
Abstract
In a Pavlovian conditioning situation, an initially neutral stimulus may be made excitatory by nonreinforced presentations in compound with an established conditioned excitor [i.e., second-order conditioning (SOC)]. The established excitor may be either a punctate cue or the training context. In four conditioned suppression experiments using rats, we investigated whether SOC phenomena parallel other cue interaction effects. In Experiment 1, we found that the response potential of a target stimulus was directly related to the intertrial interval when SOC was mediated by a punctate cue, and inversely related to the intertrial interval when SOC was mediated by the training context. Experiment 2 demonstrated that punctate- and context-mediated SOC are oppositely affected by posttraining context extinction, and Experiments 3 and 4 demonstrated that context- and punctate-mediated SOC are differentially affected by conditioned stimulus (Experiment 3) and unconditioned stimulus (Experiment 4) preexposure treatments. These findings parallel phenomena in conditioned inhibition and cue competition situations.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Pavlov (1927) discovered that positive and negative mediation effects can occur when a target cue (X) is presented in conjunction with a nontarget cue (A). Positive mediation effects refer to situations in which the presence of A during presentations of X results in X becoming more like A in terms of its behavioral control. The first documented positive mediation effect in Pavlovian conditioning was second-order conditioning (SOC), which was initially observed after pairing the target stimulus [i.e., the second-order conditioned stimulus (CS)] with an excitatory punctate (i.e., discrete) CS (Pavlov, 1927). Subsequent research provided evidence that SOC can also be mediated by an excitatory training context (e.g., Marlin, 1983). That is, contextually mediated SOC can be observed after nonreinforced presentations of the target stimulus in an excitatory training context. SOC is a robust phenomenon and has been observed in numerous preparations, including conditioned suppression (Rizley & Rescorla, 1972; Stout, Escobar & Miller, 2004; Yin, Barnet, & Miller, 1994), appetitive conditioning (Holland & Rescorla, 1975), autoshaping (Rashotte, Griffin, & Sisk, 1977), and human contingency judgment scenarios (Jara, Vila, & Maldonado, 2006). Despite the generality of positive mediation effects, the overwhelming majority of research in associative learning has focused on negative mediation effects (cue competition and conditioned inhibition), which occur when the presence of A during trials involving X makes X’s behavioral control less similar to that of A. The aim of the experiments reported here was to determine whether SOC would empirically parallel some negative mediation phenomena.
There are several important similarities between SOC and negative mediation phenomena. Most relevant to the present discussion is that negative mediation, like SOC, can be driven by interaction between a target cue and either an excitatory punctate cue or an excitatory context. For example, conditioned inhibition can be driven by a target cue interacting with an excitatory context (i.e., explicitly unpaired conditioned inhibition) and by the interaction of a target cue with an excitatory punctate cue (i.e., Pavlov’s procedure for conditioned inhibition). In the explicitly unpaired procedure for conditioned inhibition, the inhibitory response potential of a target stimulus is established by repeatedly presenting the target stimulus alone in the training context and interspersing presentations of the unconditioned stimulus (US) alone in the training context (e.g., Friedman, Blaisdell, Escobar, & Miller, 1998; Rescorla, 1966; Urcelay & Miller, 2008). This phenomenon procedurally (and potentially empirically and theoretically) parallels SOC mediated by the training context because both involve highly similar procedures. Similar to SOC mediated by a punctate cue (e.g., Pavlov, 1927), conditioned inhibition can be generated by interaction between a target cue and a punctate, excitatory nontarget cue. Pavlov’s procedure for conditioned inhibition involves pairings of the nontarget cue (A) with the US interspersed among pairings of the target cue with A (i.e., A+/AX−), which is procedurally similar to SOC mediated by a punctate cue. In cue competition situations, analagous parallels are often observed. For example, overshadowing involves interaction between a target stimulus and a nontarget punctate cue (e.g., Pavlov, 1927), and the degraded contingency effect involves interaction between a target cue and an excitatory training context (e.g., Urcelay & Miller, 2006a), with both procedures attenuating behavioral control by the target.
Among the most striking phenomena discovered in our laboratory in recent years is counteraction between the cue interaction treatments (for a review, see Wheeler & Miller, 2008), which occurs when the conjoint administration of two cue interaction treatments that can independently influence responding to a target cue result in less of an effect than when either treatment is administered alone. Notably, many examples of counteraction phenomena involve counteraction between the training context and an excitatory nontarget punctate cue. For example, Urcelay and Miller (2008) conducted an experiment in which they compared conditioned inhibition to a target stimulus after a control treatment, explicitly unpaired training for conditioned inhibition (i.e., X−/+), Pavlov’s procedure for conditioned inhibition (i.e., A+/AX−), and explicitly unpaired training plus Pavlov’s procedure (i.e., A+/+/AX−/X−). In the groups that received a single type of conditioned inhibition training, the target stimulus developed the potential to reduce responding to an independently trained excitor (i.e., negative summation) and was slower to gain excitatory behavioral control than a neutral cue (i.e., retardation). Thus, both Pavlov’s and the explicitly unpaired procedures established strong inhibitory behavioral control by the target stimulus when they were administered alone. However, when both treatments were combined, less inhibitory behavioral control by the target stimulus was observed than when either treatment alone was administered. That is, contextually mediated conditioned inhibition (i.e., explicitly unpaired inhibition) and inhibition mediated by a punctate cue (i.e., Pavlovian inhibition) counteracted in their potentials to drive inhibition to the target stimulus. Similar phenomena have been observed in cue competition situations. For instance, Urcelay and Miller (2006a) discovered that conjoint administration of degraded contingency and overshadowing treatments resulted in more behavioral control by the target CS (i.e., less cue competition) than when either degraded contingency or overshadowing was administered alone.
The aim of the experiments reported here was to determine whether counteraction, which is prevalent in cue competition and conditioned inhibition, would also occur in SOC. To maintain some degree of empirical similarity with studies of counteraction in negative mediation situations, we examined the potential for counteraction between an excitatory punctate cue and an excitatory training context, each of which can mediate SOC, at least in some situations. Counteraction in SOC would consist of strong excitatory responding (relative to unpaired control conditions) to a target stimulus trained with either an excitatory punctate cue or an excitatory context, but weak excitatory responding to a target stimulus trained with both an excitatory punctate cue and an excitatory context. Similar to prior studies of negative mediation, we addressed this question by comparing the effect of manipulations that should affect the associative status of the training context (e.g., trial massing) on SOC mediated by the training context with SOC mediated by a punctate cue.
Experiment 1—trial spacing in punctate cue-mediated and context-mediated SOC
Towards establishing a parallel between SOC and negative mediation phenomena, in Experiment 1 we investigated the extent to which two mediators of SOC would counteract each other by exploring the potentially differential effects of trial spacing on SOC mediated by the training context and SOC mediated by a punctate training excitor (A). Counteraction effects in conditioned inhibition and cue competition situations indicate that the potential of a punctate cue to influence responding to a target cue is diminished when the context–US association is strong (e.g., Urcelay & Miller, 2006a, b). Similarly, the potential of the training context to influence responding to a target cue is reduced by the presence of an excitatory punctate cue during training with the target.
Experiment 1 sought to conceptually replicate the central finding of Stout, Chang, and Miller (2003) in a SOC situation. Stout et al. observed a counteraction between overshadowing (analogous to SOC mediated by a punctate cue) and trial massing (analogous to SOC mediated by a context). That is, when trials were spaced and training was elemental (i.e., lacking a nontarget punctate cue), neither the training context nor the nontarget punctate cue were able to influence responding to the target stimulus (i.e., strong excitatory responding to the target was observed). However, when elemental trials were massed, the training context could compete with the target stimulus for behavioral control, and reduced responding to the target was observed. Moreover, when the target was trained in the presence of a nontarget punctate (overshadowing) cue and trials were spaced, the punctate cue was able to compete with the target for behavioral control, and reduced responding to the target was again observed. But when the target cue was trained in the presence of an overshadowing cue and trials were massed, the potential of both the overshadowing cue and the context to compete with the target stimulus was reduced relative to when either trial massing or overshadowing was administered alone; that is, strong responding was seen. If a parallel effect occurs in SOC, responding to a second-order CS should be weak when it is not paired with the punctate excitor and the training context is not excitatory. When the target is trained without being paired with a first-order punctate CS, SOC should be weak with spaced trials and strong with massed trials because massed trials should maintain a strong context–US association, which should drive SOC. When the target is paired with a first-order punctate CS, SOC should be relatively strong when trials are spaced because this should allow the punctate cue to drive SOC to the target. However, when massed paired training is administered, responding should be weak because both the punctate cue and the training context should be strongly excitatory, which should cause counteraction between the context and the punctate CS. We delay consideration of theoretical mechanisms that could account for such a counteraction effect until the General Discussion.
Table 1 summarizes the 2 × 2 × 2 design used in Experiment 1. The first factor was whether the target was presented in the context of reinforcement (Context 1) or in an associatively neutral context [Context 2; Condition Control (Ctrl)], which effectively reproduces control groups for SOC that involve unpaired presentations of the second-order CS (X) with the first-order CS (A and the context). The second factor was whether or not a punctate excitatory cue (A) was present during training (Punctate vs. Context). The third factor was whether trials were Massed or Spaced. Notably, we omitted a control condition in which the first-order CS (A and the context) was unpaired with respect to the US because the flashing light (CS A) often serves as an effective first-order CS in our laboratory (e.g., Witnauer & Miller, submitted).
Method
Subjects
Subjects were 24 female and 24 male Sprague-Dawley, experimentally naïve, young adult rats, bred in our colony. The body weight ranged from 182 to 249 g in females and from 243 to 352 g in males. Subjects were individually housed and maintained on a 16/8-h (light/dark) cycle with experimental sessions occurring roughly midway through the light portion. Subjects had free access to food in the home cage. One week prior to initiation of the experiment, water availability was progressively reduced to 20 min per day, provided soon after all scheduled treatments.
Apparatus
Two distinctly different types of enclosures served as the training contexts (Contexts 1 and 2), and their physical identities were counterbalanced within groups. Enclosure R was a clear, Plexiglas chamber in the shape of a rectangular box (22.75 × 8.25 × 13.00 cm, length × width × height) with a floor constructed of 0.48-cm diameter stainless steel rods set 1.5 cm apart, center-to-center, connected by NE-2 neons. The grid floor allowed a 0.5-s, 0.7-mA constant-current footshock to be delivered by means of a high voltage AC circuit in series with a 1.0-MΩ resistor. Each of the six replicates of Enclosure R had its own environmental isolation chest and was dimly illuminated by a 2-W (nominal at 120 VAC) bulb driven at 80 VAC and mounted on an inside wall of the environmental isolation chest approximately 30 cm from the center of the animal enclosure. A visual stimulus that consisted of a flashing light (0.25 s on/ 0.25 s off) could be presented. The flashing light (Stimulus A) was provided by a 25-W bulb (nominal at 120 VAC, but driven at 80 VAC). The light was located approximately 30 cm from the center of the chamber. A 45-Ω speaker could provide a 10-s white noise at 6 dB (C scale) above background. This noise served as the target CS. The background noise level (primarily from a ventilation fan) was 74 dB (C scale).
Enclosure V was a 25.5-cm-long box in the shape of a truncated-V (28 cm high; 21 cm wide at the top/5.25 cm wide at the bottom). Each of six replicates of Enclosure V had its own environmental isolation chest. The floor and long sides were constructed of stainless steel sheets. The ceiling was of clear Plexiglas and the short sides were of black Plexiglas. The floor consisted of two parallel metal plates, each 2-cm wide, with a 1.25-cm gap between them, which could deliver a 0.7-mA, 0.5-s constant-current footshock. Enclosure V was dimly illuminated by a 7-W (nominal at 120 VAC) bulb driven at 80 VAC mounted on an inside wall of the environmental isolation chest approximately 30 cm from the center of the animal enclosure, with the light entering the animal enclosure primarily reflected from the roof of the environmental chest. Due to differences in the opaqueness of the enclosures, this level of illumination roughly matched that of Enclosure R. A 10-s flashing light cue (Stimulus A) could be provided by a 100-W bulb (nominal at 120 VAC, but driven at 80 VAC). Within the animal enclosure, the bulb was mounted on an inside wall of the environmental chest, approximately 30 cm from the center of the experimental chamber. The brightness of the flashing light emitted by this bulb was similar to that of the flashing light in Enclosure R. As in Enclosure R, a 10-s white noise 6 dB (C scale) above a 74 dB (C scale) background served as the target CS (X).
A third context served as the testing context for all subjects (Context 3). Twelve identical chambers, each measuring 30 × 25 × 32 cm, length × width × height), were used. The walls of each chamber were made of Plexiglas, and the floor was constructed of 0.5-cm-diameter rods, spaced 2 cm center-to-center. Each chamber was housed in an environmental isolation chest, which was dimly illuminated by a houselight, which was a 2-W (nominal at 120 VAC) bulb driven at 80 VAC, mounted on the ceiling of the experimental chamber. Each chamber was equipped with a water-filled lick tube (opening 0.3 cm in diameter) that extended about 1 cm from the rear of a cylindrical niche (diameter 4.5 cm), which was left–right centered on one wall, with its axis perpendicular to the wall and positioned 4 cm above the grid floor. An infrared photobeam was projected horizontally across the niche, 1 cm in front of the lick tube. In order to drink from the tube, subjects had to insert their heads into the niche, thereby breaking the infrared photobeam. Thus, the amount of time the photobeam was disrupted could be monitored; this served as our dependent measure. A 45-Ω speaker mounted on the interior of each environmental chest could deliver the 10-s white noise stimulus 6 dB (C scale) above background. Ventilation fans in each enclosure provided a constant 72 dB (C scale) of background noise.
Procedure
Subjects were randomly assigned to one of eight groups: Context Massed (n = 12), Context Spaced (n = 12), Punctate Massed (n = 12), Punctate Spaced (n = 12), Context Massed Ctrl (n = 6), Context Spaced Ctrl (n = 6), Punctate Massed Ctrl (n = 6), and Punctate Spaced Ctrl (n = 6). Sex and experimental chamber were counterbalanced as completely as possible within the eight groups. Subjects were exposed to target training in Contexts 1 and 2, whereas acclimation, reacclimation, and testing were conducted in a distinctly different context (Context 3). Changing the contexts between training and testing served to minimize possible differences among groups in terms of the test context.
Acclimation
All subjects were acclimated to the test context (Context 3) during daily 60-min sessions on Days 1 and 2. During these acclimation sessions, subjects were exposed to one 10-s presentation of the target stimulus (X) at 30 min into each acclimation session. The acclimation sessions served to reduce unconditioned suppression to the test context and the test stimulus during subsequent testing.
Training
On Days 3 through 6, all subjects received training. Subjects in the Massed trial condition (i.e., Groups Context Massed, Punctate Massed, Context Massed Ctrl, Punctate Massed Ctrl) experienced 6.5-min training sessions. These groups were exposed to footshock presentations at 30, 120, 210, and 300 s into the training sessions on Days 3 and 5 (see Table 1). In the Punctate mediating cue condition (i.e., Groups Punctate Massed and Punctate Massed Ctrl), these footshock presentations were preceded by the training excitor (A, a 10-s flashing light), such that the footshock and A coterminated. These two groups were also exposed to two nonreinforced presentations of Stimulus A at 75 and 255 s into each training session. For Group Punctate Massed, the target stimulus (X) was simultaneously presented with Stimulus A on these two daily nonreinforced trials. In the Context condition in which the context served as the mediating stimulus (i.e., Groups Context Massed and Context Massed Ctrl), subjects were exposed to training identical to that administered to subjects in the Punctate condition except that the Stimulus A presentations were omitted. Finally, subjects in the Control conditions (i.e., Groups Context Massed Ctrl and Punctate Massed Ctrl) were exposed to X presentations on Days 4 and 6 in Context 2 at 75 and 255 s into each 6.5-min session. On these 2 days, all other subjects were placed in Context 2 for 6.5 min, with no nominal stimuli being presented.
The Spaced condition was similar to the Massed condition, except that the mean intertrial interval (ITI) was 546 s instead of 61 s. Sessions were 55 min in duration with reinforced trials occurring at 5, 20, 35, and 50 min into each session. Nonreinforced trials occurred at 12 and 42 min into each session.
Reacclimation
On Days 7 and 8, all subjects were exposed to Context 3 for 60 min per day in absence of any nominal stimuli. This served to restabilize their licking behavior. Subjects were given extra 30-min sessions on the first day of reacclimation if they required more than 60 s to complete an initial 5 cumulative seconds of licking.
Testing
On Day 9, all subjects were tested on Stimulus X in Context 3 during a 16-min session. When each subject completed 5 cumulative seconds of drinking, CS X was presented. This ensured that all subjects were drinking at the time of CS X onset. As is the practice in our laboratory, the data from any rat that failed to complete 5 cumulative seconds of drinking within the initial 60 s of the test trial were eliminated from the study so that suppression scores would reflect fear of X unaffected by fear of the test context. No subject met this criterion. Lick-suppression values were calculated based upon the amount of time required to complete an additional 5 cumulative seconds of drinking in the presence of X. A 15-min ceiling was placed on individual scores. A log (base 10) transformation was performed on pre-CS and CS latencies to improve the within-group normality of the data, thereby allowing for parametric statistical analysis.
Results and discussion
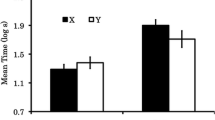
Group means for conditioned suppression during presentation of the target CS (X) are depicted in Fig. 1. SOC was observed in the Spaced trial condition when the mediating stimulus was a punctate stimulus and in the Massed trial condition when the mediating stimulus was the training context. Most important, we observed what appeared to be a counteraction between the mediating stimulus (A) and an excitatory context (in the Massed trial condition) in that no appreciable SOC was observed in Group Punctate Massed. The following analyses support these conclusions.

Experiment 1: mean time to complete 5 cumulative seconds of licking in the presence of the target conditioned stimulus (CS), X, in Context 3. Context = Contextual training excitor, Punctate = Punctate training excitor. Spaced and Massed refer to the spacing of trials. Control = Control treatment. See Table 1 for treatments of groups. Error bars: Standard errors of means (SEMs)
A 2 × 2 × 2 analysis of variance (ANOVA) on baseline behavior, with the first factor being trial spacing (Massed vs. Spaced), the second factor being mediating stimulus (Context vs. Punctate), and the third factor being whether or not the target stimulus was paired with an excitatory stimulus (Experimental vs. Control), revealed no main effects or three-way interaction in log latencies to complete the first 5 cumulative seconds of licking (i.e., prior to onset of X), all p > 0.29. Among the two-way interactions, the interaction between trial spacing and whether or not the target stimulus was paired with an excitatory stimulus alone reached significance, F(1, 64) = 7.42, p < 0.01. We acknowledge that this complicates interpretation of suppression to X, but we point out that this is probably a spurious finding and, more important, the difference is not one that could have contributed to any of the relevant differences observed in the CS scores. Moreover, subsequent experiments failed to detect such baseline differences.
For each of the following omnibus ANOVAs, Cohen’s f was calculated to serve as an estimate of effect size (Myers & Well, 2003). A 2 (Context vs. Punctate) × 2 (Massed vs. Spaced) × 2 (Experimental vs. Control) ANOVA on the log latencies in the presence of X detected a three-way interaction, F(1, 64) = 20.01, p < 0.01, f = 0.51, a main effect of whether X was paired with an excitatory stimulus (i.e., Experimental vs. Control), F(1, 64) = 22.30, p < 0.01, f = 0.54, and an interaction between the mediating stimulus and trial spacing, F(1, 64) = 10.34, p < 0.01, f = 0.36. The significant three-way interaction in conjunction with Fig. 1 suggests that the interaction between trial spacing and mediating cue was only observed among the experimental subjects. The following analyses were consistent with this inference. Within the experimental groups, a 2 × 2 ANOVA on log latencies detected an interaction between mediating stimulus (Context vs. Punctate) and trial spacing (Massed vs. Spaced), F(1, 44) = 36.98, p < 0.01, f = 0.87; this same analysis detected no appreciable interaction within the control subgroups, p > 0.25, suggesting that the interaction between trial spacing and mediating cue only occurred when the target stimulus was paired with the mediating stimulus. As no effect of trial spacing was detected in the control groups, subsequent analyses collapsed across the trial spacing variable within the control groups, thereby creating Groups Punctate Ctrl and Context Ctrl.
A planned comparison between Groups Punctate Spaced and Punctate Ctrl revealed a difference, F(1, 66) = 34.16, p < 0.01, indicating that SOC was mediated by the punctate stimulus when training trials were sufficiently spaced. Moreover, a planned comparison between Groups Context Massed and Context Ctrl proved to be significant, F(1, 66) = 22.98, p < 0.01, which indicates that SOC was mediated by the training context when training trials were massed. Furthermore, SOC was attenuated when training trials involving a punctate mediator were massed, which is supported by reliably attenuated conditioned suppression in Group Punctate Massed relative to Group Punctate Spaced, F(1, 66) = 26.30, p < .0.01. In contrast, suppression was reliably attenuated when training trials were spaced in the context-mediated condition, indicated by reduced conditioned suppression in Group Context Spaced relative to Group Context Massed, F(1, 66) = 19.93, p < 0.01. Lastly, the addition of Stimulus A in the massed condition attenuated SOC, as indicated by a comparison of Groups Punctate Massed and Context Massed, F(1, 66) = 12.69, p < 0.01, suggesting that the training context and the punctate CS A counteracted each other when both were potential mediators of SOC.
The augmented responding evoked by the target stimulus (X) in Groups Context Massed and Punctate Spaced, relative to Groups Context Ctrl and Punctate Ctrl, respectively, indicates that SOC was observed when X was paired with either an excitatory training context (Marlin, 1983) or an excitatory punctate stimulus (A; Pavlov, 1927). However, when X was paired with both of these mediating stimuli, its response potential was attenuated, as was shown by the comparisons between Groups Punctate Massed and Punctate Spaced (in which trial massing attenuated SOC) and those between Groups Punctate Massed and Context Massed (in which the pairings of X with A attenuated SOC). Thus, the excitatory context and punctate cue, each with the potential to mediate SOC when administered independently, counteracted in their potentials to drive SOC when they were trained together. This constitutes an important parallel between SOC and conditioned inhibition and cue competition.
It is possible that responding to X could have been influenced by generalization between A and X. This account assumes that generalization between A and X is specific to situations in which they are paired because little suppression to X was observed among subjects that received an unpaired control treatment. This analysis also explains the increased responding to X that was observed in the paired relative to the unpaired groups as well as the response reduction caused by massing trials within the Punctate condition. If massing trials attenuates A’s behavioral control, which would be expected on the basis of the trial massing effect, then the X’s response potential would be attenuated. The results of Experiment 1 could also have been influenced by first-order trace conditioning. The average interval between X presentations and US presentations may have been sufficiently short in the Massed condition to allow for some degree of trace conditioning to develop between the target stimulus and the US. Thus, the augmented responding due to massing training trials in the context-mediated groups (i.e., Massed Context and Spaced Context) could have been the result of first-order trace conditioning. Moreover, little trace conditioning to X would be expected in the Punctate condition because Stimulus A should compete with Stimulus X similar to the competition observed in overshadowing situations. What remains unclear in this analysis is how Stimulus A could simultaneously be both effective at competing with X and ineffective in provoking a conditioned response that would generalize to X.
It is also possible that between-group variations in suppression reflect differences in unconditioned suppression to the target stimulus. This is possible (despite unpaired controls for SOC) because the interval between the last exposure to X and testing was confounded between the experimental and unpaired control conditions. That is, control subjects received unpaired presentations of X on Day 5, but experimental subjects received X–A or X– context pairings on Day 4. Recovery from habituation to X might have occurred during the additional day of retention among control subjects, which could explain some of the increase in suppression to X. However, the amount of suppression to X observed in the experimental groups in Experiment 1 was far greater than the amount of unconditioned suppression observed in other experiments with similar stimuli in our laboratory (Witnauer & Miller, submitted).
Experiment 2—posttraining extinction of the training context
In Experiment 2, we investigated the effect of extinction of the training context subsequent to massed SOC training. This experiment was conducted to illuminate the mechanisms underlying the effects observed in Experiment 1 and to assess another potential parallel between SOC and negative mediation phenomena (e.g., cue competition and conditioned inhibition). To this end, we compared the effect of posttraining context extinction on punctate-mediated SOC with context-mediated SOC. In cue competition situations, cue competition between a target stimulus and the training context is greater with short ITIs (the trial massing effect). Moreover, conditioning a target stimulus in the presence of a highly salient nontarget punctate cue attenuates responding (overshadowing). When overshadowing and trial massing are administered conjointly, less cue competition is observed than when the treatments are administered alone. Thus, the training context and nontarget punctate (overshadowing) cue counteract when trials are massed (Stout et al., 2003). Experiment 1 replicated this finding in a positive mediation situation. An important aspect to the counteraction between overshadowing and trial massing is that overshadowing is restored (i.e., responding is attenuated) when the context is extinguished subsequent to training. Moreover, the trial massing effect is attenuated (i.e., responding increases) with posttraining extinction (Stout et al., 2003). An analogous finding in SOC would occur if, after massed training, context extinction caused a recovery in punctate-mediated SOC (i.e. responding increased) and a reduction in context-mediated SOC. Experiment 2 explored this possibility. To investigate whether posttraining extinction of the training context differentially impacted responding after massed SOC mediated by an excitatory punctate stimulus or by an excitatory training context, Experiment 2 used a 2 (20 vs. 480 min of context extinction) × 2 (Punctate vs. Context) design (see Table 2).
Method
Subjects
Subjects were 24 female (186–229 g) and 24 male (271–397 g) Sprague-Dawley, experimentally naïve, young adult rats that had been bred in our colony. They were maintained in the same manner as in Experiment 1.
Apparatus
Two distinctly different types of enclosures served as the training and test contexts (Contexts 1 and 2, respectively), the physical identities of which corresponded to Enclosures R and V from Experiment 1, counterbalanced within groups.
Procedure
Subjects were randomly assigned to one of four groups (all ns = 12): Context 20 min (Context-20), Context 480 min (Context-480), Punctate 20 min (Punctate-20), and Punctate 480 min (Punctate-480). SOC was conducted in a manner similar to the training of the Massed groups of Experiment 1 (i.e., Groups Context Massed and Punctate Massed on Days 3 and 5 of that study), except that now training was conducted over the course of 2 days (Days 3 and 4), and subjects were exposed only to the context where target training occurred, which corresponds to Context 1 in Experiment 1. Testing occurred in Context 2, which was physically identical to Context 2 in Experiment 1. Thus, subjects received target training in Context 1, whereas Acclimation, Reacclimation, and Testing were conducted in a distinctly different context (Context 2). Switching contexts between training and testing served to minimize possible group differences in fear of the test context. Acclimation (Days 1 and 2), Reacclimation (Days 9 and 10), and Testing (Day 11) were conducted in Context 2 in the same manner as in Experiment 1.
Context extinction
Days 5–8 consisted of context extinction treatment for Groups Punctate-480 and Context-480 and a handling control treatment for Groups Punctate-20 and Context-20. Groups Punctate-480 and Context-480 were exposed to Context 1 for 120 min during each daily treatment session for a total of 480 min of context exposure. Environmental chamber doors were opened once every 30 min to assure that the rats were awake. This exposure was intended to extinguish the context-US and context-A associations and perhaps the X-context and A-context associations. The handling and retention interval control treatment for the other two groups consisted of placing subjects in the same training context (Context 1) for 5 min per session, for a total of 20 min of context exposure during this phase of Experiment 2. Preliminary research in our laboratory has indicated that if context extinction is going to yield retrospective revaluation, a total extinction duration of multiple hours is required. During these context exposure sessions, no nominal stimuli were presented.
A log transformation of the pre-CS and CS test latency scores was performed as in Experiment 1. No subjects met the elimination criterion described in Experiment 1.
Results and Discussion
Figure 2 depicts the group mean log latencies to drink in the presence of Stimulus X. Reduced responding in Group Punctate-20 relative to Context-20 indicates that the presence of Stimulus A during training disrupted SOC to X and replicates the critical finding of Experiment 1. Most importantly, within the Punctate condition, context extinction yielded a recovery of conditioned suppression to X, whereas the identical treatment attenuated responding in the Context condition. The following analysis supported these observations.

Experiment 2: mean time to complete 5 cumulative seconds of licking in the presence of the target CS (X) in Context 2. Context = contextual training excitor, Punctate = punctate training excitor, 480 min = 480 min of posttraining context exposure, 20 min = 20 min of posttraining context exposure. See Table 2 for treatments of groups. Error bars: SEMs
A 2 (Context vs. Punctate) × 2 (480 vs. 20) ANOVA detected no main effect or interaction, all p > 0.43, on the log latencies to complete the first 5 cumulative seconds of licking in the test context, prior to the onset of X. A 2 × 2 ANOVA on log latencies in the presence of Stimulus X detected an interaction between training excitor and context extinction, F(1, 44) = 37.46, p < 0.01, f = 0.87. Planned comparisons were conducted to determine the source of this interaction. A planned comparison between Groups Punctate-20 and Punctate-480 revealed a difference, F(1, 44) = 20.05, p < 0.01, suggesting that the trial massing deficit within the Punctate condition was alleviated by posttraining context extinction. Moreover, context extinction subsequent to massed contextually mediated SOC reliably attenuated the response potential of the target stimulus, as evidenced by a reliable difference between Groups Context-20 and Context-480, F(1, 44) = 17.44, p < .01.
The results of Experiment 2 support the view that SOC is similar to other cue interactions. The interaction between the target cue and the punctate training excitor was enhanced when the training context was extinguished. Moreover, the interaction between the target cue and the training context was attenuated by posttraining context extinction. These effects parallel findings in cue competition (Stout et al., 2003; Urcelay & Miller, 2006a) and conditioned inhibition situations (Urcelay & Miller, 2006b). A potentially important limitation to Experiment 2 is that the unpaired control groups from Experiment 1 were omitted. However, because the only difference between Experiments 1 and 2 (aside from target manipulations) was that subjects did not receive control exposures to Context 3 in Experiment 2 and did not receive 20 min of Phase 2 context exposure in Experiment 1, it seems reasonable to assume that the control groups would perform similarly in both experiments.
Experiment 3—CS-preexposure
The results of Experiments 1 and 2 are consistent with the view that SOC parallels other cue interaction phenomena (e.g., conditioned inhibition and cue competition). Another important finding in the literature on cue competition and conditioned inhibition is that preexposure of a target stimulus (latent inhibition) attenuates the interaction between a target and a punctate companion cue. In contrast, effects driven by the training context (at least in conditioned inhibition situations) are relatively unaffected by CS preexposure. For example, conditioned inhibition established through Pavlov's procedure (analogous to punctate-mediated SOC) was attenuated, and explicitly unpaired inhibition (analogous to context-mediated SOC) was unaffected when the target cue was preexposed in the training context (Friedman et al., 1998). The purpose of the Experiment 3 was to determine whether or not a similar effect would occur in SOC. According to the view that SOC parallels conditioned inhibition, CS preexposure should not affect suppression when SOC is mediated by the context and should attenuate suppression when SOC is mediated by a punctate companion cue.
The design summarized in Table 3 was used to test these predictions. A foreseeable problem with this investigation was that the trial spacing parameters conducive to SOC mediated by the training context are considerably different from the trial spacing parameters conducive to SOC mediated by a punctate CS, as evidenced by the results of Experiment 1. Towards achieving optimal sensitivity in Experiment 3, the effect of Stimulus X preexposure on SOC was examined in two subexperiments. In Experiment 3a, the effect of Stimulus X preexposure on SOC mediated by the training context was investigated, and in Experiment 3b the effect of the same manipulation on SOC mediated by a punctate training excitor was investigated. Trial spacing parameters during training in both Experiments 3a and 3b were selected to achieve an intermediate level of responding in the groups that were not preexposed to X (i.e., Group Context in Experiment 3a and Group Punctate in Experiment 3b).
Experiment 3a
The aim of Experiment 3a was to determine whether enhancing the strength of the X-context association prior to contextually mediated SOC training would impact the response potential of Stimulus X at testing. The ITI in Experiment 3a was increased relative to the ITI used in the Massed groups of Experiments 1 and 2. Lengthening the ITI relative to the presumably optimally short ITI was done in order to slightly attenuate the response potential of Stimulus X, thereby avoiding any potential ceiling effects.
Method
Subjects and apparatus
Subjects were 12 female (179–207 g) and 12 male (261–309 g) Sprague-Dawley, experimentally naïve, young adult rats that had been bred in our colony. They were maintained in the same manner as in Experiments 1 and 2. The equipment used in Experiment 3a was identical to that used in Experiment 2.
Procedure
Subjects were randomly assigned to one of two groups (all ns = 12): Context or Context CS-pre. Groups Context and Context CS-pre were treated identically during training, which was similar to the training procedure used for groups in the Context condition of the mediating stimulus independent in Experiment 1, except that the mean ITI was greater. Acclimation, Reacclimation, and Testing proceeded in the same manner as in the previous experiments.
Preexposure
Preexposure was conducted in one 6.5-min session in Context 1 on Day 3. Subjects in Group Context CS-pre were exposed to eight presentations of X. Presentations occurred at 30, 60, 100, 160, 210, 260, 300, and 360 s into the session. Subjects in Group Context were exposed to a handling control procedure that involved placement in the context for 6.5 min. Thus, the two groups received identical exposure to the context, which implies that latent inhibition of the context is equated across conditions.
Training
In daily 12-min sessions in Context 1 on Days 4 and 5, subjects in Groups Context and Context CS-pre were exposed to contextually mediated SOC training in which Stimulus X was presented, interspersed with presentations of the footshock US. Subjects in these groups were exposed to four daily US presentations occurring at 60, 240, 420, and 600 s into each session. Two presentations of CS X were interspersed between footshock presentations (at 340 and 520 s into each training session).
Results
Group means for Experiment 3a are depicted on the left side of Fig. 3. A t-test failed to detect a difference between Groups Context and Context CS-pre on the log latencies to complete the first 5 cumulative seconds of licking in the test context, prior to the onset of X, p > 0.67. A similar analysis was used to assess the difference between these groups on log latencies to drink in the presence of X. This analysis also failed to detect a difference, p > 0.81.

Experiments 3a and 3b: mean time to complete 5 cumulative seconds of licking in the presence of the target CS (X) in Context 2. Context = contextual training excitor, Punctate = punctate training excitor. See Table 3 for treatments of groups. Error bars = SEMs
Experiment 3b
Experiment 3b sought to determine whether CS-preexposure would impact the response potential of Stimulus X after SOC training with a punctate companion stimulus. The mean ITI during training was increased by a factor of about four from the mean ITI used in Experiment 2. Moderate enhancement of the ITI was done in order to slightly enhance the response potential of Stimulus X, thereby avoiding any potential floor effects.
Method
Subjects and apparatus
Subjects were 12 female (180–216 g) and 12 male (252–294 g) Sprague-Dawley, experimentally naïve, young adult rats, bred in our colony, maintained in the same manner as in the previous experiments. The equipment used in Experiment 3b was identical to that used in Experiment 3a except for the addition of the flashing light which served as CS A.
Procedure
Subjects were randomly assigned to one of two groups (ns = 12): Punctate or Punctate CS-preexposure (Punctate CS-pre). Groups Punctate and Punctate CS-pre were treated identically during training, which was similar to the training procedure used in the Punctate condition of Experiment 2, except that the mean interval between trials was increased. Acclimation, Preexposure, Reacclimation, and Testing occurred in the same manner as in Experiment 3a.
Training
Subjects in Groups Punctate and Punctate CS-pre were exposed to daily, 30-min training sessions on Days 4 and 5. This training consisted of four daily pairings of the punctate training excitor (A) with the US, and two daily pairings of the punctate training excitor with X. Delay conditioning A-US presentations occurred at 2.5, 10.5, 19.0, and 27.0 min into each training session. Simultaneous A-X pairings occurred at 6 and 22 min into each session.
Results
Group means for Experiment 3b are depicted on the right side of Fig. 3. A t-test failed to detect a difference between Groups Punctate and Punctate CS-pre on the log latencies to complete the first 5 cumulative seconds of licking in the test context, prior to the onset of X, p > 0.21. A similar analysis was used to assess the difference between these groups on log latencies to drink in the presence of X. This analysis detected a difference, t(22) = 5.14, p < .0.01, indicating that pretraining exposure to Stimulus X attenuated the response potential of SOC mediated by a punctate training excitor.
Discussion: Experiment 3a and 3b
The results of Experiment 3 supported the view that SOC parallels conditioned inhibition with respect to target preexposure. Similar to the results of Friedman et al. (1998) in conditioned inhibition, in our experiment, target CS preexposure failed to affect the interaction between the target and the context (i.e., contextually mediated SOC was unaffected by preexposure). That is, the absence of a reliable difference between Groups Context and Context CS-pre suggests that CS preexposure did not appreciably enhance contextually mediated SOC. This null effect does not seem to be the result of either a ceiling or floor effect because the level of responding in both groups seemingly allowed for adequate sensitivity. Moreover, because the CS-preexposure treatment used in Experiment 3a was identical to the treatment used in Experiment 3b, which did reveal appreciable differences in responding, it seems unlikely that the manipulation was not strong enough to reliably impact responding. In Experiment 3b, the interaction between a target and a punctate companion cue was attenuated as a result of CS preexposure, which parallels the reduced conditioned inhibition observed by Friedman et al. when Pavlov's procedure for inhibition was preceded by preexposure of the target CS. This conclusion is supported by the reduced responding in Group Punctate CS-pre relative to Group Punctate.
Experiment 4—US preexposure
Urushihara and Miller (2006) reported that preexposure to the US attenuates the interaction between a target and a punctate companion (overshadowing) cue and, in contrast, the same preexposure enhances competition between the target the training context. Specifically, subjects received either elemental or compound (overshadowing) training. Orthogonally, subjects received either preexposure to the US or a handling control treatment in the training context. When the US was administered prior to elemental training, suppression was attenuated (i.e., competition between the context and the target was enhanced). However, when the US was preexposed prior to compound training, suppression was enhanced (competition between the target and the overshadowing cue was disrupted). Similar effects occur when the outcome in sensory preconditioning is repeatedly presented after training (Urushihara & Miller, 2006) or when unsignaled USs are interspersed among overshadowing trials (Urcelay & Miller, 2006a). Moreover, unsignaled US and CS-alone presentations interspersed among trials in Pavlov's procedure for conditioned inhibition disrupts the inhibitory potential of a target stimulus (Urcelay & Miller, 2008). Towards extending these findings to SOC, the design summarized in Table 4 was conducted, which investigated the effect of US preexposure in context-mediated SOC (Experiment 4a) and punctate-mediated SOC (Experiment 4b). Experiment 4 was divided into subexperiments for the same reasons as in Experiment 3.
Experiment 4a
Experiment 4a investigated the effect of US preexposure prior to contextually mediated SOC, with the expectation that this treatment would enhance the interaction between the target and the context (i.e., more responding would be observed after US preexposure than after a handling control treatment).
Method
Subjects and apparatus
Subjects were 12 female (193-228 g) and 12 male (219-310 g) Sprague-Dawley, experimentally naïve, young adult rats that had been bred in our colony. They were maintained in the same manner as in the previous experiments. The equipment used in Experiment 4a was identical to that used in Experiments 2 and 3.
Procedure
Subjects were randomly assigned to one of two groups (all ns = 12): Context or Context US-pre. Groups Context and Context US-pre were treated identically during training, which was the same as the training procedure used in Experiment 3a. Acclimation, Reacclimation, and Testing proceeded in the same manner as in the previous experiments. The only procedural difference between Experiments 3a and 4a was that pretraining exposure to the CS was replaced with pretraining exposure to the US in the training context.
Preexposure
Preexposure was conducted in one 6.5-min session on Day 3. Subjects in Group Context US-pre were exposed to eight presentations of the footshock US. Presentations occurred at 30, 60, 100, 160, 210, 260, 300, and 360 s into the session. Subjects in Group Context were exposed to a handling control procedure in which they were placed in the context for 6.5 min, with no nominal stimulus being presented. Thus, the two groups received identical exposure to the context. As in Experiment 3a, the short exposure to the context (6.5 min) was far too brief to have induced latent inhibition of the context (e.g., Jenkins, Barnes, & Barrera, 1981).
Results
Group means of suppression scores in the presence of X in Experiment 4a are depicted on the left side of Fig. 4. A t-test failed to detect a difference between Groups Context and Context US-pre on the log latencies to complete the first 5 cumulative seconds of licking in the test context, prior to the onset of X, p > 0.70. A similar analysis was used to assess the difference between these groups on log latencies to resume drinking, which proved reliable, t(22) = 2.07, p = 0.05, suggesting that US-preexposure augments behavioral control when SOC is mediated by the training context.

Experiment 4a and 4b: mean time to complete 5 cumulative seconds of licking in the presence of the target CS (X) in Context 2. Context = Contextual training excitor, Punctate = punctate training excitor. See Table 4 for treatments of groups. Error bars: SEMs
Experiment 4b
Experiment 4b investigated the effect of US preexposure on SOC mediated by a punctate cue, with the expectation that the target–punctate companion cue interaction would be attenuated after US preexposure (resulting in less responding) than after a handling control.
Method
Subjects and apparatus
Subjects were 12 female (196–230 g) and 12 male (235–320 g) Sprague-Dawley, experimentally naïve, young adult rats that had been bred in our colony. They were maintained in the same manner as in the previous Experiments. The equipment used in Experiment 4b was identical to that used in Experiment 4a.
Procedure
Subjects were randomly assigned to one of two groups (all ns = 12): Punctate or Punctate US-pre. Groups Punctate and Punctate US-pre were treated identically during training with the same procedure as that used in Experiment 3b. Acclimation, Reacclimation, and Testing proceeded in the same manner as in the previous experiments. The only procedural difference between Experiments 3b and 4b was that pretraining exposure to the CS was replaced by pretraining exposure to the US. Pretraining exposure to the US in Experiment 4b was conducted in exactly the same way as in Experiment 4a. As is the practice in our laboratory, all rats that failed to complete 5 cumulative seconds of drinking within the initial 60 s of the test trial were scheduled to be eliminated from the study for exhibiting excessive fear of the test context. One subject from Group Punctate US-pre met this criterion.
Results
Group means of suppression scores in the presence of X are depicted on the right side of Fig. 4. A t-test failed to detect a difference between Groups Punctate and Punctate US-pre in log latencies to complete the first 5 s of drinking during the test session, p > 0.79, suggesting that there were no appreciable differences between groups with respect to fear of the test context. A similar analysis of log latencies to drink in the presence of X detected a difference between Groups Punctate and Punctate US-pre, t(21) = 4.59, p < 0.01, which suggests that US-preexposure reliably attenuated the response potential of a target stimulus when SOC was mediated by a punctate cue.
Discussion: Experiment 4a and 4b
Experiment 4 tested the effects of US-preexposure on the response potential of a second-order CS [i.e., the target stimulus (X)] when SOC was mediated by the training context (Experiment 4a) and by an excitatory punctate companion stimulus (i.e., CS A in Experiment 4b). These data suggest that, when second-order conditioned responding was mediated by the training context, US-preexposure augmented responding to X. Moreover, when SOC responding was mediated by Stimulus A, this same treatment had deleterious effects on the response potential of X. According to the view that SOC parallels other cue interactions, enhancement of the context–US association through US-preexposure was expected to differentially affect these two SOC situations. In Experiment 4a, this treatment endowed the training context with a greater potential to mediate SOC. In contrast, this treatment increased the capacity of the training context to counteract the response mediating potential of the punctate excitor in Experiment 4b.
General discussion
The results of Experiments 1, 2, and 4 provide evidence that an excitatory training context and an excitatory punctate cue can counteract each other in their potential to mediate SOC. Experiment 1 showed attenuated responding to a second-order CS after it was paired with an excitatory punctate first-order CS (A) in an excitatory training context (achieved through massing trials). Individually, the training context and Stimulus A were each potent mediators of SOC. This was evidenced by the observation of SOC mediated by A when training trials were sufficiently spaced and of SOC mediated by the training context when training trials were massed in the absence of Stimulus A. However, when the training trials were massed and CS A was present during training, X’s behavioral control was weaker than that with either massed trials without CS A or CS A without massed trials (i.e., spaced trials). Based solely on the results of Experiment 1, one could argue that massed trials resulted in trace conditioning, which allowed Stimulus A to compete with X in a manner analogous to blocking. That is, with a shorter interval between trials in training, the average interval between X and the US was shorter in the Massed than in the Spaced condition, which might have allowed for some degree of first-order trace condition to X. However, this explanation fails to account for the results of Experiment 2 in which posttraining extinction of the context attenuated X's response potential in the Context condition. Posttraining context extinction in first-order conditioning (which is similar to trace conditioning), if anything, increases (rather than decreases) responding in elemental conditioning situations (Urcelay et al., submitted). The results of Experiment 2 suggest that associatively deflating the context after massed SOC training increases punctate-mediated SOC and decreases context-mediated SOC. The results of Experiment 3 revealed that CS preexposure attenuates punctate-mediated SOC (Experiment 3b) and does not affect context-mediated SOC (Experiment 3a). US preexposure similarly disrupts punctate-mediated SOC (Experiment 4b) and enhances context-mediated SOC (Experiment 4a).
The results of these experiments parallel effects in conditioned inhibition situations. The results of Experiment 1 in the Context groups were consistent with the results of Hearst and Franklin (1977), who found that the strength of inhibitory behavioral control by a target stimulus was inversely related to the ITI when the target stimulus and the US were explicitly unpaired in the training context. The data from Experiment 1 suggest that the excitatory response potential of the target stimulus is inversely related to the ITI when the target stimulus and the US are explicitly unpaired in the training context (i.e., when SOC is mediated by the training context). Considerable research suggests that the potential of a target stimulus to control behavior, either in the form of conditioned inhibition or SOC, can be attenuated through associative deflation of the mediating stimulus (e.g., the training context in Experiment 2). Consistent with this view, Lysle and Fowler (1985) found that the inhibitory behavioral control of a target stimulus is attenuated after associative deflation of either the training context or the punctate training excitor. Results consistent with this finding have also been found in SOC experiments when SOC was mediated by a punctate stimulus (e.g., Rashotte et al., 1977; Rescorla, 1982; but see Holland & Rescorla, 1975; Rizley & Rescorla, 1972) and when SOC was mediated by the training context (e.g., Chang, Blaisdell, & Miller, 2003). The results of Experiment 2 provide evidence that the strength of contextually mediated SOC often depends upon the excitatory status of the mediating context at the time of the test rather than at the time of acquisition.
When SOC to the target stimulus was mediated by a punctate cue (A), the response potential was inversely related to the excitatory status of the training context. This relationship was observed in Experiment 1, with the ITI serving as the independent manipulation of the excitatory status of the context; that is, long ITIs favored SOC with a punctate first-order CS. Consistent with the results of Experiment 1, Urcelay and Miller (2006b) observed greater inhibition to a target stimulus that was embedded in Pavlov’s procedure for conditioned inhibition (i.e., A+/AX−; operationally similar to punctate cue-mediated SOC) when training trials were widely spaced, relative to when trials were massed, suggesting that the inhibitory strength of the target stimulus was directly related to the ITI (see also Maisiak & Frey, 1977). Additionally, in Experiment 2 extinction of the training context augmented behavioral control by X when SOC was mediated by A. The results of Experiment 2 parallel the results of Urcelay and Miller’s (2006b) Experiment 3 in which they observed reliable behavior indicative of inhibition (as assessed by summation and retardation tests) when they extinguished the training context following massed training trials. As previously mentioned, consistent with the similarities between SOC and CI, Urcelay and Miller (2008) also reported a counteraction effect when a target stimulus was trained using both Pavlov’s procedure and the explicitly unpaired (i.e., X−/+) procedure for inhibition. Finally, the results of Experiments 3a and 3b were consistent with the general position that SOC parallels other cue interaction phenomena. Pretraining exposure to X attenuated responding when X was paired with A during training, but this same manipulation did not impact responding when the training context was the mediator of SOC. These results are consistent with the observation that CS-preexposure reduces the strength of a conditioned inhibitor trained using Pavlov’s procedure for conditioned inhibition but does not affect an inhibitor trained with the explicitly unpaired procedure for conditioned inhibition (Friedman et al., 1998).
Our findings are also in accordance with several phenomena related to cue competition. Specifically, cue competition effects seem to be dependent upon the excitatory status of the competing cue at test, as evidenced by several observations of recovery from cue competition effects after extensive posttraining extinction of competing cues (e.g., Blaisdell, Gunther, & Miller, 1999; Kaufman & Bolles, 1981). Observations of recovery from cue competition through associative deflation of the competing cue are analogous to findings of attenuated SOC and conditioned inhibition through deflation of the excitatory companion stimulus, in that both effects involve an attenuated interaction between a target stimulus and its associate. A number of recent papers suggest that, when combined, some cue competition effects can counteract each other in a manner akin to that observed in the present experiments and in the aforementioned conditioned inhibition experiments (e.g., Sissons, Urcelay, & Miller, 2009; Urushihara, Stout, & Miller, 2004; Witnauer, Urcelay, & Miller, 2008). For example, overshadowing (i.e., the attenuated response potential of a target stimulus that is paired with the US in the presence of a particularly salient stimulus) seems to be attenuated when the context–US association is especially strong due to short ITIs during training (Stout et al., 2003) or unsignaled US-presentations in the training context (Urcelay & Miller, 2006a; Urushihara & Miller, 2006). These relationships were observed in SOC in the present Experiments 1 and 4 using trial massing and US-preexposure manipulations, respectively. Moreover, overshadowing is attenuated when the target stimulus–context association is augmented through CS-preexposure (e.g., Blaisdell, Bristol, Gunther, & Miller, 1998) or through partial reinforcement or CS-duration manipulations (Urushihara & Miller, 2007). These relationships are analogous to the findings of Experiment 3a in which CS-preexposure was shown to attenuate the subsequent interaction between the target stimulus and the excitatory companion stimulus (A).
The experiments reported here were motivated by, but not intended to directly test, the predictions of Stout and Miller's (2007) sometimes competing retrieval (SOCR) model. According to this model, responding to a target is determined not only by the target CS–US association, but also by the associations between background cues that are associated with the target cue (comparator stimuli) and the US. In negative mediation situations (e.g., cue competition and conditioned inhibition), comparator stimuli function to attenuate responding to a target stimulus. In positive mediation situations, comparator stimuli facilitate responding to the target. According to this model, when a target is trained in the presence of multiple comparator stimuli (e.g., the training context and CS A), those comparator stimuli compete with each other for the potential to influence the target's response potential. Thus, the model anticipated the present findings by asserting that the two comparator stimuli to the target (the training context and CS A) counteracted each other when they are simultaneously trained in compound with the target stimulus, which resulted in less positive mediation and, consequently, less responding to the target. This theoretical interpretation of the present data is of limited value in differentiating among contemporary accounts of positive mediation because models other than SOCR can also explain the present results. Specifically, those accounts of positive mediation based on within-compound associations can explain the present findings by asserting that the context and the punctate training excitor (A) compete for associative strength both with the US and with target CS. Such competition reduces the target CS's response potential, which depends on the target CS-A, target CS-training context, CS A-US, and training context-US associations (e.g., Pineno, 2007). Thus, the principle contribution of the present results is empirical in demonstrating strong parallels between positive and negative mediation.
The results of the experiments reported here suggest that an excitatory training context and punctate cue can counteract in their potential to mediate SOC. With the exception of Experiment 3a, an inverse relationship was observed between responding to a contextually mediated second-order conditioned stimulus (i.e., X) and responding to a second-order conditioned stimulus paired with an excitatory punctate cue. This suggests that the potential of the training context to mediate SOC is directly related to the potential of the training context to counteract a punctate cue, and vice versa. These results are consistent with experiments pertaining to cue competition and conditioned inhibition, which have consistently found that contextual associations can interfere with interactions between a target stimulus and its punctate associates (e.g., Urcelay & Miller, 2006a; 2008). Finally, these results parallel findings in conditioned inhibition situations, adding to the body of evidence suggesting that the determinants of SOC closely correspond to the determinants of conditioned inhibition.
References
Blaisdell, A. P., Bristol, A. S., Gunther, L. M., & Miller, R. R. (1998). Overshadowing and latent inhibition counteract each other: Support for the comparator hypothesis. Journal of Experimental Psychology: Animal Behavior Processes, 24, 335–351.
Blaisdell, A. P., Gunther, L. M., & Miller, R. R. (1999). Recovery from blocking through deflation of the blocking stimulus. Animal Learning & Behavior, 27, 63–76.
Chang, R. C., Blaisdell, A. P., & Miller, R. R. (2003). Backward conditioning: Mediation by the context. Journal of Experimental Psychology: Animal Behavior Processes, 29, 171–183.
Friedman, B. X., Blaisdell, A. P., Escobar, M., & Miller, R. R. (1998). Comparator mechanisms and conditioned inhibition: Conditioned stimulus-preexposure disrupts Pavlovian inhibition but not explicitly unpaired inhibition. Journal of Experimental Psychology: Animal Behavior Processes, 24, 453–466.
Hearst, E., & Franklin, S. R. (1977). Positive and negative relations between a signal and food: Approach withdrawal behavior to the signal. Journal of Experimental Psychology: Animal Behavior Processes, 3, 37–52.
Holland, P. C., & Rescorla, R. A. (1975). Second-order conditioning with food unconditioned stimulus. Journal of Comparative and Physiological Psychology, 88.
Jara, E. T., Vila, J., & Maldonado, A. (2006). Second-order conditioning of human causal learning. Learning and Motivation, 37.
Jenkins, H. M., Barnes, R. A., & Barrera, F. J. (1981). Why autoshaping depends on trial spacing. In C. M. Locurto, H. S. Terrace, & J. Gibbon (Eds.), Autoshaping and conditioning theory (pp. 255–284). New York: Academic Press.
Kaufman, M. A., & Bolles, R. C. (1981). A nonassociative aspect of overshadowing. Bulletin of the Psychonomic Society, 18.
Lysle, D. T., & Fowler, H. (1985). Inhibition as a "slave" process: Deactivation of conditioned inhibition through extinction of conditioned excitation. Journal of Experimental Psychology: Animal Behavior Processes, 11, 71–94.
Maisiak, R., & Frey, P. W. (1977). Second-order conditioning: The importance of stimulus overlap on second-order trials. Animal Learning & Behavior, 11, 290–294.
Marlin, N. A. (1983). Second-order conditioning using a contextual stimulus as S1. Animal Learning & Behavior, 11, 290–294.
Myers, J. L., & Well, A.D. (2003). Research design and statistical analysis, 2nd edn. Mahwah: Lawrence Erlbaum Associates
Pavlov, I. P. (1927). Conditioned reflexes. London: Oxford Press.
Pineno, O. (2007). A response rule for positive and negative stimulus interaction in associative learning and performance. Psychonomic Bulletin & Review, 14, 1115–1124.
Rashotte, M. E., Griffin, R. W., & Sisk, C. L. (1977). Second-order conditioning of the pigeon's keypeck. Animal Learning & Behavior, 5, 25–38.
Rescorla, R. A. (1966). Predictability and number of pairings in Pavlovian fear conditioning. Pschonomic Science, 4, 383–384.
Rescorla, R. A. (1982). Simultaneous second-order conditioning produces S-S learning in conditioned suppression. Journal of Experimental Psychology: Animal Behavior Processes, 8, 23–32.
Rizley, R. C., & Rescorla, R. A. (1972). Associations in second-order conditioning and sensory preconditioning. Journal of Comparative and Physiological Psychology, 81, 1–11.
Sissons, H. T., Urcelay, G. P., & Miller, R. R. (2009). Overshadowing and CS duration: Counteraction and a reexamination of the role of within-compound associations in cue competition. Learning & Behavior, 37, 254–268.
Stout, S. C., & Miller, R. R. (2007). Sometimes competing retrieval (SOCR): A formalization of the extended comparator hypothesis. Psychological Review, 114, 759–783.
Stout, S. C., Chang, R. C., & Miller, R. R. (2003). Trial spacing is a determinant of cue interaction. Journal of Experimental Psychology: Animal Behavior Processes, 29, 23–38.
Stout, S. C., Escobar, M., & Miller, R. R. (2004). Trial number and compound stimuli temporal relationship as joint determinants of second-order conditioning and conditioned inhibition. Learning & Behavior, 32, 230–239.
Urcelay, G. P., & Miller, R. R. (2006a). Counteraction between overshadowing and degraded contingency treatments: Support for the extended comparator hypothesis. Journal of Experimental Psychology: Animal Behavior Processes, 32, 21–32.
Urcelay, G. P., & Miller, R. R. (2006b). A comparator view of Pavlovian and differential inhibition. Journal of Experimental Psychology: Animal Behavior Processes, 32, 271–283.
Urcelay, G. P., & Miller, R. R. (2008). Counteraction between two kinds of conditioned inhibition training. Psychonomic Bulletin & Review, 15, 103–107.
Urushihara, K., & Miller, R. R. (2006). Overshadowing and the outcome-alone exposure effect counteract each other. Journal of Experimental Psychology: Animal Behavior Processes, 32, 253–270.
Urushihara, K., & Miller, R. R. (2007). CS-duration and partial-reinforcement effects counteract overshadowing in select situations. Learning & Behavior, 35, 268–271.
Urushihara, K., Stout, S. C., & Miller, R. R. (2004). The basic laws of conditioning differ for elemental cues and cues trained in compound. Psychological Science, 15, 268–271.
Wheeler, D. S., & Miller, R. R. (2008). Determinants of cue interactions. Behavioral Processes, 78, 191–203.
Witnauer, J. E., Urcelay, G. P., & Miller, R. R. (2008). Reduced blocking as a result of increasing the number of blocking cues. Psychonomic Bulletin & Review, 15, 651–655.
Yin, H., Barnet, R. C., & Miller, R. R. (1994). Second-order conditioning and Pavlovian conditioned inhibition: Operational similarities and differences. Journal of Experimental Psychology: Animal Behavior Processes, 20, 419–428.
Acknowledgments
NIMH Grant 33881 provided support for this research. We thank Eric Curtis, Sean Gannon, Ryan Green, Jeremie Jozefowiez, Mario Laborda, Bridget McConnell, Mikael Molet, Lisa Ng, Heather Sissons, and Gonzalo Urcelay for comments on an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Witnauer, J.E., Miller, R.R. Some determinants of second-order conditioning. Learn Behav 39, 12–26 (2011). https://doi.org/10.1007/s13420-010-0002-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13420-010-0002-6