Table S7). Then, we performed a PCA with these parameters (Table S8). A single principal component (PC1) achieved 71% of the cumulative proportion of deviation. In other words, PC1 explained the different locomotor patterns easily distinguishable in our data with clusters associated with larval age (Figure S7). The brood during the spinning period showed locomotor patterns very similar to each other. Meanwhile, the starved brood before to spin showed more variable locomotor patterns. However, PC1 did not strongly correlate with any kinematic parameter. Therefore, none of the kinematic parameters provided redundant information and all of them partially explained the patterns and the total variance in our data. Additionally, it was not possible to differentiate between the locomotor patterns of larvae reared in vitro or a hive.
Finally, we analysed changes in locomotion due to the contact with the edge (Figure S3–S4). Only fifth instars contacted the edge after 24 h. Furthermore, they never touched the edge of the arena before 3 h. Besides, the number of those instars was the highest around the larval age with the maximum locomotor activity (Figure 4a). There was no significant difference for the delay of larvae to contact the edge in the age of maximum activity between brood from both rearing contexts. Whereas in-hive larvae showed a delay of 6.77 ± 0.75 h (CV = 54.1%), in vitro larvae showed a delay of 6.32 ± 0.7 h (CV = 54.4%). These delays decreased according to an increasing larval age [Figure 4a. GLM: delay ~ age + rearing context + age × rearing context. F (5,75) = 4.54, P = 0.001, N = 81. Post hoc pairwise comparisons were performed with Tukey test: Table S9].

Honey bee larva showed thigmotaxia and thigmokinesis in the actometry assay. Locomotion of brood located in the middle of Petri dishes (10 cm Ø) was tracked in the actometry assay. Larvae reared in vitro or in a hive from three colonies (A–C) were sampled at different ages (hours post-hatching). Brood in feeding period were tracked over 24 h in each age (72-h-old larvae in stadium IV, 96-h-old and 120-h-old larvae in stadium V). Meanwhile, larvae in spinning period (144-h-old larvae in stadium V) were tracked over 48 h. A We measured the delay of larvae to contact the edge of the arena during the test (24 h). Then, we calculated B the coefficient of angular dispersion, C linear velocity and D turning frequency of the fifth stadium larvae reared in vitro. These variables were plotted before, during and after to contact the edge of the arena. Bars indicate means ± s.e.m. Boxplots are plotted with their mean (square) and median (thick line). The number of assessed larvae and P values for each test are shown in the graphs. Groups with different letters had significantly different means (P < 0.05). Multiple pairwise comparisons were performed with Tukey test
Changes in locomotion of in vitro larvae were assessed before, during and after they contacted the edge of the arena as a potential tactile stimulus. This group was analysed because it had the largest number of larvae which contacted the edge. The change due to the stimulus was compared between the two stages in the larval development: feeding period (120-h-old larvae) and spinning period (144-h-old larvae). Average brood in both developmental stages spent similar proportions of the testing time edging the arena. The 120-h-old larvae spent 29.9% of the 24 h while the 144-h-old larvae spent 21.5%. In this sense, we observed only silk (clear secretion) and faeces (brown secretion) in the Petri dishes of the 144-h-old larvae which secreted mainly near the edge.
Those larvae which contacted the edge showed a high directionality moving away from the centre of the arena. Their coefficients of angular dispersion were above 0.8 in both ages (Figure 4b, S4). A coefficient around 1 means a concentration of the polar angles for the positional vectors. Furthermore, a significant increase in the directionality was detected during and after to contact the edge [Figure 4b. GLMM: coefficient of angular dispersion ~ age + contact with edge + age × contact with edge + (1|bee). Variance structure: 1.94% among bees. χ2 (4) = 40.67, P < 0.001, N = 109. Post hoc pairwise comparisons were performed with Tukey test: Table S10]. This effect was observed in the trajectories of larvae like a continuous movement in contact with the edge which was a sign of thigmotaxis (Figure S4, S8). Following the change in the directionality, also a change in the linear velocity and turning frequency were detected during and after the larvae contacted the edge (Figure 4c, d). These were signs of thigmokinesis due to the tactile stimulus of the edge. A significant increase in the mean linear velocity (35.5% more than before the edge) and a significant decrease in the mean turning frequency (89% less than before the edge) were measured regardless of age [Figure 4c. GLMM: linear velocity ~ contact with edge + (1|bee). Variance structure: 27.44% among bees. χ2 (2) = 32.33, P < 0.001, N = 109. Age term: χ2 (1) = 0.07, P = 0.791. Post hoc pairwise comparisons were performed with Tukey test: Table S11] [Figure 4d. GLMM: turn ~ contact with edge + log(time in displacement) + (1|bee). Variance structure: < 1% among bees. χ2 (2) = 55.09, P < 0.001. Zero-inflated term (~ contact with edge): χ2 (3) = 27.7, P < 0.001. Age term: χ2 (1) = 0.58, P = 0.446. Post hoc pairwise comparisons were performed with Tukey test: Table S12].
4 Discussion
4.1 Developmental program and food supply modulate larval behaviour
The honey bee larvae displayed locomotor behaviours inside different sites and surfaces such as a wax or plastic cell and even in a glass Petri dish before the spinning period. In these environments, larvae mainly moved in circles with transversal turns at very low speed generating circular or spiral trajectories even without food after several hours. The typical crawling in an apodous larva was recognised as their motor pattern when accelerated video recordings were observed (Brackenbury 2000; Heckscher et al. 2012; Quillin 1999; Simon et al. 2010). Moreover, the larvae increased gradually their speed with growing restrictions in the food supply. In food deprivation, fifth instars reached a maximum speed after 100 min when there was not nursing care. Indeed, 12% of the fifth instars in the brood frames in starvation left their cells after 2 h. These results are very consistent with the experiment of Heimken et al. (2009) with larvae in starvation. Therefore, the locomotor pattern of honey bee larvae depends apparently on the intensity of nursing interaction when adult bees give them food.
On one hand, old larvae with nursing care showed a mean angular velocity of 5°/min inside their cells before the sealing. This speed was measured when they had a mean frequency of interaction with adult bees of 11.7 visits/h, a mean frequency of feedings of 1.7 visits/h and a mean duration of feedings of 52.8 s. On the other hand, middle-aged larvae showed a mean angular velocity of 1.9°/min. This speed was measured when they had a mean frequency of interaction of 8.6 visits/h, a mean frequency of feedings of 1.4 visits/h and a mean duration of feedings of 32.6 s.
Furthermore, we observed acceleration in the speed of larvae during a feeding visit and some minutes after, looking at the position of the larva pre- and post-interaction. This initial acceleration is consistent with the observations of Siefert et al. (2021) where larvae mainly move if they were not fed close to their mouth. Hence, the larvae exhibit a searching behaviour in which they move to reach and eat new food depositions. Nevertheless, the larvae showed continuous transversal turns during inter-feeding periods with differences in speed among stadia. These differences could be a consequence of different frequency and duration of nursing interactions, changing properties of larval food, inherent behavioural or physiological attributes of each stadium or all of them.
The differences in the nursing interaction among larval stadia were observed in our experiment and previous studies with observation beehives or small colonies (Brouwers et al. 1987; Heimken et al. 2009; Huang and Otis 1991; Siefert et al. 2020). In all of these studies, adult bees visited more frequent old brood than middle-aged brood. Besides, old brood had the greatest number of inspections which were mainly of short duration (< 10 s). Then, the mean frequency of feedings in our experiment is coherent with the previous studies (Brouwers et al. 1987; Heimken et al. 2009; Huang and Otis 1991; Lindauer 1952). Moreover, the mean duration of feedings for old brood in our experiment is coherent with previous results (Brouwers et al. 1987; Huang and Otis 1991). However, the frequency of the total nursing visits in our experiment was ten times smaller than the result of Siefert et al. (2021). Also, the mean feeding duration of the middle-aged brood in our experiment is shorter than the result of Brouwers et al. (1987). Furthermore, both studies stated an inverse relationship between the larval age and the duration of feeding visits unlike our results (Brouwers et al. 1987; Siefert et al. 2020).
Those differences could be explained by different experimental environments. While those studies carried out their assays with observation hives, we used isolated brood frames inside an incubator. Each method has its pros and cons. Observation hives with brood cells monitored laterally with a longitudinal cut hinder recordings of larval movements. Meanwhile, in an incubator, the volatile hunger signals of starved larvae could be diluted (Pankiw 2004). Besides that, the number of nurse bees and the ratio of brood:nurse bees in our experiment were smaller in comparison with experiments performed with small colonies. Nevertheless, a higher number of adult bees hinders the monitoring of larvae. Moreover, nurse bees are not all the same and depending on their age and glandular development, they take care of different age groups of larvae as Brouwers et al. (1987) indicated. We do not know the ratio of each type of nurse bees in our and their experiments. These reasons could explain the different patterns in the duration of feedings and the low frequency of inspections. Beyond these aspects, our description allows us to relate the observed larval behaviours with physiological or environmental factors in our specific conditions. In this sense, a positive correlation was found only in old larvae between their angular velocity and the cumulative time of interaction with the adult bees.
The occurrence or not of that correlation according to larval age could be explained by the switch in larval food composition (Brouwers et al. 1987; Shuel and Dixon 1959). Although the different compounds ingested by fifth instars may induce behavioural changes in themselves, larvae may also experience physiological changes in each stadium as well as morphological ones. Besides that, the increase in the mean angular velocity of older larvae may occur because of an increase in the amount of ingested carbohydrates when larvae switch from worker jelly to modified worker jelly with honey and beebread. In this sense, under in vitro conditions, larvae were fed ad libitum with a unique diet throughout all stadia. These brood moved constantly in circles turning around the plastic cells with a low steady angular velocity. They lasted approximately from 3 to 5 h in complete one turn (360°) regardless of their age. Thus, these satiated brood at the top of the food drop exhibits slow locomotion which is linked with feeding and not with seeking.
The carbohydrate concentration of the in vitro diet is constant and similar to the worker jelly which is supplied to young and middle-aged larvae (Kaftanoglu et al. 2011). Consequently, all stadia of in vitro larvae ingested growing amounts of food and carbohydrates but in a much smaller quantity for the fifth instars than those reared in a hive. Thus, if we relativize the angular velocity of each in vitro instar with their weight associated with food intake, the relative angular velocity decreases according to age. Then, motility dropped until quiescence before pupation according to the developmental program (Myser 1954; Jay 1963, 1964a; Oertel 1930). Therefore, the inverse relationship between speeds and amount of ingested carbohydrates indicates a physiological cause apart from a nutritional one (Quillin 1999).
Up to this point, the locomotion of honey bee larvae seems to be related to the larval age and the presence of food. However, in brood frames without nursing care, old larvae with food deprivation increased their mean angular velocity 33% more than brood with nursing interaction. Certainly, they did not move to reach a distant deposition of food in deprivation. Thus, larvae are always moving even without interaction with adult bees or new food depositions. This is also confirmed in the larval actometry assay when the fourth and fifth instars explored an empty Petri dish. In addition, larvae moved with the same motor pattern in circles on the ground of the cells or arenas just as they did when were fed by nurse bees.
It is important to mention that larvae showed a different pattern of movements with longitudinal turns, secretions and excretions when they are spinning (Jay 1964a). These behavioural changes in fifth instars are a consequence of the developmental program which is physiologically modulated by larval weight (Callier and Nijhout 2013; Mirth and Riddiford 2007). The growth rate is similar among larvae of the same cohort reaching the threshold weight at a similar age when they also start to release capping pheromones (Qin et al. 2019). These pheromones induce nurse bees to seal brood cells. In our experiment in brood frames, the fifth instars with nursing care showed less than 1% of sealed cells during the monitoring. In this sense, fifth instars of all brood frames with or without adult bees had approximately the same age around 96–100 h post-hatching before the average sealing time around 120 h. Consequently, larvae with food deprivation were not in the average age to start the spinning period.
Larvae reared in a hive do not start immediately the spinning period after the sealing. On average larvae started to spin between 168 and 252 min after they contacted the seal which they use as a reference during the longitudinal turns and then to defecate (Jay 1964a). Our results showed starved fifth instars in unsealed cells that started to leave their cells after 80 min with a peak in number around 140 min. Although underfeeding fifth instars spin early after an interruption of food supply as detailed Jay (1964b), it is strange that the spinning behaviour was induced by food deprivation in such a short time in all our current experiments. Furthermore, this behaviour of dispersion was observed a day before the average sealing time and even in fourth instars in the actometry assay. Hence, we suggest a potential role of hunger in the modulation of locomotion.
4.2 Satiety and sensory inputs could modulate larval locomotion
In general, the metabolic state of an organism affects its locomotor activity which increases with food deprivation (Klowden 2007). Different neuroendocrine signals of satiety and hunger regulate motor patterns associated with feeding and food seeking in insects (Browne 1993; Saraswati et al. 2004). In the case of honey bees, brood are capable of begging for food (He et al. 2016; Heimken et al. 2009; Huang and Otis 1991). Starved larvae signal actively their food needs to nurse bees increasing the production of volatile signals such as E-β-ocimene. Genes of the biosynthetic pathway of E-β-ocimene were upregulated in larvae reaching a peak after 30 min of starvation (He et al. 2016). This period agreed with our findings in the mean duration of the inter-feeding period of 20.6 min regardless of larval age. After 2 h, starved larvae are visited significantly more frequent than satiated ones (Huang and Otis 1991), when isolated and starved larvae in our experiment started to leave their cells. Young larvae produced more hunger signals than middle-aged and old larvae after the same time of food deprivation (He et al. 2016). This might be the reason for an increasing frequency of inspections according to larval age because the begging maybe is less intense in old larvae.
The association between satiety and locomotion could be an evolutionary relict of the searching behaviour that the solitary hymenopteran ancestor of honey bees exhibited to survive as most insect larvae (Wilson and Hölldobler 2005; Wong et al. 2013). This behavioural trait may currently have a function or not. A possibility would be that locomotion acts like an enhancer of begging in old brood after reaching the maximum biosynthesis of the pheromone. The increase of amplitude, frequency or both in muscular contractions would facilitate the secretion of the hunger signal as a hypothetical mechanism. Other soft-bodied larvae, such as caterpillars and Drosophila, increase speed primarily by increasing stride frequency (Casey 1991; Heckscher et al. 2012; Quillin 1999). Apodous and polypodous larvae produce similar stride lengths during the crawling of around 25% of their body length (Brackenbury 2000). This compression of the cuticle could affect the integument glands and their secretions.
In early stadia, the locomotion was independent of satiety. This would be caused by a combination of a high begging capability and a high energy cost for crawling (Berrigan and Pepin 1995; Casey 1991). Conversely to the older larvae, brood in early stadia is very susceptible to starvation (He et al. 2016; Wang et al. 2016). In this sense, the latency to start movements in Petri dishes was shorter for older larvae. Moreover, the fourth and fifth instars exhibited locomotor patterns of increasing dispersal with random orientation inside the Petri dishes. The dispersal behaviour serves to increase the probability of finding a food source or a shelter by expanding the search area (Bell 1990; Ainsley et al. 2008; Gomez-Marin and Louis 2012). In fifth instars this searching behaviour could be initially to find food, but we cannot dismiss later an early spinning (Jay 1964b).
An average 120-h-old larva in one day could move up to half a meter inside a Petri dish. Nevertheless, they spent most of the time in rest (72%). The remaining time, the larvae moved with a low mean linear velocity of 0.75 mm/min. D. melanogaster, another apodous larva, also showed a low mean speed of 5.3 mm/min during the foraging period unlike the caterpillars (Berrigan and Pepin 1995; Heckscher et al. 2012; van Griethuijsen and Trimmer 2014). Meanwhile, an average 144-h-old larva in spinning state exhibited a greater explored distance up to one meter in 24 h, less time of resting (47%) and a higher mean linear velocity of 1.02 mm/min than younger fifth instars. Besides, the high angular velocity during the spinning in plastic cells agrees with previous studies (Jay 1964a; Siefert et al. 2021). All these characteristics are consistent with the wandering state of other insect larvae (Ainsley et al. 2008; Berrigan and Pepin 1995; Wang et al. 1997).
Finally, the fifth instars showed signs of positive thigmotaxis and thigmokinesis regardless of larval age. Indeed, honey bee larvae have shown orientation during spinning based on tactile stimuli of the seal and cell walls (Jay 1963, 1964a). All these results suggest mechanoreception in honey bee larvae. We must keep in mind that sensory feedbacks and neuroendocrine control lead insect larvae to change among different strategies of locomotion during development (Browne 1993; Dominick and Truman 1983; Gomez-Marin and Louis 2012; Saraswati et al. 2003).
4.3 Conclusion
The honey bee brood during the larval stage showed locomotion and searching behaviours which were modulated by the satiety, developmental program and tactile stimuli. Moreover, a relationship between locomotion and begging of larvae during starvation appears a feasible hypothesis. Lastly, the larval actometry assay developed in the current study was a helpful tool to analyse the locomotor patterns of larvae under controlled environmental conditions.
Supplementary information.
Data Availability
Data will be available on reasonable request.
References
Ainsley, J.A., Kim, M.J., Wegman, L.J., Pettus, J.M., Johnson, W.A. (2008). Sensory mechanisms controlling the timing of larval developmental and behavioral transitions require the Drosophila DEG/ENaC subunit, Pickpocket1. Dev. Biol. 322(1), 46-55.
Avarguès-Weber, A., Mota, T., Giurfa, M. (2012). New vistas on honey bee vision. Apidologie, 43(3), 244-268.
Bell, W.J. (1990). Searching behavior patterns in insects. Annu. Rev. Entomol. 35(1), 447-467.
Berrigan, D., Pepin, D.J. (1995). How maggots move: allometry and kinematics of crawling in larval diptera. J. Insect Physiol. 41(4), 329-337.
Brackenbury, J. (2000). Locomotory modes in the larva and pupa of Chironomus plumosus (Diptera, Chironomidae). J. Insect Physiol. 46(12), 1517-1527.
Browne, L.B. (1993). Physiologically induced changes in resource-oriented behavior. Annu. Rev. Entomol. 38(1), 1-23.
Brouwers, E.V.M., Ebert, R., Beetsma, J. (1987). Behavioural and physiological aspects of nurse bees in relation to the composition of larval food during caste differentiation in the honeybee. J. Apic. Res. 26(1), 11-23.
Bujok, B., Kleinhenz, M., Fuchs, S., Tautz, J. (2002). Hot spots in the bee hive. Naturwissenschaften. 89(7), 299-301.
Callier, V., Nijhout, H.F. (2013). Body size determination in insects: a review and synthesis of size‐and brain‐dependent and independent mechanisms. Biol. Rev. 88(4), 944-954.
Casey, T.M. (1991). Energetics of caterpillar locomotion: biomechanical constraints of a hydraulic skeleton. Science. 252(5002), 112-114.
Chapman, R.F., (1998). The insects: structure and function. Cambridge university press.
Crailsheim K., Brodschneider R., Aupinel P., Behrens D., Genersch E., et al. (2013). Standard methods for artificial rearing of Apis mellifera larvae. J. Apicult. Res., 52(1): 1-16.
Dominick, O.S., Truman, J.W. (1984). The physiology of wandering behaviour in Manduca sexta. I. Temporal organization and the influence of the internal and external environments. J. Exp. Biol. 110(1), 35–51.
Gerber, B., Stocker, R.F. (2007). The Drosophila larva as a model for studying chemosensation and chemosensory learning: a review. Chem. Senses. 32(1), 65-89.
Gomez-Marin, A., Louis, M. (2012). Active sensation during orientation behavior in the Drosophila larva: more sense than luck. Curr. Opin. Neurobiol. 22(2), 208-215.
He, X.J., Zhang, X.C., Jiang, W.J., Barron, A.B., Zhang, J.H., et al. (2016). Starving honey bee (Apis mellifera) larvae signal pheromonally to worker bees. Sci. Rep.-UK. 6(1), 1-9.
Heckscher, E.S., Lockery, S.R., Doe, C.Q. (2012). Characterization of Drosophila larval crawling at the level of organism, segment, and somatic body wall musculature. J. Neurosci. 32(36), 12460-12471.
Heimken, C., Aumeier, P., Kirchner, W.H. (2009). Mechanisms of food provisioning of honeybee larvae by worker bees. J. Exp. Biol. 212(7), 1032-1035.
Huang, Z.Y., Otis, G.W. (1991). Inspection and feeding of larvae by worker honeybees (Hymenoptera: Apidae): effect of starvation and food quantity. J. Insect Behav. 4(3), 305-317.
Human, H., Brodschneider, R., Dietemann, V., Dively, G., Ellis, J., et al. (2013). Miscellaneous standard methods for Apis mellifera research. In V. Dietemann; J.D. Ellis; P. Neumann (Eds). The COLOSS BEEBOOK, Volume I: standard methods for Apis mellifera research. J. Apicult. Res. 52(4): https://doi.org/10.3896/IBRA.1.52.4.10
SC Jay 1964a The Cocoon of the Honey Bee, Apis mellifera L Can. Entomol. 96 5 784 792
SC Jay 1964 Starvation studies of larval honey bees Can. J. Zool. 42 3 455 462
Jay, S.C. (1963). The longitudinal orientation of larval honey bees (Apis mellifera) in their cells. Can. J. Zool., 41(5), 717-723.
Kaftanoglu, O., Linksvayer, T.A., Page, Jr R.E. (2011). Rearing honeybees, Apis mellifera, in vitro I: Effects of sugar concentrations on survival and development. J. Insect Sci. 11(1):96.
Kause, A., Saloniemi, I., Haukioja, E., Hanhimäki, S. (1999). How to become large quickly: quantitative genetics of growth and foraging in a flush feeding lepidopteran larva. J. Evolution. Biol. 12(3), 471-482.
Klowden, M. J. (2007). Physiological systems in insects. Academic press.
Lahiri, S., Shen, K., Klein, M., Tang, A., Kane, E., et al. (2011). Two alternating motor programs drive navigation in Drosophila larva. PloS one. 6(8), e23180.
Lindauer, M. (1954). Temperaturregulierung und wasserhaushalt im bienenstaat. Z. Vergl. Physiol. 36(4), 391-432..
Lindauer, M. (1952). Ein beitrag zur frage der arbeitsteilung im bienenstaat. Z. Vergl. Physiol. 34(4), 299-345
Loveless, J., Lagogiannis, K., Webb, B. (2019). Modelling the mechanics of exploration in larval Drosophila. Plos Comput. Biol. 15(7), e1006635.
Mänd, M., Kuusik, A., Martin, A.J., Williams, I.H. (2006). Regular periods of abdominal contractions recorded from larvae of the bumblebee, Bombus terrestris (Hymenoptera: Apidae). Eur. J. Entomol. 103(2), 319.
Mantziaris, C., Bockemühl, T., Büschges, A. (2020). Central pattern generating networks in insect locomotion. Dev. Neurobiol. 80(1-2), 16-30.
McIndoo, N.E. (1929). Tropisms and sense organs of Lepidoptera. Smithsonian Miscellaneous Collections.
Mirth, C.K., Riddiford, L.M. (2007). Size assessment and growth control: how adult size is determined in insects. Bioessays. 29(4), 344-355.
Myser, W.C. (1954). The larval and pupal development of the honeybee Apis mellifera Linnaeus. Ann. Entomol. Soc. Am. 47(4): 683-711.
Nakamatsu, Y., Tanaka, T., Harvey, J.A. (2006). The mechanism of the emergence of Cotesia kariyai (Hymenoptera: Braconidae) larvae from the host. Eur. J. Entomol. 103(2), 355.
Oertel, E. (1930). Metamorphosis in the honeybee. J. Morphol. 50(2), 295-339.
Pankiw, T. (2004). Cued in: honey bee pheromones as information flow and collective decision-making. Apidologie. 35(2), 217–226.
Qin, Q.H., He, X.J., Barron, A.B., Guo, L., Jiang, W.J., Zeng, Z.J. (2019). The capping pheromones and putative biosynthetic pathways in worker and drone larvae of honey bees Apis mellifera. Apidologie, 50(6), 793-803.
Quillin, K.J. (1999). Kinematic scaling of locomotion by hydrostatic animals: ontogeny of peristaltic crawling by the earthworm lumbricus terrestris. J. Exp. Biol. 202(6), 661-674.
Saraswati, S., Fox, L.E., Soll, D.R., Wu, C.F. (2004). Tyramine and octopamine have opposite effects on the locomotion of Drosophila larvae. J. Neurobiol. 58(4), 425-441.
Schmehl, D.R., Tomé, H.V., Mortensen, A.N., Martins, G.F., Ellis, J.D. (2016). Protocol for the in vitro rearing of honey bee (Apis mellifera L.) workers. J. Apicult. Res. 55(2), 113–129.
Shuel, R.W., Dixon, S.E. (1959). Studies in the mode of action of royal jelly in honeybee development: II. Respiration of newly emerged larvae on various substrates. Can. J. Zool., 37(5), 803–813.
Siefert, P., Buling, N., Grünewald, B. (2021). Honey bee behaviours within the hive: Insights from long-term video analysis. Plos one, 16(3), e0247323.
Siefert, P., Hota, R., Ramesh, V., Grünewald, B. (2020). Chronic within-hive video recordings detect altered nursing behaviour and retarded larval development of neonicotinoid treated honey bees. Sci. Rep.-UK, 10(1), 1–15.
Simon, M.A., Fusillo, S.J., Colman, K., Trimmer, B.A. (2010). Motor patterns associated with crawling in a soft-bodied arthropod. J. Exp. Biol. 213(13), 2303-2309.
Smirnoff, W.A. (1960). Observations on the migration of larvae of Neodiprion swainei Midd.(Hymenoptera: Tenthredinidae). Can. Entomol. 92(12), 957–958.
Van Griethuijsen, L.I., Trimmer, B.A. (2014). Locomotion in caterpillars. Biol. Rev. 89(3), 656-670.
Vandenberg, J.D., Shimanuki, H. (1987). Technique for rearing worker honeybees in the laboratory. J. Apicult. Res. 26: 90-97.
Vazquez, D.E., Farina, W.M. (2020). Differences in pre-imaginal development of the honey bee Apis mellifera between in vitro and in-hive contexts. Apidologie. 51, 861–875
Voigt, D., Gorb, S.N. (2012). Attachment ability of sawfly larvae to smooth surfaces. Arthropod Struct. Dev. 41(2), 145-153.
Wang, J.W., Sylwester, A.W., Reed, D., Wu, D.A.J., Soll, D.R., et al. (1997). Morphometric description of the wandering behavior in Drosophila larvae: aberrant locomotion in Na+ and K+ channel mutants revealed by computer-assisted motion analysis. J. Neurogenet. 11(3-4), 231-254.
Wang, Y., Campbell, J.B., Kaftanoglu, O., Page, R.E., Amdam, G.V., et al. (2016). Larval starvation improves metabolic response to adult starvation in honey bees (Apis mellifera L.). J. Exp. Biol. 219(7), 960–968.
Wilson, E.O., Hölldobler, B. (2005). Eusociality: origin and consequences. P. Natl. Acad. Sci. USA. 102(38), 13367-13371.
Winston, M.L. (1987). The biology of the honey bee. Harvard University Press, Cambridge, Mass.
Wong, J.W., Meunier, J., Kölliker, M. (2013). The evolution of parental care in insects: the roles of ecology, life history and the social environment. Ecol. Entomol. 38(2), 123-137.
Acknowledgements
Firstly, DEV and WMF thank H. Verna and R. Velarde for their technical assistance with beekeeping. Secondly, we thank M. López Medus and A. Vidal for their technical assistance to develop the actometer and the tracking software. Lastly, we thank the University of Buenos Aires, CONICET and ANPCYT for their support.
Funding
The study was financially supported by ANPCYT (PICT 2016 2084), the University of Buenos Aires (UBACYT 2018 20020170100078BA) and CONICET (PIP 112–201501-00633) to W.M.F.
Author information
Authors and Affiliations
Contributions
Conceptualization was carried out by DEV and WMF. Design, experiments and data analysis were performed by DEV. Supervision of the project and funding acquisition by WMF. The manuscript was drafted and revised by DEV and WMF.
Corresponding author
Ethics declarations
Ethics approval
N/A.
Consent to participate
N/A.
Consent for publication
N/A.
Conflict of interest
The authors declare no competing interests
Additional information
Manuscript editor: Bernd Grünewald
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vázquez, D.E., Farina, W.M. Locomotion and searching behaviour in the honey bee larva depend on nursing interaction. Apidologie 52, 1368–1386 (2021). https://doi.org/10.1007/s13592-021-00907-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13592-021-00907-0