
Abstract
Populations of European pine marten (Martes martes) across Great Britain declined dramatically during the 18th and 19th centuries due to deforestation and human-wildlife conflicts. Pine marten recovery from their northern Scottish stronghold is limited following reintroduction in Galloway Forest District, south-west Scotland, in the 1980s. With suggestions that martens have influenced red squirrel population increases in neighbouring Ireland, marten reintroduction efforts are a priority in Great Britain. Reintroduction requires establishment of recruiting populations that depend on suitable food availability to meet the associated higher energy demands of recruitment. We, therefore, investigated the diet of reproductive denning female martens during denning (March–April), and the mixed sex population in summer for the first time in Galloway. Scats (n = 114) were analysed from clumps collected from occupied artificial den boxes in 2015 and 2016, as well as genetically verified scats (n = 44) from summer transects in 2014. We compared our results with other Scottish studies and found that carrion frequency of occurrence and biomass of prey ingested differed most significantly between the regional mixed sex spring diets and the diet of denning females in Galloway. Anurans, birds, and small mammals were the likely substitution of carrion; we hypothesise that these differences are related to sex-specific behaviours and scavenging risk. We, therefore, suggest that there are differences in the diet of denning female martens compared to mixed sex spring populations and that future translocations should consider abundances of anurans in Galloway, and even more importantly than before, small mammals and birds across Great Britain.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The European pine marten Martes martes is a semi-arboreal member of the mustelid family that is widely distributed across Europe. In Great Britain, human-wildlife conflicts, such as fur-trapping and pest control, and deforestation through the 18th to early 20th centuries led to their functional extinction across England and Wales pre-1920 (Lockie 1964). The First World War of the early twentieth century lead to the initial reduction in human-wildlife conflicts (Lockie 1964), while the Wildlife and Countryside Act of 1981 later granted legal protection of pine marten populations. However, even with an increase in woodland habitat over the last century (Scottish Heritage 2015), today, this situation is largely unchanged, with pine martens being essentially restricted to Scotland (Strachan et al. 1996) and isolated areas of northern England (Birks and Messenger 2010).
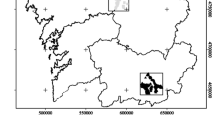
A successfully reintroduced, but limited, population of martens in the 1980s into Galloway Forest Park, Scotland (Fig. 1) (Shaw and Livingstone 1992) saw the beginnings of pine marten recovery into south-east Scotland. Other Scottish populations have since become the source for reintroductions further south, for example in mid-Wales (MacPherson 2016), with ongoing plans in England by the Vincent and Gloucestershire Wildlife Trusts (Townsend 2017).

Outline map of the Great British Isles in white in the top left-hand side with Scotland in a square enlarged and highlighted in grey with Glasgow and Edinburgh shown by triangles. The main study locations, Morangie, Kinlochewe, and Strathglass, are shown by squares while the Galloway study area is shown by the circle. The square around Galloway within south-west Scotland represents the map on the right-hand side showing the 4 study sites in and around Galloway and Galloway Forest Park (represented by dark shading labelled “Forest” in the Legend) in the Galloway Forest District. The thick black lines represent roads, while the dashed lines represent waterways. The 4 study sites are represented by the lighter shading whereby “CF” represents Carrick Forest, “GT” represents Glen Trool, “FB” represents the Fleet Basin and “AP” represents Arrecleoch and Penninghame. These maps were produced from Forestry Commission information licenced under the Open Government Licence v3.0 and a scale bar is provided
Successful reintroductions rely upon the establishment of released individuals, followed by population growth through recruitment, and subsequent spread. Reintroductions often have low success rates, for a wide variety of reasons (Griffith et al. 1989; Fischer and Lindenmayer 2000), of which dispersal and suitable food availability can be key (Bright and Morris 1994; Armstrong et al. 2007). More relevantly, Thompson and Colgan (1987) linked American pine marten (Martes americana) dispersal and reduced recruitment of young (kits) to lowered food availability.
So far, the diets of European and American pine martens have been well documented across their representative continents, covering season (De Marinis and Masseti 1995; Lynch and McCann 2007; Caryl 2008), predation pressure (e.g. Lindström et al. 1995), intraspecific competition (e.g. Lanszki et al. 2007), and sexual differences (Bull 2000; Zalewski 2007). However, little data directly focused upon the denning period (March–April) diet, despite its importance during recruitment when the energy demands of lactation to feed their dependent young are high (Bradbury and Vehrencamp 1977; Lewis and Kappeler 2005).
A Google Scholar keyword search of ‘diet’, ‘food’, ‘pine marten’, ‘martes’, ‘denning’ and ‘natal’ revealed only one paper that focused upon the denning period of pine martens, whereby wood mouse (Apodemus sylvaticus) population fluctuations and marten litter size in the Netherlands was studied by Kleef and Wijsman (2015), but this excluded any dietary assessment. This information deficiency is likely linked to the difficulty in locating dens for scat collection, with 82.8% of dens found by chance (Birks et al. 2005).
Placement of artificial den boxes, such as the wooden Vincent Wildlife Trust (VWT) and more recent “Galloway Lite” boxes—both currently used in Galloway (Croose et al. 2016b)—provide dens and enables population and dietary monitoring. During the use of den boxes for the care of litters, female martens often deposit their scat on box lids as an advertisement of use and ownership (Croose et al. 2016b).
As an opportunistic food generalist, pine marten diet in the British Isles is highly adaptable to seasonal variances and changing prey availability (e.g. Lockie 1961; Lynch and McCann 2007; Caryl 2008). In Scotland, diet varies regionally and annually. The following Scottish seasonal mixed sex dietary trends are inferred from studies by Lockie (1961), Balharry (1993) and Caryl (2008). There is significant contribution by small mammals throughout the year, especially in the form of field vole. In the winter and spring marten diet, small mammals and fruit dominate, while invertebrates and plant material dominate the summer marten diet, and bird consumption is higher in summer and spring than other months.
Pine marten also predate on red squirrels (Sciurus vulgaris), a species of conservation concern in the UK. Halliwell (1997) has documented detrimental effects of the presence of, and predation by, pine martens on reds in Scotland; however, Sheehy and Lawton (2015) compiled multiple other studies that found minimal presence or a complete absence of reds in marten diet across the British Isles. Sheehy and Lawton (2014) have also linked marten abundance to grey squirrel (Sciurus carolinensis) population crashes and showed that martens predated more frequently upon grey than red squirrels in Ireland. This is an exciting development as grey squirrels are currently viewed as a threat to native red squirrels through competition and disease transmission in the UK (Gurnell et al. 2004; Bruemmer et al. 2010; Collins et al. 2014).
When combining the above factors and an increasing focus on pine marten translocation, the need to thoroughly understand marten food ecology is fundamental. Therefore, we studied the dietary composition of female martens during denning using dietary niche breadth, frequency of occurrence of prey items, and biomass of prey ingested. Our study is the first to focus upon the diet of reproductive female pine martens during the crucial denning period, enabling comparison to the mixed sex summer diet of Galloway Forest District and mixed sex diets of martens situated in other Scottish regions.
Methodology
Study area and field vole abundance
The study area was within the Galloway Forest District—a predominantly first rotation, commercial coniferous forest area in the south-west of Scotland, U.K. (55° 6′ N, 4° 24′ W) (Fig. 1). Approximately 100 den boxes were present in the Forest District following the installation of 10 and then 40 VWT boxes in 2003 and 2013 respectively, before 50 “Galloway Lite” boxes were installed in 2014 (see Croose et al. 2016b). However, our study only includes scats from VWT boxes as “Galloway Lite” boxes had not yet been selected by females as dens. These VWT den boxes were situated across 4 different study sites in the Galloway Forest District both within Galloway Forest Park and the Park’s immediate vicinity: Carrick Forest (CF; 55° 13′ N, 4° 30′ W); Glen Trool (GT; 55°5′ N, 4° 33′ W); the Fleet Basin (FB; 55° 02′ N, 4° 16′ W) and Arrecleoch and Penninghame (AP; 50° 05′ N, 4° 48′ W and 55° 02′ N, 4° 42′ W) (Fig. 1). Due to the legal protection of den boxes only the study sites around the boxes are included in Fig. 1.
We compared our data to other Scottish studies located in Morangie, a managed plantation forest in north-east Scotland, U.K. (57° 48′ N 4° 06′ W), and Kinlochewe (57° 30′ N 5° 30′ W) and Strathglass (57° 15′ N 4° 45′ W), a mosaic of commercial forest and natural woodland, and a predominantly commercial forest, respectively, both in north-west Scotland, U.K. (Fig. 1). Due to the significance of field vole (Microtus agrestis) in the Scottish pine marten diet, such as in Kinlochewe and Strathglass (KS) (Balharry 1993) and Morangie (Caryl 2008), Birks collected field vole population abundance index data for Galloway. He used the field vole transect sign index method devised by the Mammal Society (Birks pers. comm.) across the same 4 study sites that den boxes were located (Fig. 1). This method is based on evidence of runs, feed signs and droppings recorded from 10 × 1 m quadrats placed by Birks along 100 m transects in May over 4 years (2013–2016; Online Resource 1). We averaged the 4 indices to give an average abundance index assumed to represent the entire study area for each year.
Scat collection
Springtime pine marten scats were collected by a trained and licenced team in clumps from the lids of VWT den boxes in use by reproductive female pine martens (see Croose et al. 2016b). Scats were collected from four boxes in 2015—one from each of the four study sites—and three boxes in 2016 in GT, FB, and CF, with the same box in CF in use over both years.
Without genotyping of scats, we must therefore assume that the diet of denning females represents 4–7 females. The team allowed for scat accumulation while minimising scat deterioration and pine marten disturbance by collecting clumped scats after the denning period (March–April) during mid to late May of each year. We defined the denning period as the time from which female martens give birth to when their dependent kits are no longer regularly housed in, in this case, den boxes.
Summertime scats (n = 44), which were assumed to be representative of the mixed sex summer diet of the pine marten population, were collected by two trained and licenced surveyors with trained sniffer dogs in September 2014 from along 1 km transects (see Birks 2016a). The team and surveyors placed clumped scats into one zip-seal bag per box, while individual summer scats were placed into their own bags, and then frozen.
Dietary component analysis
The trained team did not conduct genetic analysis on den box scats but assumed that they were those of females occupying the den boxes as pine martens are territorial (Powell 1979), and males are not present during the rearing of kits (Moors 1980). Birks (2016b) morphologically identified marten scats from transects with high accuracy and all scats collected were confirmed by DNA analysis in a population estimate study for the Fleet Basin by Croose et al. (2016a).
We thawed clumps in a fridge (4 °C) for 24 h, allowing the separation of individual scats. We placed these thawed clumps on a 30 × 20 × 15-cm grid system and selected intact scats at randomly generated coordinates. Due to the year-round varied nature of pine marten diet, we used the individually collected mixed sex summer scats to identify intact scats of any variation so that no scats were discarded. Balharry (1993) showed that a single prey item (squirrel) can occur across 12 scats from a single marten, and therefore, in studies such as ours, where scats are clumped on lids, it is unlikely that prey items were missed. This, therefore, allowed us to select a random and representative sample of scats from across the diet of denning females.
We then soaked selected scats for approximately 24 h to loosen material, then followed Lombardini et al. (2015) by washing them through a 0.5-mm sieve under running tap water, before drying remaining prey items in an oven (105 °C) for at least 8 h.
We then separated dried prey items into seven components; hair, feathers, invertebrate, anuran, plant material, bones, and teeth. We looked for carrion, as described by Balharry (1993), but did not find it, and we did not look for the chaetae of earthworms due to their absence from previous Scottish studies by Caryl (2008) and Balharry (1993). Examination under a dissecting microscope allowed us to identify bones to the lowest possible taxonomic level. We used teeth as a reliable indicator of species, found in 83.9% (n = 78) of denning female scats and 84.6% (n = 11) of mixed sex summer scats that contained mammal bones.
We microscopically analysed all large mammal guard hairs (> 20 mm guard hair length) found in addition to 20 guard hairs from all samples containing hair using the cuticular scale patterns and an identification key (Teerink 2003). We ensured a representative identification of hair by placing clumps of hair from each scat, in petri dishes split equally by 10, and selecting 2 guard hairs from each 10th. This enabled us to identify large and small mammal species missed by the absence of teeth across all scats containing hair from the spring (n = 112) and summer (n = 15).
We left feathers unidentified and instead used calculations by Caryl (2008) that stated a clear annual difference in small and large bird %FO (83 and 8% respectively), similarly supported by Balharry (1993). Caryl (2008) described small birds as birds up to the size of a blackbird (Turdus merula) and large birds as any bird above blackbird size based on quill diameter. Following allowances for egg shell %FO in our data, we split remaining bird %FO proportions to 91% (small bird) and 9% (large bird).
Upon identification, we grouped all components into dietary groups: bird, invertebrate, plant material, anuran, and small mammal including the pygmy shrew (Sorex minutus) and common shrew (Sorex araneus), bank vole (Myodes glareolus) and field vole, and wood mouse, and large mammal including red and grey squirrel, rabbit (Oryctolagus cuniculus), rat (Rattus rattus), and carrion (sheep and deer spp), for comparison to Morangie and KS. As we defined carrion in this way, as in other studies, carrion was assumed to be animals which were too large to be killed by martens, and instead killed by other sources and grazed upon by martens. We did not identify plant material (including fruit, seeds, and berries), egg shell, and invertebrates to a taxonomic level.
Data and statistical analysis
We analysed 158 scats comprised of 114 from spring den boxes and 44 from summer transects. We used the χ2 test for dietary comparison within Galloway, and Galloway with Morangie and KS.
We compared the diet of denning females to dietary data from March–April, and the mixed sex summer diet to corresponding summer diet. We analysed the data using altered Bonferroni confidence limits following Caryl (2008), calculated with k = 6, α = 0.05 for dietary group comparisons, k = 5, α = 0.05 for small mammal prey, and k = 3, α = 0.05 for bird prey.
Following Caryl (2008), we calculated the trophic niche breadths of the diets of denning females and mixed sex martens and the dietary groups therein using the Levin’s index (B) (Levins 1968) formula:
Where B represents diet breadth, p the proportion of the diet/dietary group comprised of dietary group/species i. We standardised (Ba) the Levin’s index using:
Where Ba represents the standardised dietary niche breadth and n the number of dietary groups. Values closer to 0 indicate a narrower niche breadth showing food specialisation, while values closer to 1 indicate a dietary generalisation. We then calculated niche breadth overlap (RO) using Pianka’s index (Pianka 1973):
Where RO represents the level of overlap between populations j and k. Values range from no overlap (0.0) to complete overlap (1.0).
We also calculated frequency of occurrence (FO), whereby a scat containing an identified prey animal indicates an occurrence, and biomass of that prey ingest (BPI). We used %BPI as a relative volumetric estimator of energy contribution by prey animals to the diet calculated following Balharry (1993):
Where CF represents corrections factors used from feeding trials of captive martens calculated by Balharry (1993) (Table 1), with an addition of the black rat (Rattus rattus). Due to their absence in previous studies and their overlap in weight range with squirrels, we gave rats the same correction factor as squirrels. We then defined %FO as the total FO of a dietary group or prey animal/total FO of all dietary groups or prey animals within the dietary group, × 100. We defined %BPI as the total BPI of a dietary group or prey animal/total BPI of all dietary groups or prey animals within the dietary group, × 100. We analysed the data using R software (Version 0.99.903 – © 2009-2016 RStudio, Inc.) and presented all results as significant differences in the data.
When we refer to seasons in Morangie and KS, both seasons represent mixed sex population diets. When we seasonally refer to Galloway, in spring, this represents the diet of denning females and in summer, the mixed sex population diet.
Results
Denning and regional mixed sex spring diet comparison
Over the two-year diet of denning females in Galloway, small mammal %FO was highest, followed equally by plant material, invertebrates, and birds. Anuran %FO was lower, but large mammal %FO was the lowest (Fig. 2). Small mammal %BPI was highest, followed by bird and anuran %BPI. Large mammal %BPI was still low, but plant material and invertebrate %BPI was the lowest (Fig. 2). The diet of denning females’ Ba was 0.67.

The diet of denning female pine martens in Galloway in 2015 and 2016 showing all present dietary groups. Data shown are expressed as a proportion of diet (%), showing frequency of occurrence (FO) and biomass of prey ingested (BPI)
Relative to the overall diet of denning females and within the bird dietary group, small bird %FO and %BPI were highest. Egg %FO was then higher than large bird %FO, but the opposite was true of their %BPI. Within the small mammal dietary group, field vole %FO and %BPI were highest, followed by wood mice, then bank voles and pygmy shrews, while common shrew were the lowest. Relative to the overall diet, the trend was similar, except bank vole and pygmy shrew %FO that was equal. Small mammal dietary group Ba was 0.43.
Between years, the diet of denning females did not differ by %FO or %BPI (χ2 = 0.8, df = 1, p = 0.9751 and χ2 = 1.3, df = 1, p = 0.939, respectively). Niche breadth overlap was high (RO = 0.99). Small mammal niche breadth overlap was also high (RO = 0.99). A single occurrence of black rat and red squirrel was observed in 2015, while a single rabbit was observed in 2016. Red squirrel only accounted for 1.1% of all mammal %BPI in the diet of denning females over the two years.
When we compared the diet of denning females in Galloway to the diet in Morangie, we found a high niche breadth overlap (RO = 0.92), however, %FO (χ2 = 22, df = 5, p = < 0.001) (Fig. 3) and %BPI (χ2 = 42.7, df = 5, p = < 0.001) differed (Fig. 4). Anuran %FO was higher (χ2 = 7.8, df = 1, p = 0.005) as was %BPI in Galloway compared to in Morangie (χ2 = 14, df = 1, p = < 0.001). Conversely, large mammals, showed a higher %FO and %BPI in Morangie (χ2 = 7, df = 1, p = < 0.008 and χ2 = 23.6, df = 1, p = < 0.001 respectively).

The six dietary groups found in the diet of denning female pine martens in Galloway compared to the mixed sex spring diet of pine martens in Morangie. Data is expressed as Frequency of Occurrence (%FO). There was a significant difference between %FO of diets. Anuran %FO was significantly higher in Galloway than in Morangie whereas large mammal %FO was significantly higher in Morangie than in Galloway. Significant differences are shown by a single asterisk “*”

The six dietary groups found in the diet of denning female pine martens in Galloway compared to the mixed sex spring diet of pine martens in Morangie and Kinlochewe and Strathglass (KS). Data is expressed as Biomass of Prey Ingested (%BPI). There was a significant difference in %BPI between Galloway and Morangie and Galloway and KS diets. Anuran %BPI was significantly higher in Galloway than in Morangie, whereas large mammal %BPI was significantly higher in Morangie and KS than in Galloway. Significant differences between Galloway and Morangie are shown by a single asterisk (*) and significant differences between Galloway and KS are shown by a double asterisk “**”
When comparing Galloway with KS, there was a difference in %BPI (χ2 = 52.3, df = 5, p = < 0.001) (Fig. 4) whereby large mammal %BPI was higher in Galloway (χ2 = 38.5, df = 1, p = < 0.001).
Mixed sex summer diet regional comparison
In the diet of mixed sex martens in Galloway in 2014, plant material and invertebrate %FO were equally highest (37.3% and 35.5%, respectively), but showed the lowest %BPI (4% and 3.8%, respectively). The small mammal dietary group %BPI (62.7%) clearly dominated the summer diet in Galloway, despite its intermediate %FO (19.1%), while anuran and bird %FO were equally lowest (4.5% and 3.6%, respectively) with an intermediate %BPI (15.4 and 14.1%, respectively). The diet of mixed sex summer martens in Galloway Ba was 0.47.
When compared regionally, we found that the summer diets of mixed sex martens in Galloway and Morangie showed high niche breadth overlap (RO = 0.88). Despite this, the diets differed by %FO (χ2 = 24.9, df = 5, p = < 0.001) and %BPI (χ2 = 53.1, df = 5, p = < 0.001) (Fig. 4).
Bird %FO was lower in Galloway (χ2 = 15.5, df = 1, p = < 0.001), reflected by a lower %BPI (χ2 = 10.8, df = 1, p = 0.001). Small mammals showed a higher %BPI in Galloway (χ2 = 9.4, df = 1, p = 0.002), and despite large mammal %FO not differing (χ2 = 1.4, df = 1, p = 0.2), large mammal %BPI was higher in Morangie (χ2 = 9.9, df = 1, p = 0.002). Anuran %BPI was higher in Galloway (χ2 = 15.1, df = 1, p = < 0.001) but did not differ by %FO with Morangie (χ2 = 4.2, df = 1, p = 0.04). Plant material %BPI was higher in Morangie (χ2 = 7.7, df = 1, p = 0.005), but again, did not differ by %FO (χ2 = 1.4, df = 1, p = 0.2).
When comparing the diet in Galloway with KS there was also a significant difference by %BPI (χ2 = 34.5, df = 5, p = < 0.001), whereby large mammal %BPI was higher in KS (χ2 = 22.5, df = 1, p = < 0.001).
Seasonal variance
Here, we analysed the seasonal variation between diets within Galloway, and data taken from Caryl (2008) and Balharry (1993) for Morangie and KS, respectively. In spring, we observed that bird %FO was higher (χ2 = 8.1, df = 1, p = 0.004), of which only egg %FO was higher (χ2 = 13, df = 1, p = < 0.001). We found no differences between any dietary group %BPI between seasons, despite an overall difference between diets (χ2 = 11.2, df = 5, p = 0.05).
In Morangie, small mammal %FO was higher in spring (χ2 = 9.5, df = 1, p = 0.002), while plant material %FO was higher in summer (χ2 = 11.7, df = 1, p = < 0.001). Whereas, large mammal %BPI was higher in spring (χ2 = 10, df = 1, p = 0.002) and plant material %BPI was higher in summer (χ2 = 15.7, df = 1, p = < 0.001). In KS, only large mammal %BPI was higher in spring (χ2 = 7.5, df = 1, p = 0.006).
Field vole abundance
Prey abundance was recorded for field voles from 2013 to 2016 in anticipation of their contribution to pine marten diet in Scotland (Fig. 5). Despite field vole relative index of abundance differing significantly during our study between 2015 and 2016 (td.f. = 0.008) we found no significant difference in their %FO in the diet (χ2 = 1.5, df = 1, p = 0.227).

Mean abundance index of field vole (Microtus agrestis) from 2013 to 2016 split by year in Galloway. J. Birks calculated average field vole (Microtus agrestis) population abundance indexes over 4 areas in Galloway Forest using the field vole transect sign index method devised by the Mammal Society (J. Birks pers. comm.). This was based on evidence of runs, feeding signs, and droppings recorded from 10 × 1 m quadrats placed along 100 m on transects in May over 4 years (2013–2016; J. Birks, unpubl. data). We averaged the 4 indices to give an average abundance index assumed to represent all of Galloway Forest for each year
Discussion
Galloway seasonal diet comparison
Although the diet of denning females and the mixed sex summer population diet in Galloway was similar, there were distinct differences. The breeding season of birds in spring plausibly explains the higher bird %FO in the diet of denning females compared to the mixed sex summer diet. During their breeding season, birds are nesting and thus birds, their eggs, and hatchlings are particularly vulnerable to an arboreal predator. We also relate the higher small mammal %FO in spring to seasonal, springtime births of these mammals. Wildlife often time their breeding to ensure sufficient resources are available to sustain females during the energetically costly lactation period (Bradbury and Vehrencamp 1977; Lewis and Kappeler 2005) or when young reach independence (Hayward et al. 2003). For a minimum of 4–7 denning female pine martens in Galloway, birds and their eggs were an important, energy-rich resource to sustain them through this period.
In the small mammal dietary group, field voles were by far the principal prey of martens in both seasons. Due to their open habitat preferences (e.g. Pearson 1985), field vole has been expressed to be an unusual preference of a highly arboreal, woodland predator by many authors, for example Balharry (1993) and Caryl (2008). Both authors, however, indicate that pine martens have a frequency independent preference for field voles. This explains the absence of annual fluctuation in field voles in the diet of denning females despite significant annual changes in their abundance within the study area.
The higher plant material and invertebrate %BPI and %FO in the mixed sex summer diet compared to the diet of denning females can be explained by seasonal availability of these dietary groups as seen in previous Scottish studies by Zalewski (2004) and Caryl (2008). Their equal %BPI within seasons indicate that these dietary groups are just as important in the diet of denning females as in the mixed sex summer population diet.
Alternatively, ground level foraging associated with invertebrates, resulting in incidental consumption of plant material, may contribute to this %BPI equality. However, as we observed that plant material consisted mostly of the same types clumped together, we considered this foraging deliberate.
We deemed any incidental ingestion of plant material from the digestive tracts of small mammals insignificant to plant material contribution as plant material frequently occurred exclusively of small mammals in the summer diet.
Denning and regional mixed sex spring diet comparison
It is evident that denning female pine martens in Galloway have a different dietary ecology to that of the mixed sex martens from other regions of Scotland. The major difference we observed was an absence of carrion in Galloway, which, despite having a low %FO in the other regions, contributed substantially to the large mammal dietary group %BPI due to the high correction factor associated with this prey item (Table 1). It is unlikely that large mammal occurrences were missed in our study due to the clumped nature of den box scats (see “Methods”) and that we identified all large mammal guard hairs found in addition to 20 guard hairs from clumped hairs from each scat. Further to this, we never observed the “tar-like” substance associated with carrion described by Balharry (1993).
The absence of carrion also likely explains the difference in %BPI of birds and small mammals, despite the relatively lower %FO of these dietary groups in Galloway compared to other Scottish regions. Nesting birds and their eggs and hatchlings associated with the breeding season of birds may have instead been deemed less risky prey by denning females. This is supported by Zalewski (2007), who showed that female martens fed less often on carrion than males based on sex-specific behavioural differences. Zalewski’s study did not focus specifically on the diet of denning females, and so this difference may be exacerbated due to denning behaviour, related to the added risk of scavenging on carrion while caring for kits, especially when considering red fox, another significant carrion scavenger (Baltrūnaitė 2002), is their main predator (Storch et al. 1990). Furthermore, Caryl (2008) observed that male martens in Morangie ranged, not only further from forest boundaries into open habitat, but for longer periods than female martens, hypothesising that due to their larger size, male martens are more willing to take risks.
Conversely, as carrion availability was not measured, it is also possible that carrion was less available to generalist martens in Galloway compared to other regions. Instead, small mammals, particularly field voles, and birds (especially in spring) may have been more reliably and regularly available to denning females during such a high energy lactation period associated with denning. We propose that a study of the mixed sex, non-denning spring diet of Galloway martens would be needed to investigate this further.
We observed further dietary variation when comparing anuran and reptile dietary groups. In Morangie, anuran, reptile and fungi were combined by Caryl (2008), while Balharry (1993) combined anuran with reptile in KS. This differs from this study in that we gave anuran its own distinct dietary group as we did not observe any reptile remains. We suggest that the opportunistic and generalist nature of martens in varying regional habitat likely explains the higher anuran %FO and %BPI and reptile absence in the Galloway diets. Firstly, anuran %FO and %BPI in the mixed sex summer diet in Galloway martens did not differ from the diet of denning females and showed a higher anuran %BPI compared to the mixed sex summer diet in Morangie. Secondly, no reptiles were observed in the Galloway diets. Alternatively, there was no difference between KS and Galloway in both seasonal diets. However, although the diet of denning females showed a higher anuran %BPI than the Morangie mixed sex spring diet, and both regional habitats were commercial coniferous woodlands, these habitats were distinctly different.
The habitat in Morangie was dominated by thinned scots pine (Pinus sylvestris) over a ground layer of heather (Calluna vulgaris) and blueberry (Vaccinium myrtillus) on relatively dry ground in the east that was more suitable for reptiles. Whereas Galloway was mainly un-thinned sitka spruce (Picea sitchensis) with open rides dominated by purple moor grass (Molinia caerulea) on wet gleyed soils in the west that was more suitable for anurans.
Mixed sex summer diet regional comparison
The mixed sex summer diet of pine martens in Galloway had not been explored before and so collections were made to further records in this area as well as for comparison with the diet of denning females and other regional mixed sex populations.
The lower large mammal %BPI in Galloway may be attributed to regional variation and correction factors, as above, as their %FO did not differ. It is worth noting, however, that our sample size of individually collected scats of the mixed sex summer diet in Galloway was small, and so some prey items may have been missed despite evidence of prey items occurring across as many as 12 different scats. The difference between Galloway and Morangie, in small mammals, likely relates to the substantially lower plant material %BPI in Galloway despite the lower small mammal %FO in Morangie. This could be attributed to the high correction factor given to pollen by Caryl (2008), not accounted for in this study.
The higher bird %FO and %BPI in Morangie compared to Galloway is likely linked to the lower anuran %FO and %BPI in Morangie. Conversely, similarities in plant material and anuran %BPI, and a higher bird %BPI in KS compared to Galloway indicates that large mammals, as in spring, are the most likely cause of %BPI fluctuations.
Squirrel predation
We hypothesise that it is unlikely denning females, and indeed the mixed sex summer population of martens in Galloway, pose a threat to the already threatened red squirrel in the area as we found only one record of red squirrel across their diets. Although our sample size of denning martens was small (n = 4–7) and the density of martens in Galloway was estimated to be low (0.13–0.15 pine marten per km2) by Croose et al. (2016a), a study by Halliwell (1997), reviewing Scottish pine marten diet from 1961, found that red squirrel accounted for no more than 0.4% of total FO, while more recently Caryl (2008) found no evidence of red squirrel in the diet of martens in Morangie. Furthermore, Sheehy and Lawton (2014) positively correlated red squirrel and pine marten abundance in Ireland. When also considering that Balharry (1993) found that a single squirrel can occur across 12 different scats, our data provides us with the plausible opinion that other denning females in Galloway likely have corresponding dietary habits. We also believe that it is possible that the culling of greys in and around the Galloway Forest District may effectively reduce grey numbers in the study area. In addition, White et al. (2015) estimate that the extensive coniferous nature of Galloway only supports very low grey and red squirrel densities, with a maximum carrying capacity of 0.006 squirrels per km2, consequently reducing their availability, and thus appearance in the pine marten diet.
Conclusions
We consider the diet of denning female pine martens in Galloway to be different to the mixed sex spring diet of pine martens in other regions of Scotland. Small mammals and birds clearly play an important role in the diet of denning females, as in the spring diets of mixed sex martens in other regions, but we propose that denning females in Galloway utilise these groups to a greater extent. We, therefore, suggest that denning females alter their foraging behaviour while caring for kits, whereby lower risk and possibly more readily available prey—such as birds and their eggs are targeted over higher risk and possibly less readily available prey —such as carrion. Furthermore, future den box placement and population restocking in Galloway to boost current low densities should especially consider locations with high anuran abundance in order to maximise food resources for denning females. In addition, due to the importance of recruitment to reintroductions, we suggest that future pine marten reintroduction and den box placement across Great Britain consider, more so than before, bird abundance, and, when habitat appropriate, anuran and reptile abundance also. As a final note, our data further supports evidence that red squirrels comprise a minimal part of pine marten diet in the British Isles.
References
Armstrong DP, Castro I, Griffiths R (2007) Using adaptive management to determine requirements of re-introduced populations: the case of the New Zealand hihi. J Appl Ecol 44(5):953–962
Balharry D (1993) Factors affecting the distribution and population density of pine martens (Martes martes L.) in Scotland. Dissertation, University of Aberdeen
Baltrūnaitė L (2002) Diet composition of the red fox (Vulpes vulpes L.), pine marten (Martes martes L.) and raccoon dog (Nyctereutes procyonoides Gray) in clay plain landscape, Lithuania. Acta Zoologica Lituanica 12(4):362–368
Birks J (2016a) Fleet basin pine marten project. Peoples trust for endangered species. https://ptes.org/wp-content/uploads/2017/01/Pine-martens-in-Scotland-final-report.pdf. Accessed 21 Apr 2017
Birks J (2016b) Pine martens in Scotland project update. Peoples trust for endangered species. https://ptes.org/wp-content/uploads/2017/01/Pine-martens-in-Scotland-final-report.pdf. Accessed 20 Apr 2017
Birks J, Messenger J (2010) Evidence of pine martens in England and Wales, 1996–2007: analysis of reported sightings and foundations for the future. Vincent Wildlife Trust, Ledbury
Birks JD, Messenger JE, Halliwell EC (2005) Diversity of den sites used by pine martens Martes martes: a response to the scarcity of arboreal cavities? Mammal Rev 35(3–4):313–320
Bradbury JW, Vehrencamp SL (1977) Social organization and foraging in emballonurid bats. Behav Ecol Sociobiol 2(1):19–29
Bright PW, Morris PA (1994) Animal translocation for conservation: performance of dormice in relation to release methods, origin and season. J Appl Ecol 31(4):699–708
Bruemmer CM, Rushton SP, Gurnell J, Lurz PWW, Nettleton P, Sainsbury AW, Duff JP, Gilray J, McInnes CJ (2010) Epidemiology of squirrelpox virus in grey squirrels in the UK. Epidemiol Infect 138(7):941–950
Bull EL (2000) Seasonal and sexual differences in American marten diet in northeastern Oregon. Northwest Science 74:186–191
Caryl FM (2008) Pine marten diet and habitat use within a managed coniferous forest. Dissertation, University of Stirling
Collins LM, Warnock ND, Tosh DG, McInnes C, Everest D, Montgomery WI, Scantlebury M, Marks N, Dick JT, Reid N (2014) Squirrelpox virus: assessing prevalence, transmission and environmental degradation. PLoS One 9(2):89521
Croose E, Birks JD, O’Reilly C, Turner P, Martin J, MacLeod ET (2016a) Sample diversity adds value to non-invasive genetic assessment of a pine marten (Martes martes) population in Galloway Forest, Southwest Scotland. Mamm Res 61(2):131–139
Croose E, Birks JD, Martin J (2016b) Den boxes as a tool for pine marten Martes martes conservation and population monitoring in a commercial forest in Scotland. Conservation Evidence 13:57–61
De Marinis AM, Masseti M (1995) Feeding habits of the pine marten Martes martes L., 1758, in Europe: a review. Hystrix 7(1–2):143–150
Fischer J, Lindenmayer DB (2000) An assessment of the published results of animal relocations. Biol Conserv 96(1):1–11
Griffith B, Scott JM, Carpenter JW, Reed C (1989) Translocation as a species conservation tool: status and strategy. Science 245(4917):477–480
Gurnell J, Wauters LA, Lurz PW, Tosi G (2004) Alien species and interspecific competition: effects of introduced eastern grey squirrels on red squirrel population dynamics. J Anim Ecol 73(1):26–35
Halliwell EC (1997) The ecology of red squirrels in Scotland in relation to pine marten predation. Dissertation, University of Aberdeen
Hayward MW, de Tores PJ, Dillon MJ, Fox BJ (2003) Local population structure of a naturally-occurring metapopulation of the quokka (Setonix brachyurus Macropodidae: Marsupialia). Biol Conserv 110:343–355
Kleef HL, Wijsman HJ (2015) Mast, mice and pine marten (Martes martes): the pine marten’s reproductive response to wood mouse (Apodemus sylvaticus) fluctuations in the Netherlands. Lutra 58:23–33
Lanszki J, Zalewski A, Horváth G (2007) Comparison of red fox Vulpes vulpes and pine marten Martes martes food habits in a deciduous forest in Hungary. Wildl Biol 13(3):258–271
Levins R (1968) Evolution in changing environments: some theoretical explorations. Princeton University Press, Princeton
Lewis RJ, Kappeler PM (2005) Seasonality, body condition, and timing of reproduction in Propithecus verreauxi verreauxi in the Kirindy Forest. Am J Primatol 67(3):347–364
Lindström ER, Brainerd SM, Helldin JO, Overskaug K (1995) Pine marten—red fox interactions: a case of intraguild predation? Ann Zool Fenn 32(1):123–130
Lockie JD (1961) The food of the pine marten martes martes in west ross-shire, Scotland. Proc Zool Soc London 136(2):187–195
Lockie JD (1964) Distribution and fluctuations of the pine marten, Martes martes (L.), in Scotland. J Anim Ecol 33(2):349–356
Lombardini M, Murru M, Repossi A, Cinerari CE, Rosin AV, Mazzoleni L, Meriggi A (2015) Spring diet of the pine marten in Sardinia, Italy. Anim Biodivers Conserv 38(2):183–190
Lynch ÁB, McCann Y (2007) The diet of the pine marten (Martes martes) in Killarney National Park. Biol Environ Proc R Irish Acad 107B(2):67–76
MacPherson J (2016) Second phase of pine marten translocations. Vincent wildlife Trust Pine marten recovery project. http://www.pine-marten-recovery-project.org.uk/blog/second-phase-of-pine-marten-translocations-from-scotland-to-wales. Accessed 19 Apr 2017
Moors PJ (1980) Sexual dimorphism in the body size of mustelids (Carnivora): the roles of food habits and breeding systems. Oikos 34(2):147–158
Pearson OP (1985) Predation. In: Tamarin RH (ed) Biology of New World Microtus. Special Publ No 8 Am Soc. Mammalog 8:535–566
Pianka ER (1973) The structure of lizard communities. Annu Rev Ecol Syst 4(1):53–74
Powell RA (1979) Mustelid spacing patterns: variations on a theme. Mustela Z Tierpsychol 50:153–165
Shaw G, Livingstone J (1992) The pine marten: its reintroduction and subsequent history in the Galloway Forest Park. Trans Dumfries Galloway Nat His Antiq Soc 67:1–7
Sheehy E, Lawton C (2014) Population crash in an invasive species following the recovery of a native predator: the case of the American grey squirrel and the European pine marten in Ireland. Biodivers Conserv 23(3):753–774
Sheehy E, Lawton C (2015) Predators of red and grey squirrels in their natural and introduced ranges. In: Shuttleworth C, Lurz PWW, Hayward M (eds) Red squirrels: ecology, conservation & management in Europe. European Squirrel Initiative, Woodbridge, pp 83–96
Storch L, Lindstrtim E, de Jounge J (1990) Diet and habitat selection of the pine marten in relation to competition with the red fox. Acta Theriol 35:311–320
Strachan R, Jefferies DJ, Paul RC (1996) Pine marten survey of England and Wales, 1987–1988. JNCC, Peterborough
Teerink BJ (2003) Hair of West European mammals: atlas and identification key. Cambridge University Press, Cambridge
Thompson ID, Colgan PW (1987) Numerical responses of martens to a food shortage in northcentral Ontario. J Wildl Manag 51(4):824–835
Townsend M (2017) Pine martens to return to the Forest of Dean? The Woodland Trust. https://www.woodlandtrust.org.uk/blogs/woodland-trust/2017/04/reintroduction-of-pine-martens/. Accessed 20 May 2017
White A, Lurz PW, Jones HE, Boots M, Bryce J, Tonkin M, Ramoo K, Bamforth L, Jarrott A (2015) The use of mathematical models in red squirrel conservation: assessing the threat from grey invasion and disease to the Fleet basin stronghold. Red squirrels: ecology, conservation & management in Europe, 4. European Squirrel Initiative, Woodbridge
Zalewski A (2004) Geographical and seasonal variation in food habits and prey size of European pine martens. In: Harrison DJ, Fuller AK, Proulx G (eds) Martens and fishers (Martes) in human-altered environments. Springer, London, pp 77–98
Zalewski A (2007) Does size dimorphism reduce competition between sexes? The diet of male and female pine martens at local and wider geographical scales. Acta Theriol 52(3):237–250
Acknowledgements
We have many people to thank for the completion of this study. To start, we would like to thank Pete Haswell, as without his provision of alternative lab space this project would not have gone as smoothly as it somehow did. This includes providing needed equipment and reference guides including his own experience and samples in dietary analysis. Emlyn Roberts was also a crucial part in supplying equipment and what was, essentially, our own fume cupboard for the entire project instead of kicking us out of the building due to the smell. Jane Lee and John Latchford were also of great help and more than deserve our thanks for permission to conduct lab work out of hours when we needed to put some extra hours in. For her moral support and encouragement, Wynn Johnston also deserves our special thanks.
From outside of the university, we are particularly grateful to Johnny Birks for patiently providing his vast knowledge of this field, answering any questions we had in depth. In addition, he and John Martin were instrumental in the collection of scats and supplementary data used in this study—without them this study would not have been able to start. Their previous and continued devotion to pine marten ecology is truly inspiring and we look forward to seeing their important work in this area in future.
I (Alexander Grabham) must also mention Leaflen and Debs for allowing me to stay with them while visiting the study site enabling me to focus on the job at hand.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors and was approved by the Bangor University ethics committee.
Additional information
Communicated by: Andrzej Zalewski
Electronic supplementary material
ESM 1
(XLSX 16.8 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Grabham, A.A., Ventress, G. & Hayward, M.W. The diet of denning female European pine martens (Martes martes) in Galloway Forest District, South West Scotland, Great Britain. Mamm Res 64, 87–97 (2019). https://doi.org/10.1007/s13364-018-0398-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13364-018-0398-5