
Abstract
Natal dispersal affects the gene flow, distribution, dynamics and social structure of a population. In many solitary mammals, dispersal is often male-biased, while females may remain philopatric. For the Eurasian lynx (Lynx lynx), dispersal distances vary greatly and may be explained by sex or by habitat characteristics, such as fragmentation. Juvenile lynx habitat selection during dispersal has not been studied before, but resident lynx select heterogeneous forested habitats and avoid human settlements and infrastructure through distinct temporal activity patterns. We studied movement patterns and habitat selection during the dispersal of 22 Eurasian lynx (7 females, 15 males) in Finland. We found no differences between the sexes in the onset age, duration, distance, route or route linearity. Dispersal took place mostly during the evening and at night but also during the day. Of the four most used habitats, the mixed forests and transitional woodlands were used more frequently than would be expected according to their availability, and conifer forests and fields less than would be expected. Housing and other human infrastructure, including roads, were the least frequently used categories. There was a highly significant difference between the used and available habitats in general. The lynx selected the habitat more carefully during the day than during the night.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Natal dispersal, which is the movement from a natal site to a breeding site, affects the gene flow within a population as well as the distribution, population dynamics and social structure (Bowler and Benton 2005). Understanding how and why individuals disperse is essential for population management and for predicting the persistence and future distribution of a species (Cote et al. 2010). In most mammal species, males disperse further than females (Greenwood 1980; Dobson 1982). This is thought to be one of the mechanisms to avoid inbreeding (Greenwood 1980; Liberg and von Schantz 1985) and to decrease male-male competition (Sandell 1989). In many solitary mammals, females may remain philopatric. Philopatry can be considered in different scales. Sometimes, philopatry is described as movement no further than ten home ranges away from the original site (Shields 1987) and sometimes simply as staying in the natal range or in a portion of it, and dispersal as movement out of the natal home range (Greenwood 1980; Waser and Jones 1983).
In the Lynx genus, male-biased dispersal has been documented in bobcats (Lynx rufus) (Janečka et al. 2006) but according to a molecular genetic study, not in Canada lynx (Lynx canadensis), possibly because of its cyclic population dynamics (Campbell and Strobeck 2006; Breitenmoser et al. 1993). A male bias has been reported based on radio-telemetry studies of Eurasian lynx (Lynx lynx) in Scandinavia (Samelius et al. 2012) but not in Central Europe (Zimmermann et al. 2005, 2007). In Eurasian lynx, natal dispersal usually occurs when new kittens are born between mid-May and mid-June; however, the age at onset varies from 8 to 24 months (Breitenmoser et al. 1993; Schmidt 1998; Zimmermann et al. 2005; Samelius et al. 2012).
In Central Europe, Eurasian lynx dispersal distances are substantially shorter than those in Scandinavia, although individual variation is large. In Central Europe, males dispersed 4.5–129 km, compared to 32–428 km in Scandinavia (Breitenmoser et al. 1993; Schmidt 1998; Zimmermann et al. 2005; Samelius et al. 2012). Females in Central Europe dispersed 2–81 km compared to 3–215 km in Scandinavia (Samelius et al. 2012). In Scandinavia, one third of female offspring did not disperse but established an overlapping home range with their natal home range (Samelius et al. 2012). Based on genetic studies, Eurasian lynx females in Finland seem to form kin clusters (Holmala et al. 2018), which are overlapping, genetically distinguishable structures suggesting at least partial philopatry.
In Central Europe, the availability of suitable habitats and the arrangement of vacant home ranges steer the direction and length of dispersal by Eurasian lynx (Schmidt 1998). Habitat selection by dispersing Eurasian lynx has not been previously studied, but studies on cougars (Puma concolor) and African leopards (Panthera pardus) reveal that dispersers select habitats non-randomly, as do resident adults in their home ranges (Zeller et al. 2014; Fattebert et al. 2015). However, dispersers are known to use a wider array of habitats and even traverse habitats thought to be impermeable for the species (Gastón et al. 2016; Vanbianchi et al. 2017). The time of day, prey availability and hunting conditions are known to influence lynx home range use (Podgórski et al. 2008; Filla et al. 2017; Gehr et al. 2017). Additionally, day resting sites are selected in areas away from all human infrastructure (Filla et al. 2017; Gehr et al. 2017; Signer et al. 2019). Lynx have a distinct circadian activity pattern (Heurich et al. 2014). The overall daily activity level in lynx is not influenced by the daylight duration, but the activity pattern is (Heurich et al. 2014). In areas with a complete day-night cycle, such as in Finland during spring and autumn, lynx activity is lowest during the day and highest at night, peaking at crepuscular times (Heurich et al. 2014). However, the bimodal differences and crepuscular peaks in activity gradually smoothen for lynx in the higher polar latitudes, and dispersing subadults are more active during the day than adults (Heurich et al. 2014).
It is unclear whether the travel between habitats is based on the nearest visible area, with route planning occurring during movement. To evaluate the dispersal and its link to habitats, we analysed the dispersal movement parameters of 22 Eurasian lynx in Finland. We hypothesised that (1) males disperse further than females, (2) dispersal movement takes place mostly at night and (3) dispersing lynx use their habitats non-randomly. No barriers to the dispersal of Eurasian lynx are known in Finland, although large roads and lakes may direct the route. Therefore, the habitat selection by dispersing lynx can be studied without specific corridors channelling selection decisions.
Material and methods
Study area
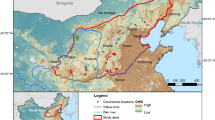
The study was conducted in southern and central Finland (61°N, 25°E; Fig. 1), encompassing about two thirds of the country (approximately 237,000 km2). We defined the study area by the national borders except in the north, where we set the limit to include the area used by the northernmost dispersing lynx. This was because we do not collar lynx in the reindeer herding area. The area was located in the boreal zone, which comprises a mosaic of agricultural land and coniferous (spruce Picea abies, pine Pinus sylvestris) and deciduous commercial forests (silver birch Betula pendula, downy birch Betula pubescens, black alder Alnus glutinosa, grey alder Alnus incana, aspen Populus tremula, bird cherry Prunus padus and rowan Sorbus aucuparia). Small settlements and several small- and medium-sized lakes are situated within the area. The mean annual temperature in the middle of the study area is 4.2 °C, ranging from − 6.5 °C in January to 16.6 °C in July. The ground is covered with snow for approximately 6 months of a year. The human population density is from 3.9 people/km2 in the north to 170.4 people/km2 in the most crowded areas in the south.

The dispersal of Eurasian lynx was studied in an area that encompassed about two thirds of Finland. In the close-ups: examples of long-distance (a, c) and short (b, d) dispersal by female (a, b) and male (c, d) Eurasian lynx. We could not present all 22 dispersals on the same map because of overlap
GPS-GSM radio-tracking
A total of 22 juvenile Eurasian lynx (15 males, 7 females) were captured and radio-tracked during 2009–2017. The lynx were captured mainly with wire or wooden baited box traps. The lynx were immobilised by using a combination of medetomidine and ketamine. The animals were weighed, sexed and fitted with GPS collars with remote GSM download technology and a programmable drop-off system (manufactured by Vectronic Aerospace GmbH, Germany; Followit, Sweden and Lotek, UK, respectively) and plastic ear-tags (Dalton, UK). Only the lynx weighing 10 kg or over and in good general health were fitted with radio-collars, the most typical collars used in our study representing approximately 3–4% (or less) of the individual’s weight. This was to ensure that mostly “normally” behaving individuals are tagged and to minimise ethical issues concerning the animal’s welfare. The age was determined by the lynx’s weight, body proportions, tooth wear and general appearance. The animals were released at the trapping location. The research plan and immobilization protocol were approved by the Finnish Animal Ethics Committee (ELLA). In addition, permits for capture were obtained from the Finnish Wildlife Agency and Finnish Ministry of Agriculture and Forestry. The collars transmitted the locations of the animals typically at 4-h intervals for a total of six times per day.
Dispersal onset, distance, route and linearity
For this study, we defined dispersal as the movement from a natal site to a potential breeding site and philopatry as staying in or in close proximity of the natal home range. We defined close proximity as a distance of a maximum of one home range radius away from the natal home range. For this description, an imaginary circular home range with 10 km radius was used, which is also the model home range used for lynx population monitoring in Finland, modified from Scandinavian monitoring method (Linnell et al. 2007; Luke 2008-2018). Thus, if the dispersal distance exceeded 20 km, an individual was considered to have dispersed.
The existence of a potential home range (either natal or post-dispersal) was estimated visually in a GIS program and was based on the spatial arrangement of the chronological point locations. If locations clustered together constantly creating a stable pattern, the individual was categorised having a clear home range. Varying time periods were used; however, the data accumulation needed to be enough to confirm a pattern in the movement behaviour and allow a calculation of a Kernel home range. For a few individuals, the natal home range site was known based on field monitoring of the marked individual moving together with its parent (known pairs of parent-offspring) before leaving the area. Typically, the takeoff was a rather straight forward movement out of the natal home range, with no return. The end of the dispersal was estimated to have been reached when the accumulation of the point locations formed a stable pattern in a specific area for over varying period of time, but so that a Kernel home range could be calculated (Table 1).
For determining the travelled distance when a natal or post-dispersal home range could not be calculated based on telemetry data, the first point location outside or on the exact border of a calculated minimum convex polygon (MCP 100%) was considered the start point of the dispersal. The start was known for 10 individuals (Table 1) and not known for 12 individuals, for which the first location of radio tracking was used. The endpoint of the dispersal was considered as the last point location outside or on the exact border of a calculated post-dispersal MCP 100% home range for 10 individuals, and for seven individuals, the last location from the radio-tracking data. Additionally, five lynx (one female and four males) with unknown dispersal endpoints were later encountered as adult hunted specimens with known death locations, thus giving the areal location of the end point of the dispersal. Three of these were encountered on their earlier dispersal routes, suggesting that some of their dispersal routes may represent a part of their post-dispersal home range. For four individuals, neither the natal nor the post-dispersal home ranges could be verified from the point location data or other information.
The dispersal distance, or the A to B distance, was calculated as the straight-line distance from the first to the last dispersal location. The route distance was calculated as the distance between all locations in the chronological order. Because of the unknown dispersal start and/or endpoints for some of the animals, all calculated distances were effectively the minimum distances except for the five individuals for which all data could be collected. The linearity of the dispersal route was calculated with a distance to route, or linearity index (Bell and Kramer 1979), which was simply the proportion of the distance length to the total route length as a percentage. The index revealed how linear the dispersal route was. A high index value implies a relatively straight path, whereas a small value indicates that the animal meandered considerably (Bell and Kramer 1979; Spencer et al. 1990). For the five individuals who were found to have been hunted, the dispersal distance was measured to both the end of the collar function (A to B) and to the death location (A to C).
Habitat data
We classified the study area’s landscape into nine different land-use classes based on the Finnish CORINE Land Cover (CLC 2012) database, a standardised land-cover classification system in the European Union, which has a 20-m pixel size (Finnish Environment Institute 2012). There are 51 land-use classes in CORINE that are present in Finland, with four robustness or resolution levels. We used level 3, which was the second most refined class, and combined some similar classes together (for example, several human infrastructure classes were combined). We used our field work experience to determine the meaningful land-use classes for lynx. The used classes and the classes that were merged into those in level 3 as well as their availabilities in the study area are listed in Table 2.
We studied habitat use and habitat selection by intersecting point locations with habitat map and by comparing the habitat composition of the dispersal point locations (used) to those of the buffered travel route (available). We combined the chronological point locations into a line that we buffered with a two-kilometre radius, forming a 4-km-wide zone which was then overlaid with the habitat map. We chose the buffer width based on the mean distance travelled per 4-h interval (approximately 500 m) of radio-tracked lynx (K. Holmala, unpublished). In theory, an individual could start from the known point and travel 2 km within the 4-h interval. However, in order for this same individual to be in the next known point at the 4th hour, an animal could actually travel only 1 km away from its trajectory and then back. Thus, we considered 2-km buffer to represent adequate potential area available for an individual. All GIS analyses were performed using ArcGIS 10.2. (Redlands, CA, USA) (ArcGIS Desktop 2014).
Statistical analysis
For the analysis of diurnal variation in the travel, the mean dispersal speed of each lynx was computed separately for each 4-h interval. The statistical significance of the differences between the time intervals in the mean speed over all lynx was evaluated by fitting an ANOVA model with individual-level random effects. The pairwise comparisons between the time intervals were obtained with Tukey’s post hoc test (see Hothorn et al. 2008).
The ratios of habitat use over their availabilities were analysed with resource selection functions that followed the approach described by Aarts et al. (2008). The details are provided in Appendix .
We also tested how well the Manly-Chesson selection index, which is the habitat use divided by the habitat availability (Manly et al. 1972; Chesson 1978), described the lynx habitat selection. This index value was < 1 if the habitat was avoided and > 1 if the habitat was selected.
Because the dispersal variables were not normally distributed, we used the Mann-Whitney U test to examine the differences in the dispersal durations, distance (A to B distance) routes and distance to route indexes and Spearman’s rank order correlation for the dispersal durations, distances (A to B distance) and routes. We set the level of significance to 0.05 and used SYSTAT 13 and IBM SPSS Statistics 25. The analyses of the diurnal variation and resource selection functions were conducted in the R environment (R Core Team 2018) using the nlme package (Pinheiro et al. 2018) to fit the models with random effects and the multcomp package (Hothorn et al. 2008) to obtain the Tukey’s tests.
Results
Dispersal age, onset, duration, distance, route and route linearity
The median age at the onset of lynx dispersal was 10 months, ranging from 7 to 16 months. Both sexes typically began to disperse in April, also when only individuals with a known dispersal start were considered. Some lynx were caught and marked while they were already dispersing. Hence, their dispersal duration, distance and route are to be considered minimum values. In general, the females travelled shorter distances according to the straight line dispersal (A to B distance), with an average of 37.8 km for females (SD = 35.61) and 65.5 km for males (SD = 48.63), but the differences between the sexes were not significant (Nfemales = 7, Nmales = 15, Mann-Whitney U = 34,000, p = 0.192, d.f. = 1; Table 3). We also counted the distance from dispersal onset to the death place for the one female and three males that were later encountered as hunted individuals (A to C distance). This increased the female average dispersal distance to 42.32 km (SD = 34.42) and 67.86 km (SD = 49.68) for males but decreased the significance between sexes (Nfemales = 7, Nmales = 15, U = 70,000, p = 0.217, d.f. = 1). For the total route length travelled during the whole dispersal period, no differences between the sexes could be distinguished (Mann-Whitney U = 36,000, p = 0.245, d.f. = 1, meanfemale = 363.6 km; SD = 383.39; meanmale = 564.8 km; SD = 370.61). This was also true for the distance/route-percentage index describing the travel linearity (Mann-Whitney U = 50,000, p = 0.860, d.f. = 1) and dispersal duration (meanfemales = 92 days, meanmales = 124 days, Mann-Whitney U = 37,000, d.f. = 1, P = 0.274). There were no statistically significant differences in any of the dispersal parameters between all individuals and those five individuals for which dispersal start and finish were known.
The observed dispersal speed was highest at 16:00–20:00 h and lowest at 08:00–12:00 (Table 4, Fig. 2). The speed was significantly higher in all 4-h intervals between 16:00 and 04:00 than in any of the other intervals between 04:00 and 16:00. The dispersal duration was highly correlated with the route (rs = 0.949, N = 22, p = 0.001) but not with straight line dispersal distance (rs = 0.161, N = 22, p = 0.161).

Individual-specific mean dispersal speeds (km/h) of Eurasian lynx in each 4-h time interval describing the distribution of activity between time intervals and individuals. Same shade of grey level indicates the same individual. The individuals are in the same order within each 4-h interval. The order is determined by their overall mean dispersal speed
Habitat use and selection
When we compared the habitat use, the lynx used four habitats most frequently during dispersal: the mixed forests (42.9%), conifer forests (27.8%), fields and other agricultural lands (14.6%) and transitional woodlands (11.8%). Of these, the mixed forests and transitional woodlands were used more frequently than would be expected according to their availability, and conifer forests and fields less than would be expected according to their availability (Fig. 3). Additionally, broad-leaved forests were used more than would be projected with their availability, but all other habitats were used less than would be expected based on their availability. The use of forests or canopy-covered areas increased during the daytime, while that of fields decreased. During the night, fields were used more than would be anticipated according to their availability.

Habitat use of Eurasian lynx (N = 22) during dispersal and the composition of available habitat types on their dispersal routes in the landscape. The habitat use was determined according to the point location data and the CORINE 2012 Land Use data. MF = mixed forest, TWL = transitional woodland, CF = conifer forest, Fields = fields and agricultural land, BL = broad-leaved forest, Peats = peatland, WB = water bodies, HI = housing, OI = other infrastructure
When looking at the resource selection functions, both the main effect of habitat and its interaction with the time of day were highly significant predictors of use-availability ratio. The transitional woodlands and mixed forests were always selected with significantly greater probability than those of the coniferous forests or water bodies (Table 5). In the daytime, all forests and transitional woodland were selected with significantly greater probability than were fields, but the difference decreased during the night. Estimates of the selection probabilities had great uncertainty for the broad-leaved forests, peatlands, housing and infrastructure because of their small areas among the available habitats (Fig. 4).

Estimates and 95% confidence intervals of habitat selection indicators derived from the estimated resource selection functions. The displayed values pjk for each habitat j (y-axis) and time-of-day k (daytime left night-time right) can be interpreted as probability to select habitat j at time-of-day k if all habitats were equally available for dispersing Eurasian lynx (sexes pooled)
Also the other used index for habitat selection, the Manly-Chesson selection index showed similar results for most of the habitats but failed to describe use for the peatlands, which several lynx did not use at all. Therefore, we omitted peatlands from the results (Fig. 5). A drop in the number of individuals that actually had used the habitat (N = 16/22) and the high use of peatland by a few individuals apparently inflated the result of this habitat compared to its availability, resulting in false overall selection.

The Manly-Chesson habitat selection index for the Eurasian lynx during dispersal. Values = 1 indicate the habitat usage in proportion to availability values > 1 indicate habitat preference and values < 1 indicate avoidance. The error bars represent the standard errors. Peatlands were omitted from the figure because of their extremely low availability. MF = mixed forest, TWL = transitional woodland, CF = conifer forest, BL = broad-leaved forest, Fields = fields and agricultural lands, WB = water bodies, HI = housing, OI = other infrastructure
Discussion
We studied movement patterns and habitat use and selection during Eurasian lynx natal dispersal in Finland in a population without known dispersal barriers. Habitat use and selection by dispersing Eurasian lynx have not been previously studied. The typical timing for the onset of dispersal was April, which was close to the birth of a new litter in late May to early June. The onset ages did, however, vary between 7 and 16 months, agreeing with those known from other European populations that were typically 9 to 11 months (Breitenmoser et al. 1993; Schmidt 1998; Zimmermann et al. 2005; Samelius et al. 2012) (between 6 and 18 months).
In our study, there was large individual variation in natal dispersal distances and duration regardless of sex. However, we acknowledge that our sample size was most likely too small to detect any sex bias in dispersal. According to our previous genetic study, lynx females seem to form spatial matrilinear clusters, suggesting at least partial philopatry (Holmala et al. 2018). In Scandinavian telemetry studies, the maximum dispersal distances were shorter for females than males (although the range was wide for both sexes), and a third of females remained philopatric (Samelius et al. 2012). Our results provided information about those females that do disperse, which remains a common trait even when a majority may remain philopatric (Holmala et al. 2018). Long-range dispersal has been associated with a species’ range expansion (Thompson and Jenks 2010) and underlying, complex local population dynamics, while stable dynamics select for short-distance dispersal (Murrell et al. 2002). Local population stability possibly differs between sexes and may thus be difficult to differentiate from inbreeding avoidance or simply the social organization. Moreover, perturbations may be affected by intensive hunting which creates vacant home ranges.
To our knowledge, this is the first time the actual route travelled during dispersal was studied in this level of detail, as other papers on lynx dispersal have only reported the distances between the start and the finish, partly because the technology used previously (mainly VHF tracking) has not permitted the description of the routes. The dispersal distances of the Finnish lynx seem to fall somewhere between the dispersal distances of Scandinavian and Central European lynx. The maximum distances in Finland were substantially shorter than those in Scandinavia (Samelius et al. 2012), probably because of the vast mountainous areas with little canopy cover in northern Scandinavian landscapes. While the distance between the start and finish may provide information about where the lynx settles, it does not tell much about the dispersal per se, as according to our results, an average of 80% of the dispersal route resulted from movement other than the straight-line distance between the start and the end points. An extreme case was female F7, which dispersed 1178 km but ended up only 30 km from its onset point.
The dispersal speed was the highest in the evening and at night, although the lynx did travel during the day as well. In a low-density lynx population in Norway, female subadults moved 2.5 km per day (mean), which was similar to daily movement distances by resident females (2 km) but less than half of the daily distance of resident males (5.9 km) (Sunde et al. 2000). Male lynx home ranges in Norway are approximately three times larger than those of females, so the daily movement distances were in line with the home range sizes (Sunde et al. 2000), as was movement speed to route length shown in our study. The daylight period varies tremendously with season and spatially within Finland. An individual dispersing during spring and summer may only experience a few hours of darkness per day, whereas an individual dispersing in autumn or winter may only experience a few hours of daylight per day, making comparisons between the different light conditions statistically unsound. That is why we settled for using the division based on time-of-day (GMT).
The lynx in our study used mostly forest habitats but some of the habitats were used disproportionally to their availability. The lynx selected mixed forests during both night and day, whereas the conifer forests were used mainly during the night. The heterogeneity of the tree species and thick undercover in mixed forests likely attracted more prey than a commercial conifer forest and provided ample ambush and hiding places for lynx. In studies of habitat use and selection involving resident Eurasian lynx, a preference for forests has been documented (Niedziałkowska et al. 2006; Basille et al. 2013; Filla et al. 2017). In Poland, resident lynx selected for high complexity forest habitats with low visibility and used conifer forests less than expected (Podgórski et al. 2008).
The studied, dispersing lynx selected areas with sparse tree-cover in transitional woodlands. These habitats included sapling stands and early successional vegetation, which attract many small mammal species (Kirkland 1977; Swanson et al. 2011). Pine and spruce saplings also provide plenty of cover. Even though open areas were not selected, fields and agricultural lands were selected during the active hunting period at night. The Finnish agricultural landscape is a relatively small-scale mosaic that provides many potential hiding places for an ambush predator and could, like transitional woodlands, also be described as habitat with high ease of prey capture.
Dispersing lynx avoided human settlement and other human infrastructure, which included roads in our study. These results are in line with previous studies on resident lynx habitat selection (Bunnefeld et al. 2006; Basille et al. 2008, 2013; Filla et al. 2017). The lynx selected the habitat more carefully during the day than during the night, which is reflected as statistically significant interaction effect of habitat and time-of-day and as smaller differences between the estimated habitat selection indicators in the night-time than during the day (Table 5). Filla et al. (2017) found that resident lynx in Central Europe selected for more rugged, closed terrain away from human infrastructure during the day and open habitats during the night, presumably due to opportunities for hunting. The avoidance of open habitats was weaker during twilight and night in a study by Gehr et al. (2017). The lynx in their study responded to the trade-off between the prey availability and human presence by selecting high prey availability areas during times of low human activity (Gehr et al. 2017).
Our study shows that the lynx select for and between tree-covered habitats, also when the landscape is mostly composed of these habitats (totalling almost 70% of the study area). This could imply that for example in Central Europe, the scarceness of tree-covered habitats effectively emphasises their significance for the dispersing lynx, therefore playing a fundamental part in connecting populations and lynx conservation.
Future studies should refine the understanding of the arrangements of vacant and non-vacant home ranges in the landscape. It is crucial to incorporate ecological and genetic methods to gain better insight of the dispersal behaviour, especially the local population structure in relation to that. It is possible that young lynx caught for research do not represent the dispersers of the total population, as has been pointed out by others regarding for example the sex ratios, behavioural traits of caught individuals and biases caused by trapping methods (Tuyttens et al. 1999; Lofroth et al. 2008; Conde et al. 2010; Stuber et al. 2013). These discoveries support the need of using different methods for catching lynx or finding alternative tracking methods altogether.
In conclusion, Eurasian lynx dispersal occurred in the crepuscular and night-time activity pattern that is typical of the species. While dispersing, young lynx seemed to select their movement habitats so that they offered cover, security and hunting possibilities. Our study could not conclude anything on the sex bias in dispersal because of high individual variation and relatively few data on separate sexes.
Data availability
The data that support the findings of this study are available from Luonnonvarakeskus (Natural Resources Institute Finland, Luke), but restrictions apply on the availability of these data, which were used under the licence for the current study, and so are not publicly available. Data are, however, available from the authors upon reasonable request and with the permission of Luke.
References
Aarts G, MacKenzie M, McConnell B, Fedak M, Matthiopoulos J (2008) Estimating space-use and habitat preference from wildlife telemetry data. Ecography 31:140–160
ArcGIS Desktop (2014) Release 10.2.2. Redlands, Environmental Systems Research Institute
Basille M, Calenge C, Marboutin E, Andersen R, Gaillard JM (2008) Assessing habitat selection using multivariate statistics: some refinements of the ecological-niche factor analysis. Ecol Model 211:233–240
Basille M, Van Moorter B, Herfindal I, Martin J, Linnell JDC, Odden J, Andersen R, Gaillard J-M (2013) Selecting habitat to survive: the impact of road density on survival in a large carnivore. PLoS One 8:e65493
Bell WJ, Kramer E (1979) Search and anemotactic orientation of cockroaches. J Insect Physiol 25:631–640
Bowler DE, Benton TG (2005) Causes and consequences of animal dispersal strategies relating individual behaviour to spatial dynamics. Biol Rev 80:205–225
Breitenmoser U, Slough BG, Breitenmoser-Würsten C (1993) Predators of cyclic prey: is the Canada lynx victim or profiteer of the snowshoe hare cycle? Oikos 66:551–554
Bunnefeld N, Linnell JD, Odden J, Van Duijn MAJ, Andersen R (2006) Risk taking by Eurasian lynx (Lynx lynx) in a human-dominated landscape: effects of sex and reproductive status. J Zool 270:31–39
Campbell V, Strobeck C (2006) Fine-scale genetic structure and dispersal in Canada lynx (Lynx canadensis) within Alberta, Canada. Can J Zool 84:1112–1119
Chesson J (1978) Measuring preference in selective predation. Ecology 59:211–215
CLC Corine Land Cover data (2012) Finnish Environment Institute. https://www.avoindata.fi/data/fi/dataset/corine-maanpeite-2012, https://land.copernicus.eu/pan-european/corine-land-cover
Conde DA, Colchero F, Zarza H, Christensen NL Jr, Sexton JO, Manterola C, Chávez C, Rivera A, Azuara D, Ceballos G (2010) Sex matters: modeling male and female habitat differences for jaguar conservation. Biol Conserv 143:1980–1988
Cote J, Clobert J, Brodin T, Fogarty S, Sih A (2010) Personality-dependent dispersal: characterization ontogeny and consequences for spatially structured populations. Philos Trans R Soc Lond B Biol Sci 365:4065–4076
Dobson FS (1982) Competition for mates and predominant juvenile male dispersal in mammals. Anim Behav 30:1183–1192
Fattebert J, Robinson HS, Balme G, Slotow R, Hunter L (2015) Structural habitat predicts functional dispersal habitat of a large carnivore: how leopards change spots. Ecol Appl 25:1911–1921
Filla M, Premier J, Magg N, Dupke C, Khorozyan I, Waltert M, Heurich M (2017) Habitat selection by Eurasian lynx (Lynx lynx) is primarily driven by avoidance of human activity during day and prey availability during night. Ecol Evol 7:6367–6381
Finnish Environment Institute (2012) https://metatieto.ymparisto.fi:8080/geoportal/catalog/search/resource/details.page?uuid=%7BA58329D8-3ED0-47B8-B789-3095EE55C255%7D
Gastón A, Blázquez-Cabrera S, Garrote G, Mateo-Sánchez MC, Beier P, Simón MA, Saura S (2016) Response to agriculture by a woodland species depends on cover type and behavioural state: insights from resident and dispersing Iberian lynx. J Appl Ecol 53:814–824
Gehr B, Hofer EJ, Muff S, Ryser A, Vimercati E, Vogt K, Keller LF (2017) A landscape of coexistence for a large predator in a human dominated landscape. Oikos 126:1389–1399
Greenwood PJ (1980) Mating systems philopatry and dispersal in birds and mammals. Anim Behav 28:1140–1162
Heurich M, Hilger A, Küchenhoff H, Andrén H, Bufka L, Krofel M, Mattisson J, Odden J, Persson J, Rauset GR, Schmidt K, Linnell JDC, Yamazaki S (2014) Activity patterns of eurasian lynx are modulated by light regime and individual traits over a wide latitudinal range. PLoS ONE 9 (12):e114143
Holmala K, Herrero A, Kopatz A, Schregel J, Eiken HG, Hagen SB (2018) Genetic evidence of female kin clusters in a continuous population of a solitary carnivore the Eurasian lynx. Ecol Evol 8:10964–10975
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50:346–363
Janečka JE, Blankenship TL, Hirth DH, Tewes ME, Kilpatric CW, Grassman LI (2006) Kinship and social structure of bobcats (Lynx rufus) inferred from microsatellite and radio-telemetry data. J Zool 269:494–501
Kirkland GL Jr (1977) Responses of small mammals to the clearcutting of northern Appalachian forests. J Mammal 58:600–609
Liberg O, von Schantz T (1985) Sex-biased philopatry and dispersal in birds and mammals: the Oedipus hypothesis. Am Nat 126:129–135
Linnell JD, Odden J, Andersen R, Brøseth H, Andrén H, Liberg O, Ahlqvist P, Moa P, Kvam T, Segerström P, Schmidt K (2007) Distance rules for minimum counts of Eurasian lynx Lynx lynx family groups under different ecological conditions. Wildl Biol 13:447–455
Lofroth EC, Klafki R, Krebs JA, Lewis D (2008) Evaluation of live-capture techniques for free-ranging wolverines. J Wildl Manag 72:1253–1261
Luke 2008–2018 population estimates at national and game management district levels (in Finnish and Swedish) http://riistahavainnot.fi/suurpedot/kannanarviointi/lausunnot
Manly BFJ, Miller P, Cook LM (1972) Analysis of a selective predation experiment. Am Nat 106:719–736
Murrell DJ, Travis JM, Dytham C (2002) The evolution of dispersal distance in spatially-structured populations. Oikos 97:229–236
Niedziałkowska M, Jędrzejewski W, Mysłajek RW, Nowak S, Jędrzejewska B, Schmidt K (2006) Environmental correlates of Eurasian lynx occurrence in Poland–Large scale census and GIS mapping. Biol Conserv 133:63–69
Pinheiro J. Bates D. DebRoy S. Sarkar D, R Core Team (2018) nlme: linear and nonlinear mixed effect models. R package version 3.1–131.1
Podgórski T, Schmidt K, Kowalczyk R, Gulczyńska A (2008) Microhabitat selection by Eurasian lynx and its implications for species conservation. Acta Theriol 53:97–110
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna Austria. URL https://www.R-project.org/
Samelius G, Andrén H, Liberg O, Linnell JDC, Odden J, Ahlqvist P, Segerström P, Sköld K (2012) Spatial and temporal variation in natal dispersal by Eurasian lynx in Scandinavia. J Zool 286:120–130. https://doi.org/10.1111/j1469-7998201100857x
Sandell M (1989) The mating tactics and spacing patterns of solitary carnivores. In: Carnivore behavior ecology and evolution, Springer, Boston, pp 164-182
Schmidt K (1998) Maternal behaviour and juvenile dispersal in the Eurasian lynx. Acta Theriol 43:391–408
Shields WM (1987) Dispersal and mating systems: investigating their causal connections. In: Chepko-Sade B, Halpin ZT (eds) Mammalian dispersal patterns: The effects of social structure on population genetics. The University of Chicago Press, pp 3–24
Signer J, Filla M, Schoneberg S, Kneib T, Bufka L, Belotti E, Heurich M (2019) Rocks rock: the importance of rock formations as resting sites of the Eurasian lynx Lynx lynx. Wildl Biol 2019(1)
Spencer SR, Cameron GN, Swihart RK (1990) Operationally defining home range: temporal dependence exhibited by hispid cotton rats. Ecology 71:1817–1822
Stuber EF, Araya-Ajoy YG, Mathot KJ, Mutzel A, Nicolaus M, Wijmenga JJ, Mueller JC, Dingemanse NJ (2013) Slow explorers take less risk: a problem of sampling bias in ecological studies. Behav Ecol 24(5):1092–1098
Sunde P, Kvam T, Moa P, Negård A, Overskaug K (2000) Space use by Eurasian lynx Lynx lynx in central Norway. Acta Theriol 45:507–524
Swanson ME, Franklin JF, Beschta RL, Crisafulli CM, DellaSala DA, Hutto RL, Swanson FJ (2011) The forgotten stage of forest succession: early-successional ecosystems on forest sites. Front Ecol Environ 9:117–125
Thompson DJ, Jenks JA (2010) Dispersal movements of subadult cougars from the Black Hills: the notions of range expansion and recolonization. Ecosphere 1:1–11
Tuyttens FAM, Macdonald DW, Delahay R, Rogers LM, Mallinson PJ, Donnelly CA, Newman C (1999) Differences in trappability of European badgers Meles meles in three populations in England. J Appl Ecol 36:1051–1062
Vanbianchi CM, Murphy MA, Hodges KE (2017) Canada lynx use of burned areas: conservation implications of changing fire regimes. Ecol Evol 7:2382–2394
Waser PM, Jones WT (1983) Natal philopatry among solitary mammals. Q Rev Biol 58:355–390
Zeller KA, McGarigal K, Beier P, Cushman SA, Vickers TW, Boyce WM (2014) Sensitivity of landscape resistance estimates based on point selection functions to scale and behavioral state: pumas as a case study. Landsc Ecol 29:541–557
Zimmermann F, Breitenmoser-Würsten C, Breitenmoser U (2005) Natal dispersal of Eurasian lynx (Lynx lynx) in Switzerland. J Zool 267:381–395
Zimmermann F, Breitenmoser-Würsten C, Breitenmoser U (2007) Importance of dispersal for the expansion of a Eurasian lynx Lynx lynx population in a fragmented landscape. Oryx 41:358–368
Acknowledgements
We would like to thank Tanjona Ramiadantsoa and Timo Pitkänen for help with data management. We are grateful for those Luke field workers and volunteer hunters who helped in catching the lynx.
Funding
Open access funding provided by Natural Resources Institute Finland (LUKE). The study was partially financed by the Raija and Ossi Tuuliainen Foundation, the Emil Aaltonen Foundation, and the Academy of Finland (decision number 310072).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
The handling protocol of lynx was approved by the Finnish Animal Ethics Committee (ELLA) and fulfils the ethical requirements for research on wild animals in Finland. In addition, permits for wild animal capture were obtained from the Finnish Wildlife Agency and Finnish Ministry of Agriculture and Forestry.
Additional information
Communicated by: Krzysztof Schmidt
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 30 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Herrero, A., Heikkinen, J. & Holmala, K. Movement patterns and habitat selection during dispersal in Eurasian lynx. Mamm Res 65, 523–533 (2020). https://doi.org/10.1007/s13364-020-00499-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-020-00499-7